
Elevated CO2 and Increased N Intensify Competition between Two Invasive Annual Plants in China
1
State Key Laboratory of Environmental Criteria and Risk Assessment, Chinese Research Academy of Environmental Sciences, Beijing 100012, China
2
China National Accreditation Service for Conformity Assessment, Beijing 100062, China
3
Command Center for Comprehensive Survey of Natural Resources, China Geological Survey Bureau, Beijing 100055, China
*
Author to whom correspondence should be addressed.
Life 2022, 12(10), 1669; https://doi.org/10.3390/life12101669
Submission received: 27 September 2022
/
Revised: 9 October 2022
/
Accepted: 17 October 2022
/
Published: 21 October 2022
(This article belongs to the Special Issue Interaction between Invasive and Native Plants)
Abstract
:As multiple invaders often co-occur, understanding the interactions between different invasive species is important. Previous studies have reported on invasional meltdown and neutral and interference relationships between invasive species. However, interspecific interactions may vary with environmental change owing to the different responses of interacting invaders. To better understand the interaction of notorious invasive alien plants under CO2 enrichment and N deposition, the growth characteristics of common ragweed (Ambrosia artemisiifolia) and redroot pigweed (Amaranthus retroflexus) were studied when they were planted in monoculture (4Rag and 4Pig) or mixture (1Rag:3Pig, 2Rag:2Pig, 3Rag:1Pig) under four environmental treatments: elevated CO2, increased N, elevated CO2 + increased N and a control. Increased N positively affected almost all the traits (basal stem diameter, height, shoot biomass, root biomass and total biomass) of common ragweed, except for branch number and root-shoot ratio. But increased N only promoted redroot pigweed’s height and basal stem diameter. interspecific competition promoted basal stem diameter and number of branches but decreased root biomass of common ragweed, and the basal stem diameter was significantly higher in 1Rag:3Pig and 2Rag:2Pig compared to the other two treatments. interspecific competition inhibited almost all the characteristics of redroot pigweed. The interaction between elevated CO2 and increased N also increased the biomass characteristics (shoot biomass, root biomass and total biomass) of common ragweed. However, elevated CO2 inhibited the root biomass of redroot pigweed. The results indicated that common ragweed was a superior competitor under conditions of elevated CO2 and increased N. Moreover, environmental change might strengthen the super-invasive plant common ragweed’s competitive ability.
1. Introduction
Invasive plants often show high adaptability and phenotypic plasticity, which allow them to thrive under altered environmental conditions [1,2]. Numerous case studies have demonstrated that invasive plants benefit from increasing N deposition and elevated CO2 [3,4,5,6]. Elevated CO2 can facilitate plant invasion by increasing plant photosynthesis, growth rates, efficient resource use, productivity, and seed production [7,8,9,10,11,12]. N deposition can facilitate plant invasion by increasing N availability, plant growth, and competitive ability [13,14,15,16]. These factors often change simultaneously. However, in most studies, these two factors are studied independently.
Understanding the interactive effect of elevated CO2 and N deposition on invasive alien plants is important. Recent research demonstrated that elevated CO2 and N deposition synergistically increased the performance (such as biomass and size, survival and reproduction, and photosynthetic rate) of invasive [4,17,18] or naturalized alien plants [6] than native. Previous studies tested the effect of elevated CO2 and N deposition on alien plants compared with native plants, but few studies have tested the effect among various invasive plants.
As biological invasions increase in frequency, most habitats are invaded by multiple invasive plants, and interactions between invasive species have attracted attention [19,20,21]. Thus far, three types of interactions have been described: (1) Establishment and impact of one alien invasive species can be facilitated by another invasive species, which is described as invasional meltdown [22]; (2) neutral interactions have also been described [23]; (3) invasion by one species can be negatively impacted by the presence of another invader, termed invasional interference [24,25,26]. Many authors have noted that a decline in one nonnative species results in a rapid increase in another, which indicates that competition among invasive plants may be common [21,27]. In some cases, these negative relationships may result in an invasive species replacing another invasive species, termed “over-invasion” [20]. For example, the replacement of Spartina anglica by S. alterniflora was found in China [28]. Interactions between invasive plants under elevated CO2 and N deposition remain unclear.
Common ragweed and redroot pigweed are two notorious invasive species both native to North America. They are both included on the list published by the Ministry of Ecology and Environment of the People’s Republic of China [29]. Common ragweed, an annual weed in crop fields, usually forms dense mono-specific stands and produces a considerable amount of pollen [30], and this weed is one of the most problematic aero-allergens [31]. It was introduced into China in the 1930s and, since then, it has spread across twenty provinces [32]. Redroot pigweed occurs in various habitats, including agricultural and ruderal habitats [25,33]. It has a negative impact on ecosystems and native species [34,35] and is regarded as the third most notorious weed in the world [25,36]. Redroot pigweed, introduced into China around 1905, has expanded its distribution in large areas [37]. These two invasive species often co-occur in crops or other habitats [38,39].
In the present study, we tested competitive interactions between these two invasive alien plants in monoculture and mixture under four environmental treatments: elevated CO2, increased N, elevated CO2 and N, and a control. We aimed to answer the following questions: (1) do the growth characteristics of these two invasive alien plants respond to elevated CO2, increased N, and replacement in the same way? (2) which invasive species is more competitive under elevated CO2, increased N deposition, and replacement? (3) how do elevated CO2, N deposition, and replacement affect the reproduction of common ragweed?
2. Materials and Methods
2.1. Plant Materials
We collected seeds of common ragweed and redroot pigweed from Mentougou District in Beijing (100 km from the experiment site) in October 2013. The seeds were treated at low temperatures (−20 °C) for two months to break dormancy and then stored at room temperature in paper bags. The seeds of the two species were sown on 7 May 2014, at a depth of 2 cm in two field plots and then watered to field capacity once to stimulate germination. After three weeks, seedlings were transplanted into pots in the OTCs according to the experimental design.
2.2. Experimental Design
The experiment was conducted at the Chinese Research Academy of Environmental Sciences field laboratory, Shunyi District, Beijing, China (116.5875° E, 40.19° N), between 5 June and 8 October 2014.
We tested two CO2 concentrations in our experiment: ambient CO2 (375 ppm) concentrations and elevated CO2 (700 ppm) based on IPCC (Intergovernmental Panel on Climate Change) [40]. Four pairs of open-top chambers (OTCs: 2.2 m in height with an octagonal ground surface area of 6.25 m2) were used. During the experiment, pure CO2 was continuously ventilated into the OTCs of elevated CO2 treatment. Elevated CO2 concentrations were measured at 5-min intervals by monitoring sensors (Qs100). In the elevated CO2 treatment, the achieved level was 695.00 ± 15.67 (mean ± SD) ppm. Ambient CO2 and elevated CO2 were randomly assigned to each pair. According to the increase in N deposition rates over the coming decades in China [41,42], two N levels were conducted: ambient N (0 addition) and increased (N 0. 8 g/pot).
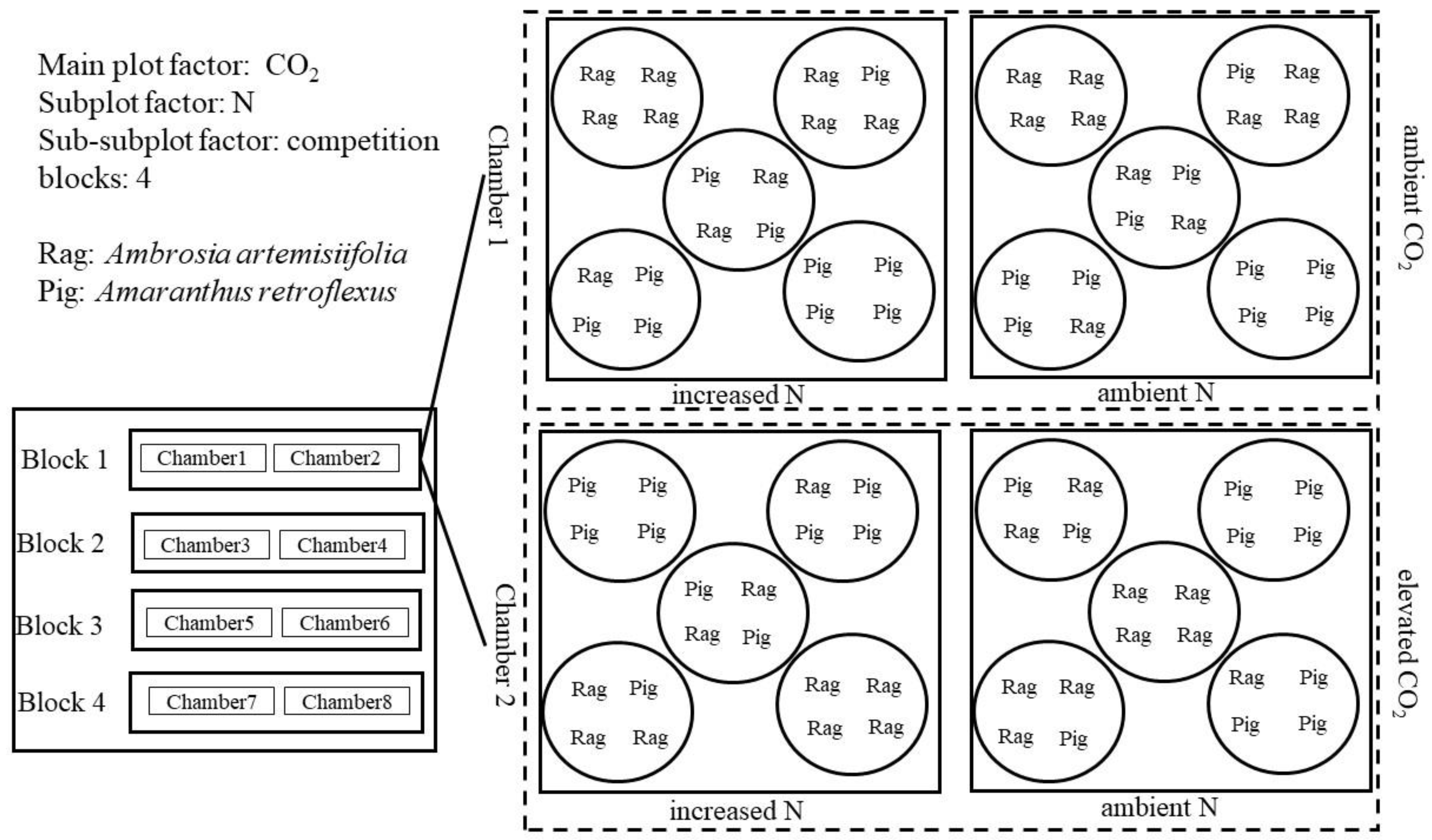
Four seedlings were planted in each pot under the replacement design: 4Rag:0Pig, 1Rag:3Pig, 2Rag:2Pig, 3Rag:1Pig, and 0Rag:4Pig (where Rag and Pig denote common ragweed and redroot pigweed, respectively). Two N levels, and five replacement levels within each N level, produced 10 pots in each chamber. Thus, 80 pots (32 cm in diameter and 38 cm in depth) were randomly arranged within 8 OTC in the following experimental design: 2 CO2 levels × 2 N levels × 5 replacement levels × 4 replicates (Figure 1). During the growing season, N was equally divided eight times and uniformly applied in the form of NH4NO3 solution, while the control pots were sprayed with the same volume of water.
To ensure homogeneity of the growing conditions, the pots were filled with a mixture of local soil (80%; collected at a depth of 3~15 cm from a weedy field neighboring the experiment base after excluding topsoil) and vermiculite (20%). Before transplanting, the following soil nutrient contents were measured: organic carbon, 8.4 g·kg−1; total N, 0.73 g·kg−1; ammonium N, 10.77 mg·kg−1; and nitrate N, 6.53 mg·kg−1. One week after transplanting, the elevated CO2 and N enrichment treatments were initiated. Plants were watered weekly as needed throughout the experiment. In each OTC, pots were rearranged randomly within the chamber every month to minimize location-specific effects.
2.3. Measurements and Calculations
Plant height, basal stem diameter, and branch number per plant of common ragweed and redroot pigweed were measured in October. Shoots of each species in every pot were harvested from above the soil surface and stored in archival paper bags. We turned over the pot, removed the soil of roots with running water, carefully separated the roots of two species in every pot, and then stored them in paper bags. The shoots and roots were oven-dried at 80 °C for 72 h. Shoot biomass and root biomass of each species in each pot were measured, and the total biomass of each species was calculated per pot.
The competitive ability of these two invasive plants was measured by relative yield. RY values > 1 indicate that one species does better when competing against the other species than when competing against itself [43]. RY values were calculated using the equation below [43,44].
where Yij is the yield of species i in the presence of species j, pi is the proportion at which species i is sown, and Yi is the yield of species i in monoculture under the same CO2 and N treatment as that for Yij.
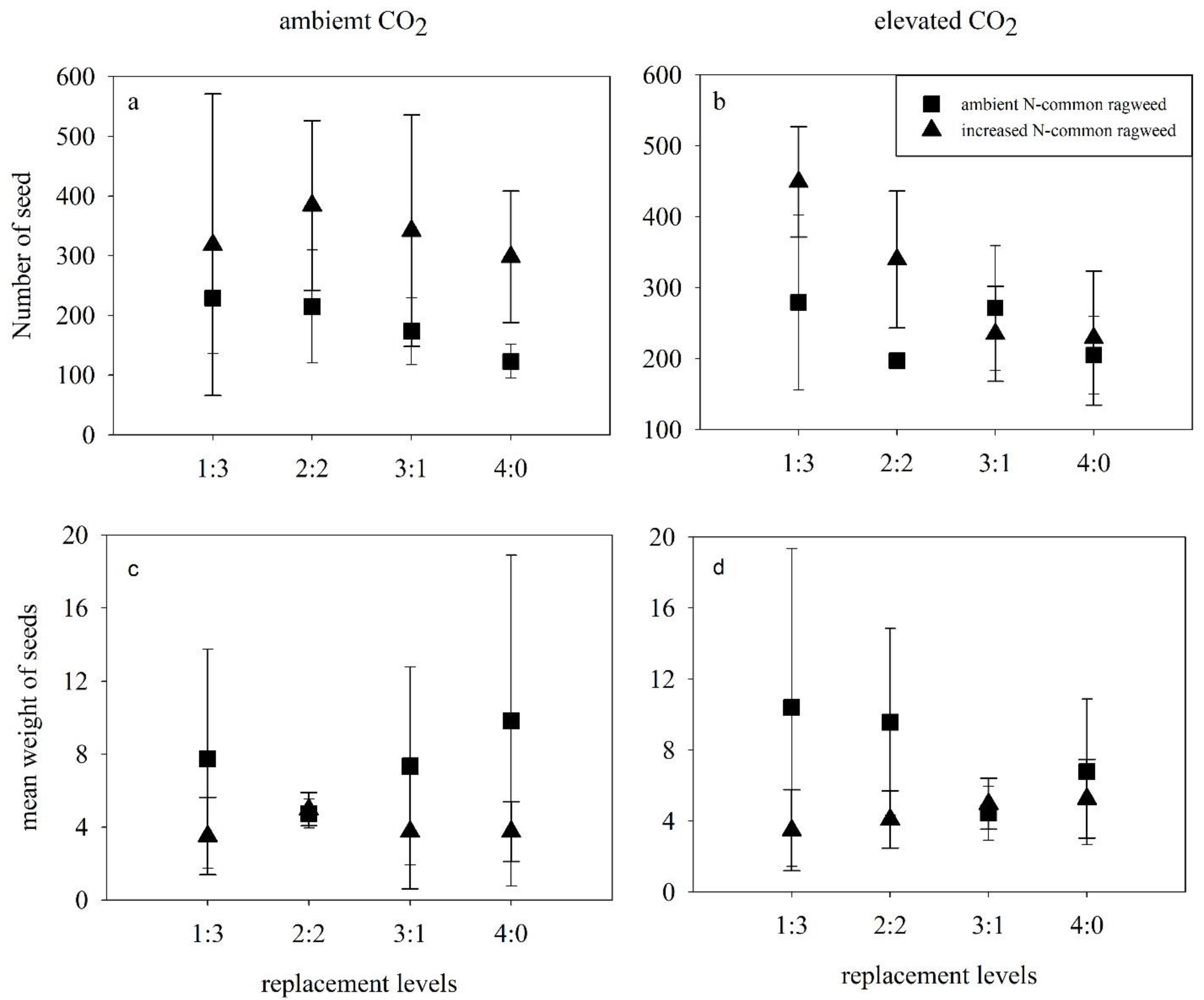
To analyze the impact of elevated CO2, increased N, and replacement on the reproduction of common ragweed, in each pot with common ragweed, seeds were collected by hand in October when the seeds were mature but had not yet begun to drop. The total seed number and the total seed weight were measured as seed yield.
2.4. Statistical Analysis
Split-plot ANOVA was employed to test the effects of CO2, N, and replacement on the plant performance and relative yield of common ragweed and redroot pigweed, with block as a random factor. When replacement level had a significant effect, significant differences between replacement levels were tested using the Tukey honesty significant difference post-hoc analyses (HSD) (p < 0.05). In all ANOVAs, data were log-transformed to conform to the assumptions of normality and homoscedasticity. All analyses were performed using IBM SPSS Statistics 19 (IBM, 2010, New York, NY, USA).
3. Results
3.1. Growth Characteristics of Two Invasive Alien Plants
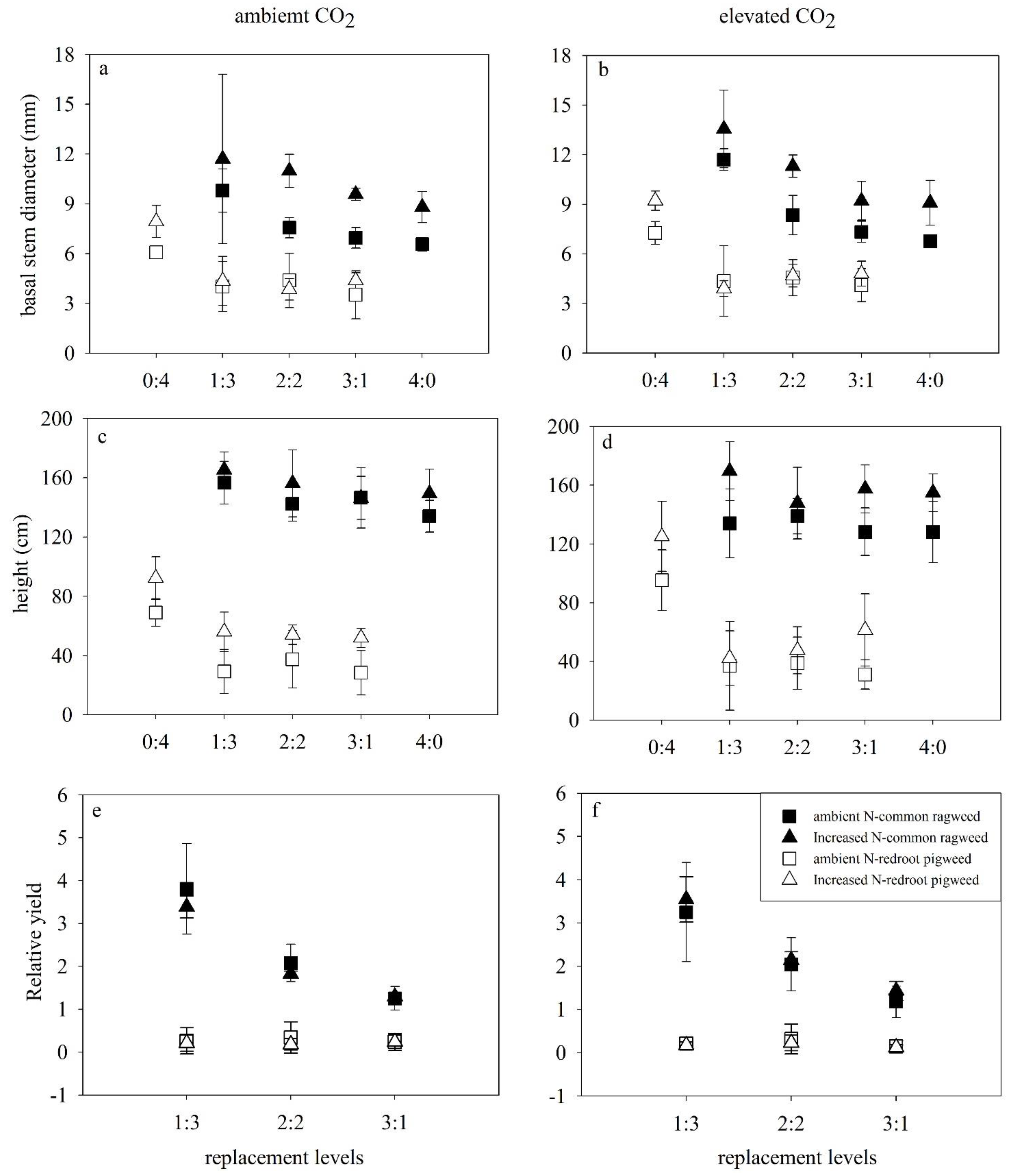
Results revealed that height and basal stem diameter of common ragweed were significantly enhanced by increased N in both monoculture and mixtures under both CO2 concentrations (p < 0.001) (Figure 2a–d; Table 1). At the same time, increased N enhanced (p < 0.05) the height and basal stem diameter (p < 0.001) of redroot pigweed.
Species replacement (p < 0.001) also promoted the basal stem diameter of common ragweed, which was significantly larger in the mixture than in monoculture. The mean stem diameter of ragweed was the largest in the 1Rag:3Pig (p < 0.05, HSD test) (Figure 2a,b), and also significantly larger in the 2Rag:2Pig than in monoculture (p = 0.014, HSD test). Species replacement (p < 0.001) decreased the height and basal stem diameter of redroot pigweed (Figure 2a–d, Table 1). These two characteristics of redroot pigweed in monoculture were all significantly larger than all mixture treatments (p < 0.001, Figure 2a–d; HSD test). Species replacement increased the branch number of common ragweed (p < 0.001), but no branches were observed for redroot pigweed during the experiment.
The impacts of elevated CO2 on these two invasive plants were not found. A significant interaction of impacts on redroot pigweed was detected in increased N and competition (p = 0.041), elevated CO2 and species replacement (p = 0.019).
3.2. Biomass Characteristics of Two Invasive Alien Plants
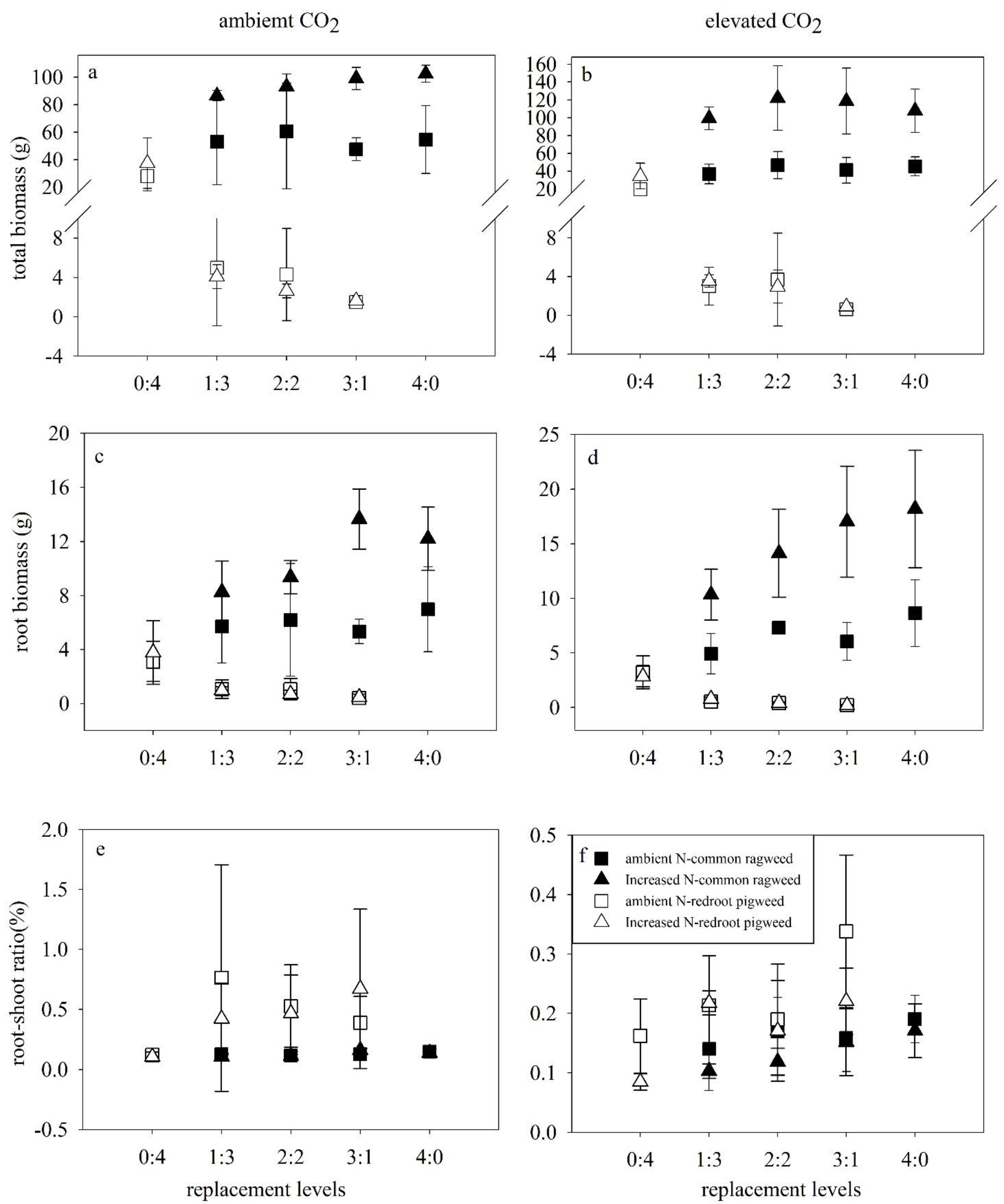
Shoot biomass, root biomass, and total biomass of common ragweed were all significantly increased by N addition in both monoculture and mixtures under both CO2 concentrations (Figure 3a–d; Table 2). No impacts of N addition were found in biomass characters of redroot pigweed.
Root biomass and the root-shoot ratio of common ragweed were affected by species replacement. Interspecific and intraspecific competition all impacted root biomass and were higher in monoculture (p = 0.001, HSD test) and decreased in 3Rag:1Pig (p = 0.020, HSD test) than in 1Rag:3Pig mixture. The root-shoot ratio of common ragweed was significantly lower in the 3Rag:1Pig mixture than in monoculture (p = 0.011, HSD test). Shoot biomass, root biomass, and total biomass of redroot pigweed were all significantly suppressed in the mixture compared with monoculture (p < 0.001) (Figure 3a–d; Table 2), and these characteristics were all decreased in 3Rag:1Pig than in the other two mixture controls (p < 0.05, HSD test). The root-shoot ratio of redroot pigweed was significantly higher in mixtures than in monoculture (p < 0.05, HSD test) (Figure 3e, f).
Elevated CO2 inhibited the root biomass (p = 0.011) of redroot pigweed in mixtures (Figure 3c, d). An interaction between elevated CO2 and increased N promoted shoot biomass (p < 0.001), total biomass (p < 0.001), and root biomass (p = 0.03) of common ragweed.
3.3. Relative Yields of Two Invasive Alien Species
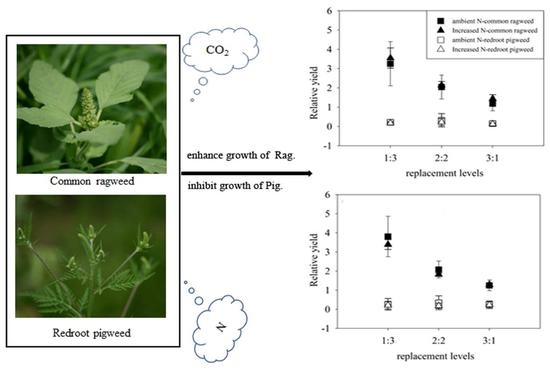
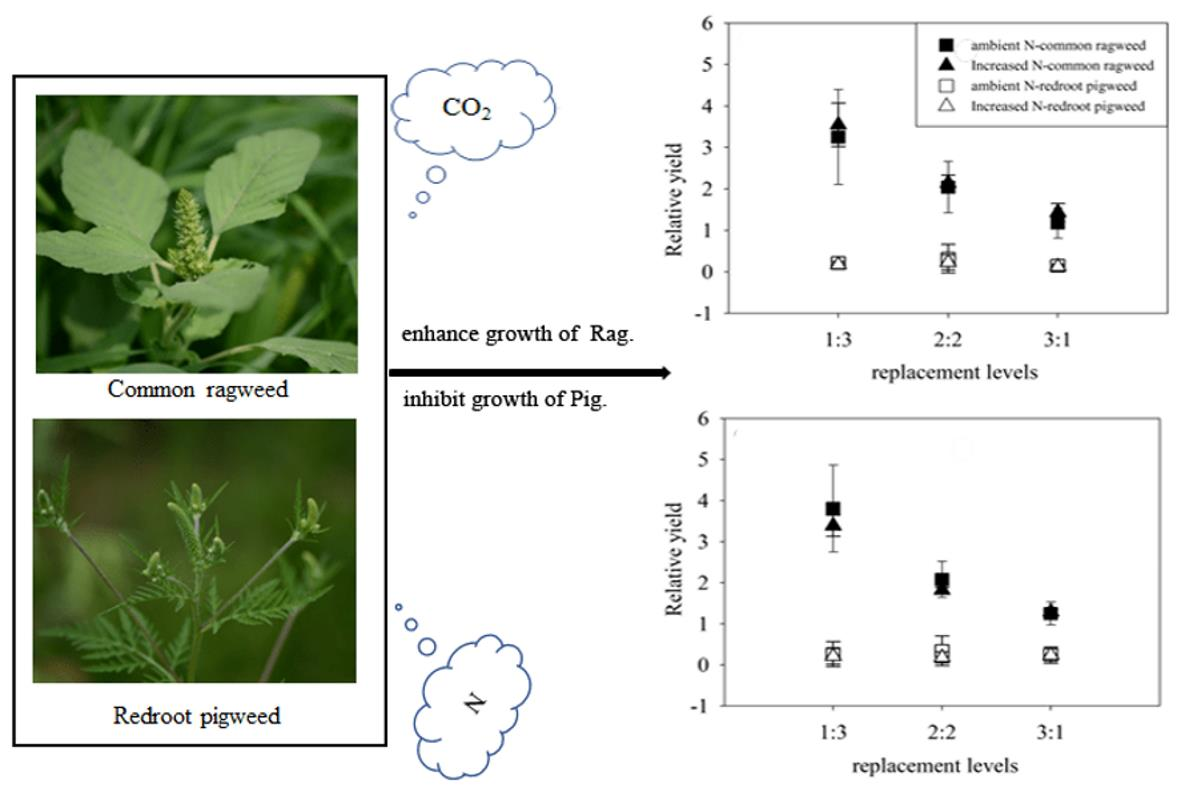
The relative yields of common ragweed were significantly higher than 1 (p < 0.001, t-test) irrespective of any treatments; while these values were significantly lower than 1 (p < 0.001, t-test) for redroot pigweed (Figure 2e,f). The highest relative yield (RY) values of common ragweed reached 4.72, and the mean values of relative yield were larger in 1Rag:3Pig compared to other replacement treatments (p < 0.001, HSD test), while these values in 2Rag:2Pig were higher than in 3Rag:1Pig (p = 0.001, HSD test). In contrast, the largest relative yield of redroot pigweed was only 0.87, and the mean values of relative yield were higher in 2Rag:2Pig compared to other replacement treatments; however, no significant difference was observed.
3.4. Reproductive Characteristics of Common Ragweed
4. Discussion
In the present study, we found common ragweed and redroot pigweed responded differently to elevated CO2, increased N, and species replacement. We revealed the interspecific competition between common ragweed and redroot pigweed under elevated CO2 and increased N.
Common ragweed showed an apparent competitive advantage over redroot pigweed under mixture treatments, where even one common ragweed plant could strongly inhibit redroot pigweed plants. Common ragweed and redroot pigweed may compete for limited resources. The tall statue of the common ragweed provided a decisive advantage in acquiring light [45]. Common ragweed was higher and more robust than redroot pigweed in mixture treatments (Figure 1 and Figure 2). The height disadvantage of redroot pigweed under competition results in a decrease in its light-capturing capacity [46]. The decline in quality and quantity of light influenced the phenology of pigweed [35,47], so the growth of redroot pigweed was inhibited under competition. We also found the height of redroot pigweed are all positively related to the other characteristics. By comparing the competition between redroot pigweed and common lamb’s quarters (Chenopodium album), it was also found that the species which shade the ground with a dense canopy will favorably compete [48]. In our study, we also found that branch numbers of common ragweed increased with competition, but no branches were observed in redroot pigweed. When two alien invasive plants live in the same habitat or require the same scarce resource [49,50], one species limiting the material or space of the other will result in competition.
Growth characteristics (except for branch number and root-shoot ratio) of common ragweed were enhanced by increased N in all treatments. Only the height and basal stem diameter of redroot pigweed increased in the mixtures under increased N (Figure 1 and Figure 2). Our results are consistent with the premise that responsiveness considerably differs between species under N addition [51]. We also found that increased N and elevated CO2 intensified the competitive advantage of common ragweed. For example, biomass characteristics of common ragweed were positively affected by the interaction of increased N and elevated CO2. Lack of an elevated CO2 direct effect on these two invasive alien plants might be explained by indirect effects of CO2 on N limitation, similar to Blumenthal et al. [52]. First, these two invasive alien plants all prefer high levels of nitrogen fertilizer. A previous study showed that the height and dry matter of common ragweed were increased by N addition in both greenhouse and field experiments [53]. Moreover, N addition stimulated the height of redroot pigweed [35] and shoot biomass and root biomass [51]. In the present study, the biomass of redroot pigweed did not increase under N addition in mixtures. Mainly because common ragweed can competitively pre-empt soil nitrogen from redroot pigweed. Secondly, the CO2 response of species might depend on local resource availability in mixed-species competition [4]. Most plants exhibit a positive growth response to elevated CO2 due to increased photosynthesis and/or efficient nutrient use when other factors (e.g., water and nutrients) are not limited [6,54]. Because most invasive plants are sensitive to N availability, the impacts of CO2 on invasive plants varied with N levels [52]. Redroot pigweed increased biomass allocation to the roots to diminish aboveground competitive disadvantage, where plants allocate more biomass to roots to acquire the most limiting resource [55,56,57]. But elevated CO2 inhibits this increase (Figure 2). A previous study on redroot pigweed revealed lower rates of photosynthesis and stomatal conductivity under elevated CO2 compared to ambient in water stress treatments [58,59]. We speculate that there is an apparent limit of available N for redroot pigweed growth under competition. Third, C3 plants are thought to take more advantage of CO2 enrichment than plants with a C4 [60,61]. In our study, common ragweed was a C3 plant and redroot was a C4 plant. Fourth, allelopathic effects also impacted interspecific competition. Bae et al. found that elevated CO2 may enhance the allelopathic potential of common ragweed [62].
The competition outcome varies with the performance [45] and resource availability of neighboring species [63]. For example, Italian ryegrass has a competitive advantage over common ragweed [64], while redroot pigweed shows higher competitiveness than Phaseolus vulgaris [65]. Competitiveness decreased with an increasing density of common ragweed. The highest competitiveness was under 1Rag:3Pig, where the relative yield of common ragweed increased to 350%. Results indicated interspecific and intraspecific generality, as detected in other experiments [4]. It is also possible that release from intraspecific competition allows common ragweed to grow higher and more robust. In contrast, their biomass decreased when redroot pigweed was released from the intraspecific competition (Figure 1 and Figure 2). Intra- or con-specific competition is always stronger than interspecific one [66,67]. Previous studies have shown that elevated CO2 and N deposition might favor performances of invasive plants relative to that of native species [3]. Our results revealed elevated CO2 and increased N prefer the super competitor when two notorious alien invasive plants grew together.
5. Conclusions
The purpose of this study was to understand the differences in growth characteristics of two alien invasive plants under elevated CO2, increased N, and species replacement. The results showed that the biomass of common ragweed was positively enhanced by increased N and species replacement, but redroot pigweed was negatively inhibited. The relative yield of these two notorious invasive plants revealed a competition interaction. In addition, our results revealed that common ragweed gained a more competitive advantage than redroot pigweed under increased N and elevated CO2. Our results indicated that common ragweed may be replaced redroot pigweed in heavily invaded regions. The competition or coexistence of redroot pigweed and common ragweed should be discussed under different invasion stages in the future, especially under environmental change. Moreover, understanding the interaction of invasive alien species will help us to manage multispecies invasions in the future.
Author Contributions
Conceptualization, J.L. and C.Z.; methodology, X.Z.; software, C.Z.; formal analysis, C.Z.; investigation, X.Z.; data curation, C.Z. and X.Z.; writing—original draft preparation, C.Z.; writing—review and editing, C.Z. and J.L.; visualization, C.Z.; supervision, J.L. and C.Z.; project administration, J.L. and C.Z.; funding acquisition, J.L. and C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2016YFC1201100, 2017YFC0506200).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; van Kleunen, M. Responses of common and rare aliens and natives to nutrient availability and fluctuations. J. Ecol. 2017, 105, 1111–1122. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Oduor, A.M.O.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Xu, X.; van Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Glob. Chang. Biol. 2017, 23, 3363–3370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Mozdzer, T.J.; Langley, J.A.; Mueller, P.; Megonigal, J.P. Deep rooting and global change facilitate spread of invasive grass. Biol. Invasions 2016, 18, 2619–2631. [Google Scholar] [CrossRef]
- Sorte, C.J.B.; Ibanez, I.; Blumenthal, D.M.; Molinari, N.A.; Miller, L.P.; Grosholz, E.D.; Diez, J.M.; D’Antonio, C.M.; Olden, J.D.; Jones, S.J.; et al. Poised to prosper? A cross-system comparison of climate change effects on native and non-native species performance. Ecol. Lett. 2013, 16, 261–270. [Google Scholar] [CrossRef]
- Dukes, J.S.; Chiariello, N.R.; Loarie, S.R.; Field, C.B. Strong response of an invasive plant species (Centaurea solstitialis L.) to global environmental changes. Ecol. Appl. 2011, 21, 1887–1894. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H. Evaluation of the growth response of six invasive species to past, present and future atmospheric carbon dioxide. J. Exp. Bot. 2003, 54, 395–404. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Belote, R.T.; Sanders, N.J. Biological invaders in a greenhouse world: Will elevated CO2 fuel plant invasions? Front. Ecol. Environ. 2003, 1, 146–153. [Google Scholar] [CrossRef]
- Smith, S.D.; Huxman, T.E.; Zitzer, S.F.; Charlet, T.N.; Housman, D.C.; Coleman, J.S.; Fenstermaker, L.K.; Seemann, J.R.; Nowak, R.S. Elevated CO2 increases productivity and invasive species success in an arid ecosystem. Nature 2000, 408, 79–82. [Google Scholar] [CrossRef]
- Stewart, J.; Potvin, C. Effects of elevated CO2 on an artificial grassland community: Competition, invasion and neighbourhood growth. Funct. Ecol. 1996, 10, 157–166. [Google Scholar] [CrossRef]
- Smith, S.D.; Charlet, T.N.; Zitzer, S.F.; Abella, S.R.; Vanier, C.H.; Huxman, T.E. Long-term response of a Mojave Desert winter annual plant community to a whole-ecosystem atmospheric CO2 manipulation (FACE). Glob. Change Biol. 2014, 20, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.-Y.; Qi, S.-S.; Zou, C.B.; Dai, Z.-C.; Ren, G.-Q.; Chen, Q.; Zhu, B.; Du, D.-L. Elevated nitrogen deposition may advance invasive weed, Solidago canadensis, in calcareous soils. J. Plant Ecol. 2019, 12, 846–856. [Google Scholar] [CrossRef]
- He, W.-M.; Yu, G.-L.; Sun, Z.-K. Nitrogen deposition enhances Bromus tectorum invasion: Biogeographic differences in growth and competitive ability between China and North America. Ecography 2011, 34, 1059–1066. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, W.; Cai, Z.; Valerie, P.; Han, F. Response of methane emission to invasion of Spartina alterniflora and exogenous N deposition in the coastal salt marsh. Atmos. Environ. 2010, 44, 4588–4594. [Google Scholar] [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. N deposition affects N availability in interstitial water, growth of Sphagnum and invasion of vascular plants in bog vegetation. New Phytol. 2003, 157, 339–347. [Google Scholar] [CrossRef]
- Lei, Y.B.; Wang, W.B.; Feng, Y.L.; Zheng, Y.L.; Gong, H.D. Synergistic interactions of CO2 enrichment and nitrogen deposition promote growth and ecophysiological advantages of invading Eupatorium adenophorum in Southwest China. Planta 2012, 236, 1205–1213. [Google Scholar] [CrossRef]
- Nackley, L.; Hough-Snee, N.; Kim, S.H. Competitive traits of the invasive grass Arundo donax are enhanced by carbon dioxide and nitrogen enrichment. Weed Res. 2017, 57, 67–71. [Google Scholar] [CrossRef]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Russell, J.C.; Sataruddin, N.S.; Heard, A.D. Over-invasion by functionally equivalent invasive species. Ecology 2014, 95, 2268–2276. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Nuñez, M.A. Negative, neutral, and positive interactions among nonnative plants: Patterns, processes, and management implications. Glob. Chang. Biol. 2015, 21, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D.; Von Holle, B. Positive Interactions of Nonindigenous Species: Invasional Meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Jackson, M.C. Interactions among multiple invasive animals. Ecology 2015, 96, 2035–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauschert, E.S.J.; Shea, K. Competition between similar invasive species: Modeling invasional interference across a landscape. Popul. Ecol. 2017, 59, 79–88. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, W.; Li, Z.; Zhu, S. Molecular Identification of a ‘Candidatus Phytoplasma ziziphi’-related Strain Infecting Amaranth (Amaranthus retroflexus L.) in China. J. Phytopathol. 2011, 159, 635–637. [Google Scholar] [CrossRef]
- Rauschert, E.S.J.; Shea, K. Invasional interference due to similar inter- and intraspecific competition between invaders may affect management. Ecol. Appl. 2012, 22, 1413–1420. [Google Scholar] [CrossRef]
- Wundrow, E.J.; Carrillo, J.; Gabler, C.A.; Horn, K.C.; Siemann, E. Facilitation and competition among invasive plants: A field experiment with alligatorweed and water hyacinth. PLoS ONE 2012, 7, e48444. [Google Scholar] [CrossRef] [Green Version]
- Zhi, Y.; Li, H.; An, S.; Zhao, L.; Zhou, C.; Deng, Z. Inter-specific competition: Spartina alterniflora is replacing Spartina anglica in coastal China. Estuar. Coast. Shelf Sci. 2007, 74, 437–448. [Google Scholar] [CrossRef]
- Gong, L.; Li, J.S.; Liu, X.Y.; Zhao, X.J.; Zhao, C.Y. Analysis of invasive alien species in Chinese national nature reserves. Ecol. Sci. 2017, 36, 210–216. [Google Scholar] [CrossRef]
- Gentili, R.; Ambrosini, R.; Montagnani, C.; Caronni, S.; Citterio, S. Effect of Soil pH on the Growth, Reproductive Investment and Pollen Allergenicity of Ambrosia artemisiifolia L. Front. Plant Sci. 2018, 9, 1335. [Google Scholar] [CrossRef]
- Ziska, L.H.; Caulfield, F.A. Rising CO2 and pollen production of common ragweed (Ambrosia artemisiifolia), a known allergy-inducing species: Implications for public health. Aust. J. Plant Physiol. 2000, 27, 893–898. [Google Scholar] [CrossRef]
- Liu, X.Y.; Li, J.S.; Zhao, C.Y.; Quan, Z.Z.; Zhao, X.J.; Gong, L. Prediction of potential suitable area of Ambrosia artemisiifolia L. in China based on MaxEnt and ArcGIS. J. Plant Proteciton 2016, 43, 1041–1048. [Google Scholar] [CrossRef]
- Valerio, M.; Tomecek, M.B.; Lovelli, S.; Ziska, L.H. Quantifying the effect of drought on carbon dioxide-induced changes in competition between a C3 crop (tomato) and a C4 weed (Amaranthus retroflexus). Weed Res. 2011, 51, 591–600. [Google Scholar] [CrossRef]
- Rezaie, F.; Yarnia, M. Allelopathic effects of Chenopodium album, Amaranthus retroflexus and Cynodon dactylon on germination and growth of safflower. J. Food Agric. Environ. 2009, 7, 516–521. [Google Scholar]
- Wang, C.; Zhou, J.; Liu, J.; Jiang, K. Differences in functional traits between invasive and native Amaranthus species under different forms of N deposition. Naturwissenschaften 2017, 104, 59. [Google Scholar] [CrossRef]
- Ma, J.; Xing, G.; Yang, W.; Ma, L.; Gao, M.; Wang, Y.; Han, Y. Inhibitory effects of leachate from Eupatorium adenophorum on germination and growth of Amaranthus retroflexus and Chenopodium glaucum. Acta Ecol. Sin. 2012, 32, 50–56. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.E.; Jiang, Y.P.; Wei, H.; Wang, F.G.; Lu, X.N.; Clements, D. Invasion process and potential spread of Amaranthus retroflexus in China. Weed Res. 2018, 58, 57–67. [Google Scholar] [CrossRef]
- Marten, G.C.; Andersen, R.N. Forage Nutritive Value and Palatability of 12 Common Annual Weeds1. Crop Sci. 1975, 15, 821–827. [Google Scholar] [CrossRef]
- Lei, T.; Cui, G.F.; Sha, H.F.; Li, F.L.; Yan, J.H. Diversity and community distribution characteristics of alien vascular plant species in wetlands of Beijing. J. Beijign For. Univ. 2010, 32, 51–57. [Google Scholar] [CrossRef]
- IPCC. IPCC Fourth Assessment Report: Climate Change 2007. Available online: https://www.ipcc.ch/assessment-report/ar4/ (accessed on 9 October 2022).
- Galloway, J.; Townsend, A.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.; Martinelli, L.; Seitzinger, S.; Sutton, M. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Wang, J.; Sang, W. Effects of nitrogen deposition on invasive and competitive abilities of an alien plant Ambrosia artemisiifolia. J. Northeast Univ. 2012, 40, 60–66. [Google Scholar] [CrossRef]
- Williams, A.C.; McCarthy, B.C. A new index of interspecific competition for replacement and additive designs. Ecol. Res. 2001, 16, 29–40. [Google Scholar] [CrossRef]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Skalova, H.; Jarosik, V.; Dvorackova, S.; Pysek, P. Effect of Intra- and Interspecific Competition on the Performance of Native and Invasive Species of Impatiens under Varying Levels of Shade and Moisture. PLoS ONE 2013, 8, e62842. [Google Scholar] [CrossRef]
- Wang, C.; Cheng, H.; Wei, M.; Wang, S.; Wu, B.; Du, D. Plant height and leaf size: Which one is more important in affecting the successful invasion of Solidago canadensis and Conyza canadensis in urban ecosystems? Urban For. Urban Green. 2021, 59, 127033. [Google Scholar] [CrossRef]
- Rajcan, I.; AghaAlikhani, M.; Swanton, C.J.; Tollenaar, M. Development of redroot pigweed is influenced by light spectral quality and quantity. Crop Sci. 2002, 42, 1930–1936. [Google Scholar] [CrossRef]
- Chu, C.-C.; Ludford, P.M.; Ozbun, J.L.; Sweet, R.D. Effects of Temperature and Competition on the Establishment and Growth of Redroot Pigweed and Common Lambsquarters. Crop Sci. 1978, 18, 308–310. [Google Scholar] [CrossRef]
- Belote, R.T.; Weltzin, J.F. Interactions between two co-dominant, invasive plants in the understory of a temperate deciduous forest. Biol. Invasions 2006, 8, 1629–1641. [Google Scholar] [CrossRef]
- Griffen, B.D.; Guy, T.; Buck, J.C. Inhibition between invasives: A newly introduced predator moderates the impacts of a previously established invasive predator. J. Anim. Ecol. 2008, 77, 32–40. [Google Scholar] [CrossRef]
- Blackshaw, R.E.; Brandt, R.N.; Janzen, H.H.; Entz, T.; Grant, C.A.; Derksen, D.A. Differential response of weed species to added nitrogen. Weed Sci. 2003, 51, 532–539. [Google Scholar] [CrossRef]
- Blumenthal, D.M.; Kray, J.A.; Ortmans, W.; Ziska, L.H.; Pendall, E. Cheatgrass is favored by warming but not CO2 enrichment in a semi-arid grassland. Glob. Chang. Biol. 2016, 22, 3026–3038. [Google Scholar] [CrossRef] [PubMed]
- LeskovŠEk, R.; Datta, A.; Knezevic, S.Z.; SimonČIČ, A. Common ragweed (Ambrosia artemisiifolia) dry matter allocation and partitioning under different nitrogen and density levels. Weed Biol. Manag. 2012, 12, 98–108. [Google Scholar] [CrossRef]
- Kimball, B.A.; Kobayashi, K.; Bindi, M. Responses of agricultural crops to free-air CO2 enrichment. In Advances in Agronomy; Sparks, D.L., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2002; Volume 77, pp. 293–368. [Google Scholar]
- Keser, L.H.; Dawson, W.; Song, Y.-B.; Yu, F.-H.; Fischer, M.; Dong, M.; van Kleunen, M. Invasive clonal plant species have a greater root-foraging plasticity than non-invasive ones. Oecologia 2014, 174, 1055–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keser, L.H.; Visser, E.J.; Dawson, W.; Song, Y.B.; Yu, F.H.; Fischer, M.; Dong, M.; van Kleunen, M. Herbaceous plant species invading natural areas tend to have stronger adaptive root foraging than other naturalized species. Front. Plant Sci. 2015, 6, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant Physiol. 2000, 27, 595–607. [Google Scholar] [CrossRef]
- Ward, J.K.; Tissue, D.T.; Thomas, R.B.; Strain, B.R. Comparative responses of model C3 and C4 plants to drought in low and elevated CO2. Glob. Change Biol. 1999, 5, 857–867. [Google Scholar] [CrossRef]
- Weller, S.; Florentine, S.; Javaid, M.M.; Welgama, A.; Chadha, A.; Chauhan, B.S.; Turville, C. Amaranthus retroflexus L. (Redroot Pigweed): Effects of Elevated CO2 and Soil Moisture on Growth and Biomass and the Effect of Radiant Heat on Seed Germination. Agron. Basel 2021, 11, 728. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Ehleringer, J. Comparative ecophysiology of C3 and C4 plants. Plant Cell Environ. 1984, 7, 1–13. [Google Scholar] [CrossRef]
- Poorter, H. Interspecific Variation in the growth-response of plants to an elevated ambient CO2 concentration. Vegetatio 1993, 104, 77–97. [Google Scholar] [CrossRef]
- Bae, J.; Byun, C.; Gyong, A.Y.; Choi, J.H.; Lee, D.; Kang, H. Effect of elevated atmospheric carbon dioxide on the allelopathic potential of common ragweed. J. Ecol. Environ. 2019, 43, 212–218. [Google Scholar] [CrossRef]
- Lonsdale, W.M. Global patterns of plant invasions and the concept of invasibility. Ecology 1999, 80, 1522–1536. [Google Scholar] [CrossRef]
- Leskovšek, R.; Eler, K.; Batič, F.; Simončič, A. The influence of nitrogen, water and competition on the vegetative and reproductive growth of common ragweed (Ambrosia artemisiifolia L.). Plant Ecol. 2012, 213, 769–781. [Google Scholar] [CrossRef]
- Amini, R.; Alizadeh, H.; Yousefi, A. Interference between red kidneybean (Phaseolus vulgaris L.) cultivars and redroot pigweed (Amaranthus retroflexus L.). Eur. J. Agron. 2014, 60, 13–21. [Google Scholar] [CrossRef]
- Golivets, M.; Wallin, K.F. Neighbour tolerance, not suppression, provides competitive advantage to non-native plants. Ecol. Lett. 2018, 21, 745–759. [Google Scholar] [CrossRef] [Green Version]
- HilleRisLambers, J.; Harpole, W.S.; Tilman, D.; Knops, J.; Reich, P.B. Mechanisms responsible for the positive diversity-productivity relationship in Minnesota grasslands. Ecol. Lett. 2004, 7, 661–668. [Google Scholar] [CrossRef]
Figure 1.
Experimental design in this study.

Figure 2.
Effects of elevated CO2 (ambient CO2, left; elevated CO2, right), increased N and replacement levels on the growth characteristics of two invasive alien plants. Bars show mean ± SD (4). (a,b). Basal stem diameter. (c,d) Height. (e,f) RY. On the horizontal axis, 1:3 refers to one common ragweed plant and three redroot pigweed plants, 2:2 refers to two common ragweed plants and two redroot pigweed plants, 3:1 refers to three common ragweed plants and one redroot pigweed plant, and 0:4 and 4:0 refer to four redroot pigweed plants or four common ragweed plants, respectively.
Figure 2.
Effects of elevated CO2 (ambient CO2, left; elevated CO2, right), increased N and replacement levels on the growth characteristics of two invasive alien plants. Bars show mean ± SD (4). (a,b). Basal stem diameter. (c,d) Height. (e,f) RY. On the horizontal axis, 1:3 refers to one common ragweed plant and three redroot pigweed plants, 2:2 refers to two common ragweed plants and two redroot pigweed plants, 3:1 refers to three common ragweed plants and one redroot pigweed plant, and 0:4 and 4:0 refer to four redroot pigweed plants or four common ragweed plants, respectively.

Figure 3.
Effects of elevated CO2 (ambient CO2, left; elevated CO2, right), N addition and replacement levels on the biomass of two invasive alien plants. Bars show mean ± SD (4). (a,b). Total biomass. (c,d). Root biomass. (e,f). Root–shoot ratio. On the horizontal axis, 1:3 refers to one common ragweed plant and three redroot pigweed plants, 2:2 refers to two common ragweed plants and two redroot pigweed plants, 3:1 refers to three common ragweed plants and one redroot pigweed plant, and 0:4 and 4:0 refer to four redroot pigweed plants or four common ragweed plants, respectively.
Figure 3.
Effects of elevated CO2 (ambient CO2, left; elevated CO2, right), N addition and replacement levels on the biomass of two invasive alien plants. Bars show mean ± SD (4). (a,b). Total biomass. (c,d). Root biomass. (e,f). Root–shoot ratio. On the horizontal axis, 1:3 refers to one common ragweed plant and three redroot pigweed plants, 2:2 refers to two common ragweed plants and two redroot pigweed plants, 3:1 refers to three common ragweed plants and one redroot pigweed plant, and 0:4 and 4:0 refer to four redroot pigweed plants or four common ragweed plants, respectively.

Figure 4.
Effects of elevated CO2, N addition and replacement levels on the reproductive characteristics of common ragweed. Bars show mean ± SD (4). (a,b) The number of seeds (c,d) mean weight of seeds. On the horizontal axis, 1:3 refers to one common ragweed plant and three redroot pigweed plants, 2:2 refers to two common ragweed plants and two redroot pigweed plants, 3:1 refers to three common ragweed plants and one redroot pigweed plant, and 4:0 refers to four common ragweed plants.
Figure 4.
Effects of elevated CO2, N addition and replacement levels on the reproductive characteristics of common ragweed. Bars show mean ± SD (4). (a,b) The number of seeds (c,d) mean weight of seeds. On the horizontal axis, 1:3 refers to one common ragweed plant and three redroot pigweed plants, 2:2 refers to two common ragweed plants and two redroot pigweed plants, 3:1 refers to three common ragweed plants and one redroot pigweed plant, and 4:0 refers to four common ragweed plants.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of three-way ANOVA for the growth characters of the alien invasive plants common ragweed and redroot pigweed according to CO2, N and replacement levels: basal stem diameter, height, branch number.
Table 1.
Results of three-way ANOVA for the growth characters of the alien invasive plants common ragweed and redroot pigweed according to CO2, N and replacement levels: basal stem diameter, height, branch number.
| Species | Factors | Basal Stem Diameter | Height | Branch Number | |
|---|---|---|---|---|---|
| common ragweed | CO2 | F | 2.756 | 3.881 | 0.84 |
| p | 0.104 | 0.055 | 0.364 | ||
| df | 1 | 1 | 1 | ||
| N | F | 32.843 | 17.834 | 0.077 | |
| p | <0.001 ** | <0.001 ** | 0.783 | ||
| df | 1 | 1 | 1 | ||
| replacement | F | 14.961 | 2.446 | 15.668 | |
| p | <0.001 ** | 0.076 | <0.001 ** | ||
| df | 3 | 3 | 3 | ||
| CO2 × N | F | 0.102 | 3.65 | 2.025 | |
| p | 0.751 | 0.062 | 0.161 | ||
| df | 1 | 1 | 1 | ||
| N × replacement | F | 1.163 | 0.398 | 2.198 | |
| p | 0.334 | 0.755 | 0.101 | ||
| df | 3 | 3 | 3 | ||
| CO2 × replacement | F | 1.093 | 0.222 | 0.905 | |
| p | 0.361 | 0.881 | 0.446 | ||
| df | 3 | 3 | 3 | ||
| CO2 × N × replacement | F | 0.128 | 0.944 | 1.094 | |
| p | 0.943 | 0.427 | 0.361 | ||
| df | 3 | 3 | 3 | ||
| redroot pigweed | CO2 | F | 0 | 3.405 | - |
| p | 0.994 | 0.071 | - | ||
| df | 1 | 1 | |||
| N | F | 4.599 | 26.336 | - | |
| p | 0.037 * | <0.001 ** | - | ||
| df | 1 | 1 | |||
| replacement | F | 36.172 | 43.233 | - | |
| p | <0.001 ** | <0.001 ** | - | ||
| df | 3 | 3 | |||
| CO2 × N | F | 0.009 | 0.249 | - | |
| p | 0.926 | 0.62 | - | ||
| df | 1 | 1 | |||
| N × replacement | F | 2.971 | 0.834 | - | |
| p | 0.041 * | 0.482 | - | ||
| df | 3 | 3 | |||
| CO2 × replacement | F | 0.896 | 3.64 | - | |
| p | 0.45 | 0.019 * | - | ||
| df | 3 | 3 | |||
| CO2 × N × replacement | F | 0.289 | 0.712 | - | |
| p | 0.833 | 0.55 | - | ||
| df | 3 | 3 |
Df represents degrees of freedom, * represents p < 0.05, ** represents p < 0.01
Table 2.
Results of three-way ANOVA for the biomass characters of the alien invasive plants common ragweed and redroot pigweed according to CO2, N and replacement levels: shoot biomass, root biomass, total biomass, and root–shoot ratio.
Table 2.
Results of three-way ANOVA for the biomass characters of the alien invasive plants common ragweed and redroot pigweed according to CO2, N and replacement levels: shoot biomass, root biomass, total biomass, and root–shoot ratio.
| Species | Factors | Shoot Biomass | Root Biomass | Total Biomass | Root-Shoot Ratio | |
|---|---|---|---|---|---|---|
| common ragweed | CO2 | F | 2.141 | 0.855 | 2.107 | 0.042 |
| p | 0.15 | 0.36 | 0.153 | 0.839 | ||
| df | 1 | 1 | 1 | 1 | ||
| N | F | 140.242 | 73.943 | 142.018 | 2.843 | |
| p | <0.001 ** | <0.001 ** | <0.001 ** | 0.099 | ||
| df | 1 | 1 | 1 | 1 | ||
| replacement | F | 1.085 | 5.75 | 1.398 | 5.158 | |
| p | 0.365 | 0.002 ** | 0.255 | 0.004 ** | ||
| df | 3 | 3 | 3 | 3 | ||
| CO2 × N | F | 13.565 | 4.989 | 13.245 | 2.609 | |
| p | 0.001 ** | 0.030 * | 0.001 ** | 0.113 | ||
| df | 1 | 1 | 1 | 1 | ||
| N × replacement | F | 0.399 | 2.804 | 0.651 | 1.275 | |
| p | 0.754 | 0.05 | 0.587 | 0.294 | ||
| df | 3 | 3 | 3 | 3 | ||
| CO2 × replacement | F | 0.361 | 0.808 | 0.346 | 0.806 | |
| p | 0.781 | 0.496 | 0.792 | 0.497 | ||
| df | 3 | 3 | 3 | 3 | ||
| CO2 × N × replacement | F | 0.354 | 0.064 | 0.271 | 0.294 | |
| p | 0.786 | 0.979 | 0.846 | 0.83 | ||
| df | 3 | 3 | 3 | 3 | ||
| redroot pigweed | CO2 | F | 1.92 | 5.08 | 2.63 | 0.037 |
| p | 0.173 | 0.029 * | 0.112 | 0.849 | ||
| df | 1 | 1 | 1 | 1 | ||
| N | F | 1.614 | 0.013 | 1.637 | 0.383 | |
| p | 0.21 | 0.909 | 0.207 | 0.539 | ||
| df | 1 | 1 | 1 | 1 | ||
| replacement | F | 100.968 | 57.074 | 110.475 | 4.281 | |
| p | <0.001 ** | <0.001 ** | <0.001 ** | 0.010 ** | ||
| df | 3 | 3 | 3 | 3 | ||
| CO2 × N | F | 0.588 | 0.054 | 0.636 | 0.088 | |
| p | 0.447 | 0.818 | 0.429 | 0.768 | ||
| df | 1 | 1 | 1 | 1 | ||
| N × replacement | F | 0.512 | 0.182 | 0.472 | 0.184 | |
| p | 0.676 | 0.908 | 0.703 | 0.907 | ||
| df | 3 | 3 | 3 | 3 | ||
| CO2 × replacement | F | 0.491 | 0.597 | 0.464 | 1.36 | |
| p | 0.69 | 0.62 | 0.709 | 0.267 | ||
| df | 3 | 3 | 3 | 3 | ||
| CO2 × N × replacement | F | 0.033 | 0.374 | 0.027 | 0.736 | |
| p | 0.992 | 0.772 | 0.994 | 0.536 | ||
| df | 3 | 3 | 3 | 3 |
Df represents degrees of freedom, * represents p < 0.05, ** represents p < 0.01
Table 3.
Results of three-way ANOVA for the seed characters of the alien invasive plants common ragweed according to CO2, N, and replacement levels: seed yield, seed mean weight.
Table 3.
Results of three-way ANOVA for the seed characters of the alien invasive plants common ragweed according to CO2, N, and replacement levels: seed yield, seed mean weight.
| Factors | Seed Number | Seed Total Weight | Seed Mean Weight | |
|---|---|---|---|---|
| CO2 | F | 0.017 | 0.344 | 0.181 |
| p | 0.896 | 0.560 | 0.673 | |
| df | 1 | 1 | 1 | |
| N | F | 14.821 | 1.241 | 12.024 |
| p | <0.001 ** | 0.271 | 0.001 ** | |
| df | 1 | 1 | 1 | |
| replacement | F | 2.316 | 2.135 | 0.330 |
| p | 0.088 | 0.109 | 0.804 | |
| df | 3 | 3 | 3 | |
| CO2 × N | F | 1.654 | 0.999 | 0.092 |
| p | 0.205 | 0.323 | 0.763 | |
| df | 1 | 1 | 1 | |
| N × replacement | F | 0.437 | 0.926 | 0.689 |
| p | 0.727 | 0.436 | 0.563 | |
| df | 3 | 3 | 3 | |
| CO2 × replacement | F | 0.806 | 0.360 | 0.083 |
| p | 0.497 | 0.782 | 0.969 | |
| df | 3 | 3 | 3 | |
| CO2 × N × replacement | F | 1.203 | 0.977 | 1.885 |
| p | 0.319 | 0.412 | 0.145 | |
| df | 3 | 3 | 3 |
Df represents degrees of freedom, * represents p < 0.05, ** represents p < 0.01
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, C.; Zhao, X.; Li, J. Elevated CO2 and Increased N Intensify Competition between Two Invasive Annual Plants in China. Life 2022, 12, 1669. https://doi.org/10.3390/life12101669
AMA Style
Zhao C, Zhao X, Li J. Elevated CO2 and Increased N Intensify Competition between Two Invasive Annual Plants in China. Life. 2022; 12(10):1669. https://doi.org/10.3390/life12101669
Chicago/Turabian StyleZhao, Caiyun, Xiangjian Zhao, and Junsheng Li. 2022. "Elevated CO2 and Increased N Intensify Competition between Two Invasive Annual Plants in China" Life 12, no. 10: 1669. https://doi.org/10.3390/life12101669
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.