
Ergosterol and Its Metabolites Induce Ligninolytic Activity in the Lignin-Degrading Fungus Phanerochaete sordida YK-624


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungus Cultivation
2.2. Screening of Ligninolytic-Inducing Compounds
2.3. Identification of Ergosterol Metabolites
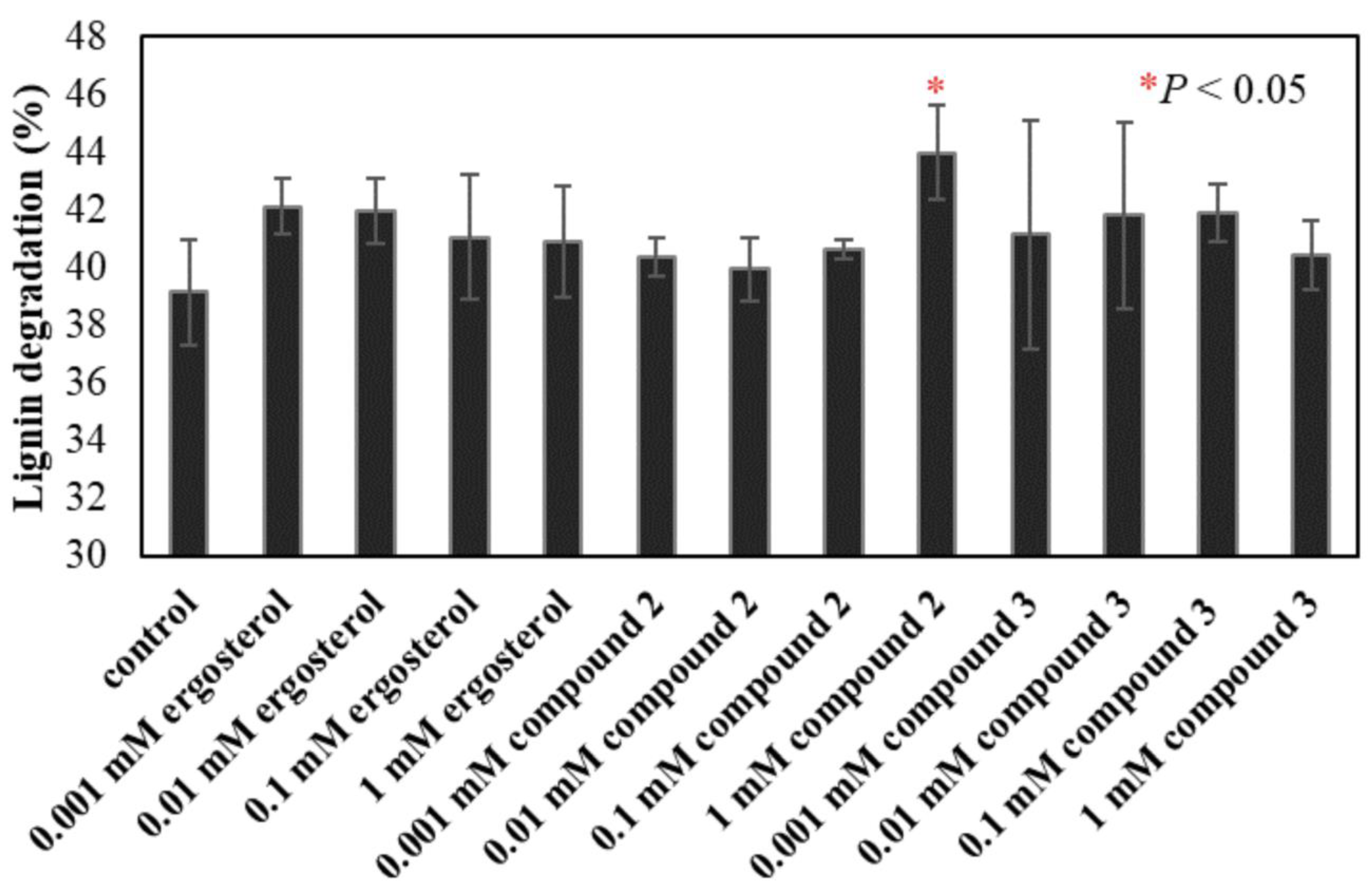
2.4. Determination of Ligninolytic Properties
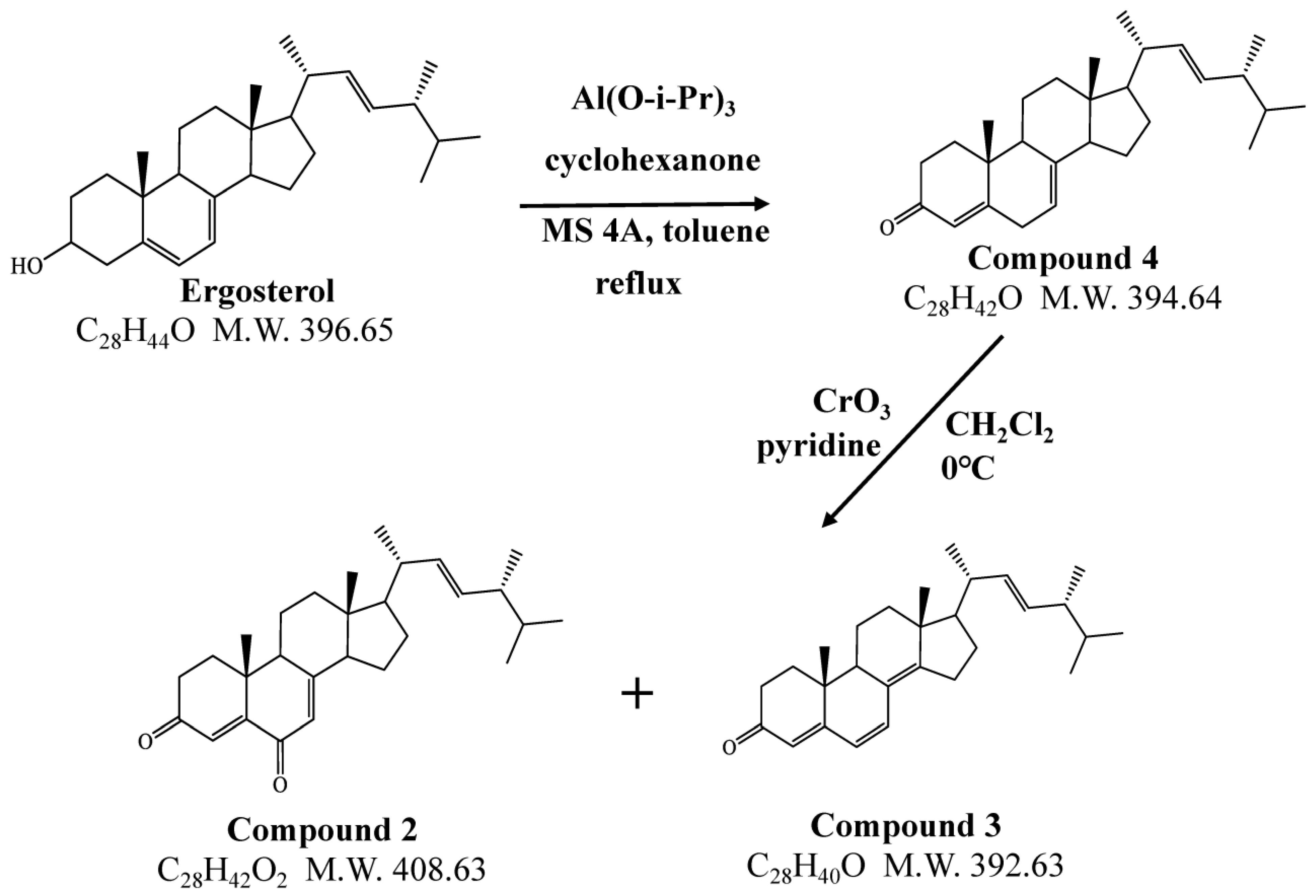
2.5. Synthesis of Ergosterol Metabolites
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J. Biological conversion of lignocellulosic biomass to ethanol. J. Biotechnol. 1997, 56, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Sarkanen, S.; Wang, Y.Y. Lignin-degrading enzyme activities. Methods Mol. Biol. 2012, 908, 251–268. [Google Scholar] [PubMed]
- Janusz, G.; Pawlik, A.; Sulej, J.; Swiderska-Burek, U.; Jarosz-Wilkolazka, A.; Paszczynski, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef] [PubMed]
- Wyman, C.E.; Dale, B.E.; Elander, R.T.; Holtzapple, M.; Ladisch, M.R.; Lee, Y.Y. Coordinated development of leading biomass pretreatment technologies. Bioresour. Technol. 2005, 96, 1959–1966. [Google Scholar] [CrossRef]
- Nakanishi, A.; Bae, J.G.; Fukai, K.; Tokumoto, N.; Kuroda, K.; Ogawa, J.; Nakatani, M.; Shimizu, S.; Ueda, M. Effect of pretreatment of hydrothermally processed rice straw with laccase-displaying yeast on ethanol fermentation. Appl. Microbiol. Biotechnol. 2012, 94, 939–948. [Google Scholar] [CrossRef]
- Bumpus, J.A.; Tien, M.; Wright, D.; Aust, S.D. Oxidation of persistent environmental pollutants by a white rot fungus. Science 1985, 228, 1434–1436. [Google Scholar] [CrossRef]
- Atiwesh, G.; Parrish, C.C.; Banoub, J.; Le, T.T. Lignin degradation by microorganisms: A review. Biotechnol. Prog. 2022, 38, e3226. [Google Scholar] [CrossRef]
- Wang, X.; Yao, B.; Su, X. Linking enzymatic oxidative degradation of lignin to organics detoxification. Int. J. Mol. Sci. 2018, 19, 3373. [Google Scholar] [CrossRef]
- Del Cerro, C.; Erickson, E.; Dong, T.; Wong, A.R.; Eder, E.K.; Purvine, S.O.; Mitchell, H.D.; Weitz, K.K.; Markillie, L.M.; Burnet, M.C.; et al. Intracellular pathways for lignin catabolism in white-rot fungi. Proc. Natl. Acad. Sci. USA 2021, 118, e2017381118. [Google Scholar] [CrossRef]
- Hirai, H.; Kondo, R.; Sakai, K. Screening of lignin degrading fungi and their ligninolytic enzyme activities during biological bleaching of kraft pulp. Mokzai Gakkaishi 1994, 40, 980–986. [Google Scholar]
- Cui, T.; Yuan, B.; Guo, H.; Tian, H.; Wang, W.; Ma, Y.; Li, C.; Fei, Q. Enhanced lignin biodegradation by consortium of white rot fungi: Microbial synergistic effects and product mapping. Biotechnol. Biofuels 2021, 14, 162. [Google Scholar] [CrossRef] [PubMed]
- Tien, M.; Kirk, T. Lignin peroxidase of Phanerochaete chrysosporium. Methods Enzymol. 1988, 161, 238–249. [Google Scholar]
- Kondo, R.; Harazono, K.; Sakai, K. Bleaching of hardwood kraft pulp with manganese peroxidase secreted from Phanerochaete sordida YK-624. Appl. Environ. Microbiol. 1994, 60, 4359–4363. [Google Scholar] [CrossRef]
- Kovganko, N.V.; Sokolov, S.N. Synthesis of (22E,24R)-ergosta-4,7-22-triene-3,6-dione and its 14α-hydroxy- and 9α,14α-dihydroxy derivatives. Chem. Nat. Compd. 1999, 35, 320–323. [Google Scholar] [CrossRef]
- Shahid, M.K.; Batool, A.; Kashif, A.; Nawaz, M.H.; Aslam, M.; Iqbal, N.; Choi, Y. Biofuels and biorefineries: Development, application and future perspectives emphasizing the environmental and economic aspects. J. Environ. Manag. 2021, 297, 113268. [Google Scholar] [CrossRef]
- Dixon, R.A.; Barros, J. Lignin biosynthesis: Old roads revisited and new roads explored. Open Biol. 2019, 9, 190215. [Google Scholar] [CrossRef]
- ten Have, R.; Teunissen, P.J. Oxidative mechanisms involved in lignin degradation by white-rot fungi. Chem. Rev. 2001, 101, 3397–3413. [Google Scholar] [CrossRef]
- Barros, J.; Temple, S.; Dixon, R.A. Development and commercialization of reduced lignin alfalfa. Curr. Opin. Biotechnol. 2019, 56, 48–54. [Google Scholar] [CrossRef]
- Wang, Y.; Gui, C.; Wu, J.; Gao, X.; Huang, T.; Cui, F.; Liu, H.; Sethupathy, S. Spatio-temporal modification of lignin biosynthesis in plants: A promising strategy for lignocellulose improvement and lignin valorization. Front. Bioeng. Biotechnol. 2022, 10, 917459. [Google Scholar] [CrossRef]
- Ralph, J.; Lapierre, C.; Boerjan, W. Lignin structure and its engineering. Curr. Opin. Biotechnol. 2019, 56, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhu, X.; Zhao, Y.; Xie, Y. The antifungal activity of o-vanillin against Aspergillus flavus via disrupting ergosterol biosynthesis and promoting oxidative stress, and an RNA-seq analysis thereof. LWT 2022, 164, 113635. [Google Scholar] [CrossRef]
- Jordá, T.; Puig, S. Regulation of ergosterol biosynthesis in Saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, Y.; Du, G.; Zhou, J.; Chen, J. Exogenous ergosterol protects Saccharomyces cerevisiae from D-limonene stress. J. Appl. Microbiol. 2013, 114, 482–491. [Google Scholar] [CrossRef]
- Sillapachaiyaporn, C.; Chuchawankul, S.; Nilkhet, S.; Moungkote, N.; Sarachana, T.; Ung, A.T.; Baek, S.J.; Tencomnao, T. Ergosterol isolated from cloud ear mushroom (Auricularia polytricha) attenuates bisphenol A-induced BV2 microglial cell inflammation. Food Res. Int. 2022, 157, 111433. [Google Scholar] [CrossRef] [PubMed]
- Rangsinth, P.; Sharika, R.; Pattarachotanant, N.; Duangjan, C.; Wongwan, C.; Sillapachaiyaporn, C.; Nilkhet, S.; Wongsirojkul, N.; Prasansuklab, A.; Tencomnao, T.; et al. Potential beneficial effects and pharmacological properties of ergosterol, a common bioactive compound in edible mushrooms. Foods 2023, 12, 2529. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Q.; Xie, Y.; Ding, Y.; Du, W.W.; Sdiri, M.; Yang, B.B. Ergosterol purified from medicinal mushroom Amauroderma rude inhibits cancer growth in vitro and in vivo by up-regulating multiple tumor suppressors. Oncotarget 2015, 6, 17832–17846. [Google Scholar] [CrossRef]
- Neto, S.L.M.; Coelho, G.D.; Ballaminut, N.; Matheus, D.R.; Thomaz, D.V.; Machado, K.M.G. Application of Deconica castanella ligninolytic enzymatic system in the degradation of hexachlorobenzene in soil. Biotechnol. Appl. Biochem. 2022, 69, 2437–2444. [Google Scholar] [CrossRef]
- Silva, R.R.; Corso, C.R.; Matheus, D.R. Effect of culture conditions on the biomass determination by ergosterol of Lentinus crinitus and Psilocybe castanella. World J. Microbiol. Biotechnol. 2010, 26, 841–846. [Google Scholar] [CrossRef]
- Zang, Q.; Chen, X.; Zhang, C.; Lin, M.; Xu, X. Improving crude protein and methionine production, selective lignin degradation and digestibility of wheat straw by Inonotus obliquus using response surface methodology. J. Sci. Food Agric. 2022, 102, 1146–1154. [Google Scholar] [CrossRef]
- Hirotani, M.; Asaka, I.; Ino, C.; Furuya, T.; Shiro, M. Ganoderic acid derivatives and ergosta-4,7,22-triene-3,6-dione from Gganoderma lucidum. Phytochemistry 1987, 26, 2797–2803. [Google Scholar] [CrossRef]
- Lee, W.; Park, Y.; Ahn, J.; Park, S.; Lee, H. Cytotoxic activity of ergosta-4,6,8,22-tetraen-3-one from the sclerotia of Polyporus umbellatus. Bull. Korean Chem. Soc. 2005, 26, 1464–1466. [Google Scholar]
- Wang, X.; Bao, H.; Bau, T. Investigation of the possible mechanism of two kinds of sterols extracted from Leucocalocybe mongolica in inducing HepG2 cell apoptosis and exerting anti-tumor effects in H22 tumor-bearing mice. Steroids 2020, 163, 108692. [Google Scholar] [CrossRef] [PubMed]
- Torres, S.; Cajas, D.; Palfner, G.; Astuya, A.; Aballay, A.; Pérez, C.; Hernández, V.; Becerra, J. Steroidal composition and cytotoxic activity from fruiting body of Cortinarius xiphidipus. Nat. Prod. Res. 2017, 31, 473–476. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Zhang, L.; Mao, J.R.; Cheng, X.H.; Lin, R.C.; Zhang, Y.; Sun, W.J. Ergosta-4,6,8,22-tetraen-3-one isolated from Polyporus umbellatus prevents early renal injury in aristolochic acid-induced nephropathy rats. J. Pharm. Pharmacol. 2011, 63, 1581–1586. [Google Scholar] [CrossRef]
- Wang, X.Y.; Jiang, S.; Liu, Y. Anti-diabetic effects of fungal ergosta-4, 6, 8, 22-tetraen-3-one from Pholiota adiposa. Steroids 2023, 192, 109185. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yin, R.; Hashizume, Y.; Todoroki, Y.; Mori, T.; Kawagishi, H.; Hirai, H. Ergosterol and Its Metabolites Induce Ligninolytic Activity in the Lignin-Degrading Fungus Phanerochaete sordida YK-624. J. Fungi 2023, 9, 951. https://doi.org/10.3390/jof9090951
Wang J, Yin R, Hashizume Y, Todoroki Y, Mori T, Kawagishi H, Hirai H. Ergosterol and Its Metabolites Induce Ligninolytic Activity in the Lignin-Degrading Fungus Phanerochaete sordida YK-624. Journal of Fungi. 2023; 9(9):951. https://doi.org/10.3390/jof9090951
Chicago/Turabian StyleWang, Jianqiao, Ru Yin, Yuki Hashizume, Yasushi Todoroki, Toshio Mori, Hirokazu Kawagishi, and Hirofumi Hirai. 2023. "Ergosterol and Its Metabolites Induce Ligninolytic Activity in the Lignin-Degrading Fungus Phanerochaete sordida YK-624" Journal of Fungi 9, no. 9: 951. https://doi.org/10.3390/jof9090951