
Discovering Tuberosin and Villosol as Potent and Selective Inhibitors of AKT1 for Therapeutic Targeting of Oral Squamous Cell Carcinoma

 ,
, 

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virtual Screening
2.2. PAINS Patterns and ADMET Properties of Compounds
2.3. Biological Activity Prediction through PASS
2.4. Interaction Analysis
2.5. MD Simulations
3. Results and Discussion
3.1. Virtual Screening
3.2. ADMET Properties and PAINS of the Compounds
3.3. Screening for Biological Activity Prediction: PASS Analysis
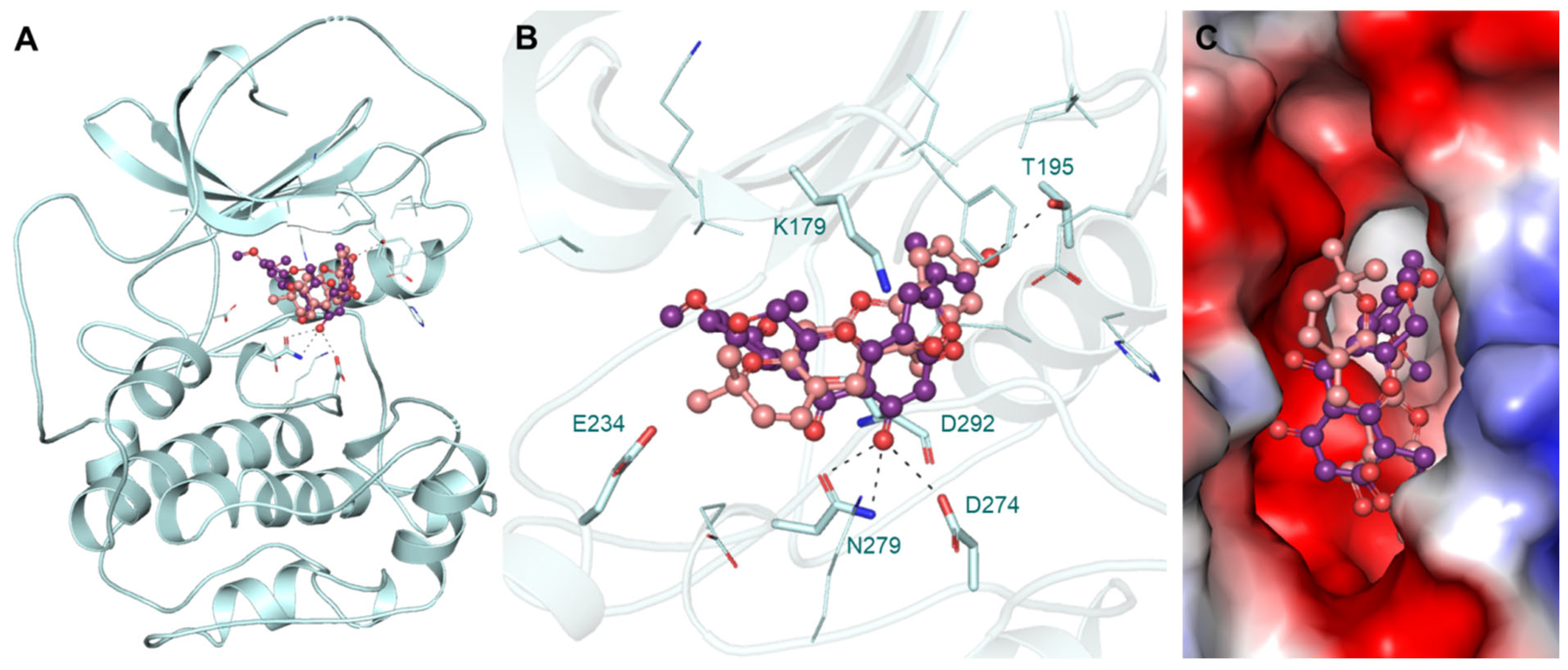
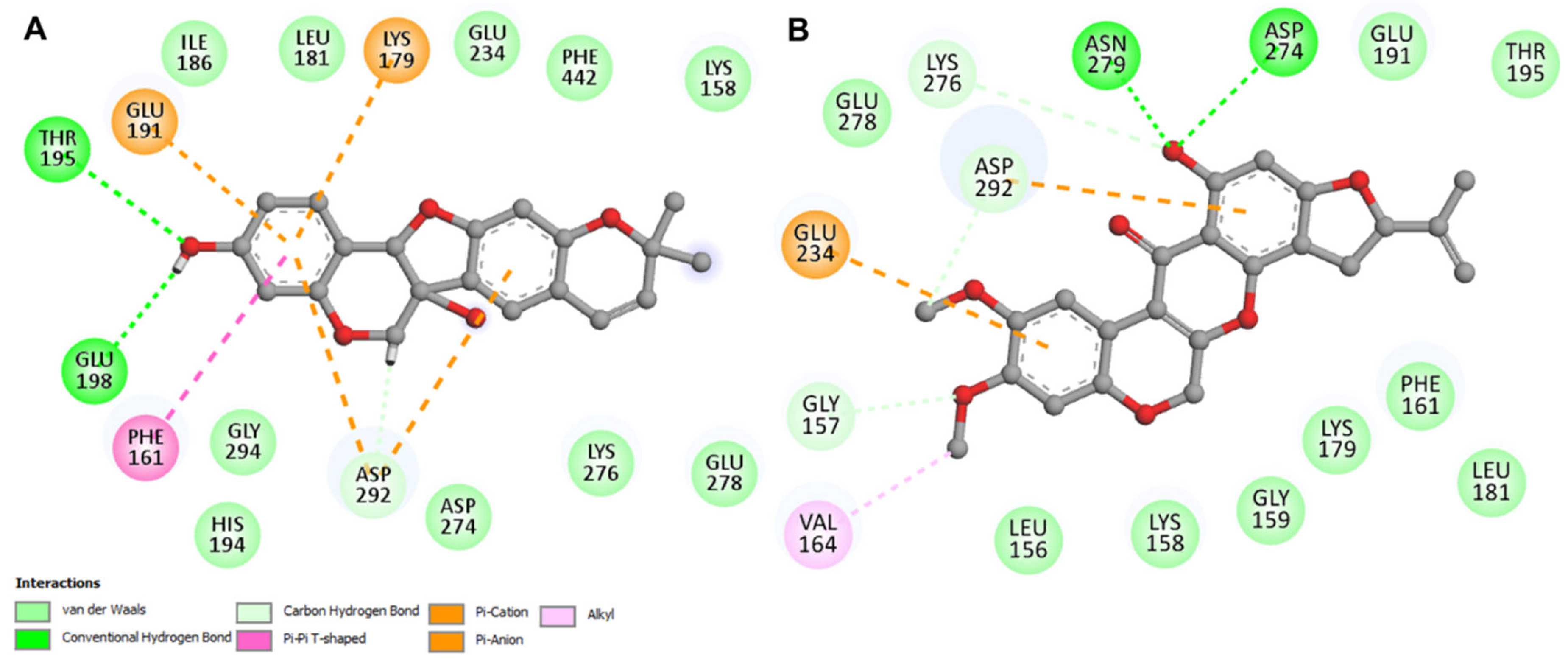
3.4. Interaction Analysis
3.5. MD Simulations
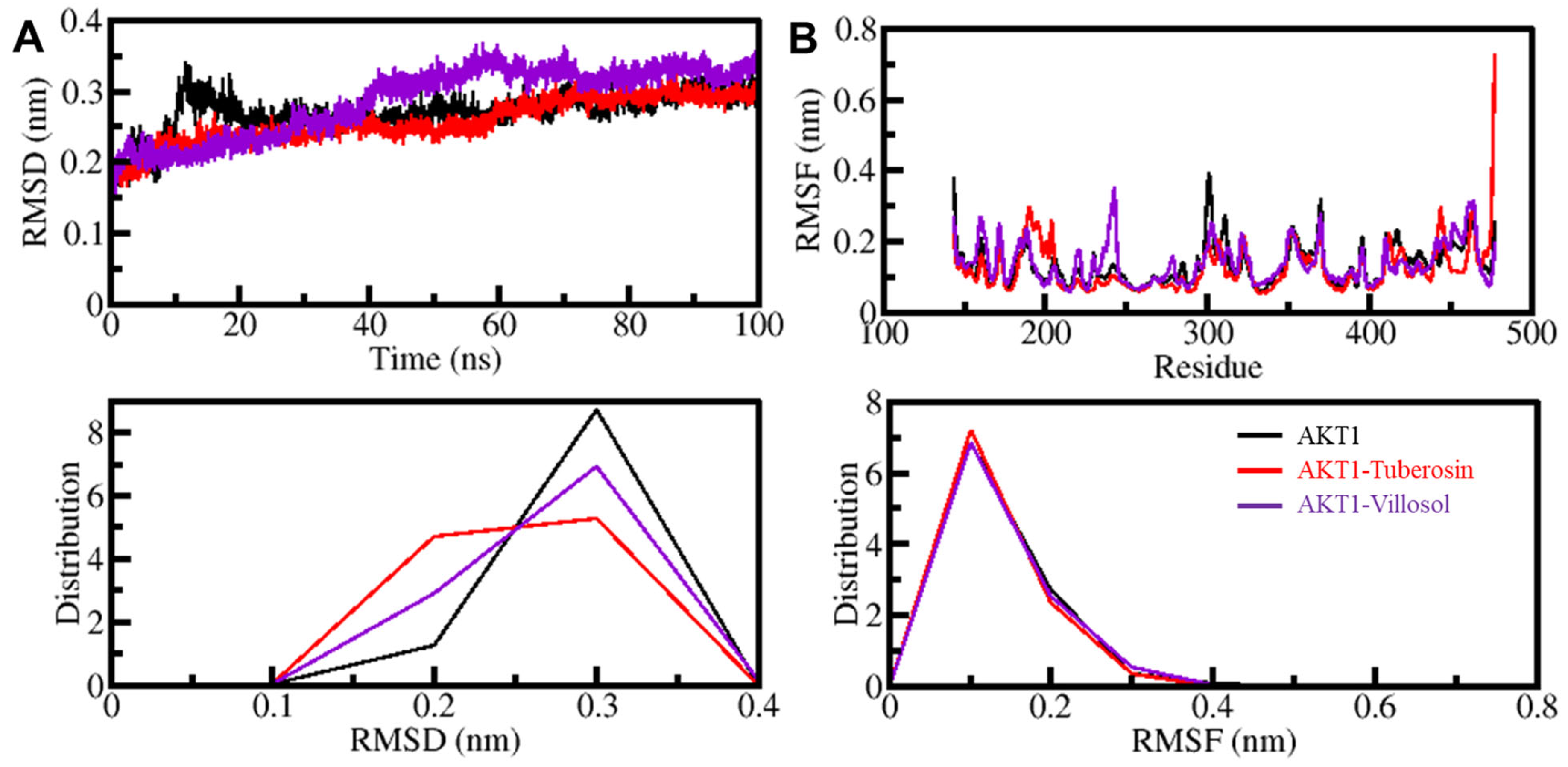
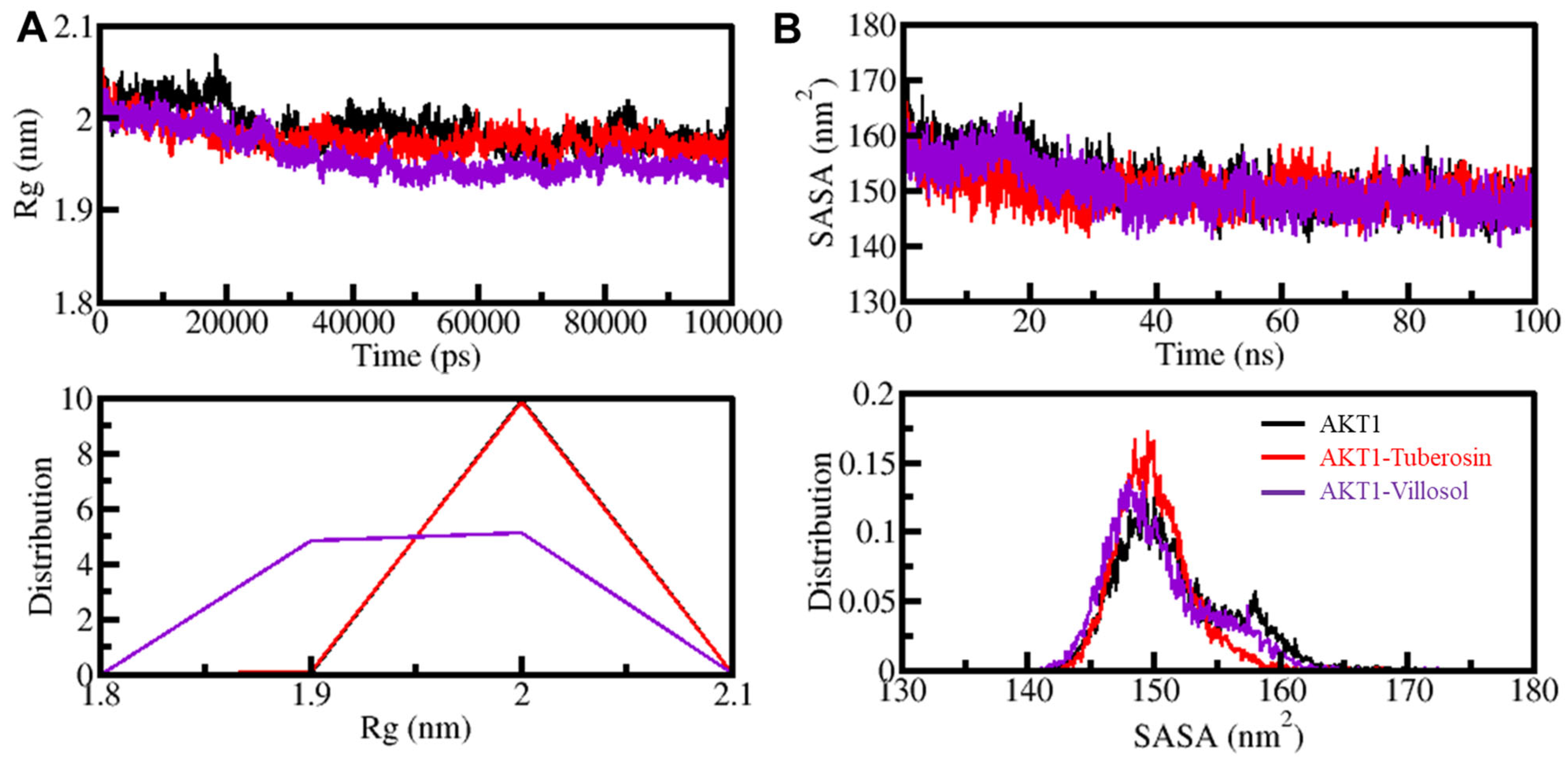
3.5.1. AKT1 Structural Changes through Ligand Binding
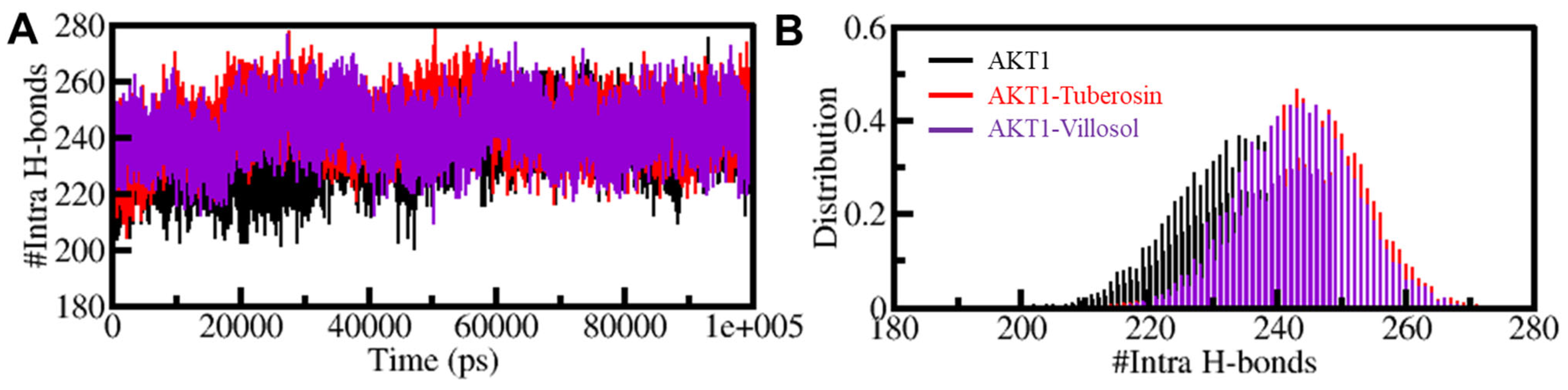
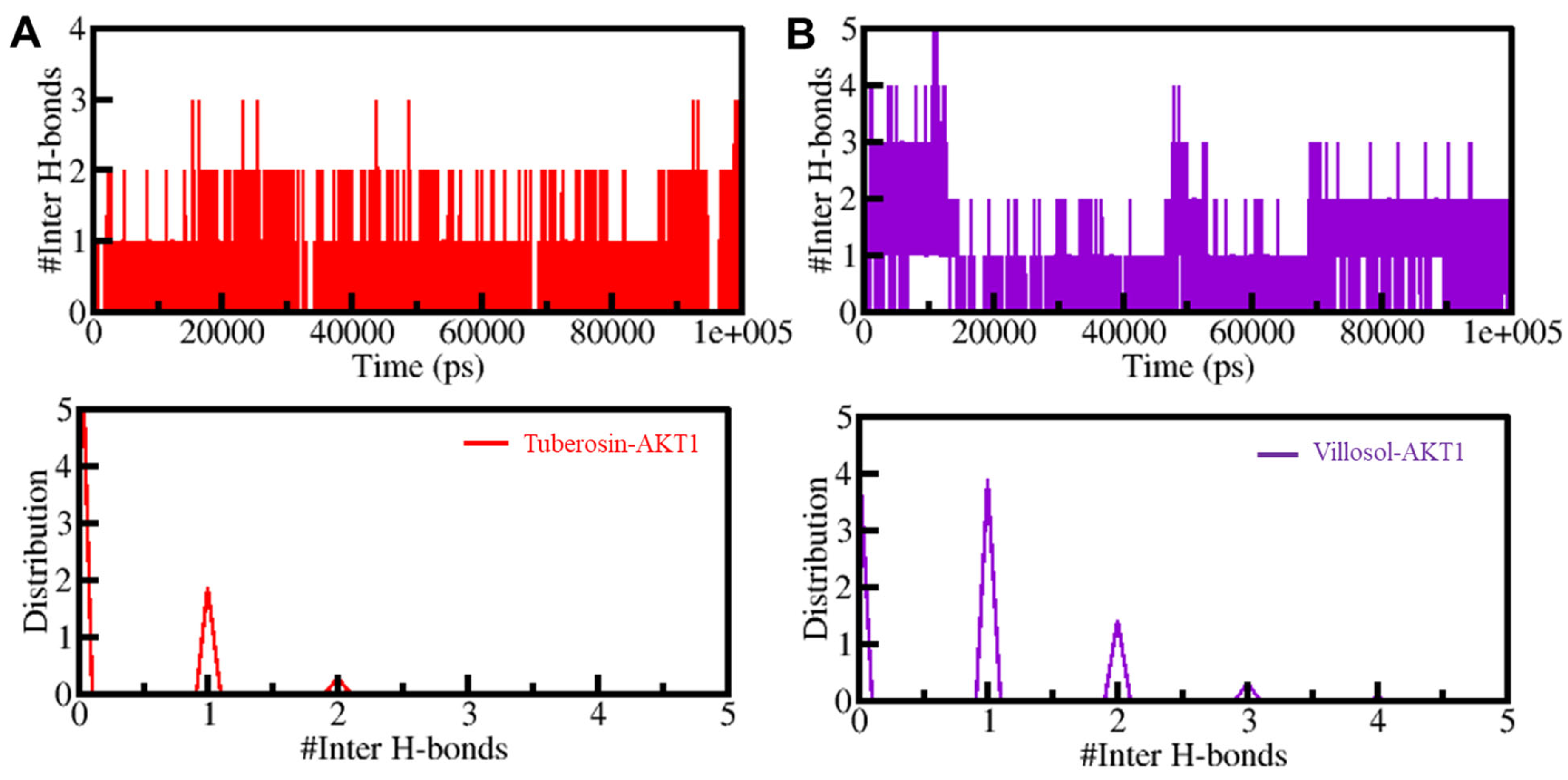
3.5.2. Dynamics of Hydrogen Bonds
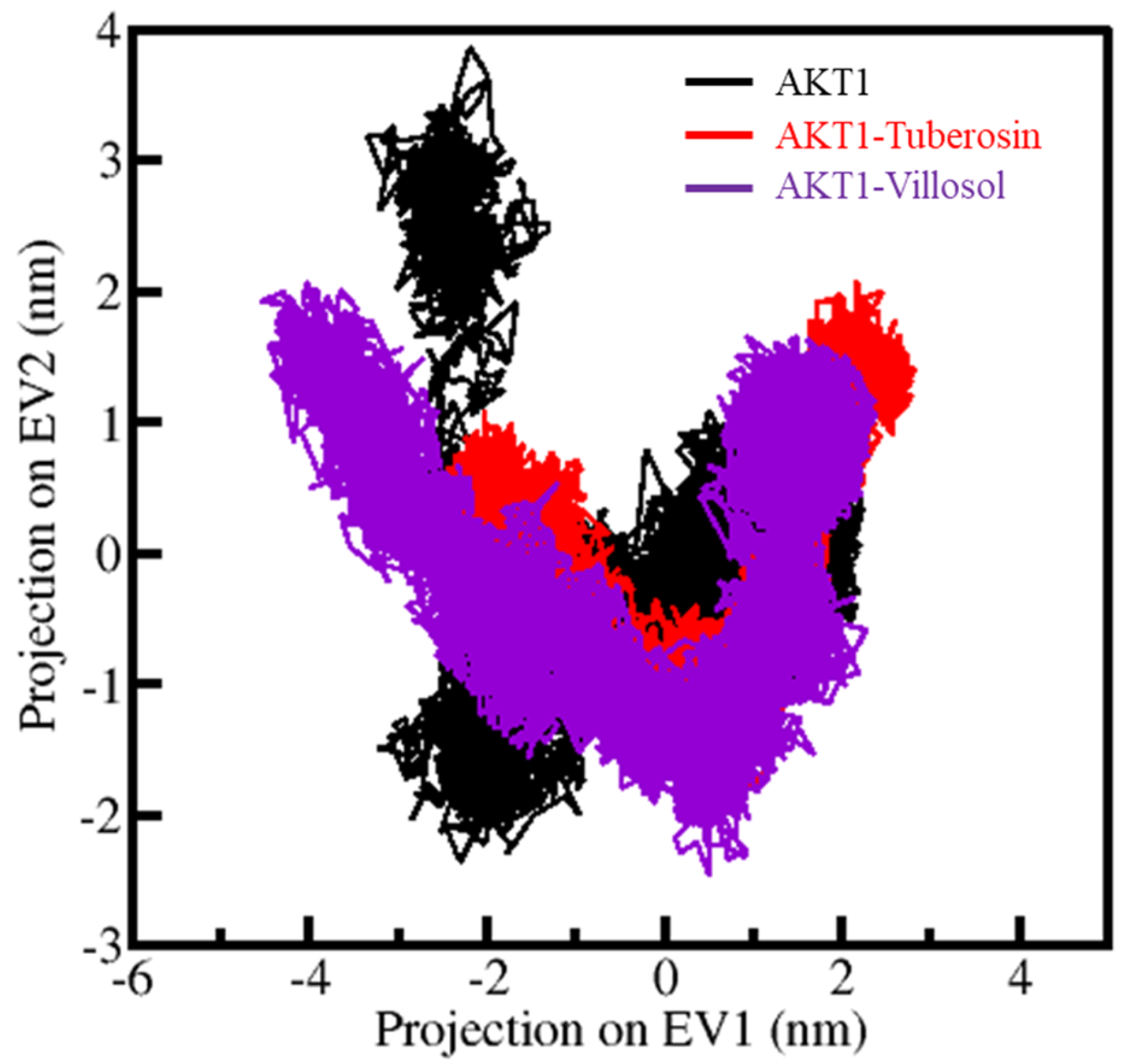
3.6. Principal Components Analysis
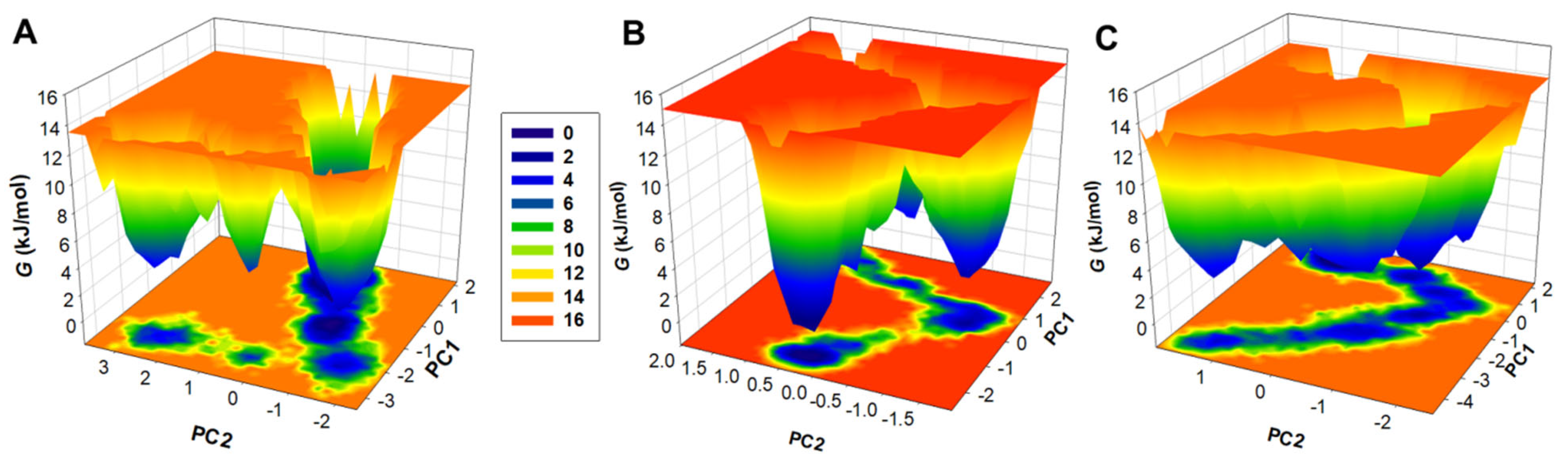
3.7. Free Energy Landscapes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giat, E.; Ehrenfeld, M.; Shoenfeld, Y. Cancer and autoimmune diseases. Autoimmun. Rev. 2017, 16, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Lawen, A. Apoptosis—An introduction. Bioessays 2003, 25, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Alwhaibi, A.; Verma, A.; Adil, M.S.; Somanath, P.R. The unconventional role of Akt1 in the advanced cancers and in diabetes-promoted carcinogenesis. Pharmacol. Res. 2019, 145, 104270. [Google Scholar] [CrossRef]
- Alwhaibi, A.; Verma, A.; Artham, S.; Adil, M.S.; Somanath, P.R. Nodal pathway activation due to Akt1 suppression is a molecular switch for prostate cancer cell epithelial-to-mesenchymal transition and metastasis. Biochem. Pharmacol. 2019, 168, 1–13. [Google Scholar] [CrossRef]
- Mishra, A.P.; Salehi, B.; Sharifi-Rad, M.; Pezzani, R.; Kobarfard, F.; Sharifi-Rad, J.; Nigam, M. Programmed cell death, from a cancer perspective: An overview. Mol. Diagn. Ther. 2018, 22, 281–295. [Google Scholar] [CrossRef]
- Cozzone, D.; Fröjdö, S.; Disse, E.; Debard, C.; Laville, M.; Pirola, L.; Vidal, H. Isoform-specific defects of insulin stimulation of Akt/protein kinase B (PKB) in skeletal muscle cells from type 2 diabetic patients. Diabetologia 2008, 51, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Somanath, P.R.; Razorenova, O.V.; Chen, J.; Byzova, T.V. Akt1 in endothelial cell and angiogenesis. Cell Cycle 2006, 5, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, F.; Lavigne, C.; Jacques, H.; Marette, A. Defective insulin-induced GLUT4 translocation in skeletal muscle of high fat–fed rats is associated with alterations in both Akt/protein kinase B and atypical protein kinase C (ζ/λ) activities. Diabetes 2001, 50, 1901–1910. [Google Scholar] [CrossRef] [Green Version]
- Nakashiro, K.-I.; Tanaka, H.; Goda, H.; Iwamoto, K.; Tokuzen, N.; Hara, S.; Onodera, J.; Fujimoto, I.; Hino, S.; Hamakawa, H. Identification of Akt1 as a potent therapeutic target for oral squamous cell carcinoma. Int. J. Oncol. 2015, 47, 1273–1281. [Google Scholar] [CrossRef] [Green Version]
- Sharif Siam, M.K.; Sarker, A.; Sayeem, M.M.S. In silico drug design and molecular docking studies targeting Akt1 (RAC-alpha serine/threonine-protein kinase) and Akt2 (RAC-beta serine/threonine-protein kinase) proteins and investigation of CYP (cytochrome P450) inhibitors against MAOB (monoamine oxidase B) for OSCC (oral squamous cell carcinoma) treatment. J. Biomol. Struct. Dyn. 2021, 39, 6467–6479. [Google Scholar] [PubMed]
- Wang, Y.W.; Lin, K.T.; Chen, S.C.; Gu, D.L.; Chen, C.F.; Tu, P.H.; Jou, Y.S. Overexpressed-eIF3I interacted and activated oncogenic Akt1 is a theranostic target in human hepatocellular carcinoma. Hepatology 2013, 58, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Priolo, C.; Pyne, S.; Rose, J.; Regan, E.R.; Zadra, G.; Photopoulos, C.; Cacciatore, S.; Schultz, D.; Scaglia, N.; McDunn, J. AKT1 and MYC induce distinctive metabolic fingerprints in human prostate cancer. Cancer Res. 2014, 74, 7198–7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Wang, G.; Paciga, J.E.; Feldman, R.I.; Yuan, Z.-Q.; Ma, X.-L.; Shelley, S.A.; Jove, R.; Tsichlis, P.N.; Nicosia, S.V. AKT1/PKBα kinase is frequently elevated in human cancers and its constitutive activation is required for oncogenic transformation in NIH3T3 cells. Am. J. Pathol. 2001, 159, 431–437. [Google Scholar] [CrossRef]
- Paoli, P.; Giannoni, E.; Chiarugi, P. Anoikis molecular pathways and its role in cancer progression. Biochim. Biophys. Acta-Mol. Cell Res. 2013, 1833, 3481–3498. [Google Scholar] [CrossRef] [Green Version]
- Alzahrani, A.S. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef]
- Oudit, G.Y.; Sun, H.; Kerfant, B.-G.; Crackower, M.A.; Penninger, J.M.; Backx, P.H. The role of phosphoinositide-3 kinase and PTEN in cardiovascular physiology and disease. J. Mol. Cell. Cardiol. 2004, 37, 449–471. [Google Scholar] [CrossRef]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [Green Version]
- Karar, J.; Maity, A. PI3K/AKT/mTOR pathway in angiogenesis. Front. Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.H.; Liu, L.Z. PI3K/PTEN signaling in angiogenesis and tumorigenesis. Adv. Cancer Res. 2009, 102, 19–65. [Google Scholar] [CrossRef]
- Mitsiades, C.S.; Mitsiades, N.; Koutsilieris, M. The Akt pathway: Molecular targets for anti-cancer drug development. Curr. Cancer Drug Targets 2004, 4, 235–256. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.Q.; Lindsley, C.W.; Cheng, G.Z.; Yang, H.; Nicosia, S.V. The Akt/PKB pathway: Molecular target for cancer drug discovery. Oncogene 2005, 24, 7482–7492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free. Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, H.R.; Jo, Y.H.; Kim, J.; Shin, Y.; Kim, S.S.; Choi, T.G. Roles of autophagy in oxidative stress. Int. J. Mol. Sci. 2020, 21, 3289. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Dutt, R.; Garg, V.; Khatri, N.; Madan, A.K. Phytochemicals in anticancer drug development. Anti-Cancer Agents Med. Chem. 2019, 19, 172–183. [Google Scholar] [CrossRef]
- Anjum, F.; Mohammad, T.; Almalki, A.A.; Akhtar, O.; Abdullaev, B.; Hassan, M.I. Phytoconstituents and Medicinal plants for anticancer drug discovery: Computational identification of potent inhibitors of PIM1 kinase. OMICS J. Integr. Biol. 2021, 25, 580–590. [Google Scholar] [CrossRef]
- Ashraf, M.A. Phytochemicals as potential anticancer drugs: Time to ponder nature’s bounty. BioMed Res. Int. 2020, 2020, 8602879. [Google Scholar] [CrossRef]
- Joshi, B.S.; Kamat, V.N. Tuberosin, a new pterocarpan from Pueraria tuberosa DC. J. Chem. Soc. Perkin Trans. 1973, 907–911. [Google Scholar] [CrossRef]
- Pandey, N.; Tripathi, Y.B. Antioxidant activity of tuberosin isolated from Pueraria tuberose Linn. J. Inflamm. 2010, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Dong, D.-D.; Huang, Q.-P.; Li, J.; Du, Y.-Y.; Li, B.; Li, H.-Q.; Huyan, T. The anti-inflammatory effect of Sonchus oleraceus aqueous extract on lipopolysaccharide stimulated RAW 264.7 cells and mice. Pharm. Biol. 2017, 55, 799–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Leister, W.H.; Robinson, R.G.; Barnett, S.F.; Defeo-Jones, D.; Jones, R.E.; Hartman, G.D.; Huff, J.R.; Huber, H.E.; Duggan, M.E. Discovery of 2, 3, 5-trisubstituted pyridine derivatives as potent Akt1 and Akt2 dual inhibitors. Bioorg. Med. Chem. Lett. 2005, 15, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, T.; Khan, F.I.; Lobb, K.A.; Islam, A.; Ahmad, F.; Hassan, M.I. Identification and evaluation of bioactive natural products as potential inhibitors of human microtubule affinity-regulating kinase 4 (MARK4). J. Biomol. Struct. Dyn. 2019, 37, 1813–1829. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, T.; Siddiqui, S.; Shamsi, A.; Alajmi, M.F.; Hussain, A.; Islam, A.; Ahmad, F.; Hassan, M. Virtual screening approach to identify high-affinity inhibitors of serum and glucocorticoid-regulated kinase 1 among bioactive natural products: Combined molecular docking and simulation studies. Molecules 2020, 25, 823. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Khan, F.I.; Mohammad, T.; Lan, D.; Hassan, M.; Wang, Y. Identification and evaluation of inhibitors of lipase from Malassezia restricta using virtual high-throughput screening and molecular dynamics studies. Int. J. Mol. Sci. 2019, 20, 884. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, A.A.; Mohammad, T.; Hasan, G.M.; Hassan, M. Advancements in docking and molecular dynamics simulations towards ligand-receptor interactions and structure-function relationships. Curr. Top. Med. Chem. 2018, 18, 1755–1768. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Olson, A.J. Using autodock for ligand-receptor docking. Curr. Protoc. Bioinform. 2008, 24, 8–14. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- Mohammad, T.; Mathur, Y.; Hassan, M.I. InstaDock: A single-click graphical user interface for molecular docking-based virtual high-throughput screening. Brief. Bioinform. 2021, 22, 279. [Google Scholar] [CrossRef]
- Discovery Studio; Version 2.1; Accelrys: San Diego, CA, USA, 2008.
- Mohanraj, K.; Karthikeyan, B.S.; Vivek-Ananth, R.; Chand, R.B.; Aparna, S.; Mangalapandi, P.; Samal, A. IMPPAT: A curated database of I ndian M edicinal P lants, P hytochemistry A nd T herapeutics. Sci. Rep. 2018, 8, 4329. [Google Scholar] [CrossRef] [Green Version]
- Lipinski, C.A. Lead-and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Reichel, A.; Lienau, P. Pharmacokinetics in drug discovery: An exposure-centred approach to optimising and predicting drug efficacy and safety. In New Approaches to Drug Discover; Springer: Cham, Switzerland, 2015; pp. 235–260. [Google Scholar]
- Yadav, D.K.; Kumar, S.; Choi, E.-H.; Chaudhary, S.; Kim, M.-H. Computational modeling on aquaporin-3 as skin cancer target: A virtual screening study. Front. Chem. 2020, 8, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waseem, R.; Anwar, S.; Khan, S.; Shamsi, A.; Hassan, M.; Anjum, F.; Shafie, A.; Islam, A.; Yadav, D.K. MAP/Microtubule affinity regulating kinase 4 inhibitory potential of irisin: A new therapeutic strategy to combat cancer and Alzheimer’s disease. Int. J. Mol. Sci. 2021, 22, 10986. [Google Scholar] [CrossRef]
- Cheon, J.H.; Kim, K.S.; Yadav, D.K.; Kim, M.; Kim, H.S.; Yoon, S. The JAK2 inhibitors CEP-33779 and NVP-BSK805 have high P-gp inhibitory activity and sensitize drug-resistant cancer cells to vincristine. Biochem. Biophys. Res. Commun. 2017, 490, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.K. Recent advances on small molecule medicinal chemistry to treat human diseases-Part III. Curr. Top. Med. Chem. 2021, 21, 1517–1518. [Google Scholar] [CrossRef]
- Baell, J.B. Feeling nature’s PAINS: Natural products, natural product drugs, and pan assay interference compounds (PAINS). J. Nat. Prod. 2016, 79, 616–628. [Google Scholar] [CrossRef]
- Pires, D.E.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.K.; Coghi, P.; Agrawal, P.; Shyamlal, B.R.K.; Yang, L.J.; Yadav, L.; Peng, Y.; Sharma, R.; Yadav, D.K.; Sahal, D. Design, synthesis, structure-activity relationship and docking studies of novel functionalized arylvinyl-1, 2, 4-trioxanes as potent antiplasmodial as well as anticancer agents. ChemMedChem 2020, 15, 1216–1228. [Google Scholar] [CrossRef]
- Kalani, K.; Yadav, D.K.; Alam, S.; Khan, F.; Kashyap, M.P.; Srivastava, S.K.; Pant, A.B. In-silico studies and wet-lab validation of camptothecin derivatives for anti-cancer activity against liver (HepG2) and lung (A549) cancer cell lines. Curr. Top. Med. Chem. 2021, 21, 908–919. [Google Scholar] [CrossRef]
- Teli, M.K.; Kumar, S.; Yadav, D.K.; Kim, M.-h. In silico identification of hydantoin derivatives: A novel natural prolyl hydroxylase inhibitor. J. Biomol. Struct. Dyn. 2021, 39, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Shyamlal, B.R.K.; Yadav, L.; Tiwari, M.K.; Mathur, M.; Prikhodko, J.I.; Mashevskaya, I.V.; Yadav, D.K.; Chaudhary, S. Synthesis, bioevaluation, structure-activity relationship and docking studies of natural product inspired (Z)-3-benzylideneisobenzofuran-1 (3H)-ones as highly potent antioxidants and antiplatelet agents. Sci. Rep. 2020, 10, 2307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teli, M.K.; Kumar, S.; Yadav, D.K.; Kim, M.h. In silico identification of prolyl hydroxylase inhibitor by per-residue energy decomposition-based pharmacophore approach. J. Cell. Biochem. 2021, 122, 1098–1112. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.K.; Kumar, S.; Teli, M.K.; Kim, M.H. Ligand-based pharmacophore modeling and docking studies on vitamin D receptor inhibitors. J. Cell. Biochem. 2020, 121, 3570–3583. [Google Scholar] [CrossRef] [PubMed]
- Lagunin, A.; Stepanchikova, A.; Filimonov, D.; Poroikov, V. PASS: Prediction of activity spectra for biologically active substances. Bioinformatics 2000, 16, 747–748. [Google Scholar] [CrossRef]
- Anjum, F.; Ali, F.; Mohammad, T.; Shafie, A.; Akhtar, O.; Abdullaev, B.; Hassan, M.I. Discovery of Natural Compounds as Potential Inhibitors of Human Carbonic Anhydrase II: An Integrated Virtual Screening, Docking, and Molecular Dynamics Simulation Study. OMICS J. Integr. Biol. 2021, 25, 513–524. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Schüttelkopf, A.W.; Van Aalten, D.M. PRODRG: A tool for high-throughput crystallography of protein–ligand complexes. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef] [Green Version]
- Turner, P. XMGRACE: Center for Coastal and Land-Margin Research; Version 5.1. 19; Oregon Graduate Institute of Science and Technology: Beaverton, OR, USA, 2005. [Google Scholar]
- Yadav, D.K.; Kumar, S.; Misra, S.; Yadav, L.; Teli, M.; Sharma, P.; Chaudhary, S.; Kumar, N.; Choi, E.H.; Kim, H.S. Molecular insights into the interaction of RONS and Thieno [3,2-c] pyran analogs with SIRT6/COX-2: A molecular dynamics study. Sci. Rep. 2018, 8, 4777. [Google Scholar] [CrossRef]
- Addie, M.; Ballard, P.; Buttar, D.; Crafter, C.; Currie, G.; Davies, B.R.; Debreczeni, J.; Dry, H.; Dudley, P.; Greenwood, R. Discovery of 4-Amino-N-[(1S)-1-(4-chlorophenyl)-3-hydroxypropyl]-1-(7 H-pyrrolo [2,3-d] pyrimidin-4-yl) piperidine-4-carboxamide (AZD5363), an Orally Bioavailable, Potent Inhibitor of Akt Kinases. J. Med. Chem. 2013, 56, 2059–2073. [Google Scholar] [CrossRef]
- Khan, A.; Mohammad, T.; Shamsi, A.; Hussain, A.; Alajmi, M.F.; Husain, S.A.; Iqbal, M.A.; Hassan, M.I. Identification of plant-based hexokinase 2 inhibitors: Combined molecular docking and dynamics simulation studies. J. Biomol. Struct. Dyn. 2021, 2021, 1942217. [Google Scholar] [CrossRef] [PubMed]
- Amir, M.; Mohammad, T.; Prasad, K.; Hasan, G.M.; Kumar, V.; Dohare, R.; Islam, A.; Ahmad, F.; Imtaiyaz Hassan, M. Virtual high-throughput screening of natural compounds in-search of potential inhibitors for protection of telomeres 1 (POT1). J. Biomol. Struct. Dyn. 2020, 38, 4625–4634. [Google Scholar] [CrossRef]
- Naz, F.; Khan, F.I.; Mohammad, T.; Khan, P.; Manzoor, S.; Hasan, G.M.; Lobb, K.A.; Luqman, S.; Islam, A.; Ahmad, F. Investigation of molecular mechanism of recognition between citral and MARK4: A newer therapeutic approach to attenuate cancer cell progression. Int. J. Biol. Macromol. 2018, 107, 2580–2589. [Google Scholar] [CrossRef]
- Fatima, S.; Mohammad, T.; Jairajpuri, D.S.; Rehman, M.T.; Hussain, A.; Samim, M.; Ahmad, F.J.; Alajmi, M.F.; Hassan, M.I. Identification and evaluation of glutathione conjugate gamma-l-glutamyl-l-cysteine for improved drug delivery to the brain. J. Biomol. Struct. Dyn. 2020, 38, 3610–3620. [Google Scholar] [CrossRef] [PubMed]
- Lobanov, M.Y.; Bogatyreva, N.; Galzitskaya, O. Radius of gyration as an indicator of protein structure compactness. Mol. Biol. 2008, 42, 623–628. [Google Scholar] [CrossRef]
- Hubbard, R.E.; Haider, M.K. Hydrogen Bonds in Proteins: Role and Strength; eLS: Cologne, Germany, 2010. [Google Scholar]
- Dahiya, R.; Mohammad, T.; Roy, S.; Anwar, S.; Gupta, P.; Haque, A.; Khan, P.; Kazim, S.N.; Islam, A.; Ahmad, F. Investigation of inhibitory potential of quercetin to the pyruvate dehydrogenase kinase 3: Towards implications in anticancer therapy. Int. J. Biol. Macromol. 2019, 136, 1076–1085. [Google Scholar] [CrossRef]
- Maisuradze, G.G.; Liwo, A.; Scheraga, H.A. Principal component analysis for protein folding dynamics. J. Mol. Biol. 2009, 385, 312–329. [Google Scholar] [CrossRef] [Green Version]
- Altis, A.; Otten, M.; Nguyen, P.H.; Hegger, R.; Stock, G. Construction of the free energy landscape of biomolecules via dihedral angle principal component analysis. J. Chem. Phys. 2008, 128, 6B620. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Compound ID | Phytochemical Name | Affinity (kcal/mol) | pKi | Ligand Efficiency |
|---|---|---|---|---|---|
| 1. | 174880 | Lactupicrin | −10.1 | 7.41 | 0.34 |
| 2. | 120698 | Capnoidine | −10.0 | 7.33 | 0.37 |
| 3. | 308140 | Diospyrin | −10.0 | 7.33 | 0.36 |
| 4. | 101277405 | Arboreol | −10.0 | 7.33 | 0.36 |
| 5. | 104940 | Withanolide S | −9.9 | 7.26 | 0.39 |
| 6. | 3822 | Cubebol | −9.9 | 7.26 | 0.34 |
| 7. | 5315739 | N-Acetyldehydroanonaine | −9.8 | 7.19 | 0.43 |
| 8. | 20055371 | - | −9.7 | 7.11 | 0.40 |
| 9. | 14630495 | Tuberosin | −9.7 | 7.11 | 0.39 |
| 10. | 146680 | (2,2′-Binaphthalene)-5,5′,8,8′-tetrone, 1,1′-dihydroxy-6,6′-dimethyl- | −9.7 | 7.11 | 0.35 |
| 11. | 16745513 | 4-Hydroxysesamin | −9.7 | 7.11 | 0.36 |
| 12. | 5317297 | 5-(6,7-Dihydroxy-2-oxochromen-5-yl)-6,7-dihydroxychromen-2-one | −9.7 | 7.11 | 0.37 |
| 13. | 101746 | Sesamolin | −9.6 | 7.04 | 0.35 |
| 14. | 12309624 | Papaverrubine B | −9.6 | 7.04 | 0.34 |
| 15. | 196979 | Frangulin A | −9.6 | 7.04 | 0.32 |
| 16. | 2913 | Acora-4,10-diene | −9.5 | 6.97 | 0.43 |
| 17. | 73393 | Lagerine | −9.5 | 6.97 | 0.36 |
| 18. | 21723446 | Pseudostrychnine | −9.5 | 6.97 | 0.36 |
| 19. | 5316097 | Corylin | −9.5 | 6.97 | 0.39 |
| 20. | 329584 | 3,3′-Bisjuglone | −9.5 | 6.97 | 0.36 |
| 21. | 5281806 | Psoralidin | −9.5 | 6.97 | 0.38 |
| 22. | 119204 | Roemerine | −9.4 | 6.89 | 0.45 |
| 23. | 5320486 | Gomezine | −9.4 | 6.89 | 0.47 |
| 24. | 5459059 | Ribasine | −9.4 | 6.89 | 0.36 |
| 25. | 91572 | Phaseolin | −9.4 | 6.89 | 0.39 |
| 26. | 94577 | Cepharadione A | −9.3 | 6.82 | 0.40 |
| 27. | 101595 | Vomicine | −9.3 | 6.82 | 0.33 |
| 28. | 5320772 | Psoralenol | −9.3 | 6.82 | 0.37 |
| 29. | 5490819 | Villosol | −9.3 | 6.82 | 0.31 |
| 30. | 11438278 | Cryptodorine | −9.2 | 6.75 | 0.40 |
| S. No. | Compound ID | Phytochemical | Absorption | Distribution | Metabolism | Excretion | Toxicity |
|---|---|---|---|---|---|---|---|
| GI Absorption (%) | BBB Permeation | CYP2D6 Inhibitor | OCT2 Substrate | AMES and Hepatotoxicity | |||
| 1. | 329584 | 3,3′-Bisjuglone | High | No | No | No | No |
| 2. | 5320772 | Psoralenol | High | No | Yes | No | No |
| 3. | 5459059 | Ribasine | High | Yes | Yes | No | No |
| 4. | 5490819 | Villosol | High | No | Yes | No | No |
| 5. | 12309624 | Papaverrubine B | High | Yes | Yes | No | No |
| 6. | 14630495 | Tuberosin | High | Yes | Yes | No | No |
| S. No. | Phytochemical | Molecular Structure | Pa | Pi | Activity |
|---|---|---|---|---|---|
| 1. | 3,3′-Bisjuglone |  | 0866 | 0003 | Antimutagenic |
| 0879 | 0016 | Membrane integrity agonist | |||
| 0828 | 0009 | Antineoplastic | |||
| 0799 | 0006 | Kinase inhibitor | |||
| 0773 | 0007 | Caspase-3 stimulant | |||
| 2. | Psoralenol |  | 0704 | 0008 | Anticarcinogenic |
| 0631 | 0023 | Apoptosis agonist | |||
| 0535 | 0013 | Antimutagenic | |||
| 0564 | 0053 | Antineoplastic | |||
| 0506 | 0033 | Kinase inhibitor | |||
| 3. | Ribasine |  | 0878 | 0002 | Neurotransmitter uptake inhibitor |
| 0512 | 0005 | Antineoplastic alkaloid | |||
| 0474 | 0023 | Caspase-8 stimulant | |||
| 0345 | 0079 | Caspase-3 stimulant | |||
| 0276 | 0029 | Antineoplastic (lymphoma) | |||
| 4. | Villosol |  | 0993 | 0001 | TP53 expression enhancer |
| 0902 | 0004 | Apoptosis agonist | |||
| 0854 | 0007 | Antineoplastic | |||
| 0662 | 0007 | Antineoplastic (breast cancer) | |||
| 0556 | 0006 | Prostate cancer treatment | |||
| 5. | Papaverrubine B |  | 0683 | 0029 | Antineoplastic |
| 0526 | 0028 | Antineoplastic (non-Hodgkin lymphoma) | |||
| 0503 | 0006 | Antineoplastic alkaloid | |||
| 0474 | 0009 | Antimitotic | |||
| 0376 | 0027 | Antineoplastic (lung cancer) | |||
| 6. | Tuberosin |  | 0792 | 0013 | Antineoplastic |
| 0770 | 0005 | Antineoplastic (breast cancer) | |||
| 0727 | 0021 | TP53 expression enhancer | |||
| 0627 | 0023 | Apoptosis agonist | |||
| 0571 | 0005 | Antineoplastic (ovarian cancer) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adnan, M.; Jairajpuri, D.S.; Chaddha, M.; Khan, M.S.; Yadav, D.K.; Mohammad, T.; Elasbali, A.M.; Abu Al-Soud, W.; Hussain Alharethi, S.; Hassan, M.I. Discovering Tuberosin and Villosol as Potent and Selective Inhibitors of AKT1 for Therapeutic Targeting of Oral Squamous Cell Carcinoma. J. Pers. Med. 2022, 12, 1083. https://doi.org/10.3390/jpm12071083
Adnan M, Jairajpuri DS, Chaddha M, Khan MS, Yadav DK, Mohammad T, Elasbali AM, Abu Al-Soud W, Hussain Alharethi S, Hassan MI. Discovering Tuberosin and Villosol as Potent and Selective Inhibitors of AKT1 for Therapeutic Targeting of Oral Squamous Cell Carcinoma. Journal of Personalized Medicine. 2022; 12(7):1083. https://doi.org/10.3390/jpm12071083
Chicago/Turabian StyleAdnan, Mohd, Deeba Shamim Jairajpuri, Muskan Chaddha, Mohd Shahnawaz Khan, Dharmendra Kumar Yadav, Taj Mohammad, Abdelbaset Mohamed Elasbali, Waleed Abu Al-Soud, Salem Hussain Alharethi, and Md. Imtaiyaz Hassan. 2022. "Discovering Tuberosin and Villosol as Potent and Selective Inhibitors of AKT1 for Therapeutic Targeting of Oral Squamous Cell Carcinoma" Journal of Personalized Medicine 12, no. 7: 1083. https://doi.org/10.3390/jpm12071083