


Remote Sensing and GIS-Based Suitability Mapping of Termite Habitat in the African Savanna: A Case Study of the Lowveld in Kruger National Park


Abstract
:1. Introduction
2. Material and Methods
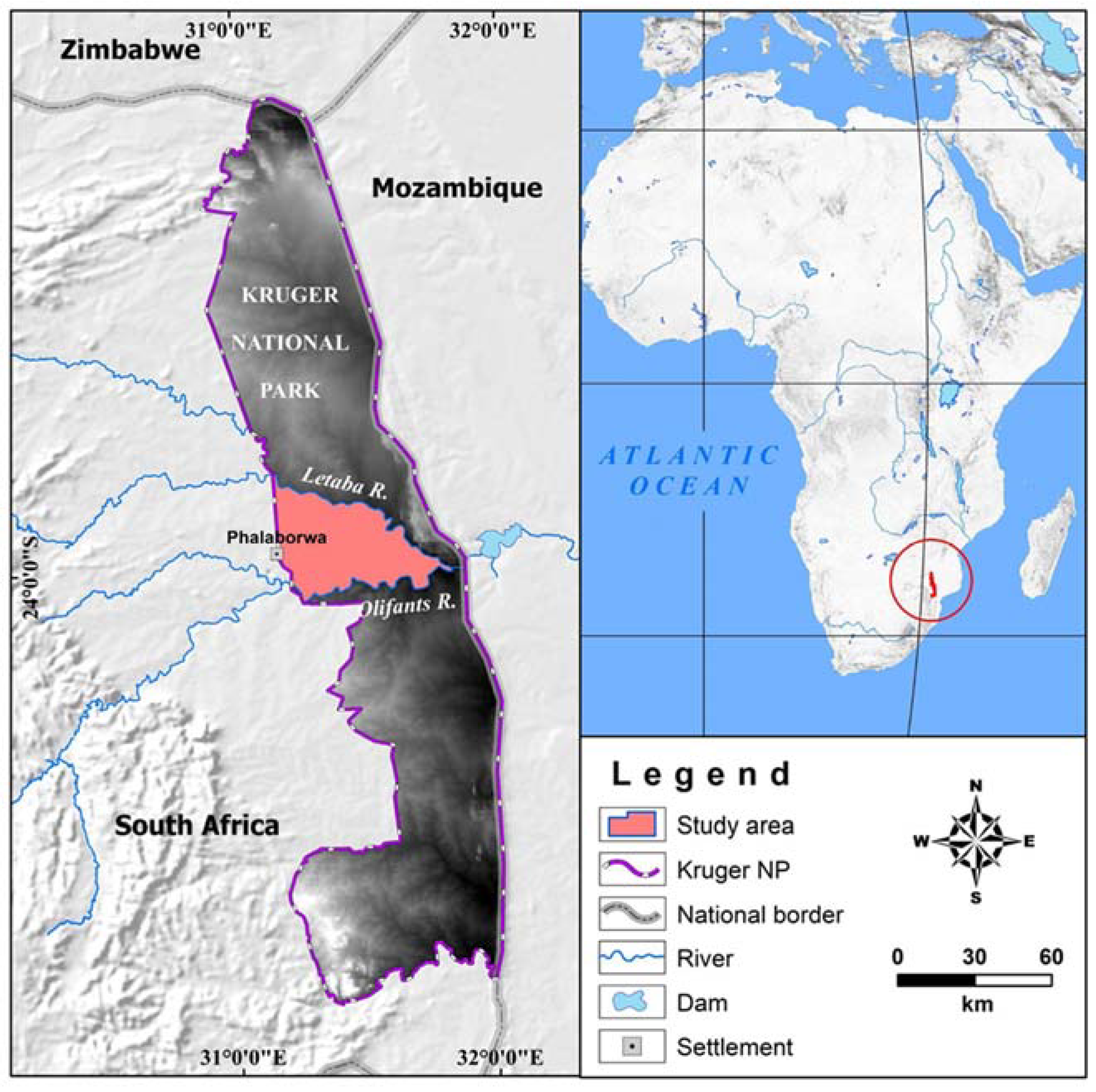
2.1. Study Area
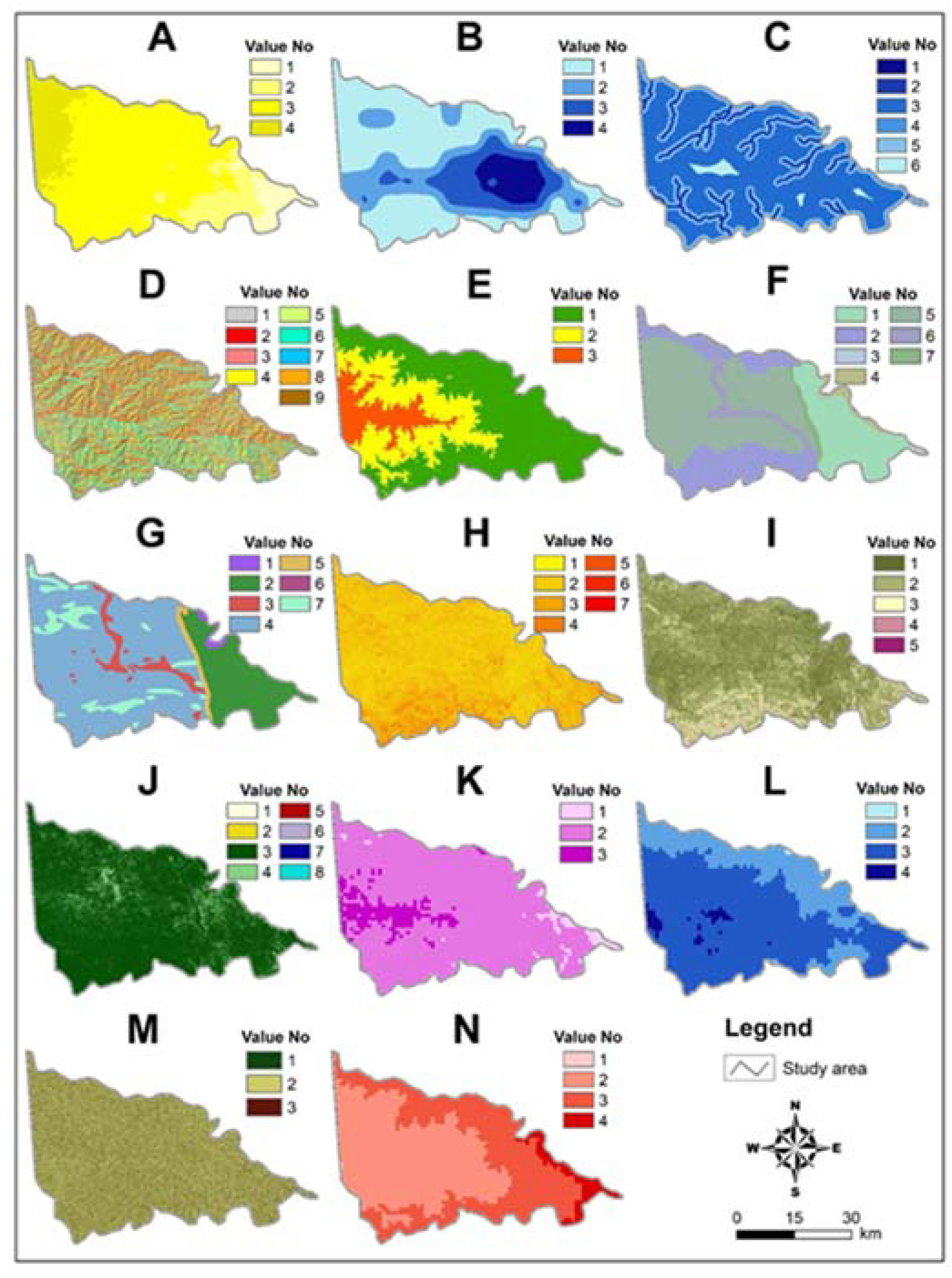
2.2. Material
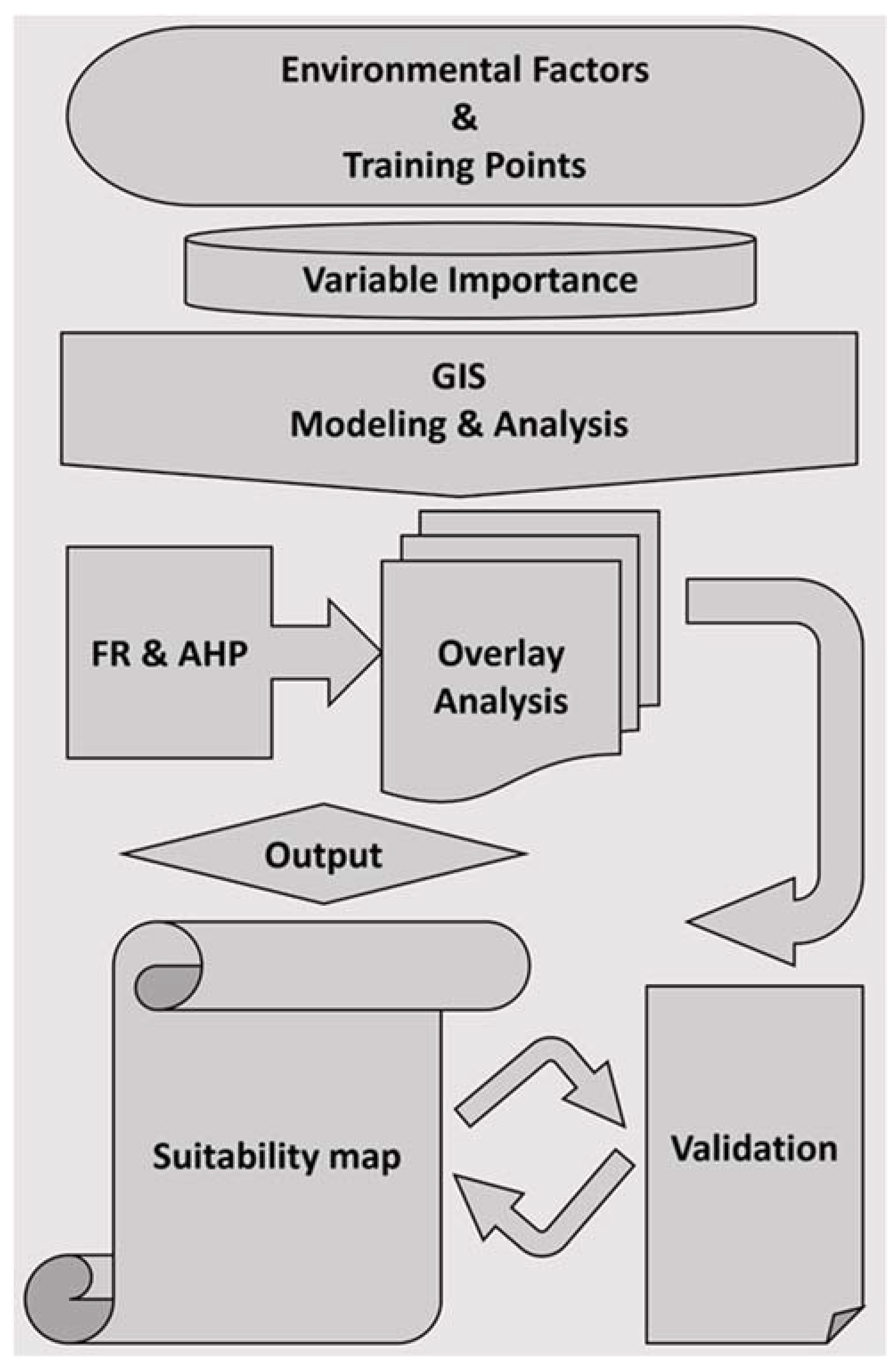
2.3. Methods
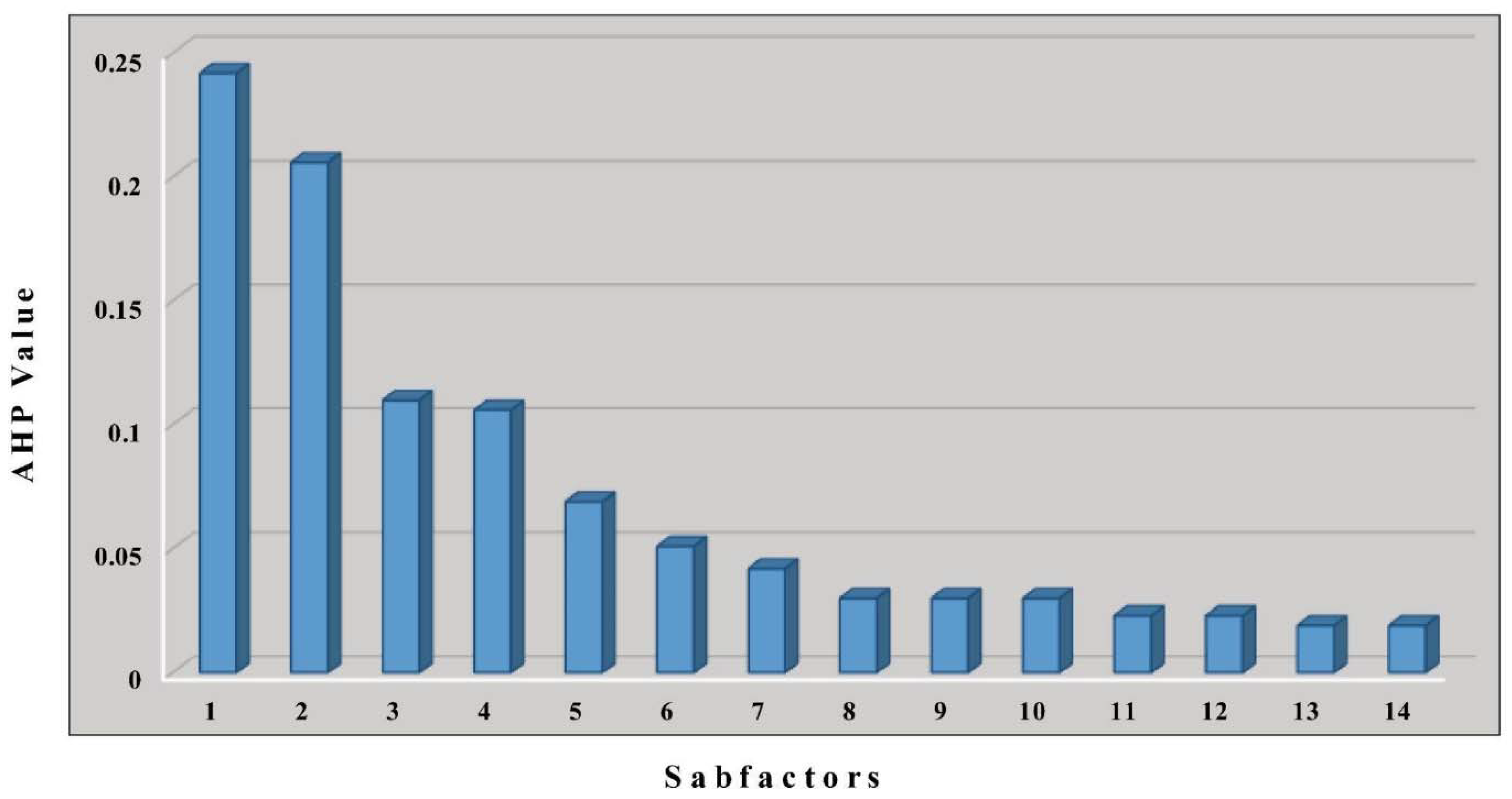
2.3.1. Variable Importance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Factors | Data Type | Sources | Var. Importance (%) |
|---|---|---|---|---|
| 1 | Solar radiation | DEM/Grid | Fick and Hijmans, 2017 [67] | 15 |
| 2 | Groundwater depth | Raster | MacDonald et al., 2012 [68] | 14 |
| 3 | Distance to the Drainage | Polyline | Andreadis et al., 2013 [69] | 13 |
| 4 | Aspect | DEM/Grid | ASF DAAC, 2015 [70] | 12 |
| 5 | Elevation | DEM/Grid | ASF DAAC, 2015 [70] | 9 |
| 6 | Soil | Polygon | Viljoen, 2015 [54] | 8 |
| 7 | Lithology | Polygon | Viljoen, 2015 [54] | 6 |
| 8 | Slope | DEM/Grid | ASF DAAC, 2015 [70] | 5 |
| 9 | Topographic relief | DEM/Grid | ASF DAAC, 2015 [70] | 4 |
| 10 | Land cover | Raster | SAPAD, 2020 [71] | 4 |
| 11 | Wind speed | DEM/Grid | Fick and Hijmans, 2017 [67] | 3 |
| 12 | Precipitation | DEM/Grid | Fick and Hijmans, 2017 [67] | 3 |
| 13 | Slope Position | DEM/Grid | ASF DAAC, 2015 [70] | 2 |
| 14 | Temperature | DEM/Grid | ASF DAAC, 2015 [70] | 2 |
2.3.2. Frequency Ratio (FR)
| Factor No | Factor | Value No | Variables/Subfactors | FR Value |
|---|---|---|---|---|
| 1 | Solar radiation (MJ m−2 day−1) | 1 | <17.500 | 0.00 |
| 2 | 17.500–17.750 | 0.00 | ||
| 3 | 17.750–18.000 | 1.27 | ||
| 4 | 18.000-> | 0.96 | ||
| 2 | Groundwater depth (m) | 1 | Very shallow (0–25 m) | 0.57 |
| 2 | Shallow (25–50 m) | 2.00 | ||
| 3 | Mid (50–100 m) | 0.95 | ||
| 4 | Deep (100-> m) | 0.00 | ||
| 3 | Distance to drainage network (m) | 1 | <100 | 0.37 |
| 2 | 100–250 | 0.60 | ||
| 3 | 250–500 | 0.80 | ||
| 4 | 500–1000 | 0.80 | ||
| 5 | 1000–5000 | 1.10 | ||
| 6 | 5000> | 2.36 | ||
| 4 | Aspect | 1 | Flat | 0.98 |
| 2 | North | 0.82 | ||
| 3 | Northeast | 0.78 | ||
| 4 | East | 0.93 | ||
| 5 | Southeast | 1.09 | ||
| 6 | South | 1.03 | ||
| 7 | Southwest | 1.21 | ||
| 8 | West | 1.17 | ||
| 9 | Northwest | 1.06 | ||
| 5 | Elevation (m) | 1 | Very low (<350 m) | 0.14 |
| 2 | Low (350–400 m) | 1.85 | ||
| 3 | Moderate (400>) | 3.37 | ||
| 6 | Soil type | 1 | Weakly developed shallow soil | 0.00 |
| 2 | Weakly developed shallow & Lithosol | 0.30 | ||
| 3 | Lithosol | 0.00 | ||
| 4 | Alluvial | 0.00 | ||
| 5 | Fersiallitic | 1.85 | ||
| 6 | Smectitic clay & weakly developed shallow | 0.88 | ||
| 7 | Fersiallitic & Lithosols | 0.00 | ||
| 7 | Lithology | 1 | Alluvium | 0.00 |
| 2 | Basalt | 0.00 | ||
| 3 | Gabbro | 0.91 | ||
| 4 | Granite/Gneiss | 1.49 | ||
| 5 | Sandstone | 0.00 | ||
| 6 | Pyroxenite/Carbonite | 0.00 | ||
| 7 | Greenstone belt | 0.21 | ||
| 8 | Slope (%) | 1 | 0–2 | 1.01 |
| 2 | 2–6 | 1.05 | ||
| 3 | 6–12 | 1.01 | ||
| 4 | 12–20 | 0.27 | ||
| 5 | 20–30 | 0.00 | ||
| 6 | 30–45 | 0.00 | ||
| 7 | 45+ | 0.00 | ||
| 9 | Topographic relief (m) | 1 | 0–2 | 0.97 |
| 2 | 2–5 | 1.12 | ||
| 3 | 5–9 | 0.64 | ||
| 4 | 9–16 | 0.15 | ||
| 5 | 16–47 | 0.00 | ||
| 10 | Land-cover type | 1 | Barren land | 0.00 |
| 2 | Settlements | 0.00 | ||
| 3 | Forested land | 1.02 | ||
| 4 | Grassland | 0.90 | ||
| 5 | Mines & Quarries | 0.00 | ||
| 6 | Scrublands | 0.00 | ||
| 7 | Water bodies | 0.00 | ||
| 8 | Wetlands | 0.00 | ||
| 11 | Wind speed (m/s) | 1 | 1.0–1.6 | 0.00 |
| 2 | 1.6–2.3 | 0.74 | ||
| 3 | 2.3–3.0 | 3.68 | ||
| 12 | Precipitation (mm) | 1 | 438–470.7 | 0.00 |
| 2 | 470.7–503.5 | 0.09 | ||
| 3 | 503.5–536.2 | 0.60 | ||
| 4 | 536.2–569 | 3.52 | ||
| 13 | Slope Position | 1 | Concave (<−0.1) | 0.90 |
| 2 | Flat (−0.1–+0.1) | 1.01 | ||
| 3 | Convex (>+0.1) | 1.06 | ||
| 14 | Temperature (°C) | 1 | 21.9–22.3 | 2.65 |
| 2 | 22.3–22.6 | 0.58 | ||
| 3 | 22.6–23.0 | 0.00 | ||
| 4 | 23.0–23.3 | 0.00 |
2.3.3. The Analytic Hierarchy Process (AHP)
3. Results
3.1. The Most Effective Factors in the Distribution of Termitaria
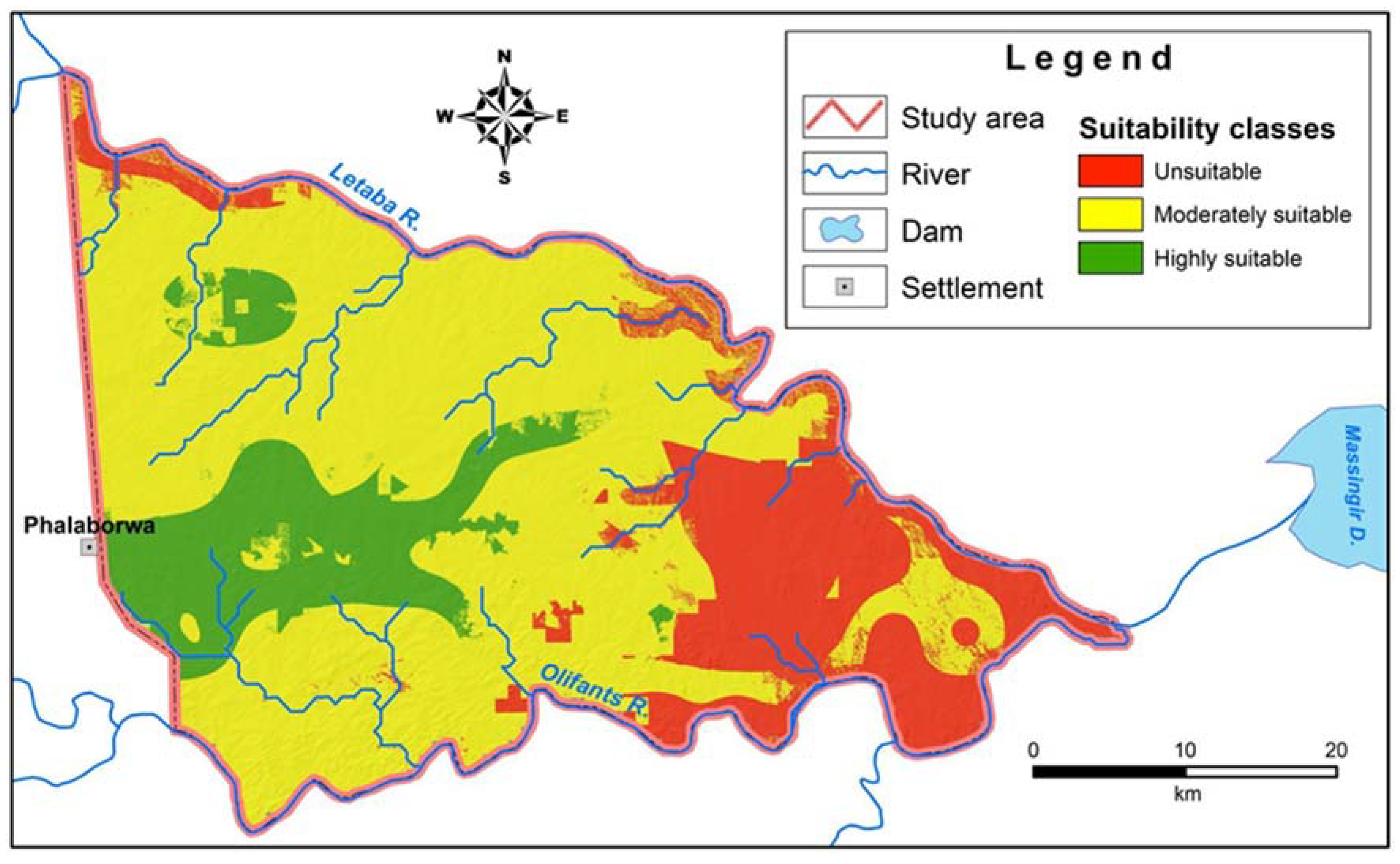
3.2. The Distribution of Land Suitability for Termitaria
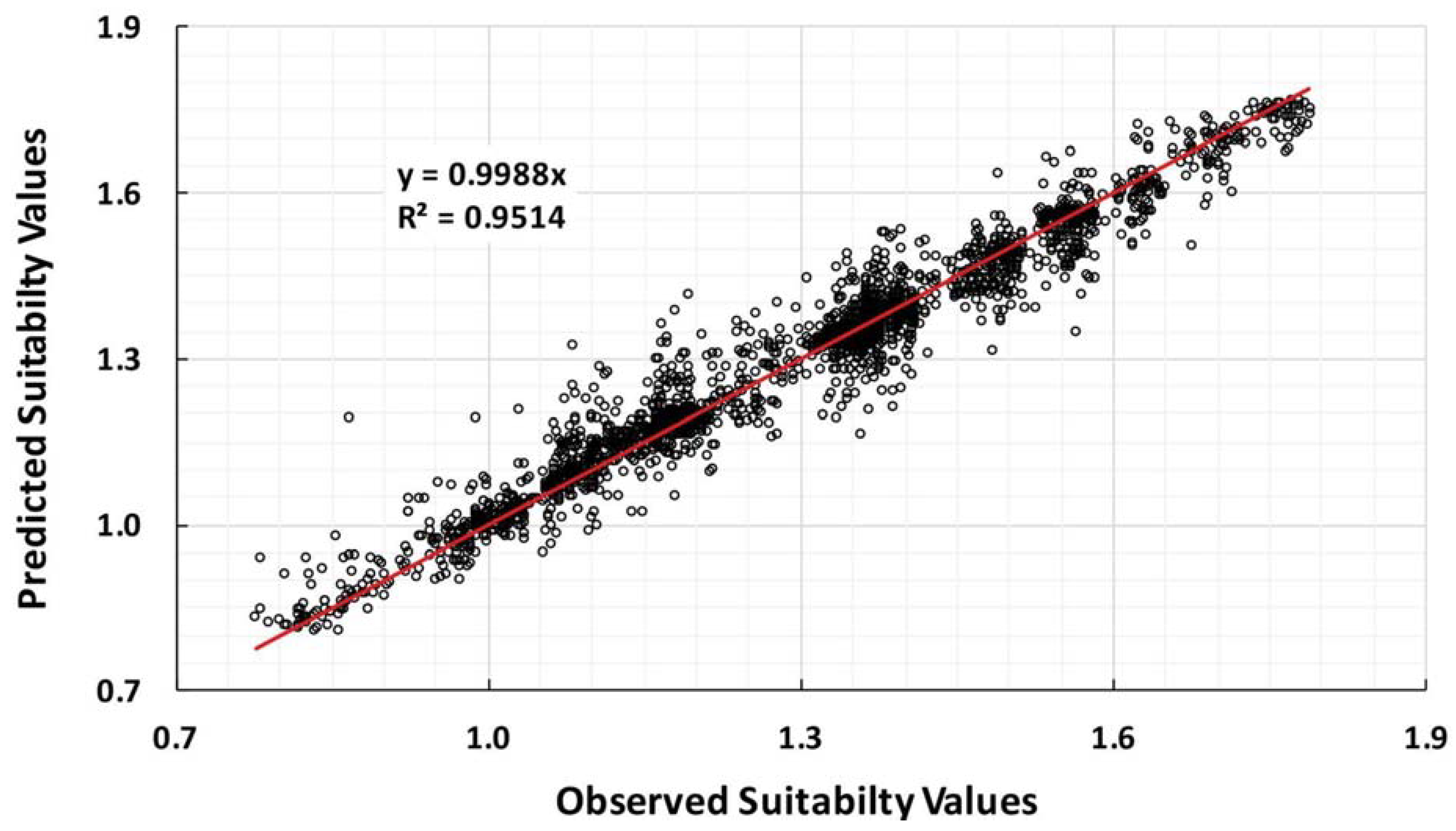
3.3. Validation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eggleton, P. Global Patterns of Termite Diversity. In Termites: Evolution, Sociality, Symbioses, Ecology; Springer Science & Business Media: Berlin, Germany, 2000. [Google Scholar]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.P. Soil Invertebrates and Ecosystem Services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Black, H.I.J.; Okwakol, M.J.N. Agricultural Intensification, Soil Biodiversity and Agroecosystem Function in the Tropics: The Role of Termites. Appl. Soil Ecol. 1997, 6, 37–53. [Google Scholar] [CrossRef]
- Jouquet, P.; Dauber, J.; Lagerlöf, J.; Lavelle, P.; Lepage, M. Soil Invertebrates as Ecosystem Engineers: Intended and Accidental Effects on Soil and Feedback Loops. Appl. Soil Ecol. 2006, 32, 153–164. [Google Scholar] [CrossRef]
- Bignell, D.E. Termites as Soil Engineers and Soil Processors. In Intestinal Microorganisms of Termites and Other Invertebrates; Springer: Berlin, Germany, 2005; pp. 183–220. [Google Scholar] [CrossRef]
- Jouquet, P.; Traoré, S.; Choosai, C.; Hartmann, C.; Bignell, D. Influence of Termites on Ecosystem Functioning. Ecosystem Services Provided by Termites. Eur. J. Soil Biol. 2011, 47, 215–222. [Google Scholar] [CrossRef]
- Ashton, L.A.; Griffiths, H.M.; Parr, C.L.; Evans, T.A.; Didham, R.K.; Hasan, F.; Teh, Y.A.; Tin, H.S.; Vairappan, C.S.; Eggleton, P. Termites Mitigate the Effects of Drought in Tropical Rainforest. Science 2019, 363, 174–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.T.; Susilo, F.X.; Bignell, D.E.; Hardiwinoto, S.; Gillison, A.N.; Eggleton, P. Termite Assemblage Collapse along a Land-Use Intensification Gradient in Lowland Central Sumatra, Indonesia. J. Appl. Ecol. 2003, 40, 380–391. [Google Scholar] [CrossRef] [Green Version]
- Albano Araújo, A.P.; Cristaldo, P.F.; Florencio, D.F.; Araújo, F.S.; DeSouza, O. Resource Suitability Modulating Spatial Co-occurrence of Soil-forager Termites (Blattodea: Termitoidea). Wiley Online Libr. 2016, 56, 235–243. [Google Scholar] [CrossRef]
- Jouquet, P.; Bottinelli, N.; Shanbhag, R.R.; Bourguignon, T.; Traoré, S.; Abbasi, S.A. Termites: The Neglected Soil Engineers of Tropical Soils. Soil Sci. 2016, 181, 157–165. [Google Scholar] [CrossRef]
- Shanbhag, R.R.; Kabbaj, M.; Sundararaj, R.; Jouquet, P. Rainfall and Soil Properties Influence Termite Mound Abundance and Height: A Case Study with Odontotermes obesus (Macrotermitinae) Mounds in the Indian Western Ghats Forests. Appl. Soil Ecol. 2017, 111, 33–38. [Google Scholar] [CrossRef]
- Jamilu Bala Ahmed, I.I.; Pradhan, B.; Mansor, S.; Tongjura, J.D.C.; Yusuf, B. Multi-Criteria Evaluation of Suitable Sites for Termite Mounds Construction in a Tropical Lowland. Catena 2019, 178, 359–371. [Google Scholar] [CrossRef]
- Lavelle, P.; Bignell, D.; Lepage, M.; Wolters, V.; Roger, P.A.; Ineson, P.O.W.H.; Heal, O.W.; Dhillion, S. Soil Function in a Changing World: The Role of Invertebrate Ecosystem Engineers. Eur. J. Soil Biol. 1997, 33, 159–193. [Google Scholar]
- Lavelle, P. Functional Domains in Soils. Ecol. Res. 2002, 17, 441–450. [Google Scholar] [CrossRef]
- Korb, J.; Linsenmair, K.E. Ventilation of Termite Mounds: New Results Require a New Model. Behav. Ecol. 2000, 11, 486–494. [Google Scholar] [CrossRef]
- Wang, X.D.; Zhong, X.H.; Liu, S.Z.; Liu, J.G.; Wang, Z.Y.; Li, M.H. Regional Assessment of Environmental Vulnerability in the Tibetan Plateau: Development and Application of a New Method. J. Arid. Environ. 2008, 72, 1929–1939. [Google Scholar] [CrossRef]
- Toro, J.; Duarte, O.; Requena, I.; Zamorano, M. Determining Vulnerability Importance in Environmental Impact Assessment. The Case of Colombia. Environ. Impact Assess. Rev. 2012, 32, 107–117. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, G.; Fang, L.; Nie, Y.; Rosen, M.A. GIS-Based Integration of Subjective and Objective Weighting Methods for Regional Landslides Susceptibility Mapping. Sustainability 2016, 8, 334. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Zhang, Q.; Ye, S.; Ren, B. A Novel Hybrid Approach for Landslide Susceptibility Mapping Integrating Analytical Hierarchy Process and Normalized Frequency Ratio Methods with the Cloud Model. Geomorphology 2019, 327, 170–187. [Google Scholar] [CrossRef]
- Tan, Y.; Shuai, C.; Jiao, L.; Shen, L. An Adaptive Neuro-Fuzzy Inference System (ANFIS) Approach for Measuring Country Sustainability Performance. Environ. Impact Assess. Rev. 2017, 65, 29–40. [Google Scholar] [CrossRef]
- Barro, A.S.; Fegan, M.; Moloney, B.; Porter, K.; Muller, J.; Warner, S.; Blackburn, J.K. Redefining the Australian anthrax belt: Modeling the ecological niche and predicting the geographic distribution of Bacillus anthracis. PLoS Negl. Trop. Dis. 2016, 10, e0004689. [Google Scholar] [CrossRef]
- Lissovsky, A.; Dudov, S. Species-Distribution Modeling: Advantages and Limitations of Its Application. 2. MaxEnt. Biol. Bull. Rev. 2021, 11, 265–275. [Google Scholar] [CrossRef]
- Akhter, S.; McDonald, M.A.; van Breugel, P.; Sohel, S.; Kjaer, E.D.; Mariott, R. Habitat distribution modelling to identify areas of high conservation value under climate change for Mangifera sylvatica Roxb. of Bangladesh. Land Use Policy 2017, 60, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Escobar, L.E.; Craft, M.E. Advances and limitations of disease biogeography using ecological niche modeling. Front. Microbiol. 2017, 7, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahabi, H.; Hashim, M.; Ahmad, B. bin Remote Sensing and GIS-Based Landslide Susceptibility Mapping Using Frequency Ratio, Logistic Regression, and Fuzzy Logic Methods at the Central Zab Basin, Iran. Environ. Earth Sci. 2015, 73, 8647–8668. [Google Scholar] [CrossRef]
- Levick, S.R.; Asner, G.P.; Kennedy-Bowdoin, T.; Knapp, D.E. The Spatial Extent of Termite Influences on Herbivore Browsing in an African Savanna. Biol. Conserv. 2010, 143, 2462–2467. [Google Scholar] [CrossRef]
- Ahmed II, J.B.; Pradhan, B. Spatial Assessment of Termites Interaction with Groundwater Potential Conditioning Parameters in Keffi, Nigeria. J. Hydrol. 2019, 578, 124012. [Google Scholar] [CrossRef]
- Ahmed, J.B.; Pradhan, B. Termite Mounds as Bio-Indicators of Groundwater: Prospects and Constraints. Pertanika J. Sci. Technol. 2018, 26, 479–498. [Google Scholar]
- Bala, J.; Ii, A.; Pradhan, B.; Mansor, S.; Yusoff, Z.M.; Ekpo, S.A. Aquifer Potential Assessment in Termites Manifested Locales Using Geo-Electrical and Surface Hydraulic Measurement Parameters. Sensors 2019, 19, 2107. [Google Scholar] [CrossRef] [Green Version]
- Chiri, E.; Greening, C.; Lappan, R.; Waite, D.W.; Jirapanjawat, T.; Dong, X.; Arndt, S.K.; Nauer, P.A. Termite Mounds Contain Soil-Derived Methanotroph Communities Kinetically Adapted to Elevated Methane Concentrations. ISME J. 2020, 14, 2715–2731. [Google Scholar] [CrossRef]
- Chen, Q.L.; Hu, H.W.; Yan, Z.Z.; Li, C.Y.; Nguyen, B.A.T.; Sun, A.Q.; Zhu, Y.G.; He, J.Z. Deterministic Selection Dominates Microbial Community Assembly in Termite Mounds. Soil Biol. Biochem. 2021, 152, 108073. [Google Scholar] [CrossRef]
- Sinclair, A.; Walker, B. The Kruger Experience: Ecology and Management of Savanna Heterogeneity; Island Press: Washington, DC, USA, 2003. [Google Scholar]
- Coaton, W.G.H. Survey of the Termites of the Kruger National Park. Koedoe 1962, 5, 144–156. [Google Scholar] [CrossRef]
- Davies, A.B.; Eggleton, P.; van Rensburg, B.J.; Parr, C.L. The Pyrodiversity-Biodiversity Hypothesis: A Test with Savanna Termite Assemblages. J. Appl. Ecol. 2012, 49, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.B.; Lind, B.M. A Checklist of the Termites of Kruger National Park, South Africa. Koedoe Afr. Prot. Area Conserv. Sci. 2019, 61, a1531. [Google Scholar] [CrossRef] [Green Version]
- Meyer, V.W.; Braack, L.E.O.; Biggs, H.C.; Ebersohn, C. Distribution and Density of Termite Mounds in the Northern Kruger National Park, with Specific Reference to Those Constructed by Macrotermes Holmgren (Isoptera: Termitidae). Afr. Entomol. 1999, 7, 123–130. [Google Scholar]
- Braack, L.E.O. Seasonal Activity of Savanna Termites during and after Severe Drought. Koedoe 1995, 38, 73–82. [Google Scholar] [CrossRef]
- Meyer, V.W.; Braack, L.E.O.; Biggs, H.C. Distribution and Density of Cubitermes Wasmann (Isoptera: Termitidae) Mounds in the Northern Kruger National Park. Koedoe 2000, 43, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Meyer, V.W.; Crewe, R.M.; Braack, L.E.O. Estimates of Food Consumption by the Fungus-Growing Termite Macrotermes Natalensis in a South African Savanna-Woodland. S. Afr. J. Sci. 2003, 99, 207–208. [Google Scholar] [CrossRef]
- Meyer, V.W.; Crewe, R.M.; Braack, L.E.O.; Groeneveld, H.T.; van der Linde, M.J. Intracolonial Demography of the Mound-Building Termite Macrotermes Natalensis (Haviland) (Isoptera, Termitidae) in the Northern Kruger National Park, South Africa. Insectes Soc. 2000, 47, 390–397. [Google Scholar] [CrossRef]
- Meyer, V.W.; Crewe, R.M.; Braack, L.E.O.; Groeneveld, H.T.; van der Linde, M.J. Biomass of Macrotermes Natalensis in the Northern Kruger National Park, South Africa- the Effects of Land Characteristics. Sociobiology 2001, 38, 431–438. [Google Scholar]
- Sponheimer, M.; Lee-Thorp, J.; de Ruiter, D.; Codron, D.; Codron, J.; Baugh, A.T.; Thackeray, F. Hominins, Sedges, and Termites: New Carbon Isotope Data from the Sterkfontein Valley and Kruger National Park. J. Hum. Evol. 2005, 48, 301–312. [Google Scholar] [CrossRef]
- Grant, C.C.; Scholes, M.C. The Importance of Nutrient Hot-Spots in the Conservation and Management of Large Wild Mammalian Herbivores in Semi-Arid Savannas. Biol. Conserv. 2006, 130, 426–437. [Google Scholar] [CrossRef]
- Levick, S.R.; Asner, G.P.; Chadwick, O.A.; Khomo, L.M.; Rogers, K.H.; Hartshorn, A.S.; Kennedy-Bowdoin, T.; Knapp, D.E. Regional Insight into Savanna Hydrogeomorphology from Termite Mounds. Nat. Commun. 2010, 1, 65. [Google Scholar] [CrossRef] [PubMed]
- Coetsee, C.; Jacobs, S.; Govender, N. An Overview of Nitrogen Cycling in a Semiarid Savanna: Some Implications for Management and Conservation in a Large African Park. Environ. Manag. 2012, 49, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.B.; Parr, C.L.; Eggleton, P. A Global Review of Termite Sampling Methods. Insectes Soc. 2021, 68, 3–14. [Google Scholar] [CrossRef]
- Davies, A.B.; Baldeck, C.A.; Asner, G.P. Termite Mounds Alter the Spatial Distribution of African Savanna Tree Species. J. Biogeogr. 2016, 43, 301–313. [Google Scholar] [CrossRef]
- Davies, A.B.; Brodrick, P.G.; Parr, C.L.; Asner, G.P. Resistance of Mound-Building Termites to Anthropogenic Land-Use Change. Environ. Res. Lett. 2020, 15, 094038. [Google Scholar] [CrossRef]
- Davies, A.B.; Eggleton, P.; van Rensburg, B.J.; Parr, C.L. Seasonal Activity Patterns of African Savanna Termites Vary across a Rainfall Gradient. Insectes Soc. 2015, 62, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.B.; Levick, S.R.; Asner, G.P.; Robertson, M.P.; van Rensburg, B.J.; Parr, C.L.; Davies, A.B.; Robertson, M.P.; van Rensburg, B.J. Spatial Variability and Abiotic Determinants of Termite Mounds throughout a Savanna Catchment. Wiley Online Libr. 2014, 37, 852–862. [Google Scholar] [CrossRef]
- Davies, A.B.; Levick, S.R.; Robertson, M.P.; van Rensburg, B.J.; Asner, G.P.; Parr, C.L. Termite Mounds Differ in Their Importance for Herbivores across Savanna Types, Seasons and Spatial Scales. Wiley Online Libr. 2015, 125, 726–734. [Google Scholar] [CrossRef]
- Joseph, G.S.; Seymour, C.L.; Coetzee, B.W.T.; Ndlovu, M.; De, A.; Torre, L.; Suttle, R.; Hicks, N.; Oxley, S.; Foord, S.H. Microclimates Mitigate against Hot Temperatures in Dryland Ecosystems: Termite Mounds as an Example. Wiley Online Libr. 2016, 7, e01509. [Google Scholar] [CrossRef]
- Leitner, M.; Davies, A.B.; Robertson, M.P.; Parr, C.L.; van Rensburg, B.J. Termite Mounds Create Heterogeneity in Invertebrate Communities across a Savanna Rainfall Gradient. Biodivers. Conserv. 2020, 29, 1427–1441. [Google Scholar] [CrossRef]
- Viljoen, M. The Kruger National Park: Geology and Geomorphology of the Wilderness. World Geomorphol. Landsc. 2015, 111–120. [Google Scholar] [CrossRef]
- Venter, F.J.; Scholes, R.J.; Eckhardt, H.C. The Abiotic Template and Its Associated Vegetation Pattern. Kruger Exp. Ecol. Manag. Savanna Heterog. 2003, 83, 129. [Google Scholar]
- Phillips, G.; Mketeni, F.G.; Yawitch, J. Kruger National Park Management Plan. Available online: https://www.sanparks.org/assets/docs/conservation/park_man/knp/draft-plan.pdf (accessed on 26 April 2022).
- Pribadi, T.; Raffiudin, R.; Harahap, I.S. Termites Community as Environmental Bioindicators in Highlands: A Case Study in Eastern Slopes of Mount Slamet, Central Java. Biodivers. J. Biol. Divers. 2011, 12, 235–240. [Google Scholar] [CrossRef]
- Agarwal, V.B. Temperature and Relative Humidity inside the Mound of Odontotermes obesus (Rambur) (Isoptera: Termitidae). Proc. Anim. Sci. 1980, 89, 91–99. [Google Scholar] [CrossRef]
- Dibog, L.; Eggleton, P.; Norgrove, L.; Bignell, D.E.; Hauser, S. Impacts of Canopy Cover on Soil Termite Assemblages in an Agrisilvicultural System in Southern Cameroon. Bull. Entomol. Res. 1999, 89, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Fagundes, T.M.; Ordonez, J.C.; Yaghoobian, N. How the Thermal Environment Shapes the Structure of Termite Mounds. R. Soc. Open Sci. 2020, 7, 191332. [Google Scholar] [CrossRef] [Green Version]
- D’hont, B.; Calders, K.; Bartholomeus, H.; Whiteside, T.; Bartolo, R.; Levick, S.; Moorthy, S.M.K.; Terryn, L.; Verbeeck, H.; Wallace, L. Characterising Termite Mounds in a Tropical Savanna with UAV Laser Scanning. Remote Sens. 2021, 13, 476. [Google Scholar] [CrossRef]
- Grohmann, C.; Oldeland, J.; Stoyan, D.; Linsenmair, K.E. Multi-Scale Pattern Analysis of a Mound-Building Termite Species. Insectes Soc. 2010, 57, 477–486. [Google Scholar] [CrossRef]
- Florencio, D.; Diehl, E. Termitofauna (Insecta, Isoptera) Em Remanescentes de Floresta Estacional Semidecidual Em São Leopoldo, Rio Grande Do Sul, Brasil. SciELO Bras. 2006, 50, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Lima, S.S.; Ceddia, M.B.; Zuchello, F.; de Aquino, A.M.; Mercante, F.M.; Alves, B.J.R.; Urquiaga, S.; Martius, C.; Boddey, R.M. Variabilidade Espacial e Vitalidade de Ninhos Epígeos Detérmitas Em Pastagens de Mato Grosso Do Sul, Brasil. Rev. Bras. De Cienc. Do Solo 2015, 39, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Korb, J.; Linsenmair, K.E. The Architecture of Termite Mounds: A Result of a Trade-off between Thermoregulation and Gas Exchange? Behav. Ecol. 1999, 10, 312–316. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Maiti, R. Application of Analytical Hierarchy Process (AHP) and Frequency Ratio (FR) Model in Assessing Landslide Susceptibility and Risk. In Semi-Quantitative Approaches for Landslide Assessment and Prediction; Springer: Singapore, 2015; pp. 191–226. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Wiley Online Libr. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- MacDonald, A.M.; Bonsor, H.C.; Dochartaigh, B.É.Ó.; Taylor, R.G. Quantitative Maps of Groundwater Resources in Africa. Environ. Res. Lett. 2012, 7, 24009–24016. [Google Scholar] [CrossRef]
- Andreadis, K.M.; Schumann, G.J.P.; Pavelsky, T. A Simple Global River Bankfull Width and Depth Database. Water Resour. Res. 2013, 49, 7164–7168. [Google Scholar] [CrossRef]
- ASF DAAC. ALOS PALSAR Radiometric Terrain Corrected Low_Resolution. Available online: https://asf.alaska.edu (accessed on 26 April 2022).
- SA National Land-Cover Datasets|Environmental Geographical Information Systems (E-GIS). Available online: https://egis.environment.gov.za/sa_national_land_cover_datasets (accessed on 26 April 2022).
- Saaty, T.L.; Vargas, L.G.; Dellmann, K. The Allocation of Intangible Resources: The Analytic Hierarchy Process and Linear Programming. Socio-Econ. Plan. Sci. 2003, 37, 169–184. [Google Scholar] [CrossRef]
- Amin Salehi, A.; Navabian, M.; Varaki, M.E.; Pirmoradian, N. Evaluation of HYDRUS-2D Model to Simulate the Loss of Nitrate in Subsurface Controlled Drainage in a Physical Model Scale of Paddy Fields. Paddy Water Environ. 2017, 15, 433–442. [Google Scholar] [CrossRef]
- Freymann, B.P.; de Visser, S.N.; Olff, H.; Freymann, B.P.; Olff, H. Spatial and Temporal Hotspots of Termite-driven Decomposition in the Serengeti. Wiley Online Libr. 2010, 33, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Mège, D.; Rango, T. Permanent Groundwater Storage in Basaltic Dyke Fractures and Termite Mound Viability. J. Afr. Earth Sci. 2010, 57, 127–142. [Google Scholar] [CrossRef]
- Ocko, S.A.; King, H.; Andreen, D.; Bardunias, P.; Turner, J.S.; Soar, R.; Mahadevan, L. Solar-Powered Ventilation of African Termite Mounds. J. Exp. Biol. 2017, 220, 3260–3269. [Google Scholar] [CrossRef] [Green Version]
- Vesala, R.; Harjuntausta, A.; Hakkarainen, A.; Rönnholm, P.; Pellikka, P.; Rikkinen, J. Termite Mound Architecture Regulates Nest Temperature and Correlates with Species Identities of Symbiotic Fungi. PeerJ 2019, 2019, e6237. [Google Scholar] [CrossRef]
- Crist, T.O. The Spatial Distribution of Termites in Shortgrass Steppe: A Geostatistical Approach. Oecologia 1998, 114, 410–416. [Google Scholar] [CrossRef] [PubMed]
- King, H.; Ocko, S.; Mahadevan, L. Termite Mounds Harness Diurnal Temperature Oscillations for Ventilation. Proc. Natl. Acad. Sci. USA 2015, 112, 11589–11593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Schijff, H.P. Die Invloed van Termitaria Op Die Plantegroei van Die Nasionale Krugerwildtuin. Available online: weblink (accessed on 26 April 2022).
- Ahmed, B.M.; Nkunika, P.O.Y.; Sileshi, G.W.; French, J.R.J.; Nyedo, P.; Jain, S. Potential Impact of Climate Change on Termite Distribution in Africa. Br. J. Environ. Clim. Chang. 2011, 1, 172–189. [Google Scholar]
- Herrero, H.V.; Southworth, J. Special Issue on Dynamics of the Global Savanna and Grasslands Biomes. Appl. Sci. 2020, 10, 43. [Google Scholar] [CrossRef]
- Herrero, H.; Waylen, P.; Southworth, J.; Khatami, R.; Yang, D.; Child, B. A Healthy Park Needs Healthy Vegetation: The Story of Gorongosa National Park in the 21st Century. Remote Sens. 2020, 12, 476. [Google Scholar] [CrossRef] [Green Version]






| Suitability Class | Index Value | Area (ha) | Ratio (%) |
|---|---|---|---|
| Highly suitable | 1.27–1.78 | 25,078.11 | 14.26 |
| Moderately suitable | 0.76–1.27 | 114,205.06 | 64.93 |
| Unsuitable | 0.25–0.76 | 36,609.22 | 20.81 |
| TOTAL | 175,892.00 | 100 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozsahin, E.; Ozdes, M.; Smith, A.C.; Yang, D. Remote Sensing and GIS-Based Suitability Mapping of Termite Habitat in the African Savanna: A Case Study of the Lowveld in Kruger National Park. Land 2022, 11, 803. https://doi.org/10.3390/land11060803
Ozsahin E, Ozdes M, Smith AC, Yang D. Remote Sensing and GIS-Based Suitability Mapping of Termite Habitat in the African Savanna: A Case Study of the Lowveld in Kruger National Park. Land. 2022; 11(6):803. https://doi.org/10.3390/land11060803
Chicago/Turabian StyleOzsahin, Emre, Mehmet Ozdes, Audrey C. Smith, and Di Yang. 2022. "Remote Sensing and GIS-Based Suitability Mapping of Termite Habitat in the African Savanna: A Case Study of the Lowveld in Kruger National Park" Land 11, no. 6: 803. https://doi.org/10.3390/land11060803