
Rescue IVM of Denuded GV- and MI-Stage Oocytes of Premenopausal Rats with Oncostatin M, Insulin-like Growth Factor I, and Growth Hormone

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
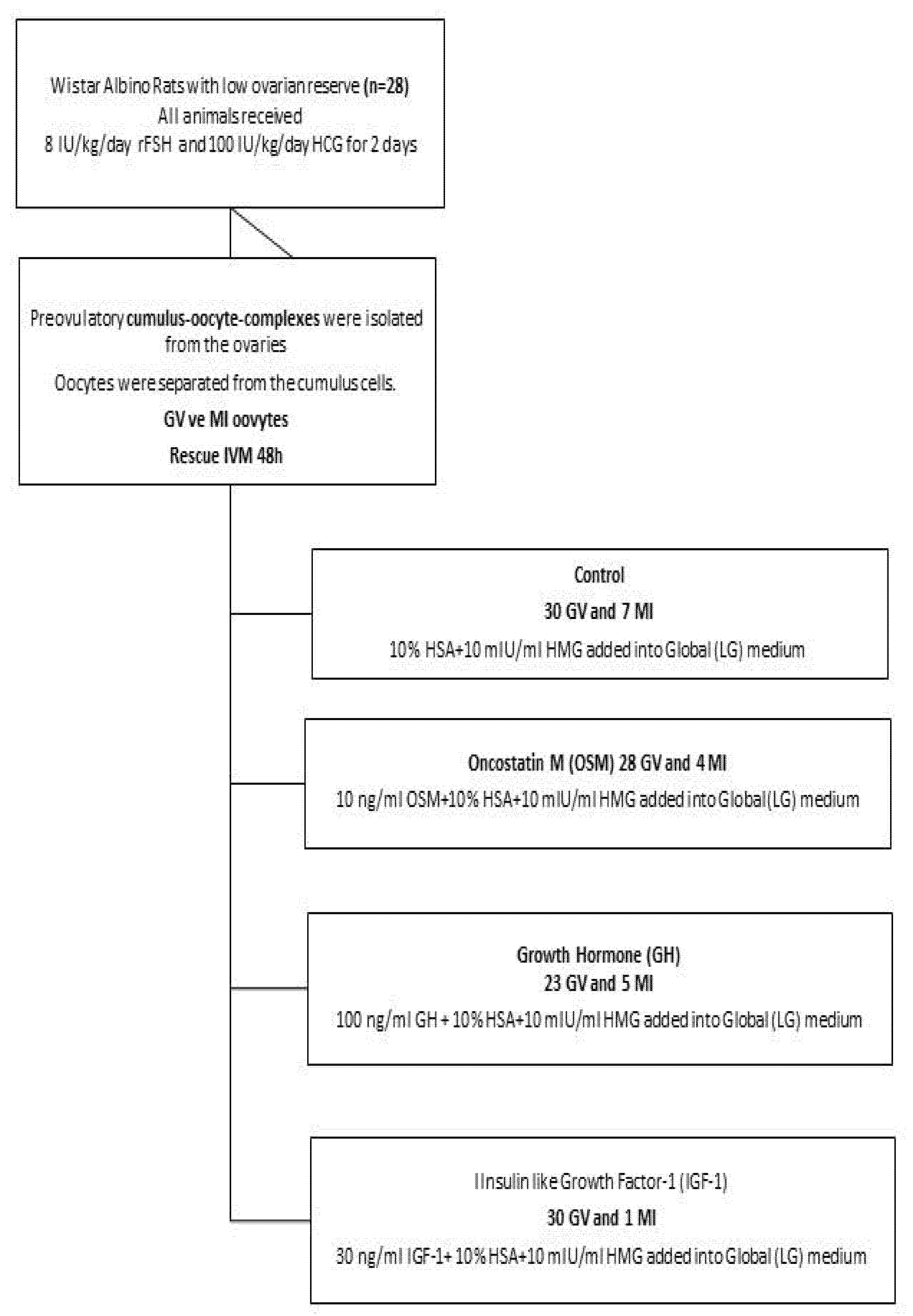
2.1. Animals and Oocyte Recovery
2.2. In Vitro Maturation of Oocytes
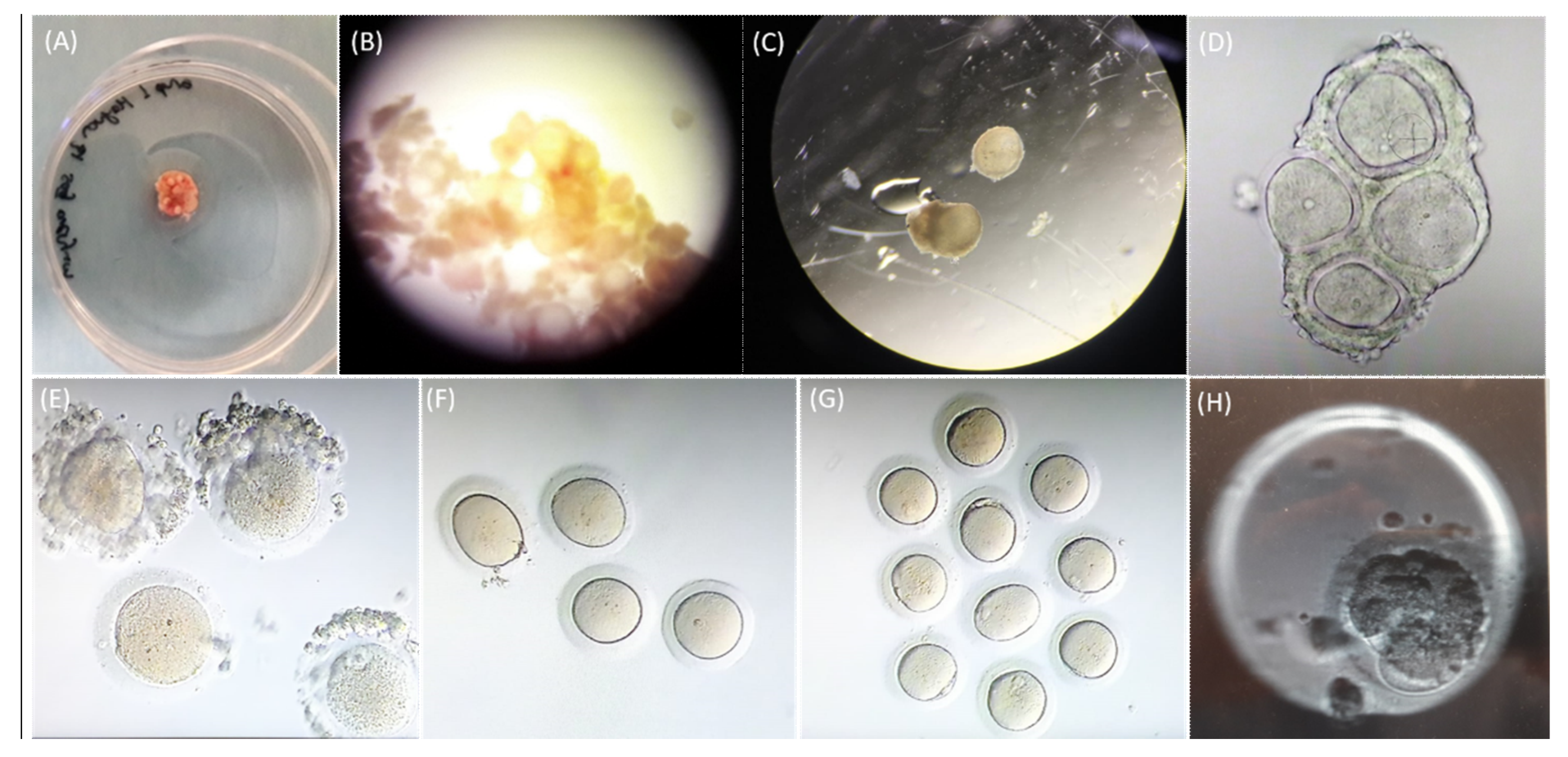
2.3. Assessment of Oocyte Maturation
2.4. Measurement of FSH and Anti-Müllerian Hormone (AMH)
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arroyo, A.; Kim, B.; Yeh, J. Luteinizing hormone action in human oocyte maturation and quality: Signaling pathways, regulation, and clinical impact. Reprod. Sci. 2020, 27, 1223–1252. [Google Scholar] [CrossRef] [PubMed]
- Braga, D.; Zanetti, B.F.; Setti, A.S.; Iaconelli, A., Jr.; Borges, E., Jr. Immature oocyte incidence: Contributing factors and effects on mature sibling oocytes in intracytoplasmic sperm injection cycles. JBRA Assist. Reprod. 2020, 24, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Halvaei, I.; Ali Khalili, M.; Razi, M.H.; Nottola, S.A. The effect of immature oocytes quantity on the rates of oocytes maturity and morphology, fertilization, and embryo development in ICSI cycles. J. Assist. Reprod. Genet. 2012, 29, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Parrella, A.; Irani, M.; Keating, D.; Chow, S.; Rosenwaks, Z.; Palermo, G.D. High proportion of immature oocytes in a cohort reduces fertilization, embryo development, pregnancy and live birth rates following ICSI. Reprod. Biomed. Online 2019, 39, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Borges, E., Jr.; Zanetti, B.F.; Setti, A.S.; Braga, D.P.; Figueira, R.d.C.S.; Iaconelli, A., Jr. FSH dose to stimulate different patient’ ages: When less is more. JBRA Assist. Reprod. 2017, 21, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Ashourzadeh, S.; Khalili, M.A.; Omidi, M.; Mahani, S.N.N.; Kalantar, S.M.; Aflatoonian, A.; Habibzadeh, V. Noninvasive assays of in vitro matured human oocytes showed insignificant correlation with fertilization and embryo development. Arch. Gynecol. Obstet. 2015, 292, 459–463. [Google Scholar] [CrossRef]
- Jie, H.; Zhao, M.; Alqawasmeh, O.A.M.; Chan, C.P.S.; Lee, T.L.; Li, T.; Chan, D.Y.L. In vitro rescue immature oocytes—A literature review. Hum. Fertil. 2021, 1–20. [Google Scholar] [CrossRef]
- Palmerini, M.G.; Antonouli, S.; Macchiarelli, G.; Cecconi, S.; Bianchi, S.; Khalili, M.A.; Nottola, S.A. Ultrastructural evaluation of the human oocyte at the germinal vesicle stage during the application of assisted reproductive technologies. Cells 2022, 11, 1636. [Google Scholar] [CrossRef]
- Rienzi, L.F.; Ubaldi, F.M. Oocyte retrieval and selection. In Textbook of Assisted Reproductive Techniques; CRC Press: Florida, FL, USA, 2012; pp. 136–153. [Google Scholar]
- Edwards, R.G. Maturation in vitro of mouse, sheep, cow, pig, rhesus monkey and human ovarian oocytes. Nature 1965, 208, 349–351. [Google Scholar] [CrossRef]
- De Vos, M.; Grynberg, M.; Ho, T.M.; Yuan, Y.; Albertini, D.F.; Gilchrist, R.B. Perspectives on the development and future of oocyte IVM in clinical practice. J. Assist. Reprod. Genet. 2021, 38, 1265–1280. [Google Scholar] [CrossRef]
- Hatırnaz, Ş.; Ata, B.; Hatırnaz, E.S.; Dahan, M.H.; Tannus, S.; Tan, J.; Tan, S.L. Oocyte in vitro maturation: A sytematic review. Turk. J. Obstet. Gynecol. 2018, 15, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Barad, D.H.; Kushnir, V.A.; Shohat-Tal, A.; Lazzaroni-Tealdi, E.; Wu, Y.G.; Gleicher, N. Rescue in vitro maturation (IVM) of immature oocytes in stimulated cycles in women with low functional ovarian reserve (LFOR). Endocrine 2016, 52, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Son, W.-Y.; Henderson, S.; Cohen, Y.; Dahan, M.; Buckett, W. Immature Oocyte for Fertility Preservation. Front. Endocrinol. 2019, 10, 464. [Google Scholar] [CrossRef]
- Hatırnaz, Ş.; Akarsu, S.; Hatırnaz, E.S.; Işık, A.Z.; Dahan, M.H. The use of in vitro maturation in stimulated antagonist in vitro fertilization cycles of normo-hyperresponder women due to arrested follicular development: A rescue procedure. Turk. J. Obstet. Gynecol. 2018, 15, 141–146. [Google Scholar] [CrossRef]
- Madkour, A.; Bouamoud, N.; Kaarouch, I.; Louanjli, N.; Saadani, B.; Assou, S.; Aboulmaouahib, S.; Sefrioui, O.; Amzazi, S.; Copin, H.; et al. Follicular fluid and supernatant from cultured cumulus-granulosa cells improve in vitro maturation in patients with polycystic ovarian syndrome. Fertil. Steril. 2018, 110, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, H.; Yu, Q.; Liu, H.; Huang, T.; Zhao, S.; Ma, J.; Zhao, H. Growth Hormone Promotes in vitro Maturation of Human Oocytes. Front. Endocrinol. 2019, 10, 485. [Google Scholar] [CrossRef]
- Menezo, Y.J.R.; Nicollet, B.; Rollet, J.; Hazout, A. Pregnancy and delivery after in vitro maturation of naked ICSI-GV oocytes with GH and transfer of a frozen thawed blastocyst: Case report. J. Assist. Reprod. Genet. 2006, 23, 47–49. [Google Scholar] [CrossRef]
- Apa, R.; Lanzone, A.; Miceli, F.; Mastrandrea, M.; Caruso, A.; Mancuso, S.; Canipari, R. Growth hormone induces in vitro maturation of follicle- and cumulus-enclosed rat oocytes. Mol. Cell. Endocrinol. 1994, 106, 207–212. [Google Scholar] [CrossRef]
- Toori, M.A.; Mosavi, E.; Nikseresht, M.; Barmak, M.J.; Mahmoudi, R. Influence of Insulin-Like Growth Factor-I on Maturation and Fertilization Rate of Immature Oocyte and Embryo Development in NMRI Mouse with TCM199 and α-MEM Medium. J. Clin. Diagn. Res. 2014, 8, AC05–AC08. [Google Scholar] [CrossRef]
- Lorenzo, P.L.; Illera, M.J.; Illera, J.C.; Illera, M. Enhancement of cumulus expansion and nuclear maturation during bovine oocyte maturation in vitro by the addition of epidermal growth factor and insulin-like growth factor I. J. Reprod. Fertil. 1994, 101, 697–701. [Google Scholar] [CrossRef]
- Sato, A.; Sarentonglaga, B.; Ogata, K.; Yamaguchi, M.; Hara, A.; Atchalalt, K.; Sugane, N.; Fukumori, R.; Nagao, Y. Effects of insulin-like growth factor-1 on the in vitro maturation of canine oocytes. J. Reprod. Dev. 2018, 64, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Oberlender, G.; Murgas, L.D.; Zangeronimo, M.G.; da Silva, A.C.; Menezes Tde, A.; Pontelo, T.P.; Vieira, L.A. Role of insulin-like growth factor-I and follicular fluid from ovarian follicles with different diameters on porcine oocyte maturation and fertilization in vitro. Theriogenology 2013, 80, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.M.; De, M.D.G.; Jackson, J.A.; Palmer, S.S.; Tran, C.A.T. Use of IL-6 Type Cytokines to Mature. Oocytes. Patent DK2325201T3, 26 November 2004. [Google Scholar]
- Nikanfar, S.; Hamdi, K.; Haiaty, S.; Samadi, N.; Shahnazi, V.; Fattahi, A.; Nouri, M. Oncostatin M and its receptor in women with polycystic ovary syndrome and association with assisted reproductive technology outcomes. Reprod. Biol. 2022, 22, 100633. [Google Scholar] [CrossRef] [PubMed]
- Abir, R.; Ao, A.; Jin, S.; Barnett, M.; Van den Hurk, R.; Freimann, S.; Fisch, B. Immunocytochemical detection and reverse transcription polymerase chain reaction expression of oncostatin M (OSM) and its receptor (OSM-Rbeta) in human fetal and adult ovaries. Fertil. Steril. 2005, 83 (Suppl. 1), 1188–1196. [Google Scholar] [CrossRef]
- Lacoste, C.R.; Clemenson, A.; Lima, S.; Lecointre, R.; Peoc’h, M.; Chene, G. Tubo-ovarian dysplasia in relationship with ovulation induction in rats. Fertil. Steril. 2013, 99, 1768–1773. [Google Scholar] [CrossRef]
- Ozcan, Z.; Celik, H.; Gurates, B.; Ozercan, H.I.; Hanay, F.; Nalbant, M.; Dogan, Z. Effects of ovulation induction agents on ovarian surface epithelium in rats. Reprod. Biomed. Online 2009, 19, 314–318. [Google Scholar] [CrossRef]
- Taketsuru, H.; Kaneko, T. In vitro maturation of immature rat oocytes under simple culture conditions and subsequent developmental ability. J. Reprod. Dev. 2016, 62, 521–526. [Google Scholar] [CrossRef]
- Picut, C.A.; Dixon, D.; Simons, M.L.; Stump, D.G.; Parker, G.A.; Remick, A.K. Postnatal ovary development in the rat: Morphologic study and correlation of morphology to neuroendocrine parameters. Toxicol. Pathol. 2015, 43, 343–353. [Google Scholar] [CrossRef]
- Pastuschek, J.; Poetzsch, J.; Morales-Prieto, D.M.; Schleußner, E.; Markert, U.R.; Georgiev, G. Stimulation of the JAK/STAT pathway by LIF and OSM in the human granulosa cell line COV434. J. Reprod. Immunol. 2015, 108, 48–55. [Google Scholar] [CrossRef]
- Jimenez, C.R.; de Azevedo, J.L.; Silveira, R.G.; Penitente-Filho, J.; Carrascal-Triana, E.L.; Zolini, A.M.; Araujo, V.R.; Torres, C.; Gonçalves, W.G. Effects of IGF-1 on In Vitro Culture of Bovine Preantral Follicles are Dose-Dependent. Reprod. Domest. Anim. Zuchthyg. 2016, 51, 435–444. [Google Scholar] [CrossRef]
- Kiapekou, E.; Loutradis, D.; Drakakis, P.; Zapanti, E.; Mastorakos, G.; Antsaklis, A. Effects of GH and IGF-I on the in vitro maturation of mouse oocytes. Hormones 2005, 4, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Faramarzi, A.; Khalili, M.A.; Ashourzadeh, S.; Palmerini, M.G. Does rescue in vitro maturation of germinal vesicle stage oocytes impair embryo morphokinetics development? Zygote 2018, 26, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Alcoba, D.D.; Pimentel, A.M.; Brum, I.S.; Corleta, H.E. Developmental potential of in vitro or in vivo matured oocytes. Zygote 2015, 23, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.A.; Azab, H.; Rahman, A.A.; Nafee, T.M. Clinical Assisted Reproduction: Effects of Growth Hormone on in Vitro Maturation of Germinal Vesicle of Human Oocytes Retrieved from Small Antral Follicles. J. Assist. Reprod. Genet. 2001, 18, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Benkhalifa, M.; Madkour, A.; Louanjli, N.; Bouamoud, N.; Saadani, B.; Kaarouch, I.; Chahine, H.; Sefrioui, O.; Merviel, P.; Copin, H. From global proteome profiling to single targeted molecules of follicular fluid and oocyte: Contribution to embryo development and IVF outcome. Expert Rev. Proteom. 2015, 12, 407–423. [Google Scholar] [CrossRef]
- Rajabi, Z.; Khokhar, Z.; Yazdekhasti, H. The Growth of Preantral Follicles and the Impact of Different Supplementations and Circumstances: A Review Study with Focus on Bovine and Human Preantral Follicles. Cell. Reprogram. 2018, 20, 164–177. [Google Scholar] [CrossRef]
- Ménézo, Y.J.; el Mouatassim, S.; Chavrier, M.; Servy, E.J.; Nicolet, B. Human oocytes and preimplantation embryos express mRNA for growth hormone receptor. Zygote 2003, 11, 293–297. [Google Scholar] [CrossRef]
- Lobie, P.E.; Breipohl, W.; Waters, M.J. Growth hormone receptor expression in the rat gastrointestinal tract. Endocrinology 1990, 126, 299–306. [Google Scholar] [CrossRef]
- Zhao, J.; Taverne, M.A.; van der Weijden, G.C.; Bevers, M.M.; van den Hurk, R. Immunohistochemical localisation of growth hormone (GH), GH receptor (GHR), insulin-like growth factor I (IGF-I) and type I IGF-I receptor, and gene expression of GH and GHR in rat pre-antral follicles. Zygote 2002, 10, 85–94. [Google Scholar] [CrossRef]
- Izadyar, F.; Van Tol, H.T.; Colenbrander, B.; Bevers, M.M. Stimulatory effect of growth hormone on in vitro maturation of bovine oocytes is exerted through cumulus cells and not mediated by IGF-I. Mol. Reprod. Dev. 1997, 47, 175–180. [Google Scholar] [CrossRef]
- Iga, K.; Niwa, K.; Bartke, A. Recombinant Bovine Growth Hormone Stimulates Nuclear Maturation of Bovine Oocytes In Vitro and Promotes Subsequent Embryonic Development. J. Reprod. Dev. 1998, 44, 45–52. [Google Scholar] [CrossRef]
- Eddie, S.L.; Childs, A.J.; Jabbour, H.N.; Anderson, R.A. Developmentally regulated IL6-type cytokines signal to germ cells in the human fetal ovary. Mol. Hum. Reprod. 2011, 18, 88–95. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Control Group | +OSM | +IGF-1 | +GH | p | |
|---|---|---|---|---|---|
| Weight (g) | 324.28 ± 13.94 | 330.00 ± 3.08 | 344.28 ± 17.30 | 320.00 ± 7.86 | 0.405 |
| Right ovarian weight (g) | 106.61 ± 9.91 | 98.45 ± 9.09 | 93.47 ± 15.22 | 95.62 ± 9.03 | 0.580 |
| Left ovarian weight (g) | 105.82 ± 7.53 | 91.68 ± 9.11 | 78.31 ± 6.96 | 92.35 ± 8.19 | 0.237 |
| FSH (mIU/mL) | 7.75 ± 0.56 | 7.07 ± 0.25 | 7.81 ± 0.37 | 8.75 ± 0.52 | 0.103 |
| AMH (ng/mL) | 7.20 ± 0.31 | 7.35 ± 0.38 | 7.81 ± 0.43 | 8.06 ± 0.32 | 0.515 |
| No. of GV Oocytes | No. (%) of Oocytes after Rescue IVM | |||
|---|---|---|---|---|
| GV→GV | GV→MI | GV→MII | ||
| Control group | 30 | 8 (26.6%) | 5 (16.6) | 17 (56.6%) |
| +OSM | 28 | 7 (25%) | 6 (21.4%) | 15 (53.5%) |
| +IGF-1 | 30 | 18 (60%) | 2 (6.6%) | 10 (33.3%) |
| +GH | 23 | 16 (69.5%) | 0 | 7 (30.3%) |
| p | 0.001 | 0.001 | 0.001 | |
| No. of MI Oocytes | No. (%) of Oocytes after Rescue IVM | ||
|---|---|---|---|
| MI→MI | MI→MII | ||
| Control group | 7 | 0 | 7 (100%) |
| +OSM | 4 | 0 | 4 (100%) |
| +IGF-1 | 1 | 0 | 1 (100%) |
| +GH | 5 | 4 (80%) | 1 (20%) |
| p | 0.004 | 0.004 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akdemir, Y.; Donmez Cakil, Y.; Selam, B.; Sitar, M.E.; Cincik, M. Rescue IVM of Denuded GV- and MI-Stage Oocytes of Premenopausal Rats with Oncostatin M, Insulin-like Growth Factor I, and Growth Hormone. Life 2022, 12, 1247. https://doi.org/10.3390/life12081247
Akdemir Y, Donmez Cakil Y, Selam B, Sitar ME, Cincik M. Rescue IVM of Denuded GV- and MI-Stage Oocytes of Premenopausal Rats with Oncostatin M, Insulin-like Growth Factor I, and Growth Hormone. Life. 2022; 12(8):1247. https://doi.org/10.3390/life12081247
Chicago/Turabian StyleAkdemir, Yesim, Yaprak Donmez Cakil, Belgin Selam, Mustafa Erinc Sitar, and Mehmet Cincik. 2022. "Rescue IVM of Denuded GV- and MI-Stage Oocytes of Premenopausal Rats with Oncostatin M, Insulin-like Growth Factor I, and Growth Hormone" Life 12, no. 8: 1247. https://doi.org/10.3390/life12081247