
The Potential of Endophytic Fungi for Enhancing the Growth and Accumulation of Phenolic Compounds and Anthocyanin in Maled Phai Rice (Oryza sativa L.)

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sampling Site
2.2. Isolation of Endophytic Fungi
2.3. In Vitro Screening of Endophytic Fungi for Plant Growth-Promoting (PGP) Properties
2.3.1. Phosphate Solubilization Assay
2.3.2. Indole-3-Acetic Acid (IAA) Production
2.3.3. Gibberellic Acid (GA3) Production
2.3.4. Ammonia Production
2.3.5. Siderophore Production
2.3.6. Determination of Extracellular Enzyme Production
2.4. Identification of Endophytic Fungi
2.4.1. Morphological Identification
2.4.2. Molecular Identification
2.5. Effect of Endophytic Fungi on the Growth Promotion of Maled Phai
2.5.1. Endophytic Fungi Inoculum Preparation
2.5.2. Preparation of Rice Seeds
2.5.3. Experimental Design and Treatment
2.5.4. Determination of Plant Growth Parameters
2.5.5. Determination of Functional Substances
2.6. Statistical Analysis
3. Results
3.1. Isolation of Endophytic Fungi
3.2. In Vitro Screening of Endophytic Fungi for Plant Growth-Promoting (PGP) Properties
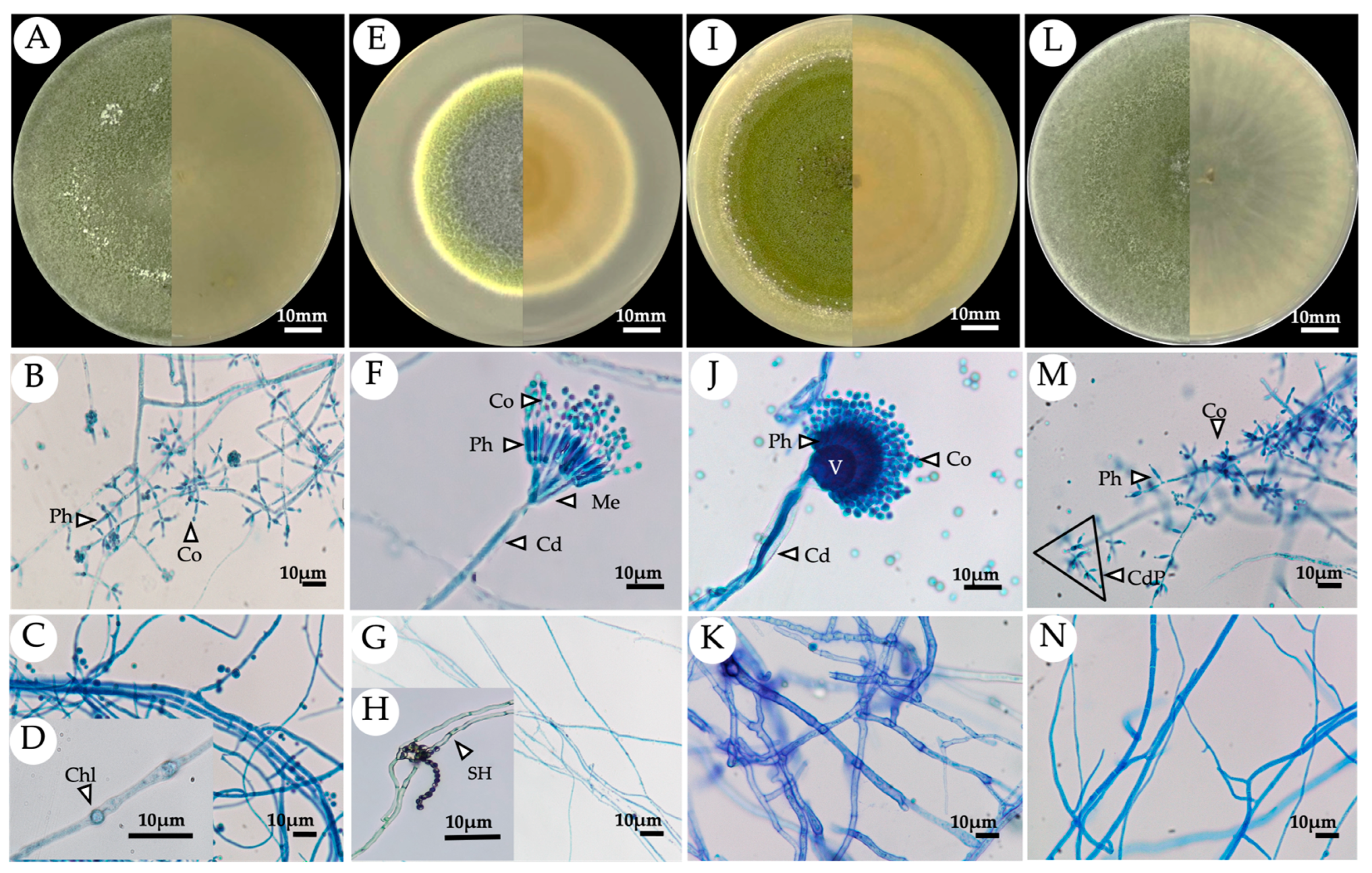
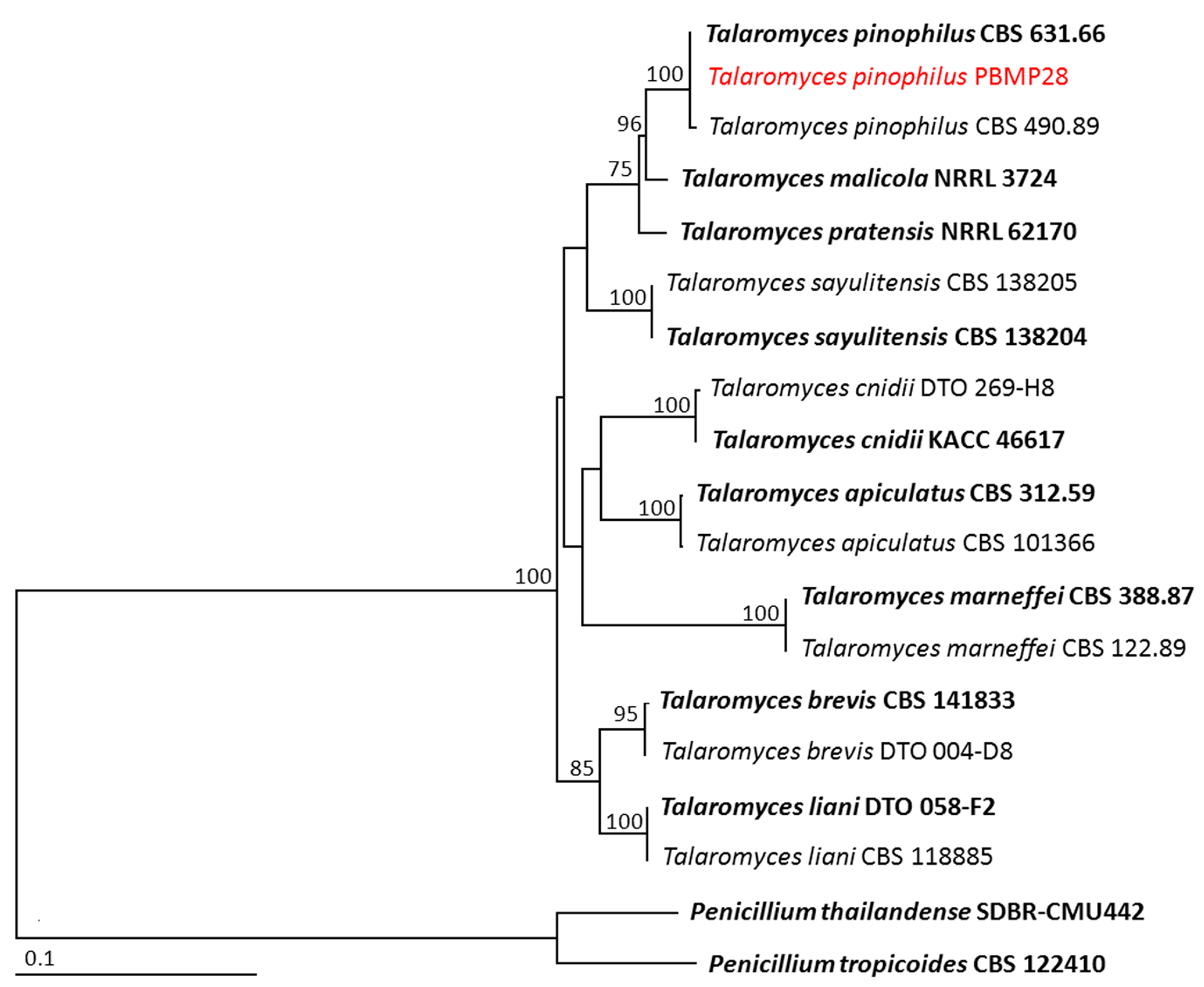
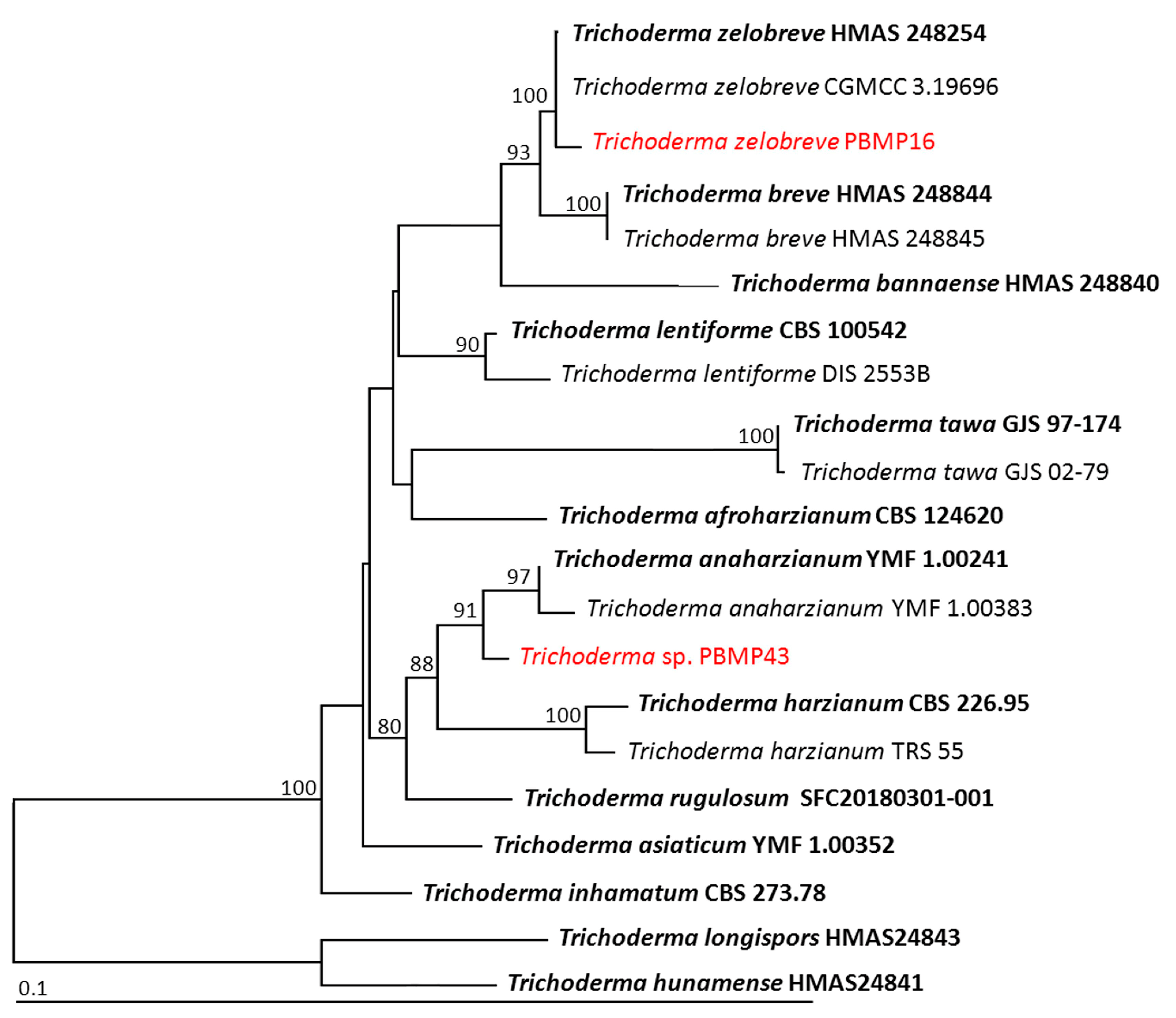
3.3. Identification of the Selected Endophytic Fungal Isolates
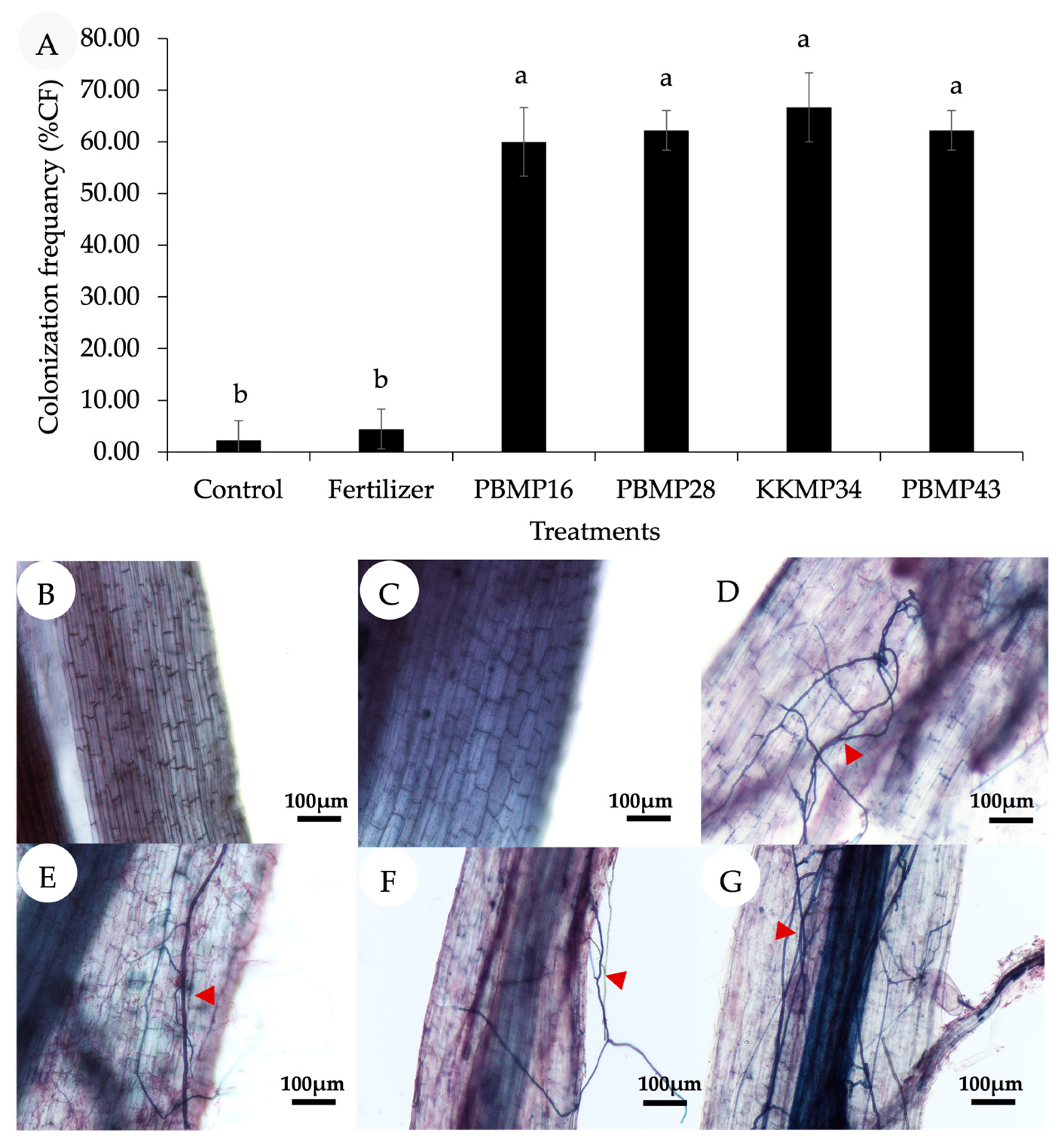
3.4. Effect of Endophytic Fungi on the Growth Promotion of Upland Rice in Pot under Greenhouse Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaudhari, P.R.; Tamrakar, N.; Singh, L.; Tandon, A.; Sharma, D.; Prabha Chaudhari, C.R. Rice nutritional and medicinal properties: A review article. J. Pharmacogn. Phytochem. 2018, 7, 150–156. [Google Scholar]
- Verma, D.K.; Srivastav, P.P. Bioactive compounds of rice (Oryza sativa L.): Review on paradigm and its potential benefit in human health. Trends Food Sci. Technol. 2020, 97, 355–365. [Google Scholar] [CrossRef]
- Sripanidkulchai, B.; Junlatat, J.; Tuntiyasawasdikul, S.; Fangkrathok, N.; Sanitchon, J.; Chankaew, S. phytochemical and bioactivity investigation of Thai pigmented-upland rice: Dam-Mong and Ma-Led-Fy varieties. Agric. Nat. Resour. 2022, 56, 889–898. [Google Scholar] [CrossRef]
- Kapcum, N.; Uriyapongson, J.; Alli, I.; Phimphilai, S. Anthocyanins, phenolic compounds and antioxidant activities in colored corn cob and colored rice bran. Int. Food Res. J. 2016, 23, 2347–2356. [Google Scholar]
- Lazze, M.C.; Savio, M.; Pizzala, R.; Cazzalini, O.; Perucca, P.; Scovassi, A.I.; Stivala, L.A.; Bianchi, L. Anthocyanins induce cell cycle perturbations and apoptosis in different human cell lines. Carcinogenesis 2004, 25, 1427–1433. [Google Scholar] [CrossRef] [PubMed]
- Srisompun, O. Rice Economy in Northeast Thailand: Current Status and Challenges. Available online: https://ap.fftc.org.tw/article/1863 (accessed on 14 April 2023).
- Narenoot, K.; Monkham, T.; Chankaew, S.; Songsri, P.; Pattanagul, W.; Sanitchon, J. Evaluation of the tolerance of Thai indigenous upland rice germplasm to early drought stress using multiple selection criteria. Plant Genet. Resour. 2017, 15, 109–118. [Google Scholar] [CrossRef]
- Knapp, D.G.; Imrefi, I.; Boldpurev, E.; Csíkos, S.; Akhmetova, G.; Berek-Nagy, P.J.; Otgonsuren, B.; Kovács, G.M. Root-colonizing endophytic fungi of the dominant grass Stipa krylovii from a Mongolian steppe grassland. Front. Microbiol. 2019, 10, 2565. [Google Scholar] [CrossRef]
- Mengistu, A.A. Endophytes: Colonization, behaviour, and their role in defense mechanism. Int. J. Microbiol. 2020, 2020, 6927219. [Google Scholar] [CrossRef] [PubMed]
- Toppo, P.; Subba, R.; Roy, K.; Mukherjee, S.; Mathur, P. Elucidating the strategies for isolation of endophytic fungi and their functional attributes for the regulation of plant growth and resilience to stress. J. Plant Growth Regul. 2023, 42, 1342–1363. [Google Scholar] [CrossRef]
- Khaekhum, S.; Ekprasert, J.; Suebrasri, T.; Mongkolthanaruk, W.; Riddech, N.; Jogloy, S.; Boonlue, S. The first member of Exserohilum rostratum beneficial for promoting growth and yield of sunchoke (Helianthus tuberosus L.). Rhizosphere 2021, 19, 100379. [Google Scholar] [CrossRef]
- Poveda, J.; Baptista, P.; Sacristán, S.; Velasco, P. Editorial: Beneficial effects of fungal endophytes in major agricultural crops. Front Plant Sci. 2022, 13, 1061112. [Google Scholar] [CrossRef] [PubMed]
- Varga, T.; Hixson, K.K.; Ahkami, A.H.; Sher, A.W.; Barnes, M.E.; Chu, R.K.; Battu, A.K.; Nicora, C.D.; Winkler, T.E.; Reno, L.R.; et al. Endophyte-promoted phosphorus solubilization in populus. Front. Plant Sci. 2020, 11, 1585. [Google Scholar] [CrossRef] [PubMed]
- Caruso, D.J.; Palombo, E.A.; Moulton, S.E.; Zaferanloo, B. Exploring the promise of endophytic fungi: A review of novel antimicrobial compounds. Microorganisms 2022, 10, 1990. [Google Scholar] [CrossRef]
- Chand, K.; Shah, S.; Sharma, J.; Paudel, M.R.; Pant, B. Isolation, characterization, and plant growth-promoting activities of endophytic fungi from a wild orchid Vanda cristata. Plant Signal. Behav. 2020, 15, 1744294. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C. Endophytic fungi: A tool for plant growth promotion and sustainable agriculture. Mycology. 2022, 13, 39–55. [Google Scholar] [CrossRef]
- Suebrasri, T.; Somteds, A.; Harada, H.; Kanokmedhakul, S.; Jogloy, S.; Ekprasert, J.; Lumyong, S.; Boonlue, S. Novel endophytic fungi with fungicidal metabolites suppress Sclerotium disease. Rhizosphere 2020, 16, 100250. [Google Scholar] [CrossRef]
- Yu, M.; Chen, J.C.; Qu, J.Z.; Liu, F.; Zhou, M.; Ma, Y.M.; Xiang, S.Y.; Pan, X.X.; Zhang, H.B.; Yang, M.Z. Exposure to endophytic fungi quantitatively and compositionally alters anthocyanins in grape cells. Plant Physiol. Biochem. 2020, 149, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Kandar, M.; Suhandono, S.; Pugeg Aryantha, I.N. Growth promotion of rice plant by endophytic fungi. J. Pure Appl. Microbiol. 2018, 12, 1569–1577. [Google Scholar] [CrossRef]
- Mingma, R.; Duangmal, K.; Mai Sci, C.J. Characterization, antifungal activity and plant growth promoting potential of endophytic actinomycetes isolated from rice (Oryza sativa L.). Chiang Mai J. Sci. 2018, 45, 2652–2665. [Google Scholar]
- Jahn, L.; Storm-Johannsen, L.; Seidler, D.; Noack, J.; Gao, W.; Schafhauser, T.; Wohlleben, W.; Van Berkel, W.J.H.; Jacques, P.; Kar, T.; et al. The endophytic fungus Cyanodermella asteris influences growth of the nonnatural host plant Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2022, 35, 49–63. [Google Scholar] [CrossRef]
- Yodsing, N.; Kanokmedhakul, S.; Mongkolthanaruk, W.; Aimi, T.; Srisapoomi, T.; Boonlue, S. Diversity of endophytic fungi isolated from Thai medicinal plants (Casearia grewiifolia) in Khon Kaen, Thailand, and their bioactive compounds. Chiang Mai J. Sci. 2018, 45, 45–59. [Google Scholar]
- Turbat, A.; Rakk, D.; Vigneshwari, A.; Kocsubé, S.; Thu, H.; Szepesi, Á.; Bakacsy, L.; Škrbić, B.D.; Jigjiddorj, E.A.; Vágvölgyi, C.; et al. Characterization of the plant growth-promoting activities of endophytic fungi isolated from Sophora Flavescens. Microorganisms 2020, 8, 683. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.A.M.E.S.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Saengket, M.; Hyde, K.D.; Kumar, V.; Doilom, M.; Brooks, S.; Deng Pa Pae, P.T.; Mae Taeng, A.; Mai, C.; Mai Sci, C.J. Endophytic fungi from Oncosperma sp. with promising in vitro plant growth promotion and antagonistic activities. Chiang Mai J. Sci. 2021, 48, 837–852. [Google Scholar]
- Syamsia, S.; Idhan, A.; Firmansyah, A.P.; Noerfitryani, N.; Rahim, I.; Kesaulya, H.; Armus, R. Combination on endophytic fungal as the plant growth-promoting fungi (PGPF) on cucumber (Cucumis sativus). Biodiversitas 2021, 22, 1194–1202. [Google Scholar] [CrossRef]
- Mahfooz, M.; Dwedi, S.; Bhatt, A.; Raghuvanshi, S.; Bhatt, M.; Agrawal, P.K. Evaluation of antifungal and enzymatic potential of endophytic fungi isolated from Cupressus torulosa D. Don. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 4084–4100. [Google Scholar] [CrossRef]
- Passari, A.K.; Mishra, V.K.; Gupta, V.K.; Yadav, M.K.; Saikia, R.; Singh, B.P. In vitro and in vivo plant growth promoting activities and DNA fingerprinting of antagonistic endophytic actinomycetes associates with medicinal plants. PLoS ONE 2015, 10, e0139468. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Potshangbam, M.; Indira Devi, S.; Sahoo, D.; Strobel, G.A. Functional characterization of endophytic fungal community associated with Oryza sativa L. and Zea mays L. Front. Microbiol. 2017, 8, 325. [Google Scholar] [CrossRef]
- Kumla, J.; Nundaeng, S.; Suwannarach, N.; Lumyong, S. Evaluation of Multifarious Plant Growth Promoting Trials of Yeast Isolated from the Soil of Assam Tea (Camellia sinensis var. assamica) Plantations in Northern Thailand. Microorganisms 2020, 8, 1168. [Google Scholar] [CrossRef]
- Tiwari, K. The future products: Endophytic fungal metabolites. J. Biodivers. Biopros. Dev. 2015, 2, 2–7. [Google Scholar] [CrossRef]
- Niu, C.; Kebede, H.; Auld, D.L.; Woodward, J.E.; Burow, G.; Wright, R.J. A safe inexpensive method to isolate high-quality plant and fungal DNA in an open laboratory environment. Afr. J. Biotechnol. 2008, 7, 2818–2822. [Google Scholar]
- Nuangmek, W.; Aiduang, W.; Kumla, J.; Lumyong, S.; Suwannarach, N. Evaluation of a Newly Identified Endophytic Fungus, Trichoderma Phayaoense for Plant Growth Promotion and Biological Control of Gummy Stem Blight and Wilt of Muskmelon. Front. Microbiol. 2021, 12, 634772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.K.; Wang, X.C.; Zhuang, W.Y.; Cheng, X.H.; Zhao, P. New Species of Talaromyces (Fungi) Isolated from Soil in Southwestern China. Biology 2021, 10, 745. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus Section Flavi and Their Production of Aflatoxins, Ochratoxins and Other Mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Sennoi, R.; Jogloy, S.; Saksirirat, W.; Patanothai, A. Pathogenicity test of sclerotium rolfsii, a causal agent of Jerusalem artichoke (Helianthus tuberosus L.) stem rot. Asian J. Plant Sci. 2010, 9, 281–284. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Mehmood, A.; Hussain, A.; Irshad, M.; Hamayun, M.; Iqbal, A.; Khan, N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis 2018, 77, 225–235. [Google Scholar] [CrossRef]
- Leong, L.P.; Shui, G. An investigation of antioxidant capacity of fruits in Singapore markets. Food Chem. 2002, 76, 69–75. [Google Scholar] [CrossRef]
- Khan, M.S.; Gao, J.; Munir, I.; Zhang, M.; Liu, Y.; Moe, T.S.; Xue, J.; Zhang, X. Characterization of endophytic fungi, Acremonium sp., from Lilium davidii and analysis of its antifungal and plant growth-promoting effects. Biomed. Res. Int. 2021, 2021, 9930210. [Google Scholar] [CrossRef]
- Wijesooriya, W.A.D.K.; Deshappriya, N. An inoculum of endophytic fungi for improved growth of a traditional ricevariety in Sri Lanka. Trop. Plant Res. 2016, 3, 470–480. [Google Scholar] [CrossRef]
- Mehta, P.; Sharma, R.; Putatunda, C.; Walia, A. Endophytic fungi: Role in phosphate solubilization. Fungal Biol. 2019, 183–209. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Khan, A.L.; Kang, S.M.; Lee, I.J. A comparative study of phosphate solubilization and the host plant growth promotion ability of Fusarium verticillioides RK01 and Humicola sp. KNU01 under salt stress. Ann. Microbiol. 2015, 65, 585–593. [Google Scholar] [CrossRef]
- Noorjahan, A.; Aiyamperumal, B.; Anantharaman, P. Isolation and charecterisation of seaweed endophytic fungi as an efficient phosphate solubiizers. Biosci. Biotechnol. Res. Asia 2019, 16, 33–39. [Google Scholar] [CrossRef]
- Castro-Camba, R.; Sánchez, C.; Vidal, N.; Vielba, J.M. Plant development and crop yield: The role of gibberellins. Plants 2022, 11, 2650. [Google Scholar] [CrossRef]
- Bader, A.N.; Salerno, G.L.; Covacevich, F.; Consolo, V.F. Native Trichoderma harzianum strains from Argentina produce indole-3 acetic acid and phosphorus solubilization, promote growth and control wilt disease on tomato (Solanum lycopersicum L.). J. King Saud Univ. Sci. 2020, 32, 867–873. [Google Scholar] [CrossRef]
- Mazrou, Y.S.A.; Makhlouf, A.H.; Elseehy, M.M.; Awad, M.F.; Hassan, M.M. Antagonistic activity and molecular characterization of biological control agent Trichoderma harzianum from Saudi Arabia. Egypt. J. Biol. Pest Control 2020, 30, 120. [Google Scholar] [CrossRef]
- Nath, R.; Sharma, G.D.; Barooah, M. Plant growth promoting endophytic fungi isolated from tea (Camellia sinensis) shrubs of assam, India. Appl. Ecol. Environ. Res. 2015, 13, 877–891. [Google Scholar] [CrossRef]
- Hassan, S.E.D. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Kumar, V.; Dhaliwal, H.S.; Prasad, R.; Saxena, A.K. Microbiome in crops: Diversity, distribution, and potential role in crop improvement. In Crop Improvement through Microbial Biotechnology; Prasad, R., Gill, S.S., Tuteja, N., Eds.; Elsevier: Noita, India, 2018; pp. 305–332. [Google Scholar]
- Jadhav, H.P.; Sayyed, R.Z. Hydrolytic Enzymes of Rhizospheric Microbes in Crop Protection. MOJ Cell Sci. Rep. 2016, 3, 135–136. [Google Scholar] [CrossRef]
- Wei, H.; Wang, W.; Yarbrough, J.M.; Baker, J.O.; Laurens, L.; van Wychen, S.; Chen, X.; Taylor, L.E.; Xu, Q.; Himmel, M.E.; et al. Genomic, Proteomic, and Biochemical Analyses of Oleaginous Mucor Circinelloides: Evaluating Its Capability in Utilizing Cellulolytic Substrates for Lipid Production. PLoS ONE 2013, 8, e71068. [Google Scholar] [CrossRef]
- Hawar, S.N. Extracellular Enzyme of Endophytic Fungi Isolated from Ziziphus Spina Leaves as Medicinal Plant. Int. J. Biomater. 2022, 2022, 2135927. [Google Scholar] [CrossRef]
- Srinivas, C.; Sunitha, V.H.; Devi, D.N. Extracellular Enzymatic Activity of Endophytic Fungal Strains Isolated from Medicinal Plants. World J. Agric. Sci. 2013, 9, 1–9. [Google Scholar]
- Mohamed, A.H.; Abd El-Megeed, F.H.; Hassanein, N.M.; Youseif, S.H.; Farag, P.F.; Saleh, S.A.; Abdel-Wahab, B.A.; Alsuhaibani, A.M.; Helmy, Y.A.; Abdel-Azeem, A.M. Native rhizospheric and endophytic fungi as sustainable sources of plant growth promoting traits to improve wheat growth under low nitrogen input. J. Fungi 2022, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Khaekhum, S.; Ekprasert, J.; Suebrasri, T.; Seemakram, W.; Mongkolthanaruk, W.; Riddech, N.; Jogloy, S.; Boonlue, S. Co-inoculation of an endophytic and arbuscular mycorrhizal fungus improve growth and yield of Helianthus tuberosus L. under field condition. J. Fungi 2021, 7, 976. [Google Scholar] [CrossRef] [PubMed]
- Velasco, P.; Rodríguez, V.M.; Soengas, P.; Poveda, J. Trichoderma hamatum increases productivity, glucosinolate content and antioxidant potential of different leafy Brassica vegetables. Plants 2021, 10, 2449. [Google Scholar] [CrossRef]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma applications on strawberry plants modulate the physiological processes positively affecting fruit production and quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef]
- Ortega-García, J.G.; Montes-Belmont, R.; Rodríguez-Monroy, M.; Ramírez-Trujillo, J.A.; Suárez-Rodríguez, R.; Sepúlveda-Jiménez, G. Effect of Trichoderma asperellum applications and mineral fertilization on growth promotion and the content of phenolic compounds and flavonoids in onions. Sci. Hortic. 2015, 195, 8–16. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Prom-U-Thai, C. The potential of high-anthocyanin purple rice as a functional ingredient in human health. Antioxidants 2021, 10, 833. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Phosphate Solubilization (μg·mL−1) | IAA Production (μg·mL−1) | GA3 Production (μg·mL−1) | Ammonia Production (μg·mL−1) | Siderophore Production | Extracellular Enzyme Production | ||
|---|---|---|---|---|---|---|---|---|
| Endoglucanase | Pectinase | Xylanase | ||||||
| PBMP16 | 1176.67 de | 307.17 a | 298.46 a | 51.87 b | − | + | + | − |
| KKDM22 | 897.31 f | 266.90 b | 89.23 g | 35.41 d | − | nd | nd | nd |
| PBMP28 | 1082.53 e | 273.29 b | 162.86 d | 44.27 c | + | + | − | − |
| KKDM04 | 1350.00 c | 318.48 a** | 23.64 i | 12.45 g | + | nd | nd | nd |
| KKMP34 | 1055.83 ef | 160.84 d | 241.34 b | 45.23 c | + | − | + | − |
| KKMP28 | 1470.14 c | 235.49 c | 132.76 e | 32.71 d | − | nd | nd | nd |
| KKMP10 | 1692.50 b | 111.73 e | 76.21 h | 18.50 f | − | nd | nd | nd |
| PBMP43 | 2325.83 a** | 314.17 a | 301.35 a** | 65.69 a** | − | + | + | − |
| KKMP27 | 1336.67 cd | 217.77 c | 106.97 f | 27.97 e | − | nd | nd | nd |
| KKDM45 | 1085.00 e | 109.67 e | 212.50 c | 32.54 d | − | nd | nd | nd |
| Endophytic Fungi Species | Isolate | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|
| ITS | CaM | BenA | rpb2 | tef-1 | ||
| Aspergillus flavus | KKMP34 | LC738615 | OR488618 | OR488616 | – | – |
| Talaromyces pinophilus | PBMP28 | LC738614 | OR488619 | OR488617 | – | – |
| Trichodermazelobreve | PBMP16 | LC738613 | – | – | OR488620 | OR488622 |
| Trichoderma sp. | PBMP43 | LC738616 | – | – | OR488621 | OR488623 |
| Treatments | Height (cm) | Number of Tillers | Total Chlorophyll (mg·L−1) | Pn (μmol CO2·m−2·s−1) | Tr (mmol H2O·m−2·s−1) | Sc (mmol H2O·m−2·s−1) | WUE (μmol CO2 mmol·H2O−1) |
|---|---|---|---|---|---|---|---|
| Control | 85.00 e | 4.50 c | 29.83 e | 16.55 d | 3.08 b | 0.25 b | 5.37 |
| Fertilizer | 86.33 d | 15.50 a | 39.23 d | 17.30 d | 3.77 ab | 0.31 b | 4.59 |
| A. flavus KKMP34 | 94.42 bc | 7.50 b | 38.25 d | 21.10 b | 3.77 ab | 0.29 b | 5.62 |
| T. pinophilus PBMP28 | 93.00 c | 7.00 b | 41.98 c | 19.97 bc | 3.60 b | 0.28 b | 5.59 |
| Tr. zelobreve PBMP16 | 97.00 a | 7.33 b | 48.59 b | 22.35 a | 4.42 a | 0.44 a | 5.15 |
| Trichoderma sp. PBMP43 | 95.40 ab | 7.83 b | 50.47 a | 19.70 c | 3.56 b | 0.29 b | 5.58 |
| F-test | ** | ** | ** | ** | * | * | ns |
| %CV | 5.12 | 13.77 | 17.16 | 11.08 | 14.49 | 20.92 | 11.61 |
| Treatments | Number of Panicles | Total Seed Weight (g·Plant−1) | Total Shoot Weight (g·Plant−1) | Root (g·Plant−1) | |
|---|---|---|---|---|---|
| Fresh Weight | Dry Weight | ||||
| Control | 3.83 d | 3.59 c | 15.69 d | 102.57 c | 11.78 d |
| Fertilizer | 9.17 a | 8.85 b | 36.16 a | 131.08 a | 28.23 a |
| A. flavus KKMP34 | 5.83 bc | 8.77 b | 27.15 c | 121.96 ab | 20.67 bc |
| T. pinophilus PBMP28 | 5.00 cd | 7.75 b | 26.53 c | 114.29 bc | 17.55 c |
| Tr. zelobreve PBMP16 | 6.50 b | 10.69 a | 32.66 ab | 123.14 ab | 20.84 bc |
| Trichoderma sp. PBMP43 | 6.50 b | 8.79 b | 28.58 bc | 123.55 ab | 21.12 b |
| F-test | ** | ** | ** | * | ** |
| %CV | 30.49 | 30.53 | 26.83 | 12.43 | 27.45 |
| Treatments | Length (cm) | Surface Area (cm2) | Diameter (mm) | Volume (cm3) |
|---|---|---|---|---|
| Control | 3418.27 c | 673.08 c | 0.64 b | 13.50 c |
| Fertilizer | 3952.24 b | 910.09 a | 0.78 a | 18.99 ab |
| A. flavus KKMP34 | 3943.44 b | 865.81 b | 0.75 a | 17.70 ab |
| T. pinophilus PBMP28 | 3979.19 b | 868.38 b | 0.72 a | 18.97 bc |
| Tr. zelobreve PBMP16 | 4252.58 a | 862.94 b | 0.82 a | 19.05 a |
| Trichoderma sp. PBMP43 | 4260.40 a | 927.50 a | 0.83 a | 19.41 a |
| F-test | ** | ** | ** | ** |
| %CV | 7.86 | 10.25 | 12.75 | 10.27 |
| Treatments | TPC (mg·gallic eq·100 g−1 DW) | TAC (mg·Cy-3-G eq·100 g−1 DW) | Antioxidant (% Radical Scavenging) | Nutrient Uptake (mg·g−1) | ||
|---|---|---|---|---|---|---|
| N | P | K | ||||
| Control | 92.84 e | 33.06 c | 87.83 d | 0.68 d | 0.25 d | 1.37 d |
| Fertilizer | 111.61 cd | 45.09 b | 90.94 c | 1.84 ab | 0.89 a | 3.25 a |
| A. flavus KKMP34 | 116.00 bc | 47.09 b | 97.49 a | 1.59 c | 0.54 c | 2.65 c |
| T. pinophilus PBMP28 | 110.39 d | 45.09 b | 91.80 b | 1.61 c | 0.52 c | 2.56 c |
| Tr. zelobreve PBMP16 | 122.32 a | 55.77 a | 98.07 a | 1.93 a | 0.67 b | 3.18 a |
| Trichoderma sp. PBMP43 | 119.86 a | 53.10 a | 93.25 a | 1.76 b | 0.66 b | 2.94 b |
| F-test | ** | ** | ** | ** | ** | ** |
| %CV | 9.01 | 16.53 | 4.06 | 27.70 | 28.64 | 24.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gateta, T.; Nacoon, S.; Seemakram, W.; Ekprasert, J.; Theerakulpisut, P.; Sanitchon, J.; Suwannarach, N.; Boonlue, S. The Potential of Endophytic Fungi for Enhancing the Growth and Accumulation of Phenolic Compounds and Anthocyanin in Maled Phai Rice (Oryza sativa L.). J. Fungi 2023, 9, 937. https://doi.org/10.3390/jof9090937
Gateta T, Nacoon S, Seemakram W, Ekprasert J, Theerakulpisut P, Sanitchon J, Suwannarach N, Boonlue S. The Potential of Endophytic Fungi for Enhancing the Growth and Accumulation of Phenolic Compounds and Anthocyanin in Maled Phai Rice (Oryza sativa L.). Journal of Fungi. 2023; 9(9):937. https://doi.org/10.3390/jof9090937
Chicago/Turabian StyleGateta, Thanawan, Sabaiporn Nacoon, Wasan Seemakram, Jindarat Ekprasert, Piyada Theerakulpisut, Jirawat Sanitchon, Nakarin Suwannarach, and Sophon Boonlue. 2023. "The Potential of Endophytic Fungi for Enhancing the Growth and Accumulation of Phenolic Compounds and Anthocyanin in Maled Phai Rice (Oryza sativa L.)" Journal of Fungi 9, no. 9: 937. https://doi.org/10.3390/jof9090937