
Phylogenetic and Morphological Evidence Reveal Five New Species of Boletes from Southern China

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
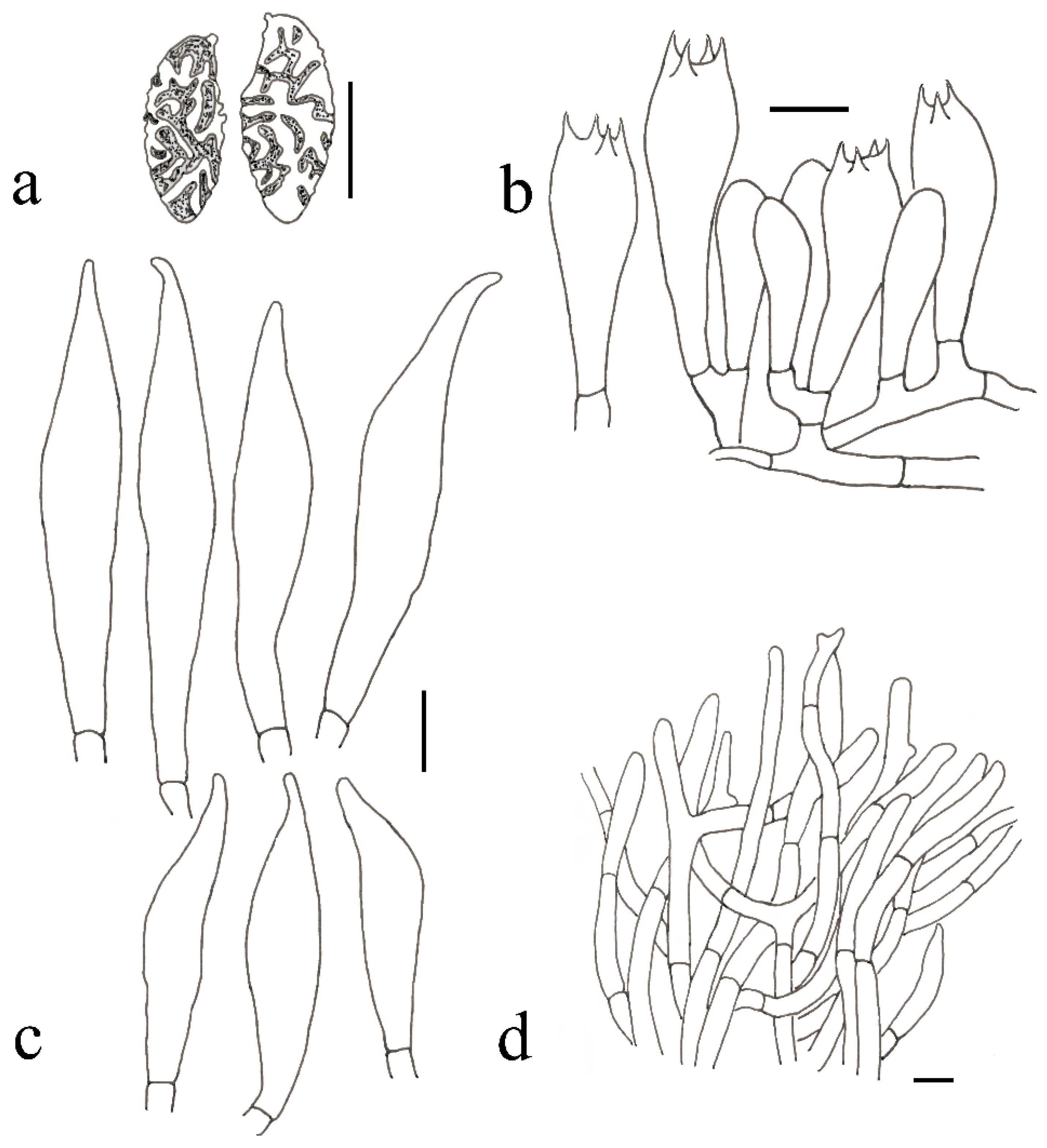
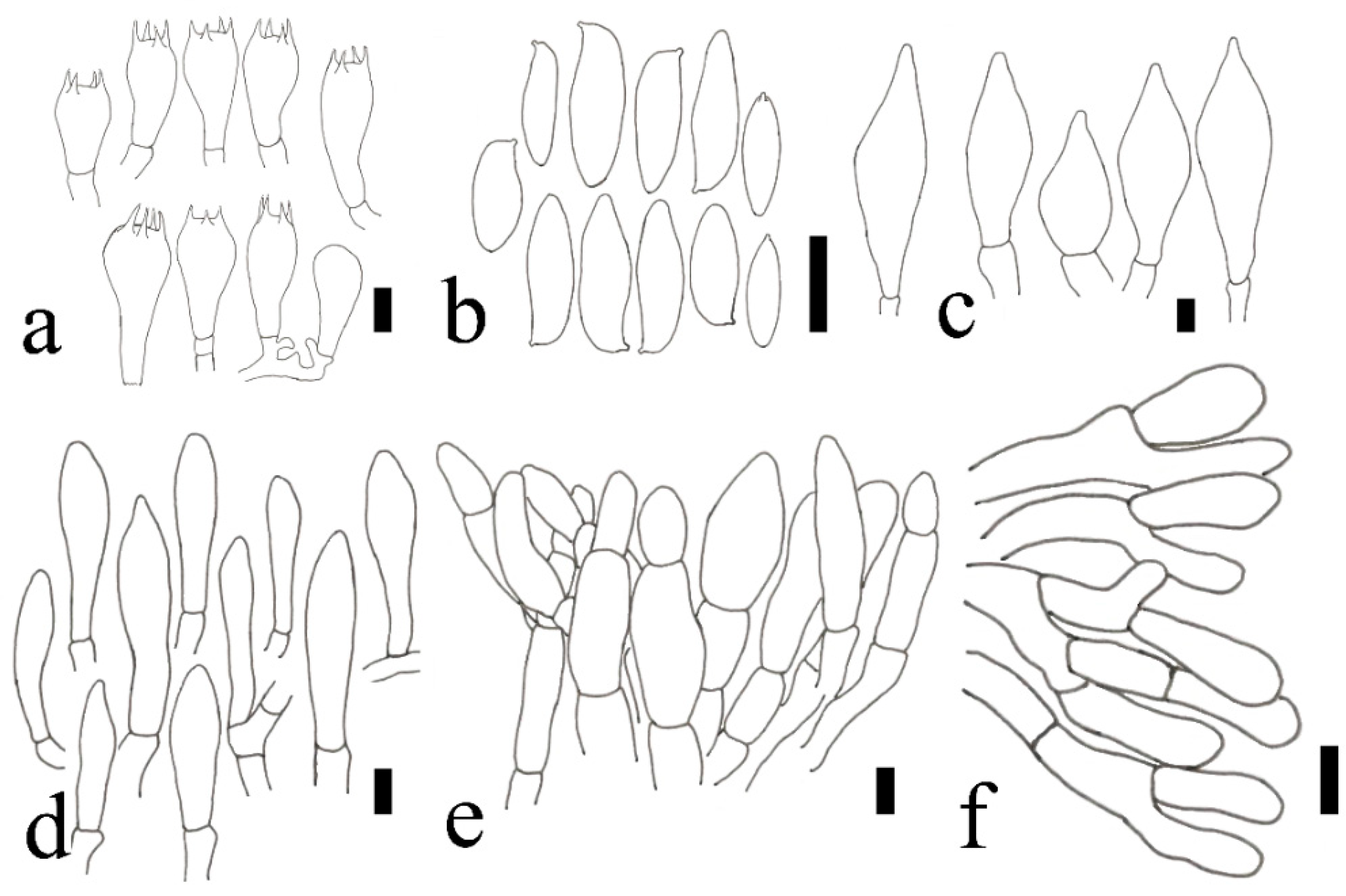
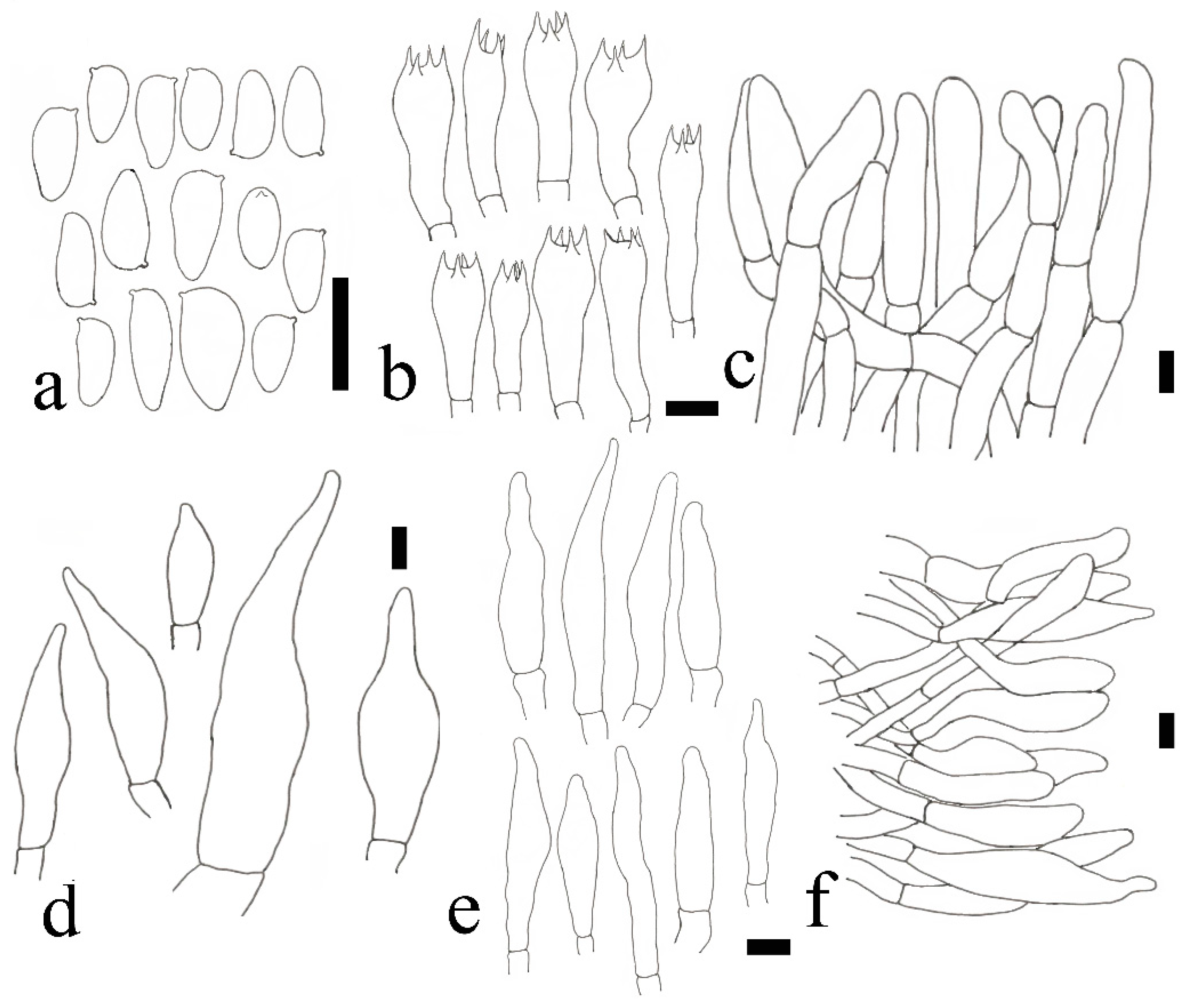
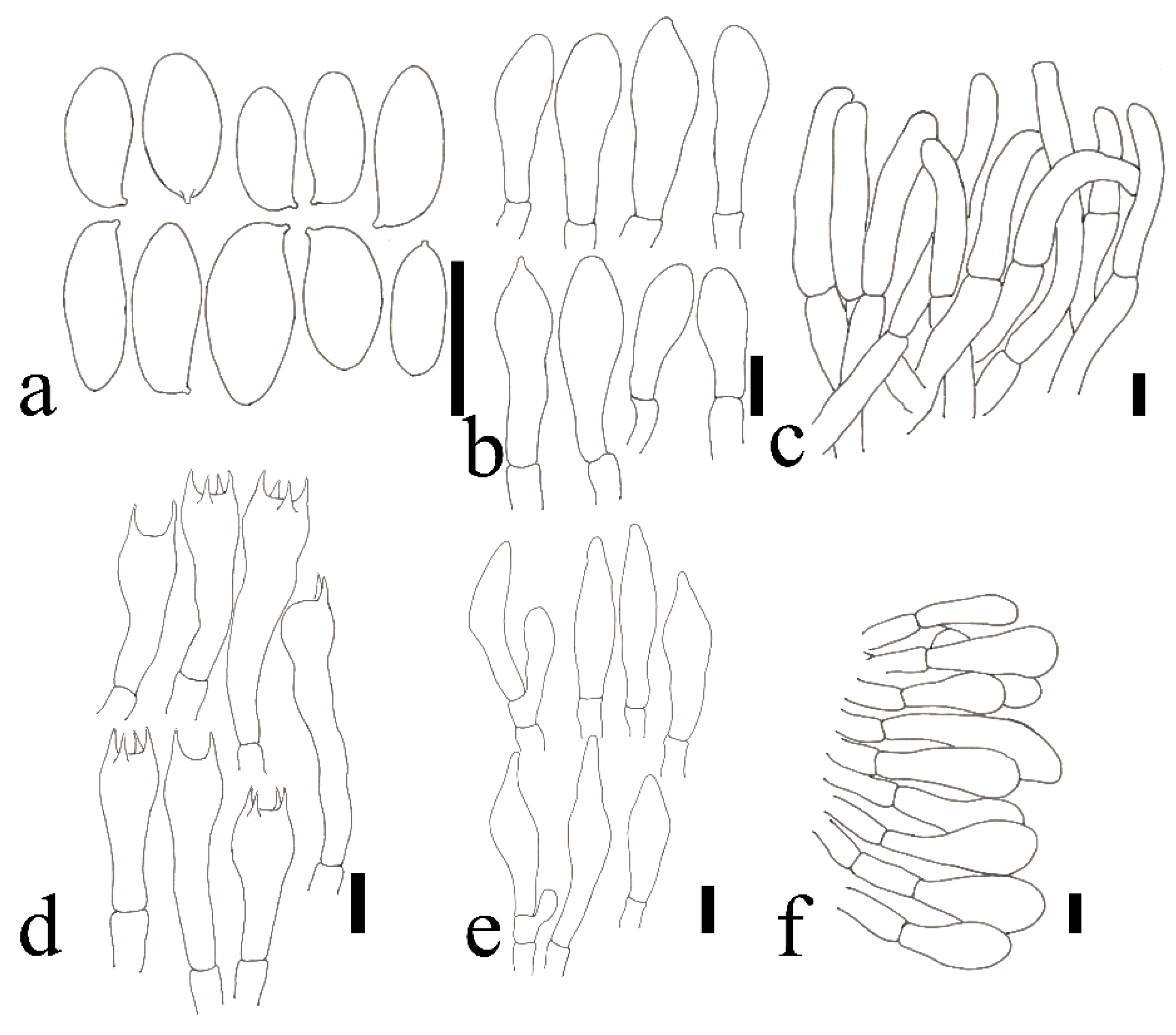
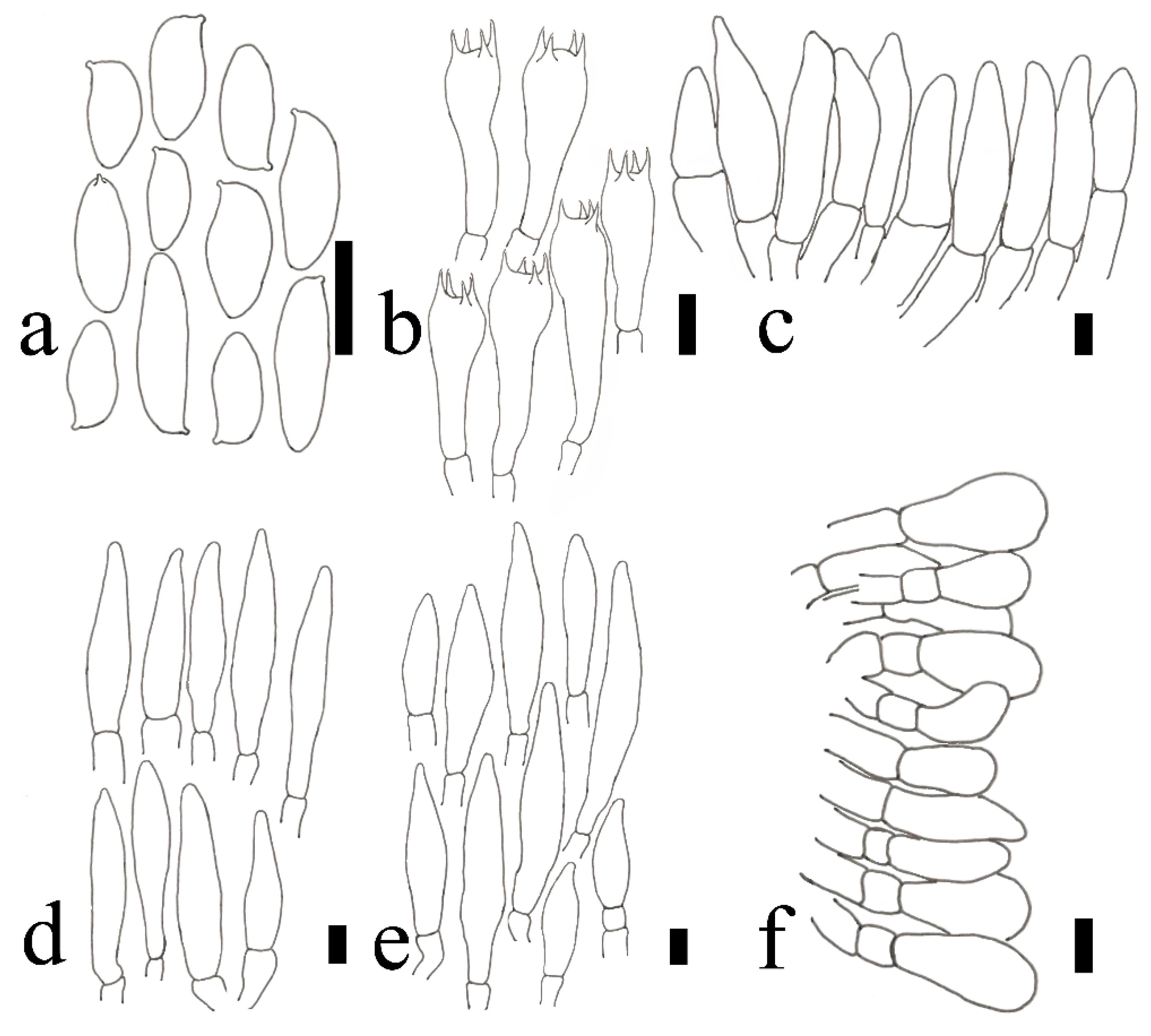
2.2. Morphological Studies
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Phylogenetic Analyses
3. Results
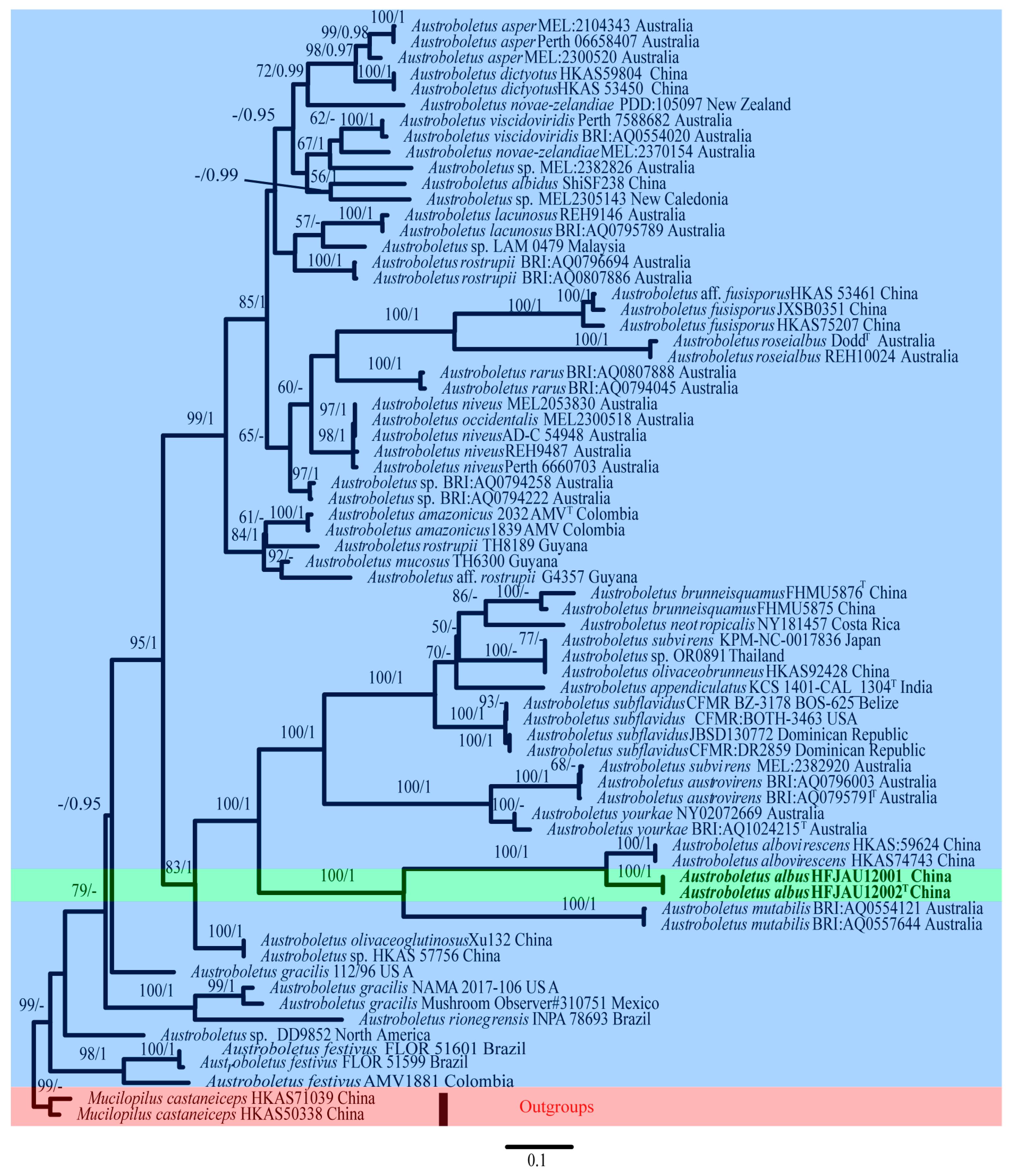
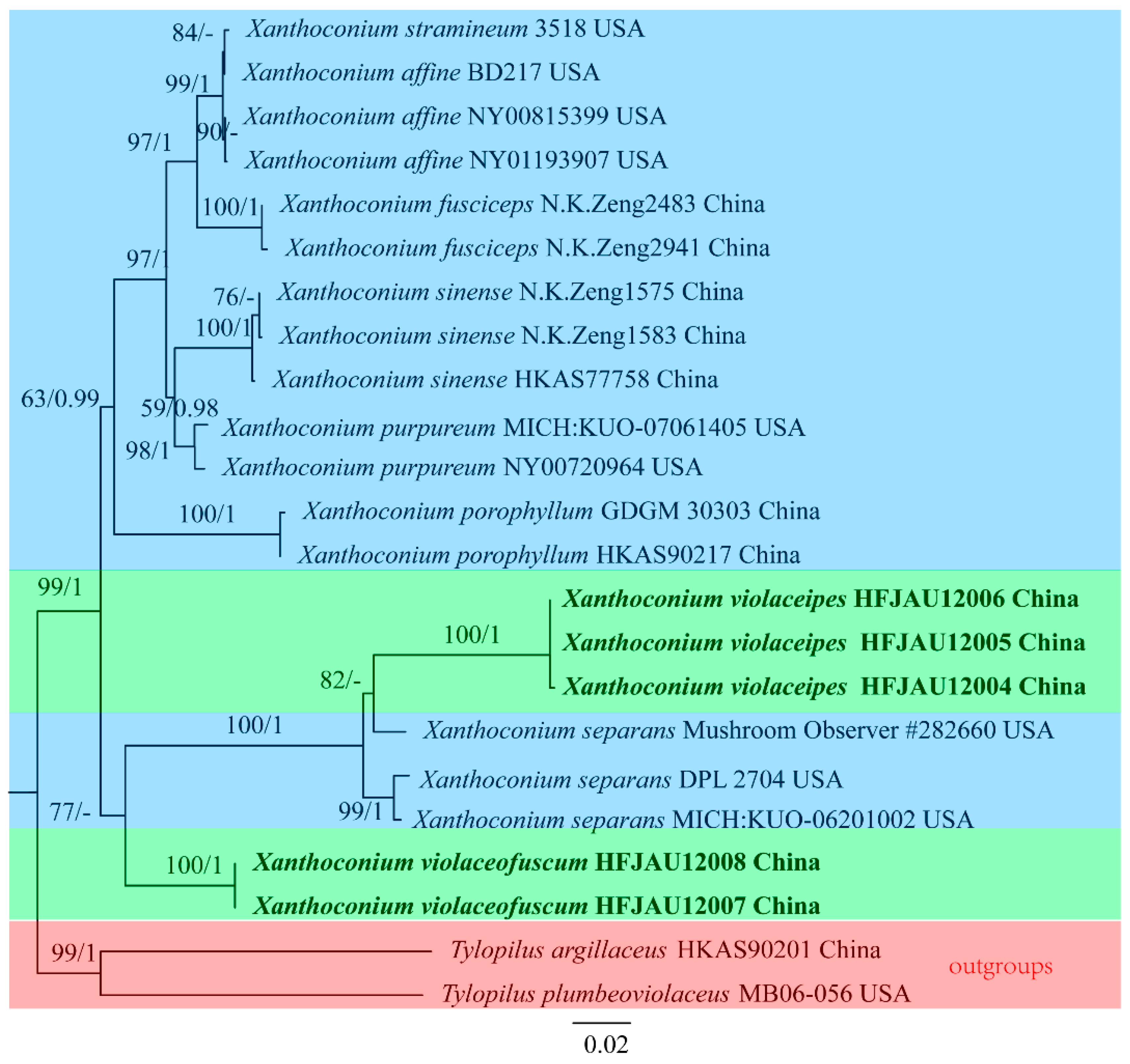
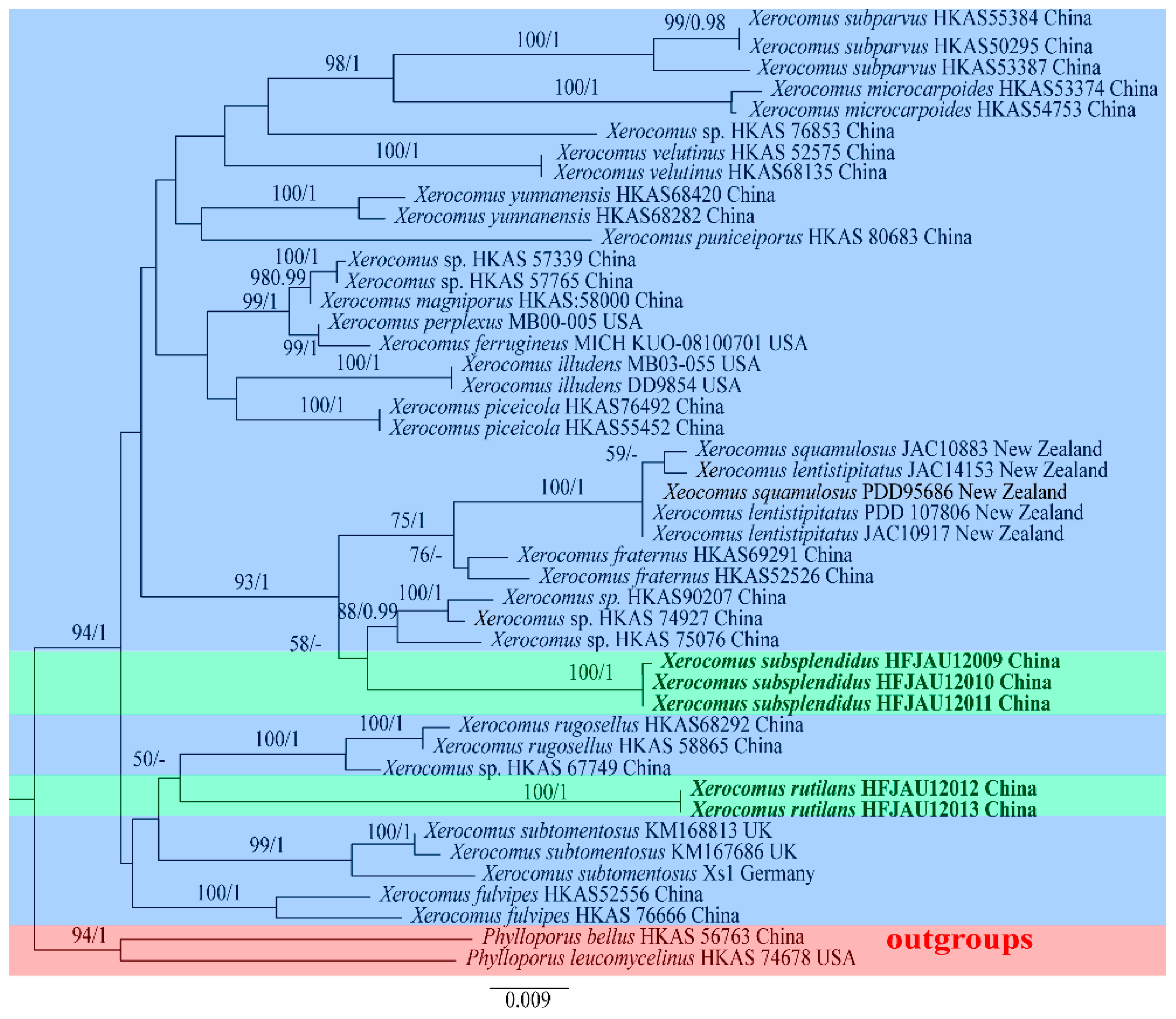
3.1. Molecular Phylogenetic Results
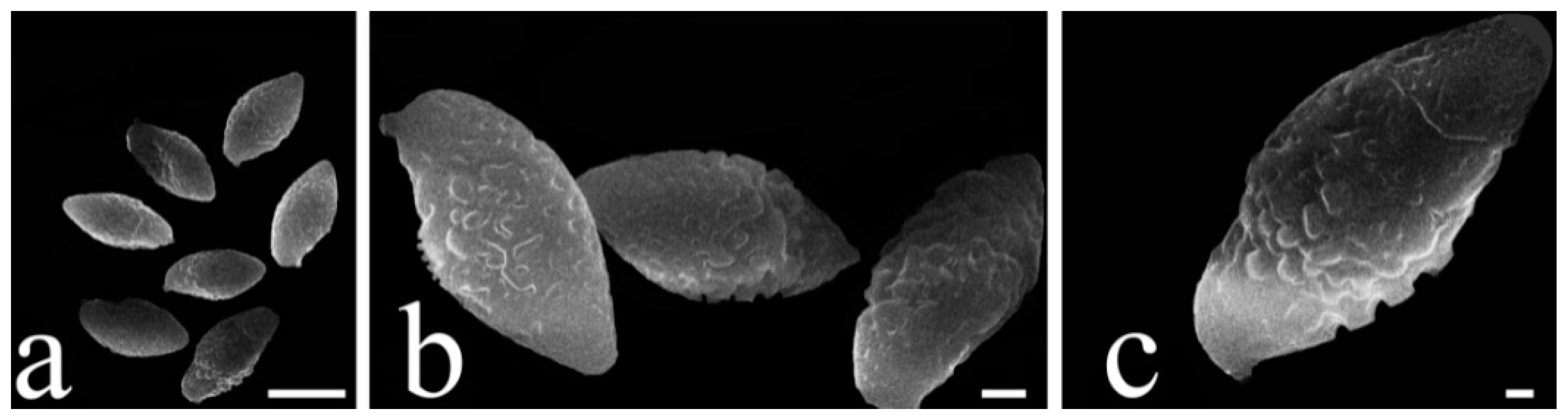
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Li, Y.C.; Yang, Z.L. The Boletes of China—Tylopilus s.l.; Springer Nature: Berlin/Heidelberg, Germany; Beijing Science Press: Beijing, China, 2021; p. 418. [Google Scholar]
- Chiu, W.F. The Boletes of Yunnan. Mycologia 1948, 40, 199–231. [Google Scholar] [CrossRef]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963. [Google Scholar]
- Yeh, K.W.; Chen, Z.C. The Boletes of Taiwan (I). Taiwania 1980, 25, 166–184. [Google Scholar]
- Bi, Z.; Lu, D.; Zheng, G. Basidiomycetes from Dinghu Mountain of China. II. Some species of Boletaceae (1). Acta Bot. Yunnanica 1982, 4, 55–64. [Google Scholar]
- Bi, Z.; Li, T.; Zheng, G.; Li, C. Basidiomycetes from Dinghu Mountain of China. III. Some species of Boletaceae (2). Acta Mycol. Sin. 1984, 3, 199–206. [Google Scholar]
- Ying, J.Z.; Ma, Q.M. New taxa and records of the genus Strobilomyces in China. Acta Mycol. Sin. 1985, 4, 95–102. [Google Scholar]
- Zang, M. Notes on the Boletales from eastern Himalayas and adjacent of China. Acta Bot Yunnanica 1985, 7, 383–401. [Google Scholar]
- Ying, J.Z. New species of the genus Xerocomus from China. Acta Microbiol. Sin. Suppl. 1986, 1, 309–315. [Google Scholar]
- Zang, M. Notes on the Boletales from eastern Himalayas and adjacent of China (2). Acta Bot Yunnanica 1986, 8, 1–22. [Google Scholar]
- Bi, Z.; Li, T. New taxon and new records of the genus Suillus from Guangdong. Acta Microbiol. Sin. 1990, 9, 20–24. [Google Scholar]
- Bi, Z.S.; Zheng, G.Y.; Li, T.H.; Wang, Y.Z. Macrofungus Flora of the Mountainous District of North Guangdong; Guangdong Science and Technology Press: Guangdong, China, 1990. [Google Scholar]
- Zang, M. Sinoboletus, a new genus of Boletaceae from China. Mycotaxon 1992, 45, 223–227. [Google Scholar]
- Bi, Z.S.; Zheng, G.Y.; Li, T.H. The Macrofungus Flora of China’s Guangdong Province; Chinese University Press: Hong Kong, China, 1993. [Google Scholar]
- Zang, M.; Li, B.; Xi, J.; Zhang, D. Fungi of Hengduan Mountains; Science Press: Beijing, China, 1996. [Google Scholar]
- Zang, M.; Li, T.; Petersen, R. Five new species of Boletaceae from China. Mycotaxon 2001, 80, 481–488. [Google Scholar]
- Zang, M. Flora Fungorum Sinicorum. Boletaceae (I); Science Press: Beijing, China, 2006; Volume 22, p. 215. [Google Scholar]
- Li, Y.C.; Feng, B.; Yang, Z.L. Zangia, a new genus of Boletaceae supported by molecular and morphological evidence. Fungal Divers. 2011, 49, 125–143. [Google Scholar] [CrossRef]
- Zhang, M.; Li, T.H.; Bau, T.; Song, B. A new species of Xerocomus from Southern China. Mycotaxon 2012, 121, 23–27. [Google Scholar] [CrossRef]
- Zang, M. Flora Fungorum Sinicorum. Boletaceae (II); Science Press: Beijing, China, 2013; Volume 44, p. 152. [Google Scholar]
- Zeng, N.K.; Liang, Z.Q.; Yang, Z.L. Boletus orientialbus, a new species with white basidioma from subtropical China. Mycoscience 2013, 55, 159–163. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, F.; Zeng, N.K.; Cui, Y.Y.; Yang, Z.L. A new genus Pseudoaustroboletus (Boletaceae, Boletales) from Asia as inferred from molecular and morphological data. Mycol. Prog. 2014, 13, 1207–1216. [Google Scholar] [CrossRef]
- Li, Y.C.; Santana, B.O.; Zeng, N.K.; Feng, B.; Yang, Z.L. Molecular phylogeny and taxonomy of the genus Veloporphyrellus. Mycologia 2014, 106, 291–306. [Google Scholar] [CrossRef]
- Zhang, M.; Li, T.H.; Song, B. A new slender species of Aureoboletus from southern China. Mycotaxon 2014, 128, 195–202. [Google Scholar] [CrossRef]
- Wu, G.; Zhao, K.; Li, Y.C.; Zeng, N.K.; Feng, B.; Halling, R.E.; Yang, Z.L. Four new genera of the fungal family Boletaceae. Fungal Divers. 2015, 81, 1–24. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Feng, B.; Wu, G.; Xu, J.P.; Yang, Z.L. Porcini mushrooms (Boletus sect. Boletus) from China. Fungal Divers. 2015, 81, 189–212. [Google Scholar] [CrossRef]
- Zeng, N.K.; Tang, L.P.; Li, Y.C.; Tolgor, B.; Zhu, X.T.; Zhao, Q.; Yang, Z.L. The genus Phylloporus (Boletaceae, Boletales) from China: Morphological and multilocus DNA sequence analyses. Fungal Divers. 2012, 58, 73–101. [Google Scholar] [CrossRef]
- Snell, W. The genera of the Boletaceae. Mycologia 1941, 33, 415–423. [Google Scholar] [CrossRef]
- Horak, E. Revision of Malaysian Species of Boletales s.l. (Basidiomycota) Described by EJH Corner (1972, 1974); Forest Research Institute Malaysia: Kuala Lumpur, Malaysia, 2011; Volume 51, p. 283. [Google Scholar]
- Wu, G.; Feng, B.; Xu, J.P.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Qi, L.L.; Fu, Y.P.; Wang, F.J.; Song, B.; Li, Y. Suillus foetidus (Boletales, Basidiomycota), a new species from northeast China. Phytotaxa 2016, 260, 167–175. [Google Scholar] [CrossRef]
- Liang, Z.Q.; Chai, H.; Jiang, S.; Ye, Z.K.; Zeng, N.K. The genus Xanthoconium (Boletaceae, Boletales) in tropical China. Phytotaxa 2017, 295, 246–254. [Google Scholar] [CrossRef]
- Qi, L.L.; Fu, Y.P.; Lang, N.; Bai, X.; Li, Y. A new species of Gomphidius from Northeast China. Phytotaxa 2017, 316, 181–188. [Google Scholar] [CrossRef]
- Chai, H.; Liang, Z.Q.; Xue, R.; Jiang, S.; Luo, S.H.; Wang, Y.; Wu, L.L.; Tang, L.P.; Chen, Y.; Hong, D.; et al. New and noteworthy boletes from subtropical and tropical China. MycoKeys 2019, 46, 55–96. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Wu, G.; Zhao, K. Aureoboletus rubellus, a new species of bolete from Jiangxi Province, China. Phytotaxa 2019, 420, 72–78. [Google Scholar] [CrossRef]
- Xue, R.O.U.; Liang, Z.Q.; Chai, H.U.I.; Jiang, S.; Tang, L.P.; Fu, Y.Q.; Fan, Y.G.; Wu, L.L.; Zeng, N.K. The Suillus spraguei complex (Suillaceae, Boletales): New taxon, new hosts and amended descriptions. Phytotaxa 2019, 401, 239–256. [Google Scholar] [CrossRef]
- Zhang, M.; Li, T.H.; Wang, C.Q.; Zeng, N.K.; Deng, W.Q. Phylogenetic overview of Aureoboletus (Boletaceae, Boletales), with descriptions of six new species from China. MycoKeys 2019, 61, 111–145. [Google Scholar] [CrossRef]
- Han, L.H.; Wu, G.; Horak, E.; Halling, R.E.; Xu, J.; Ndolo, E.S.T.; Sato, H.; Fechner, N.; Sharma, Y.P.; Yang, Z.L. Phylogeny and species delimitation of Strobilomyces (Boletaceae), with an emphasis on the Asian species. Persoonia 2020, 44, 113–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.I.; Su, M.S.; Jiang, S.; Xue, R.O.U.; Wu, L.L.; Xie, H.J.; Zhang, Y.Z.; Liang, Z.Q.; Zeng, N.K. The genus Hourangia in China and a description of Aureoboletus erythraeus sp. nov. Phytotaxa 2020, 472, 87–106. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, M.; Wu, X.L.; Wu, G.; Xu, J.P.; Yang, Z.L.; Li, Y.C. Cyanescent Gyroporus (Gyroporaceae, Boletales) from China. MycoKeys 2021, 81, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Mi, H.X.; Xie, H.J.; Zhang, X.; Chen, Y.; Liang, Z.Q.; Zeng, N.K. Neoboletus infuscatus, a new tropical bolete from Hainan, southern China. Mycoscience 2021, 62, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xie, D.C.; Wang, C.Q.; Deng, W.Q.; Li, T.H. New insights into the genus Gyroporus (Gyroporaceae, Boletales), with establishment of four new sections and description of five new species from China. Mycology 2022, 13, 223–242. [Google Scholar] [CrossRef]
- Zhou, F.; Gao, Y.; Song, H.Y.; Hu, H.J.; Li, S.X.; Liu, J.H.; Hu, D.M. Retiboletus atrofuscus (Boletaceae, Boletales), a new species from China. Arch. Microbiol. 2022, 204, 381. [Google Scholar] [CrossRef]
- Chen, X.N.; Zhang, M.; Li, T.H.; Zeng, N.K. A new species of Heimioporus (Boletaceae) from southern China. Phytotaxa 2019, 415, 179–188. [Google Scholar] [CrossRef]
- Zeng, N.K.; Liang, Z.Q.; Wu, G.; Li, Y.C.; Yang, Z.L. The genus Retiboletus in China. Mycologia 2016, 108, 363–380. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, F.M.; Zeng, Q.Q.; Han, L.H.; Li, Y.C. Tylopilus jiangxiensis, a new species of Tylopilus s. str. from China. Phytotaxa 2020, 434, 281–291. [Google Scholar] [CrossRef]
- Huo, G.H.; Yan, J.Q.; Zhang, L.P.; Hu, D.M. Illustrated Handbook of Macrofungi in Jiangxi; Jiangxi Science And Technology Press: Nanchang, China, 2020; p. 296. [Google Scholar]
- Zhang, J.; Song, H.; Hu, D. A Checklist of Macrofungi in Jiangxi Province. Biol. Disaster Sci. 2016, 39, 1–13. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben 3; Aufl; Muster-Schmid: Northeim, Germany, 1981; p. 242. [Google Scholar]
- Zeng, N.K.; Cai, Q.; Yang, Z.L. Corneroboletus, a new genus to accommodate the southeastern Asian Boletus indecorus. Mycologia 2012, 104, 1420–1432. [Google Scholar] [CrossRef] [PubMed]
- Hosen, I.; Feng, B.; Wu, G.; Zhu, X.-T.; Li, Y.-C.; Yang, Z.L. Borofutus, a new genus of Boletaceae from tropical Asia: Phylogeny, morphology and taxonomy. Fungal Divers. 2013, 58, 215–226. [Google Scholar] [CrossRef]
- Zeng, N.K.; Chai, H.; Jiang, S.; Xue, R.; Wang, Y.; Hong, D.; Liang, Z.Q. Retiboletus nigrogriseus and Tengioboletus fujianensis, two new boletes from the south of China. Phytotaxa 2018, 367, 45–54. [Google Scholar] [CrossRef]
- Doyle, J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Huang, J.C.; Ge, X.J.; Sun, M. Modified CTAB Protocol Using a Silica Matrix for Isolation of Plant Genomic DNA. BioTechniques 2000, 28, 432–434. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W.; Innis, M.A.; Gelfand, D.H.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academic Press: Cambridge, MA, USA, 1990; Volume 31, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Gonzalez, D. Organization of ribosomal DNA in the basidiomycete Thanatephorus praticola. Curr. Genet. 1990, 18, 277–280. [Google Scholar] [CrossRef]
- James, T.Y.; Kauff, F.; Schoch, C.L.; Matheny, P.B.; Hofstetter, V.; Cox, C.J.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Joseph, F. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef]
- Gelardi, M.; Angelini, C.; Costanzo, F.; Ercole, E.; Ortiz-Santana, B.; Vizzini, A. Outstanding Pinkish Brown-Spored Neotropical Boletes: Austroboletus subflavidus and Fistulinella gloeocarpa (Boletaceae, Boletales) from the Dominican Republic. Mycobiology 2020, 49, 24–45. [Google Scholar] [CrossRef]
- Fechner, N.; Bonito, G.; Bougher, N.L.; Lebel, T.; Halling, R.E. New species of Austroboletus (Boletaceae) in Australia. Mycol. Prog. 2017, 16, 769–775. [Google Scholar] [CrossRef]
- Halling, R.E.; Osmundson, T.W.; Neves, M.A. Austroboletus mutabilis sp. nov. from northern Queensland. Muelleria 2006, 24, 31–36. [Google Scholar] [CrossRef]
- Simonini, G.; Floriani, M.; Binder, M.; Besl, H. Two close extraeuropean Boletes: Boletus violaceofuscus and Boletus separans. Micol. E Veg. Mediterr. 2001, 16, 148–170. [Google Scholar]
- Shi, X.F.; Liu, P.G. Aureoboletus zangii (Boletaceae), a new species from China. Mycotaxon 2013, 123, 451–456. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Locality | GenBank Accession No. | ||
|---|---|---|---|---|---|
| ITS | LSU | TEF1-α | |||
| Austroboletus | |||||
| Austroboletus aff. fusisporus | HKAS53461 | China | — | KF112486 | KF112214 |
| Austroboletus aff. rostrupii | G4357 | Guyana | — | KJ786636 | — |
| Austroboletus albus | HFJAU12001 | China | ON207028 | ON207254 | ON221310 |
| Austroboletus albus | HFJAU12002 | China | ON207029 | ON207255 | ON221311 |
| Austroboletus albidus | ShiSF238 | China | — | MT154756 | — |
| Austroboletus albovirescens | HKAS:59624 | China | — | KF112485 | KF112217 |
| Austroboletus albovirescens | HKAS74743 | China | — | KT990527 | KT990730 |
| Austroboletus amazonicus | 2032 AMV | Colombia | KF937309 | KF714510 | — |
| Austroboletus amazonicus | 1839 AMV | Colombia | KF937307 | KF714508 | — |
| Austroboletus appendiculatus | KCS 1401-CAL_1304 | India | KX530028 | — | — |
| Austroboletus asper | Perth 06658407 | Australia | KP242216 | KP242277 | — |
| Austroboletus asper | MEL:2104343 | Australia | KP242174 | KP242260 | — |
| Austroboletus asper | MEL:2300520 | Australia | KP242186 | KP242253 | — |
| Austroboletus austrovirens | BRI:AQ0796003 | Australia | KP242212 | KP242228 | — |
| Austroboletus austrovirens | BRI:AQ0795791 | Australia | KP242211 | KP242225 | — |
| Austroboletus brunneisquamus | FHMU5875 | China | MZ855494 | MW506828 | — |
| Austroboletus brunneisquamus | FHMU5876 | China | MZ855495 | MW506829 | MW512637 |
| Austroboletus dictyotus | HKAS59804 | China | — | JX901138 | — |
| Austroboletus dictyotus | HKAS53450 | China | — | KF112487 | KF112215 |
| Austroboletus festivus | FLOR 51599 | Brazil | KY886202 | KY888001 | — |
| Austroboletus festivus | FLOR 51601 | Brazil | KY886203 | KY888000 | — |
| Austroboletus festivus | AMV1881 | Colombia | KT724086 | KT724095 | — |
| Austroboletus fusisporus | JXSB0351 | China | — | MK765810 | — |
| Austroboletus fusisporus | HKAS75207 | China | JX889719 | JX889720 | JX889718 |
| Austroboletus gracilis | 112/96 | USA | — | DQ534624 | KF030425 |
| Austroboletus gracilis | NAMA 2017-106 | USA | MH979242 | — | — |
| Austroboletus gracilis | Mushroom Observer#310751 | Mexico | MH167935 | — | — |
| Austroboletus lacunosus | BRI:AQ0795789 | Australia | KP242162 | KP242271 | — |
| Austroboletus lacunosus | REH9146 | Australia | — | JX889669 | JX889709 |
| Austroboletus mucosus | TH6300 | Guyana | — | AY612798 | — |
| Austroboletus mutabilis | BRI:AQ0554121 | Australia | KP242192 | KP242241 | — |
| Austroboletus mutabilis | BRI:AQ0557644 | Australia | KP242196 | KP242237 | — |
| Austroboletus neotropicalis | NY181457 | Costa Rica | JQ924301 | JQ924334 | — |
| Austroboletus niveus | AD-C 54948 | Australia | KP242220 | KP242280 | — |
| Austroboletus niveus | Perth 6660703 | Australia | KP242217 | KP242279 | — |
| Austroboletus niveus | MEL2053830 | Australia | KC552016 | KC552058 | KC552099 |
| Austroboletus niveus | REH9487 | Australia | — | JX889668 | JX889708 |
| Austroboletus novae-zelandiae | PDD:105097 | New Zealand | — | — | MH594051 |
| Austroboletus novae-zelandiae | MEL:2370154 | Australia | KP242175 | KP242256 | — |
| Austroboletus occidentalis | MEL2300518 | Australia | KC552017 | KC552059 | KC552100 |
| Austroboletus olivaceobrunneus | HKAS92428 | China | — | — | MT110363 |
| Austroboletus olivaceoglutinosus | Xu132 | China | — | MT154753 | MW165263 |
| Austroboletus rarus | BRI:AQ0794045 | Australia | KP242197 | KP242236 | — |
| Austroboletus rarus | BRI:AQ0807888 | Australia | KP242200 | — | — |
| Austroboletus rionegrensis | INPA 78693 | Brazil | KY886201 | — | — |
| Austroboletus roseialbus | Dodd | Australia | KY872653 | KY872650 | — |
| Austroboletus roseialbus | REH10024 | Australia | KY872652 | KY872651 | — |
| Austroboletus rostrupii | BRI:AQ0807886 | Australia | KP242163 | KP242270 | — |
| Austroboletus rostrupii | BRI:AQ0796694 | Australia | KP242179 | KP242258 | — |
| Austroboletus rostrupii | TH8189 | Guyana | JN168683 | — | — |
| Austroboletus sp. | MEL:2382826 | Australia | KP242213 | KP242283 | — |
| Austroboletus sp. | BRI:AQ0794258 | Australia | KP242182 | KP242255 | — |
| Austroboletus sp. | BRI:AQ0794222 | Australia | KP242215 | KP242234 | — |
| Austroboletus sp. | MEL2305143 | New Caledonia | KC552018 | KC552060 | KC552101 |
| Austroboletus sp. | HKAS57756 | China | — | KF112383 | KF112212 |
| Austroboletus sp. | OR0891 | Thailand | — | — | MH614706 |
| Austroboletus sp. | LAM 0479 | Malaysia | — | KY091070 | — |
| Austroboletus sp. | DD9852 | North America | — | AY612797 | — |
| Austroboletus subflavidus | CFMR BZ-3178 BOS-625 | Belize | — | MK601716 | MK721070 |
| Austroboletus subflavidus | JBSD130772 | Dominican Republic | MT581526 | MT580903 | — |
| Austroboletus subflavidus | CFMR:DR2859 | Dominican Republic | MT581523 | MT580901 | — |
| Austroboletus subflavidus | CFMR:BOTH-3463 | USA | MT581521 | MT580900 | — |
| Austroboletus subvirens | KPM-NC-0017836 | Japan | — | JN378518 | JN378458 |
| Austroboletus subvirens | MEL:2382920 | Australia | KP012789 | — | — |
| Austroboletus viscidoviridis | Perth 7588682 | Australia | KP242219 | KP242282 | — |
| Austroboletus viscidoviridis | BRI:AQ0554020 | Australia | KP242189 | KP242243 | — |
| Austroboletus yourkae | BRI:AQ1024215 | Australia | — | MZ358814 | — |
| Austroboletus yourkae | NY02072669 | Australia | — | MZ358815 | — |
| Mucilopilus castaneiceps | HKAS50338 | China | — | KT990555 | KT990755 |
| Mucilopilus castaneiceps | HKAS71039 | China | — | KT990547 | KT990748 |
| Xanthoconium | |||||
| Xanthoconium affine | NY00815399 | USA | / | KT990661 | KT990850 |
| Xanthoconium affine | NY01193907 | USA | / | KT990660 | KT990849 |
| Xanthoconium affine | BD217 | USA | / | HQ161854 | — |
| Xanthoconium fusciceps | N.K.Zeng2941 | China | / | KY271035 | — |
| Xanthoconium fusciceps | N.K.Zeng2483 | China | / | KY271034 | KY271046 |
| Xanthoconium porophyllum | HKAS90217 | China | / | KT990662 | KT990851 |
| Xanthoconium porophyllum | GDGM 30303 | China | / | KC561775 | — |
| Xanthoconium purpureum | NY00720964 | USA | / | KT990663 | KT990852 |
| Xanthoconium purpureum | MICH:KUO-07061405 | USA | / | MK601816 | MK721170 |
| Xanthoconium separans | Mushroom Observer #282660 | USA | / | MH244206 | MH347319 |
| Xanthoconium separans | DPL 2704 | USA | / | KF030329 | KF030431 |
| Xanthoconium separans | MICH KUO-06201002 | USA | / | MK601723 | MK721077 |
| Xanthoconium sinense | N.K.Zeng1583 | China | / | KY271032 | KY271044 |
| Xanthoconium sinense | HKAS77758 | China | / | KT990665 | KT990854 |
| Xanthoconium sinense | N.K.Zeng1575 | China | / | KY271031 | KY271043 |
| Xanthoconium stramineum | 3518 | USA | / | KF030353 | KF030428 |
| Xanthoconium violaceipes | HFJAU12004 | China | / | OQ146964 | OQ162207 |
| Xanthoconium violaceipes | HFJAU12005 | China | / | OQ146965 | OQ162208 |
| Xanthoconium violaceipes | HFJAU12006 | China | / | OQ146966 | OQ162209 |
| Xanthoconium violaceofuscum | HFJAU12007 | China | / | OQ146967 | OQ162210 |
| Xanthoconium violaceofuscum | HFJAU12008 | China | / | OQ146968 | OQ162211 |
| Tylopilus argillaceus | HKAS90201 | China | / | KT990588 | KT990783 |
| Tylopilus plumbeoviolaceus | MB06-056 | USA | / | KF030350 | KF030439 |
| Xerocomus | |||||
| Xerocomus ferrugineus | MICH KUO-08100701 | USA | / | MK601820 | MK721174 |
| Xerocomus fraternus | HKAS69291 | China | / | KT990683 | KT990871 |
| Xerocomus fraternus | HKAS52526 | China | / | KT990682 | KT990870 |
| Xerocomus fulvipes | HKAS52556 | China | / | KT990672 | KT990860 |
| Xerocomus fulvipes | HKAS76666 | China | / | KF112390 | KF112292 |
| Xerocomus illudens | MB03-055 | USA | / | JQ003705 | — |
| Xerocomus illudens | DD9854 | USA | / | AY612840 | — |
| Xerocomus lentistipitatus | PDD 107806 | New Zealand | / | OP141603 | — |
| Xerocomus lentistipitatus | JAC14153 | New Zealand | / | OP141550 | — |
| Xerocomus lentistipitatus | JAC10917 | New Zealand | / | OP141509 | — |
| Xerocomus magniporus | HKAS:58000 | China | / | KF112392 | KF112293 |
| Xerocomus microcarpoides | HKAS54753 | China | / | KT990680 | KT990868 |
| Xerocomus microcarpoides | HKAS53374 | China | / | KT990679 | KT990867 |
| Xerocomus perplexus | MB00-005 | USA | / | JQ003702 | KF030438 |
| Xerocomus piceicola | HKAS76492 | China | / | KT990684 | KT990872 |
| Xerocomus piceicola | HKAS55452 | China | / | KT990685 | — |
| Xerocomus puniceiporus | HKAS80683 | China | / | KU974141 | KU974138 |
| Xerocomus rugosellus | HKAS58865 | China | / | KF112389 | KF112294 |
| Xerocomus rugosellus | HKAS68292 | China | / | KT990686 | KT990873 |
| Xerocomus rutilans | HFJAU12012 | China | / | OQ146972 | OQ162215 |
| Xerocomus rutilans | HFJAU12013 | China | / | OQ146973 | OQ162216 |
| Xerocomus squamulosus | JAC10883 | New Zealand | / | OP141507 | — |
| Xerocomus squamulosus | PDD95686 | New Zealand | / | JQ924327 | — |
| Xerocomus subparvus | HKAS53387 | China | / | KF112397 | KF112297 |
| Xerocomus subparvus | HKAS50295 | China | / | KT990667 | — |
| Xerocomus subparvus | HKAS55384 | China | / | KT990687 | KT990874 |
| Xerocomus subtomentosus | KM168813 | UK | / | KC215223 | KC215249 |
| Xerocomus subsplendidus | HFJAU12009 | China | / | OQ146969 | OQ162212 |
| Xerocomus subsplendidus | HFJAU12010 | China | / | OQ146970 | OQ162213 |
| Xerocomus subsplendidus | HFJAU12011 | China | / | OQ146971 | OQ162214 |
| Xerocomus subtomentosus | KM167686 | UK | / | KC215222 | KC215248 |
| Xerocomus subtomentosus | Xs1 | Germany | / | AF139716 | JQ327035 |
| Xerocomus velutinus | HKAS68135 | China | / | KT990673 | KT990861 |
| Xerocomus velutinus | HKAS52575 | China | / | KF112393 | KF112295 |
| Xerocomus sp. | HKAS67749 | China | / | KT990676 | KT990864 |
| Xerocomus sp. | HKAS76853 | China | / | KF112394 | KF112296 |
| Xerocomus sp. | HKAS75076 | China | / | KF112387 | KF112290 |
| Xerocomus sp. | HKAS57339 | China | / | KT990674 | KT990862 |
| Xerocomus sp. | HKAS57765 | China | / | KT990675 | KT990863 |
| Xerocomus sp. | HKAS74927 | China | / | KF112395 | KF112291 |
| Xerocomus sp. | HKAS90207 | China | / | KT990677 | KT990865 |
| Xerocomus yunnanensis | HKAS68420 | China | / | KT990690 | KT990877 |
| Xerocomus yunnanensis | HKAS68282 | China | / | KT990691 | KT990878 |
| Phylloporus bellus | HKAS56763 | China | / | JQ967196 | JQ967153 |
| Phylloporus leucomycelinus | HKAS74678 | USA | / | JQ967206 | JQ967163 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, F.; Gao, Y.; Song, H.-Y.; Hu, H.-J.; Yang, W.-J.; Zhang, W.; Liao, L.-Y.; Fang, Y.; Cheng, L.; Hu, D.-M. Phylogenetic and Morphological Evidence Reveal Five New Species of Boletes from Southern China. J. Fungi 2023, 9, 814. https://doi.org/10.3390/jof9080814
Zhou F, Gao Y, Song H-Y, Hu H-J, Yang W-J, Zhang W, Liao L-Y, Fang Y, Cheng L, Hu D-M. Phylogenetic and Morphological Evidence Reveal Five New Species of Boletes from Southern China. Journal of Fungi. 2023; 9(8):814. https://doi.org/10.3390/jof9080814
Chicago/Turabian StyleZhou, Fan, Yang Gao, Hai-Yan Song, Hai-Jing Hu, Wen-Juan Yang, Wei Zhang, Li-Yu Liao, Yi Fang, Lin Cheng, and Dian-Ming Hu. 2023. "Phylogenetic and Morphological Evidence Reveal Five New Species of Boletes from Southern China" Journal of Fungi 9, no. 8: 814. https://doi.org/10.3390/jof9080814