
Mutations in the Second Alternative Oxidase Gene: A New Approach to Group Aspergillus niger Strains
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mining of Alternative Oxidase and Type-II NADH:Ubiquinone Oxidoreductase Genes, Gene Synteny around Gene Loci and Confirmation of Expression
2.2. Maximum-Likelihood Phylogenetic Analysis
2.3. Aspergillus niger Strains Used for Sequence Verification of aoxB Alleles Reported in this Study
2.4. Isolation of A. niger Genomic DNA; Polymerase Chain Reaction (PCR), Vector Cloning and Sequence Determination
2.5. Isolation of A. niger Total RNA for cDNA Sequence Analysis
3. Results and Discussion
3.1. A Rare Alternative Oxidase Paralogous Gene (aoxB) Occurs in the Aspergillus niger Species Complex (Sensu Stricto) as well as in Distally Related Aspergillus calidoustus
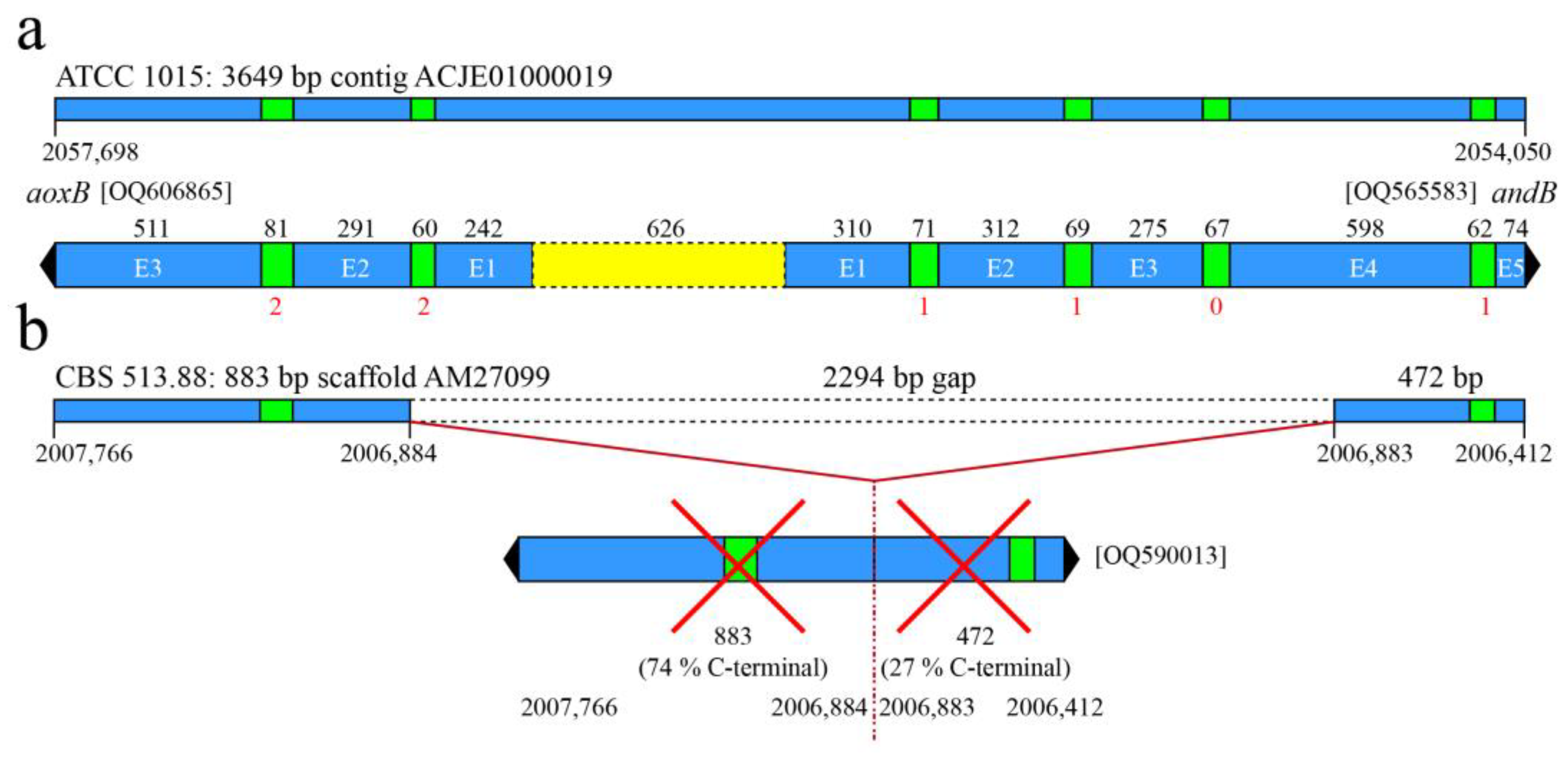
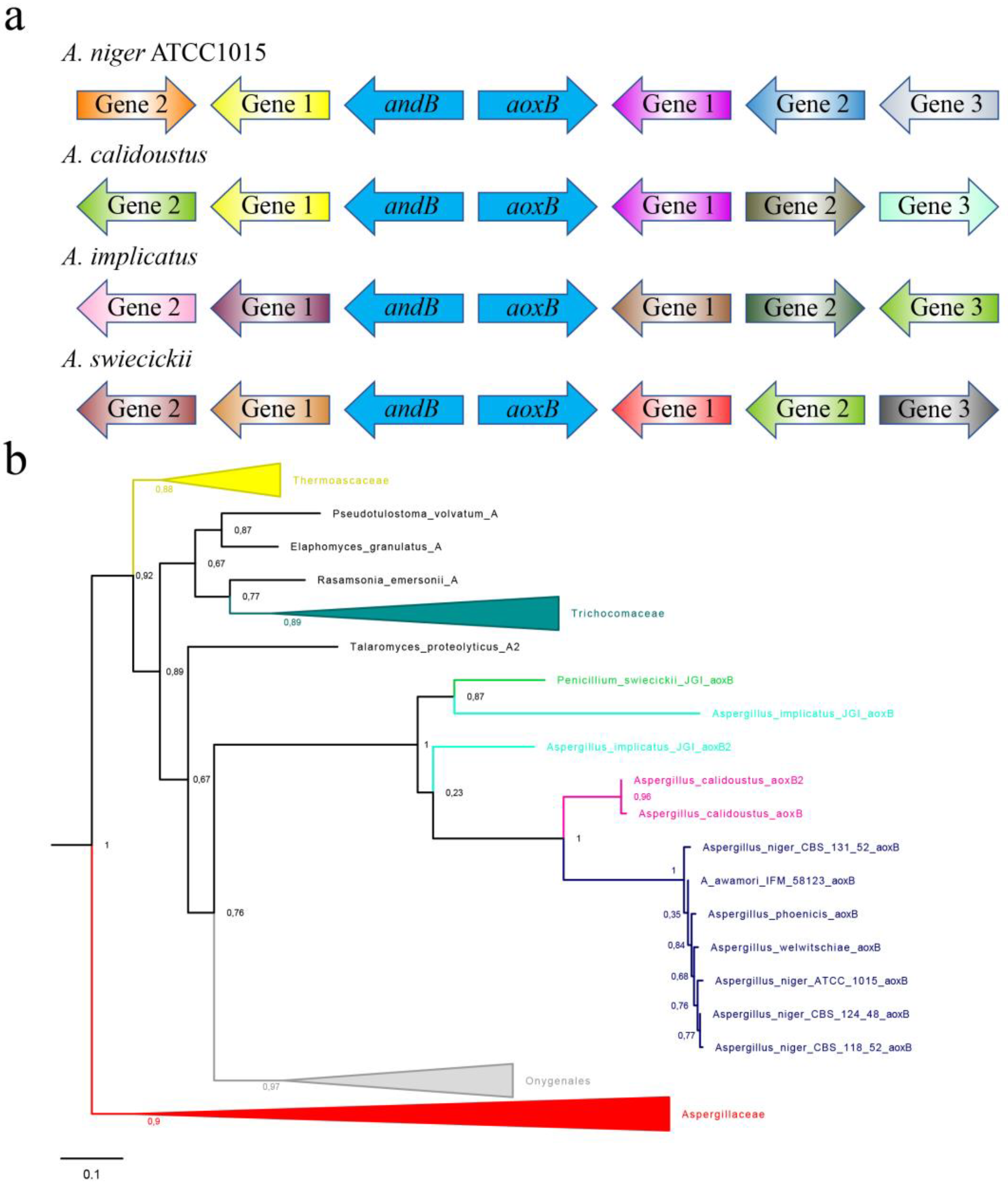
3.2. aoxB Is Divergently Orientated from a Rare Paralogous Gene Encoding a Type-II Alternative NADH:Ubiquinone Reductase
3.3. The aoxB and andB Gene Couple Occurs Sporadically in Four Dispersed Taxa of Aspergillaceae
3.4. The Rare aoxB Paralogue Appears to Originate from an Unidentified Host Taxon within the Eurotiomycetidae but Not an Aspergillaceae Taxon
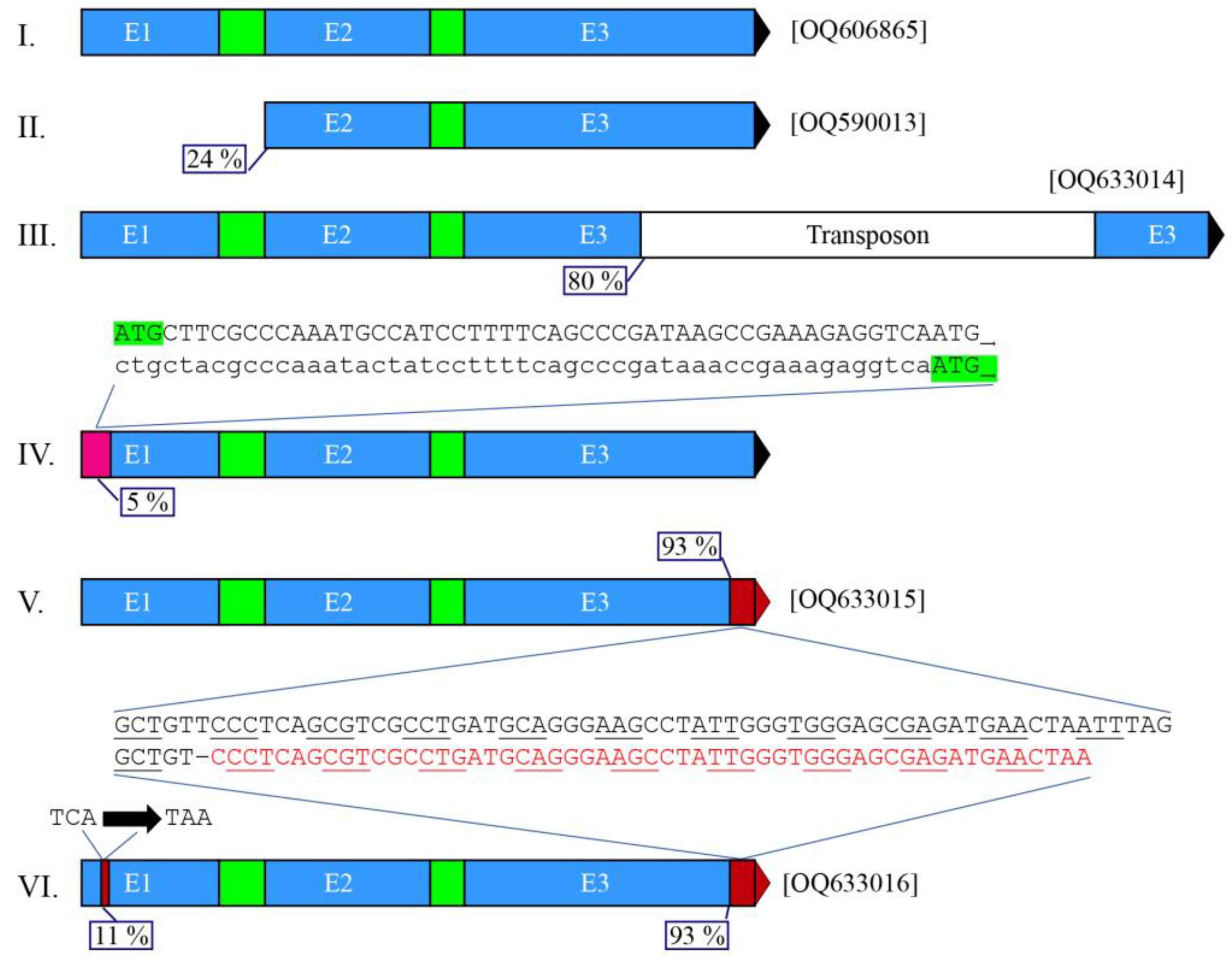
3.5. Multiple Alleles for the Acquired aoxB Paralogue in the Aspergillus niger Sensu Stricto Complex
3.6. aoxB Pseudogenes Occur in (Most) Aspergillus tubingensis but Not in Other Named Members of the Aspergillus luchuensis Clade
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmed, A.S.; Farag, S.S.; Hassan, I.A.; Botros, H.W. Production of gluconic acid by using some irradiated microorganisms. J. Radiat. Res. Appl. Sci. 2015, 8, 374–380. [Google Scholar] [CrossRef]
- Yoshioka, I.; Kobayashi, K.; Kirimura, K. Overexpression of the gene encoding alternative oxidase for enhanced glucose consumption in oxalic acid producing Aspergillus niger expressing oxaloacetate hydrolase gene. J. Biosci. Bioeng. 2020, 129, 172–176. [Google Scholar] [CrossRef]
- Xu, Y.; Shan, L.; Zhou, Y.; Xie, Z.; Ball, A.S.; Cao, W.; Liu, H. Development of a Cre-loxP-based genetic system in Aspergillus niger ATCC1015 and its application to construction of efficient organic acid-producing cell factories. Appl. Microbiol. Biotechnol. 2019, 103, 8105–8114. [Google Scholar] [CrossRef]
- Andersen, M.R.; Vongsangnak, W.; Panagiotou, G.; Salazar, M.P.; Lehmann, L.; Nielsen, J. A trispecies Aspergillus microarray: Comparative transcriptomics of three Aspergillus species. Proc. Natl. Acad. Sci. USA 2008, 105, 4387–4392. [Google Scholar] [CrossRef]
- Culleton, H.; McKie, V.; de Vries, R.P. Physiological and molecular aspects of degradation of plant polysaccharides by fungi: What have we learned from Aspergillus? Biotechnol. J. 2013, 8, 884–894. [Google Scholar] [CrossRef]
- Balajee, S.A.; Kano, R.; Baddley, J.W.; Moser, S.A.; Marr, K.A.; Alexander, B.D.; Andes, D.; Kontoyiannis, D.P.; Perrone, G.; Peterson, S.; et al. Molecular identification of Aspergillus species collected for the Transplant-Associated Infection Surveillance Network. J. Clin. Microbiol. 2009, 47, 3138–3141. [Google Scholar] [CrossRef]
- Taccone, F.S.; Van den Abeele, A.M.; Bulpa, P.; Misset, B.; Meersseman, W.; Cardoso, T.; Paiva, J.-A.; Blasco-Navalpotro, M.; De Laere, E.; Dimopoulos, G.; et al. Epidemiology of invasive aspergillosis in critically ill patients: Clinical presentation, underlying conditions, and outcomes. Crit. Care 2015, 19, 7. [Google Scholar] [CrossRef]
- D’hooge, E.; Becker, P.; Stubbe, D.; Normand, A.C.; Piarroux, R.; Hendrickx, M. Black aspergilli: A remaining challenge in fungal taxonomy? Med. Mycol. 2019, 57, 773–780. [Google Scholar] [CrossRef]
- da Silva, J.J.; Iamanaka, B.T.; Ferranti, L.S.; Massi, F.P.; Taniwaki, M.H.; Puel, O.; Lorber, S.; Frisvad, J.C.; Fungaro, M.H.P. Diversity within Aspergillus niger clade and description of a new species: Aspergillus vinaceus sp. nov. J. Fungi 2020, 6, 371. [Google Scholar] [CrossRef]
- Gits-Muselli, M.; Hamane, S.; Verillaud, B.; Cherpin, E.; Denis, B.; Bondeelle, L.; Touratier, S.; Alanio, A.; Garcia-Hermoso, D.; Bretagne, S. Different repartition of the cryptic species of black aspergilli according to the anatomical sites in human infections, in a French University hospital. Med. Mycol. 2021, 59, 985–992. [Google Scholar] [CrossRef]
- McDonald, A.E.; Vanlerberghe, G.C.; Staples, J.F. Alternative oxidase in animals: Unique characteristics and taxonomic distribution. J. Exp. Biol. 2009, 212, 2627–2634. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Lee, S.Y.; Woo, S.Y.; Chun, H.S. Alternative oxidase: A potential target for controlling aflatoxin contamination and propagation of Aspergillus flavus. Front. Microbiol. 2020, 11, 419. [Google Scholar] [CrossRef]
- Del-Saz, N.F.; Ribas-Carbo, M.; McDonald, A.E.; Lambers, H.; Fernie, A.R.; Florez-Sarasa, I. An in vivo perspective of the role(s) of the alternative oxidase pathway. Trends Plant Sci. 2018, 23, 206–219. [Google Scholar] [CrossRef]
- Karaffa, L.; Sándor, E.; Kozma, J.; Szentirmai, A. Cephalosporin-C production, morphology and alternative respiration of Acremonium chrysogenum in glucose-limited chemostat. Biotechnol. Lett. 1996, 18, 701–706. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Zehentgruber, O.; El-Kalak, H.; Röhr, M. Regulation of citric acid production by oxygen: Effect of dissolved oxygen tension on adenylate levels and respiration in Aspergillus niger. Eur. J. Appl. Microbiol. Biotechnol. 1980, 9, 101–115. [Google Scholar] [CrossRef]
- Karaffa, L.; Kubicek, C.P. Production of Organic Acids by Fungi. In Encyclopedia of Mycology, 1st ed.; Zaragoza, O., Casadevall, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 2, pp. 406–419. [Google Scholar]
- Molnár, Á.P.; Németh, Z.; Kolláth, I.S.; Fekete, E.; Flipphi, M.; Ág, N.; Soós, Á.; Kovács, B.; Sándor, E.; Kubicek, C.P.; et al. High oxygen tension increases itaconic acid accumulation, glucose consumption, and the expression and activity of alternative oxidase in Aspergillus terreus. Appl. Microbiol. Biotechnol. 2018, 102, 8799–8808. [Google Scholar] [CrossRef]
- Kozma, J.; Karaffa, L. Effect of oxygen on the respiratory system and cephalosporin-C production in Acremonium chrysogenum. J. Biotechnol. 1996, 48, 59–66. [Google Scholar] [CrossRef]
- Molnár, Á.P.; Németh, Z.; Fekete, E.; Flipphi, M.; Keller, N.P.; Karaffa, L. Analysis of the relationship between alternative respiration and sterigmatocystin formation in Aspergillus nidulans. Toxins 2018, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Huh, W.K.; Kang, S.O. Molecular cloning and functional expression of alternative oxidase from Candida albicans. J. Bacteriol. 1999, 181, 4098–4102. [Google Scholar] [CrossRef] [PubMed]
- Huh, W.K.; Kang, S.O. Characterization of the gene family encoding alternative oxidase from Candida albicans. Biochem. J. 2001, 356, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ritzel, R.G.; McLean, L.L.T.; McIntosh, L.; Ko, T.; Bertrand, H.; Nargang, F.E. Cloning and analysis of the alternative oxidase gene of Neurospora crassa. Genetics 1996, 142, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Tanton, L.L.; Nargang, C.E.; Kessler, K.E.; Li, Q.; Nargang, F.E. Alternative oxidase expression in Neurospora crassa. Fungal Genet. Biol. 2003, 39, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, K.; Yoda, M.; Usami, S. Cloning and expression of the cDNA encoding an alternative oxidase gene from Aspergillus niger WU-2223L. Curr. Genet. 1999, 34, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, K.; Ogawa, S.; Hattori, T.; Kino, K. Expression analysis of alternative oxidase gene (aox1) with enhanced green fluorescent protein as marker in citric acid-producing Aspergillus niger. J. Biosci. Bioeng. 2006, 102, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Pel, H.J.; de Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; de Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.R.; Salazar, M.P.; Schaap, P.J.; van de Vondervoort, P.J.I.; Culley, D.; Thykaer, J.; Frisvad, J.C.; Nielsen, K.F.; Albang, R.; Albermann, K.; et al. Comparative genomics of citric-acid-producing Aspergillus niger ATCC 1015 versus enzyme-producing CBS 513.88. Genome Res. 2011, 21, 885–897. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Seekles, S.J.; Punt, M.; Savelkoel, N.; Houbraken, J.; Wösten, H.A.B.; Ohm, R.A.; Ram, A.F.J. Genome sequences of 24 Aspergillus niger sensu stricto strains to study strain diversity, heterokaryon compatibility, and sexual reproduction. G3-Genes Genomes Genet. 2022, 12, jkac124. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Chorev, M.; Carmel, L. The function of introns. Front. Genet. 2012, 3, 55. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Dang, C.C.; Vinh, L.S.; Lanfear, R. QMaker: Fast and accurate method to estimate empirical models of protein evolution. Syst. Biol. 2021, 70, 1046–1060. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Horn, F.; Linde, J.; Mattern, D.J.; Walther, G.; Guthke, R.; Scherlach, K.; Martin, K.; Brakhage, A.A.; Petzke, L.; Valiante, V. Draft genome sequences of fungus Aspergillus calidoustus. Genome Announc. 2016, 4, e00102-16. [Google Scholar] [CrossRef]
- Marreiros, B.C.; Sena, F.V.; Sousa, F.M.; Oliveira, A.S.F.; Soares, C.M.; Batista, A.P.; Pereira, M.M. Structural and functional insights into the catalytic mechanism of the Type II NADH:quinone oxidoreductase family. Sci. Rep. 2017, 7, 42303. [Google Scholar] [CrossRef]
- Dinamarco, T.M.; Figueiredo Pimentel, B.d.C.; Savoldi, M.; Malavazi, I.; Soriani, F.M.; Uyemura, S.A.; Ludovico, P.; Goldman, M.H.S.; Goldman, G.H. The roles played by Aspergillus nidulans apoptosis-inducing factor (AIF)-like mitochondrial oxidoreductase (AifA) and NADH-ubiquinone oxidoreductases (NdeA-B and NdiA) in farnesol resistance. Fungal Genet. Biol. 2010, 47, 1055–1069. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, P.; Duarte, M.; Videira, A. The main external alternative NAD(P)H dehydrogenase of Neurospora crassa mitochondria. Biochim. Biophys. Acta Bioenerg. 2004, 1608, 45–52. [Google Scholar] [CrossRef]
- Ellena, V.; Seekles, S.J.; Vignolle, G.A.; Ram, A.F.J.; Steiger, M.G. Genome sequencing of the neotype strain CBS 554.65 reveals the MAT1–2 locus of Aspergillus niger. BMC Genom. 2021, 22, 679. [Google Scholar] [CrossRef]
- Duarte, E.A.A.; Damasceno, C.L.; de Oliveira, T.A.S.; Barbosa, L.d.O.; Martins, F.M.; de Silva, J.R.Q.; de Lima, T.E.F.; da Silva, R.M.; Kato, R.B.; Bortolini, D.E.; et al. Putting the mess in order: Aspergillus welwitschiae (and not A. niger) is the etiological agent of sisal bole rot disease in Brazil. Front. Microbiol. 2018, 9, 1227. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Kusuya, Y.; Alimu, Y.; Bian, C.; Takahashi, H.; Yaguchi, T. Draft Genome sequence of Aspergillus awamori IFM 58123NT. Microbiol. Resour. Announc. 2019, 8, e01453-18. [Google Scholar] [CrossRef] [PubMed]
- Vesth, T.C.; Nybo, J.L.; Theobald, S.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F.; Hoof, J.B.; Brandl, J.; Salamov, A.; Riley, R.; et al. Investigation of inter- and intraspecies variation through genome sequencing of Aspergillus section Nigri. Nat. Genet. 2018, 50, 1688–1695. [Google Scholar] [CrossRef]
- Daboussi, M.-J.; Capy, P. Transposable elements in filamentous fungi. Annu. Rev. Microbiol. 2003, 57, 275–299. [Google Scholar] [CrossRef]
- Braumann, I.; van den Berg, M.; Kempken, F. Transposons in biotechnologically relevant strains of Aspergillus niger and Penicillium chrysogenum. Fungal Genet. Biol. 2007, 44, 1399–1414. [Google Scholar] [CrossRef]
- Hong, S.-B.; Lee, M.; Kim, D.-H.; Varga, J.; Frisvad, J.C.; Perrone, G.; Gomi, K.; Yamada, O.; Machida, M.; Houbraken, J.; et al. Aspergillus luchuensis, an industrially important black Aspergillus in East Asia. PLoS ONE 2013, 8, e63769. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, I.; Takahashi, H.; Kusuya, Y.; Yaguchi, T.; Kirimura, K. Draft genome sequence of Aspergillus tubingensis WU-2223L, a citric acid-producing filamentous fungus belonging to Aspergillus section Nigri. Microbiol. Resour. Announc. 2020, 9, e00702-20. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Cheng, Z.; Zhang, H.; Liu, L.; Gao, P.; Wang, L. Draft genome sequence of Aspergillus niger strain An76. Genome Announc. 2016, 4, e01700-15. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Valerio, M.; Álvarez-Uría, A.; Olmedo, M.; Veintimilla, C.; Padilla, B.; De la Villa, S.; Guinea, J.; Escribano, P.; Ruiz-Serrano, M.J.; et al. Invasive pulmonary aspergillosis in the COVID-19 era: An expected new entity. Mycoses 2021, 64, 132–143. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Aspergillus niger Strain | Genome Master Accession | Source | Gene Allele | GenBank Accession Number(s) |
|---|---|---|---|---|
| ATCC 1015 | ACJE01 | Our laboratory | Wild type aoxB (Group I) | OQ606865 OQ686795 |
| ATCC 1015 | ACJE01 | Our laboratory | Wild type andB * | OQ565583 * OQ689783 * |
| CBS 554.65 | JAGRPH01 | Matthias Steiger | Deletion part of aoxB and andB (Group II) | OQ590013 |
| CBS 147343 | JAKJMC01 | Arthur Ram | Transposon insertion (Group III) | OQ633014 |
| CBS 630.78 | JAKJLX01 | Arthur Ram | Frameshift mutation (Group V) | OQ633015 |
| CBS 769.97 | JAKJMK01 | Arthur Ram | Frameshift mutation plus nonsense mutation (Group VI) | OQ633016 |
| Group I | Group II | Group III | Group IV | Group V | Group VI | |
|---|---|---|---|---|---|---|
| Wild Type Allele aoxB | Allele 2 aoxB [Partial Deletion] | Allele 3 aoxB [Transposon Insertion] | Allele 4 aoxB [Missense Mutation Met1Leu] | Allele 5 aoxB [Frameshift near 3′ of CDS] | Allele 6 aoxB [Frameshift Plus Nonsense Mutation] | aoxB Pseudogene in A. tubingensis [A. luchuensis Clade] |
| IFM 58123 A. awamori | ATCC 13496 | CBS 113.50 | ATCC 13157 A. phoenicis | CBS 630.78 | CBS 131.52 | “A. niger” strain An76 |
| ATCC 1015 | CBS 101883 A. lacticoffeatus | CBS 118.52 | JSC-093350089 | CBS 147347 | CBS 769.97 | “A. niger” strain 3.316 |
| ATCC 64974 [N402] | CBS 112.32 | CBS 124.48 | strain F1702 | A. tubingensis WU-2223L | ||
| CBS 147345 | CBS 115988 | CBS 147322 | strain M3604 | A. costaricensis FKII-L6-BK-DRAB1 | ||
| CBS 147346 | CBS 115989 | CBS 147323 | strain R1650 | A. tubingensis S/N-304-OC-P1 | ||
| CBS 147482 | CBS 147321 | CBS 147324 | strain R20-06 | A. tubingensis S/N-308-IC-B1 | ||
| CBS_139.54b A. welwitschiae | CBS 513.88 | CBS 147343 | strain RG13B1 | A. tubingensis VS III B KN t | ||
| CCMB 663 A. welwitschiae | CBS 554.65 A. niger neotype | CBS 147344 | strain S1603 | A. tubingensis S/N-302-OC-P2 | ||
| CCMB 674 A. welwitschiae | strain A1 | CBS 147371 | strain Y1650 | A. tubingensis strain JS3-R1 | ||
| DSM 1957 | strain H915-1 | strain F3_1F3_F | A. tubingensis strain JS3-P2 | |||
| FDAARGOS_311 | strain L2 | ATCC 10864 (2 contigs) # | A. tubingensis strain C2-2 | |||
| FGSC A1279 | strain LDM3 | CBS 133816 (2 contigs) # | strain Y4002A | |||
| IHEM 2864 A. welwitschiae | strain SH-2 | CBS 147320 (2 contigs) # | strain S3103 | |||
| ITEM 11945 (A. welwitschiae) | CBS 147352 (2 contigs) # | strain BSC-1 | ||||
| NRRL 3 [CBS 120.49; N400] | CBS 147353 (2 contigs) # | strain S1133 | ||||
| “Penicillium fimorum” S/N-308-OC-P1 * | strain MOD1-FUNGI2 (2 contigs) # | strain F8013-2 | ||||
| strain F3_4F1_F | strain S1118 | |||||
| strain F3_4F2_F | strain P1003-2 | |||||
| strain L14 | strain PG3607 | |||||
| strain P1402 | strain B7004A | |||||
| strain S1 | ||||||
| strain Y2001A1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flipphi, M.; Márton, A.; Bíró, V.; Ág, N.; Sándor, E.; Fekete, E.; Karaffa, L. Mutations in the Second Alternative Oxidase Gene: A New Approach to Group Aspergillus niger Strains. J. Fungi 2023, 9, 570. https://doi.org/10.3390/jof9050570
Flipphi M, Márton A, Bíró V, Ág N, Sándor E, Fekete E, Karaffa L. Mutations in the Second Alternative Oxidase Gene: A New Approach to Group Aspergillus niger Strains. Journal of Fungi. 2023; 9(5):570. https://doi.org/10.3390/jof9050570
Chicago/Turabian StyleFlipphi, Michel, Alexandra Márton, Vivien Bíró, Norbert Ág, Erzsébet Sándor, Erzsébet Fekete, and Levente Karaffa. 2023. "Mutations in the Second Alternative Oxidase Gene: A New Approach to Group Aspergillus niger Strains" Journal of Fungi 9, no. 5: 570. https://doi.org/10.3390/jof9050570