
Calcium Ion Channels in Saccharomyces cerevisiae
College of Life and Health, Dalian University, Dalian 116622, China
J. Fungi 2023, 9(5), 524; https://doi.org/10.3390/jof9050524
Submission received: 29 March 2023
/
Revised: 25 April 2023
/
Accepted: 25 April 2023
/
Published: 28 April 2023
(This article belongs to the Section Fungi in Agriculture and Biotechnology)
Abstract
:Regulating calcium ion (Ca2+) channels to improve the cell cycle and metabolism is a promising technology, ensuring increased cell growth, differentiation, and/or productivity. In this regard, the composition and structure of Ca2+ channels play a vital role in controlling the gating states. In this review, Saccharomyces cerevisiae, as a model eukaryotic organism and an essential industrial microorganism, was used to discuss the effect of its type, composition, structure, and gating mechanism on the activity of Ca2+ channels. Furthermore, the advances in the application of Ca2+ channels in pharmacology, tissue engineering, and biochemical engineering are summarized, with a special focus on exploring the receptor site of Ca2+ channels for new drug design strategies and different therapeutic uses, targeting Ca2+ channels to produce functional replacement tissues, creating favorable conditions for tissue regeneration, and regulating Ca2+ channels to enhance biotransformation efficiency.
1. Introduction
As cell membrane proteins, calcium ion (Ca2+) channels carry out some key functions, including Ca2+ signal transduction, cell cycle, transport motility, and gene expression [1,2]. Different stimuli or stresses can induce the opening or closing of channel proteins distributed in the plasma membrane and various organelles. Additionally, any disruption in Ca2+ homeostasis can lead to alterations of certain essential functions in the cells.
As a model unicellular eukaryote, the budding yeast Saccharomyces cerevisiae is one of the most studied organisms worldwide, with its genome being thoroughly mapped both genetically and physically [3]. Moreover, S. cerevisiae provides a myriad of compatible plasmids and phages for genetic engineering. Notably, S. cerevisiae is the key microorganism in the industrial production of bioethanol, a clean, renewable, and sustainable alternative fuel [4]. As such, S. cerevisiae is widely used in theoretical research and production practice.
Ca2+ channels in S. cerevisiae act as the structural homologue and/or a functional analogue of the part of the Ca2+ channels in human or animal cells [3]. The deletion of genes encoding these channel proteins leads to an observable phenotype in yeast. Thus, S. cerevisiae is often used to introduce mutations in homologous genes associated with diseases or metabolic alterations in humans. Furthermore, various techniques for isolating and measuring channel activity, such as patch clamp, are well established in yeast. In this context, many studies have been conducted to understand the structure–function relationship of yeast proteins in disease or altered metabolism states. In this review, the development of novel regulation strategies for Ca2+ channels in S. cerevisiae is summarized comprehensively.
2. Types and Characteristics of Ca2+ Channels in S. cerevisiae
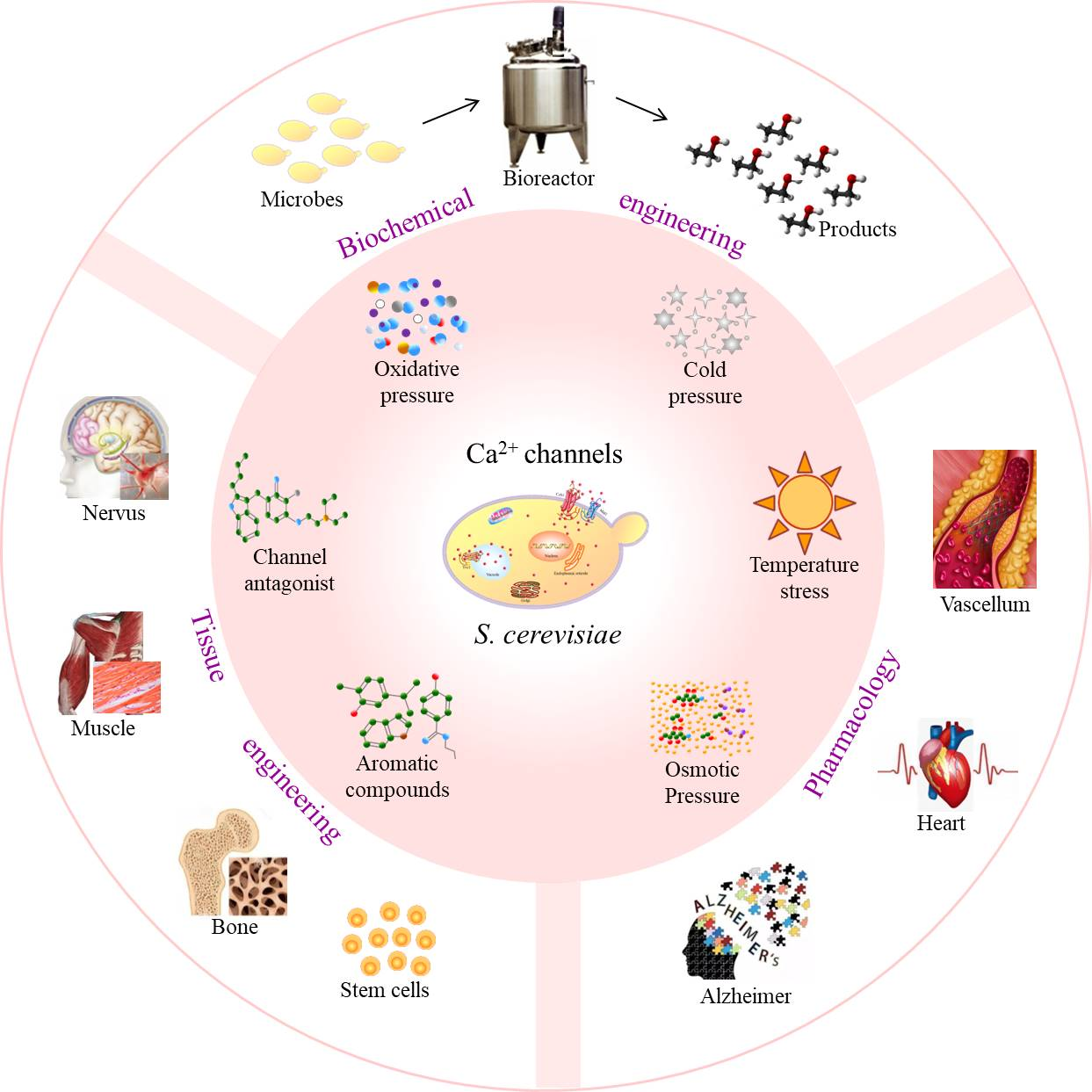
Based on their activation mechanisms, Ca2+ channels in S. cerevisiae are classified into three types: (i) Cch1 of the voltage-gated calcium channel (VGCC); (ii) Mid1 of the stretch-activated calcium channel (SACC); and (iii) Yvc1 of the transient receptor potential (TRP) channel [5,6,7] (Figure 1A). Cch1 and Mid1 are located in the cell membrane, whereas Yvc1 is located in the vacuole membrane. Ca2+-permeable channels regulate the passive flow of Ca2+ across the plasma membrane into the cytoplasm.
2.1. VGCC
In S. cerevisiae, the gene CCH1 encodes the Cch1 protein, comprising 2039 amino acid residues with a molecular weight of 234.6 kDa, which has been predicted as a multiple membrane-spanning transmembrane protein (Figure 1B). Cch1 contains four structurally similar hydrophobic domains (I-IV), with each domain having six transmembrane domain (TMD) segments [8]. All four domains contain the amino acid residue indicative of the Ca2+-selective P segment (channel specificity), whereas three of the four domains (i.e., II, III, and IV) contain a highly conserved glutamate residue, which plays a critical role in Ca2+ coordination. Each S4 segment of domains I, II, and III contains repeated motifs of a positively charged amino acid residue, followed by two hydrophobic amino acid residues (voltage dependence).
Cch1 is a homolog of the pore-forming subunit α1 of the dihydropyridine-sensitive (L-type) family of mammalian VGCCs [9], retaining several key features, including the size, topology, and domain structure of α1 subunits. Cch1 contains a total of 28 cysteine residues, of which four are found in an internal loop connecting TMD4 and TMD5 of segment I, and two at each pore region of segment III and segment IV are highly conserved across species, from yeast to humans [10].
The overall amino acid sequence similarity between Cch1 and α1 subunits of L-type VGCC in mammal cells is low, i.e., 24% for Cch1 compared to a mammalian L-type VGCC. Generally, the mammalian VGCCs contain 19 lysine and arginine residues located in the voltage sensing S4 domains, whereas Cch1 and its homologs in fungi contain 11 to 12 lysine and arginine residues at these corresponding sites [9,11], which can potentially alter the membrane voltage by changing the response speed and sensitivity. Additionally, in mammalian voltage-gated calcium (Cav) channels, a Ca2+-binding site contributing to the ionic selectivity is formed by a quartet of pore-localized glutamate residues (EEEE locus), one from the selectivity filter region of each of the four domains [12]. Moreover, each of the Cch1 channel homologues identified in the fungi possesses a similarly placed acidic motif but with three acidic residues rather than four [13].
2.2. SACC
The gene MID1 encodes Mid1, a transmembrane polypeptide comprising 548 amino acid residues with a molecular weight of 61.5 kDa. As the major and independent Ca2+ entry route, Mid1 is mostly involved in Ca2+ uptake at a low Ca2+ concentration of 100 μM and mating. The Mid1 channel has four hydrophobic regions, corresponding to the amino acid residues of 2–22, 92–111, 337–356, and 366–388, namely, H1, H2, H3, and H4, respectively (Figure 1C).
Mid1 shares no extensive similarity with any known proteins. The H1 segment is a signal sequence, whereas the H2 and H4 segments are partially similar to the transmembrane segments of known ion channels, and the H3 segment is similar to that of the pore-forming regions of several cation channels [6]. In the H2 segment, the glycine residue plays a vital role in channel gating [14]. According to the protease protection experiments on intact cells, the region between the H3 and H4 segments is often located outside the plasma membrane, suggesting that the Phe356 residue would be located outside the plasma membrane. The hydrophobicity of the Phe356 residue is essential for viability maintenance and Ca2+ uptake, while its size significantly affects viability maintenance [15].
Mid1 is a plasma and endoplasmic reticulum (ER) membrane protein with 16 N-glycosylation sites and a C-terminal cysteine (Cys)-rich region comprising 12 Cys residues [16]. The Cys-rich region is located in the domain between the H4 segment and the Thr526 residue, with at least three motifs, including a putative casein kinase 2 phosphorylation motif, a sheet–turn–sheet structure, and an EF-hand motif of helix–loop–helix structure [14]. The sheet–turn–sheet structure significantly contributes to the complementing ability and Ca2+ uptake activity, whereas the EF-hand motif contributes to the sites of interaction with other proteins or Mid1 itself.
In ER, Mid1 is located as a 200-kDa oligomer by covalent cysteine bonding, probably through the Cys-rich region. There is evidence that Mid1 is N-glycosylated in the ER, and then transported to the plasma membrane. The hypothesis is supported by the presence of a putative N-terminal signal peptide and several potential transmembrane (TM) α-helices [17].
2.3. TRPY1
The gene YVC1 encodes S. cerevisiae vacuolar transient receptor potential yeast1 (TRPY1, also known as yeast vacuolar conductance 1, or Yvc1). Yvc1 comprises 675 amino acid residues with a molecular weight of 78 kDa. As an ion channel responsible for the efflux of vacuolar Ca2+ to the cytoplasm, Yvc1 is the only member of the TRP superfamily expressed in S. cerevisiae [18]. As a low-copy-number protein, the number of Yvc1 molecules per cell during the exponential growth phase is approximately 1300 [19]. Additionally, the whole vacuole currents, combined with a single-channel recording, show at least 100 channels per vacuole [5].
Similar to all TRP members, Yvc1 contains six predicted TMDs with cytosolic N- and C-termini and a hydrophobic pore region located between TMD5 and TMD6. TMD6 forms the part of the ion conduction pathway and takes part in the deactivation of cation channel gating [5] (Figure 1D). The short amino acid sequence motif, VILLNILIALY, between the residues 448–458 in TMD6 in S. cerevisiae, is highly conserved in the corresponding TMD6 sequence of most mammalian TRPs. Additionally, the C-terminus of Yvc1 contains a cluster of four acidic residues involved in Ca2+-dependent regulation, which is analogous to the Ca2+-binding bowl in the big-conductance Ca2+-activated K+ (BK) channels [20]. Yvc1 contains nine cysteine residues, eight of which are cytoplasmic, and only Cys-343 is located within TMD2. Among these cysteine residues, only Cys-191 is conserved across yeast and fungi, and three cysteine residues of Cys-17, Cys-79, and Cys-191 are specifically glutathionylated. It is reported that glutathione depletion in yeast leads to the activation of Yvc1 [21]. Recently, the full-length cryoelectron microscopy structure of Yvc1 has been elucidated, which is consistent with most previously studied TRP channel proteins but has distinct structural folds in the cytosolic N and C termini [22].
3. Gating Mechanisms of Ca2+ Channels
The open–close status of Ca2+ channels is key to achieving the structure–function relationship. Some physiological activities of cells, such as the generation of pacemaker potential, the contraction of the muscle cells, the plateau phase of the cardiac action potential, and gene expression, are mainly mediated by fine-tuning Ca2+ entry through Ca2+ channels [23]. Moreover, the opening (activation) and closing (inactivation) of various types of channels depend on different gating mechanisms.
3.1. Voltage-Dependent Gating
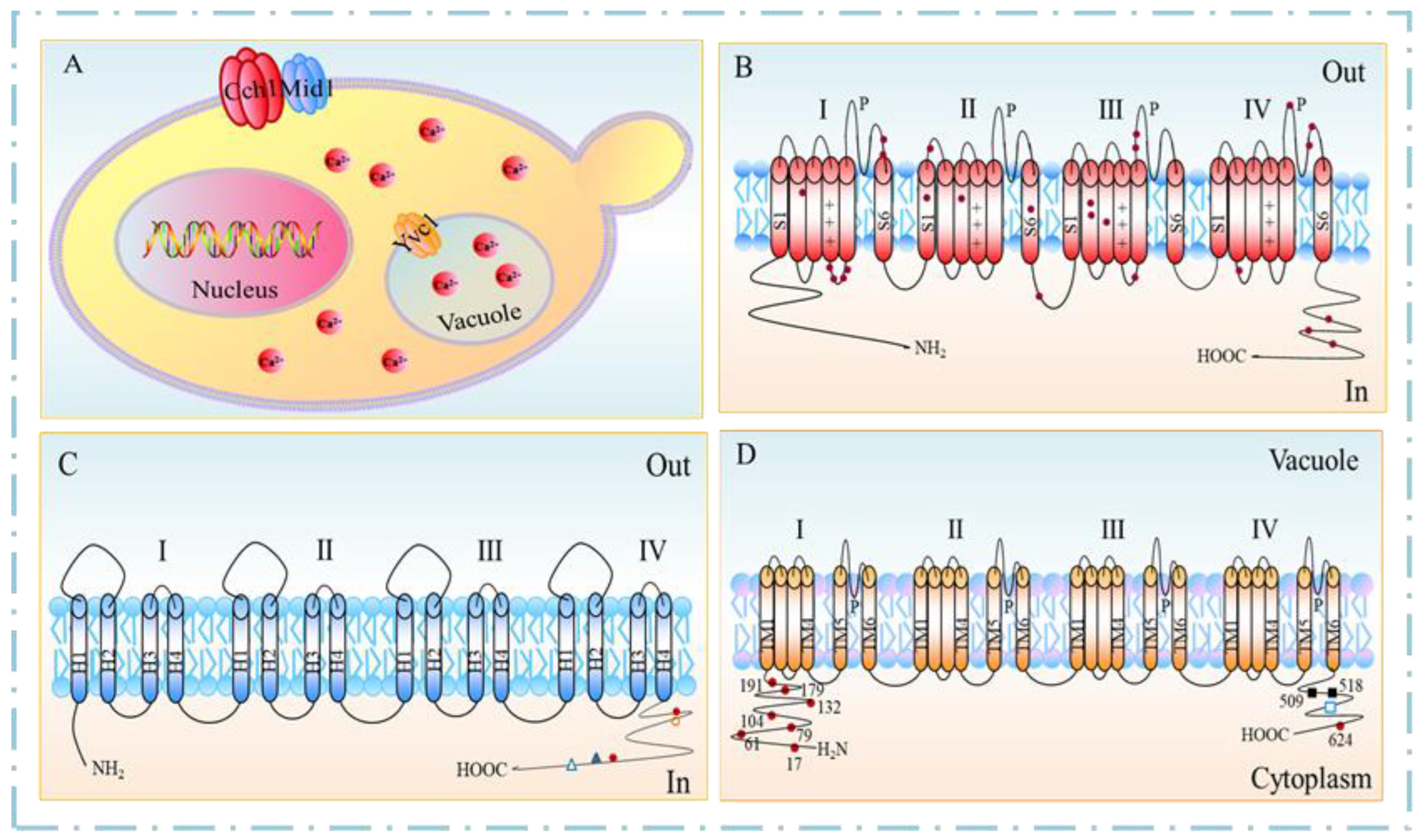
It is reported that the gating mechanism of Cch1 is consistent with the α-subunit of L-type VGCC in mammalian cells in terms of sequence similarity [24]. The atomic model of the α1-subunit comprises the pore domain, the four voltage-sensing domains (VSDs), the intracellular α1-interacting domains (AID)-containing helix between the I and II domains (the I-II helix), and the intact III-IV linker (Figure 2A). The selectivity filter (SF) in the pore domain located at the extracellular side of TMD contains the EEEE locus (Glu292/614/1014/1323) [25,26] (Figure 2B). The selectivity filter extends to the channel cavity surrounded by a tetrameric arrangement of segments S5 and S6 (Figure 2C). In all four VSDs, the gating charges are aligned on one side of the 310 helices of S4 segments. Moreover, the R5 residues and R6II are underneath, whereas the R1-R4 residues are above the conserved occluding Phe residue in the charge transfer center, representing the depolarized or “up” conformation of VSDs (Figure 2D). In contrast, the negative residue on S3 responsible for the charge transfer center is located in the VSDII and VSDIV, but not in the other two VSDs. Collectively, these may indicate a potentially inactivated state of Cav1.1.
VSDs and the pore domain are mainly lined by the S4–S5 linkers, which directly interact with all four S6 segments through the residues G432 (IS6), A780 (IIS6), G1193 (IIIS6), and A1503 (IVS6), designated as the G/A/G/A ring. Thus, mutations in the G/A/G/A residues affect the VSDs movement [27], in particular, the S6 helices form the activation gate at the intracellular end of the TMD. In the closed state, the pore-linking S6 helices converge at the intracellular flank to obstruct ion permeation. Upon opening, the intracellular ends of the S6 helices diverge from one another, thereby creating a wide opening to enable ion passage [28].
It is reported that all eight conserved extracellular Cys residues are associated with the Cch1 activity. However, replacing the Asn1066 residue with a negatively charged amino acid might change the conformation of the cytoplasmic II-III interdomain loop, thereby turning Cch1 into a closed structure prone to opening even in the absence of stimuli [8]. Conversely, replacing Gly1265 in the S2–S3 linker of domain III perturbs the spatial arrangement between S2 and S3, resulting in a loss of activity of the Cch1 [29]. Additionally, the residue Pro1228 is located in the extracellular S1-S2 linker of domain III and is well conserved across fungi and humans. Therefore, the mutation in this residue leads to a significant loss of activity of Cch1 [8].
3.2. Stretch-Activated Gating
Mid1 is the only known SACC in S. cerevisiae. So far, many mutation experiments have been employed to investigate the structure–function relationship of Mid1. Studies have shown that the mutant without the whole H3 or H4 segment and H3De or H4De loses the complement capacity in the lethality and shows low Ca2+ accumulation activity. In particular, the C373D and C373R proteins completely lost the Mid1 function related to viability and Ca2+ accumulation [6]. Therefore, segments H3 and H4 are essential for the Mid1 function. A recent study has proved that the Cys-498 residue at the C terminus of Mid1 is essential for Ca2+ channel activity [30].
3.3. Transient Receptor Gating
Yvc1, as a prominent voltage-dependent channel, possesses two gating systems for incorporated channels [31]. Gate one responds relatively fast to voltage fluctuations, and the probability of opening decreases as the potential tends towards positive but increases in the presence of the stilbene derivative 4,4-diisothiocyanato-stilbene-2,2-disulfonic acid (DIDS) at the cytoplasmic flank. In contrast, gate two responds relatively slowly to voltage fluctuations, and the probability of opening increases as the voltage potential tends toward positive. Moreover, Ca2+ is required at the cytoplasmic flank, which shifts voltage dependence towards negative.
Furthermore, Yvc1 is a mechanosensitive channel which can be activated to release Ca2+ from the vacuole to the cytoplasm by a transient osmotic force. The increase in osmolarity near the vacuole causes the shrinkage of the vacuole within seconds, inducing water withdrawal from the cytoplasm and vacuole, and this temporary osmotic imbalance produces an osmotic pressure across the vacuolar membrane [32]. It is reported that the membrane is stretched along its plane upon exerting pressure, either mechanical or osmotic, on the membrane regardless of the direction [33], which forces Yvc1 molecules into an open conformation. Eventually, the net water flux and osmotic pressure across the membrane recedes and ceases [32].
In a previous study, a novel structure–function model of mechanical force and cytoplasmic Ca2+ activating Yvc1 in parallel has been reported in S. cerevisiae [20]. In this model, the mechanical force acts through the membrane-associated domains, and Ca2+ binds to the negatively charged residues in the cytoplasmic domains, generating energy that reaches the gate in parallel. Recent studies have elucidated that Yvc1 gating in yeast is regulated by Ca2+, PI(3)P lipids, and membrane stretch. Furthermore, the mechanical force induces the release of Pl(3)P lipids from their binding sites and alters channel conformation, thereby promoting pore opening and Ca2+ flux from the vacuole to the cytosol [22].
3.4. Factors Affecting Ca2+ Channel Gating States
Several environmental factors can affect Ca2+ channel gating, including Ca2+ channel antagonists, aromatic compounds, oxidative stress, osmotic pressure, alkaline stress [34], temperature stress [35], cold stress [36], etc. Due to space limitations, only four main factors affecting Ca2+ channel gating have been discussed in this review (Table 1).
4. Applications of Ca2+ Channels in S. cerevisiae
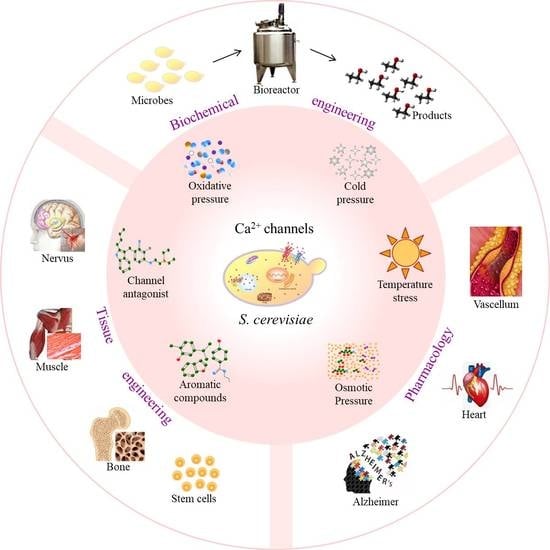
Studying the gating mechanisms and regulators of Ca2+ channels will significantly contribute to various applications in pharmacology, tissue engineering, and biochemical engineering (Figure 3).
4.1. Pharmacology
Ca2+ channels are essential targets for pharmaceutical research due to their implication in various diseases. S. cerevisiae has been widely used to study the pharmacological mechanism of drugs, including alkoxy-phenols of anti-tumor drugs and dihydropyridines of calcium channel blockers, by exploring biochemical alterations [7] due to the similarity between Cch1p and L-type VGCCs. Drugs in the dihydropyridine calcium channel blocker class, e.g., nifedipine and nimodipine, are likely to induce a permanent opening of Ca2+ channels by binding to the subunit α1, thus generating a non-controlled influx of Ca2+ into cells, which leads to a collapse of the membrane potential in S. cerevisiae [39]. Amiodarone, one of the most effective antiarrhythmic agents for chronic arrhythmias, can stimulate the activity of Mid1 but not Cch1. It is reported that a sustained increase in Ca2+ influx can cause cardiac arrhythmia. Thus, exploring the mechanism of Ca2+ homeostasis in yeast upon amiodarone inducement can provide insights into cardiac diseases [50].
S. cerevisiae has also been used as a model in studies exploring channelopathies. Particularly, Yvc1 is used as a model to study mucolipidosis type IV [3]. Moreover, the response of Cch1 to stress within the secretory pathway is consistent with the phenomenon observed in the mammalian cells, which is coupled to Ca2+ influx. Therefore, it is speculated that fungal and animal cells use a similar mechanism to trigger Ca2+ influx in response to secretory stress. Moreover, disrupting the function of presenilin-1 in the endoplasmic reticulum of neurons might affect several aspects of Ca2+ influx and signaling [51]. Accordingly, the study of Ca2+ channels in the yeast provided insights into familial Alzheimer’s disease [52].
4.2. Tissue Engineering
Current research on tissue engineering is mainly focused on producing functional replacement tissues and developing favorable conditions for tissue regeneration. L-VGCCs are significant for the osteogenic, myogenic, and neural differentiation of several types of stem cells. Thus, these L-VGCCs are involved in the function regulation of various stem cell types, including mesenchymal stem cells (MSCs) from the bone marrow, the oral cavity, adipose, and skin tissues, etc. Moreover, the corresponding regulatory mechanisms have been shown to influence cell proliferation and multipotent differentiation [53].
Recent studies have successfully demonstrated that targeting L-VGCCs modulates neurogenic activities due to the indispensable role of L-VGCCs in neurogenesis. Additionally, exposure to extremely low-frequency electromagnetic fields (i.e., 50 Hz) could increase the expression of VGCCs and modulate their function, especially CaV1.2 and CaV1.3 [54]. Ultimately, large-amplitude Ca2+ influx and a higher percentage of responsive neurons result in increased cell proliferation and neural differentiation. Overall, these results provide a reference for further research on tissue regeneration and development and broadening the potential applications of L-VGCCs in tissue engineering in the future.
4.3. Biochemical Engineering
It is well known that Ca2+ can significantly improve cell metabolism, thereby increasing the yield of the target product in microbial fermentation. Therefore, regulating the Ca2+ channels is a potential strategy for enhancing microbial productivity in green bio-manufacturing [45]. To date, very few reports have explored the regulation of Ca2+ channels in industrial fermentation.
It has been reported that ethanol production in S. cerevisiae was improved under the treatment of air cold plasma. In the further study, it has been explored that the Cch1/Mid1 in the cell membrane and Yvc1 in the vacuole membrane could be activated opening by the air cold plasma, and the increased cytoplasmic Ca2+ promotes the expression and activity of Pma1 H+-ATPase. Consequently, the cofactor metabolism from ATP and NADH is disturbed, enhancing ethanol production by S. cerevisiae [45]. In immobilized fermentation, biofilm plays a key role in improving industrial fermentation efficiency. In fact, Ca2+ channels CchA and MidA have been shown to upregulate biofilm formation in Aspergillus niger [55]. Therefore, regulating the Ca2+ channels in industrial microbes could provide a novel and promising strategy for bioprocess enhancement.
While elucidating the gating mechanism, exploring the crystal structures of Cch1 and Mid1 in S. cerevisiae is still challenging. Moreover, cloning of the CCH1 gene has been considered a major challenge due to its toxicity to E. coli. Consequently, direct studies on the CCH1 channel activity in the heterologous expression systems are inexecutable. In this regard, Vu et al. established a novel approach in which the shuttle plasmid CCH1-GFP, prepared in vitro and propagated in yeast, was successfully expressed in the HEK293 mammalian cell line [56]. This method has significantly increased the possibility of cloning and expressing Ca2+ channel genes in S. cerevisiae.
In the author’s research group, studies on Ca2+ channels in S. cerevisiae are currently ongoing. Moreover, the properties of genes and proteins related to the three Ca2+ channels have been analyzed by bioinformatics tools, with certain functional fragments of these genes being cloned, expressed, and purified successfully [57]. The used methods and obtained data in these studies will lay a theoretical and technical foundation for the further exploration of the crystal structures of Cch1 and Mid1 in S. cerevisiae.
5. Conclusions
The regulation of Ca2+ channels has emerged as a promising technology in the field of novel drug design, disease treatment, tissue regeneration, and microbial fermentation. The increasing amount of ongoing research on the regulation mechanisms of Ca2+ channels will significantly contribute to the rapid development of this promising technology. The present review provides a comprehensive summary of the protein composition, channel conformation, gating mechanism, and practical applications of three types of Ca2+ channels in S. cerevisiae, namely, Cch1, Mid1, and Yvc1, which have been considered as important models forscientific studies. Overall, this review will provide a reference for further studies and broaden the possible applications of Ca2+ channels in various fields.
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 21246012, 21306015, and 21476032).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article as no datasets were generated or analyzed in the course of the current study.
Acknowledgments
The author would like to extend her heartful gratitude to Barbara E. Ehrlich for providing the space to finish the original manuscript for her at Yale.
Conflicts of Interest
The author declares no conflict of interest.
References
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signaling: Dynamics, homeostasis and remodeling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Sze, H.; Liang, F.; Hwang, I.; Curran, A.C.; Harper, J.F. Diversity and regulation of plant Ca2+ pumps: Insights from expression in yeast. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 2000, 51, 433–462. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, D.M.; Pearce, D.A. Channeling studies in yeast as a model for channelopathies? Neuromol. Med. 2006, 8, 279–306. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Y. Enhanced ethanol production of Saccharomyces cerevisiae induced by cold plasma at atmospheric air. In Fuel Ethanol Production from Sugarcane, 1st ed.; Basso, T.P., Basso, L.C., Eds.; IntechOpen: London, UK, 2019; pp. 155–175. [Google Scholar]
- Palmer, C.P.; Zhou, X.L.; Lin, J.; Loukin, S.H.; Kung, C.; Saimi, Y. A TRP homolog in Saccharomyces cerevisiae forms an intracellular Ca2+-permeable channel in the yeast vacuolar membrane. Proc. Natl. Acad. Sci. USA 2001, 98, 7801–7805. [Google Scholar] [CrossRef]
- Tada, T.; Ohmori, M.; Iida, H. Molecular dissection of the hydrophobic segments H3 and H4 of the yeast Ca2+ channel component Mid1. J. Biol. Chem. 2003, 278, 9647–9654. [Google Scholar] [CrossRef]
- Teng, J.; Goto, R.; Iida, K.; Kojima, I.; Iida, H. Ion-channel blocker sensitivity of voltage-gated calcium-channel homologue Cch1 in Saccharomyces cerevisiae. Microbiology 2008, 154, 3775–3781. [Google Scholar] [CrossRef]
- Teng, J.; Iida, K.; Imai, A.; Nakano, M.; Tada, T.; Iida, H. Hyperactive and hypoactive mutations in Cch1, a yeast homologue of the voltage-gated calcium-channel pore-forming subunit. Microbiology 2013, 159, 970–979. [Google Scholar] [CrossRef]
- Fischer, M.; Schnell, N.; Chattaway, J.; Davies, P.; Dixon, G.; Sanders, D. The Saccharomyces cerevisiae CCH1 gene is involved in calcium influx and mating. FEBS Lett. 1997, 419, 259–262. [Google Scholar] [CrossRef]
- Chandel, A.; Bachhawat, A.K. Redox regulation of the yeast voltage-gated Ca2+ channel homolog Cch1p by glutathionylation of specific cysteine residues. J. Cell Sci. 2017, 130, 2317–2328. [Google Scholar] [CrossRef]
- Hong, M.P.; Vu, K.; Bautos, J.; Gelli, A. Cch1 restores intracellular Ca2+ in fungal cells during endoplasmic reticulum stress. J. Biol. Chem. 2010, 285, 10951–10958. [Google Scholar] [CrossRef]
- Cibulsky, S.M.; Sather, W.A. The EEEE locus is the sole high-affinity Ca2+ binding structure in the pore of a voltage-gated Ca2+ channel: Block by Ca2+ entering from the intracellular pore entrance. J. Gen. Physiol. 2000, 116, 349–362. [Google Scholar] [CrossRef]
- Prole, D.L.; Taylor, C.W. Identification and analysis of cation channel homologues in human pathogenic fungi. PLoS ONE 2012, 7, e42404. [Google Scholar] [CrossRef]
- Maruoka, T.; Nagasoe, Y.; Inoue, S.; Mori, Y.; Goto, J.; Ikeda, M.; Iida, H. Essential hydrophilic carboxyl-terminal regions including cysteine residues of the yeast stretch-activated calcium-permeable channel Mid1. J. Biol. Chem. 2002, 277, 11645–11652. [Google Scholar] [CrossRef]
- Tada, T.; Ohmori, M.; Iida, H. Phe356 in the yeast Ca2+ channel component Mid1 is a key residue for viability after exposure to α-factor. Biochem. Biophys. Res. Commun. 2004, 313, 752–757. [Google Scholar] [CrossRef]
- Iida, K.; Teng, J.; Cho, T.; Yoshikawa-Kimura, S.; Iida, H. Post-translational processing and membrane translocation of the yeast regulatory Mid1 subunit of the Cch1/VGCC/NALCN cation channel family. J. Biol. Chem. 2017, 292, 20570–20582. [Google Scholar] [CrossRef]
- Yoshimura, H.; Tada, T.; Iida, H. Subcellular localization and oligomeric structure of the yeast putative stretch-activated Ca2+ channel component Mid1. Exp. Cell. Res. 2004, 293, 185–195. [Google Scholar] [CrossRef]
- Christensen, A.P.; Corey, D.P. TRP channels in mechanosensation: Direct or indirect activation? Nat. Rev. Neurosci. 2007, 8, 510–521. [Google Scholar] [CrossRef]
- Chang, Y.; Schlenstedt, G.; Flockerzi, V.; Beck, A. Properties of the intracellular transient receptor potential (TRP) channel in Yeast, Yvc1. FEBS Lett. 2010, 584, 2028–2032. [Google Scholar] [CrossRef]
- Su, Z.; Zhou, X.; Loukin, S.H.; Saimi, Y.; Kung, C. Mechanical force and cytoplasmic Ca2+ activate yeast TRPY1 in parallel. J. Membr. Biol. 2009, 227, 141–150. [Google Scholar] [CrossRef]
- Chandel, A.; Das, K.K.; Bachhawat, A.K. Glutathione depletion activates the yeast vacuolar transient receptor potential channel, Yvc1p, by reversible glutathionylation of specific cysteines. Mol. Biol. Cell. 2016, 27, 3913–3925. [Google Scholar] [CrossRef]
- Ahmed, T.; Nisler, C.R.; Fluck, E.C., 3rd; Walujkar, S.; Sotomayor, M.; Moiseenkova-Bell, V.Y. Structure of the ancient TRPY1 channel from Saccharomyces cerevisiae reveals mechanisms of modulation by lipids and calcium. Structure 2022, 30, 139–155.e5. [Google Scholar] [CrossRef] [PubMed]
- Zamponi, G.W.; Striessnig, J.; Koschak, A.; Dolphin, A.C. The physiology, pathology, and pharmacology of voltage-gated calcium channels and their future therapeutic potential. Pharmacol. Rev. 2015, 67, 821–870. [Google Scholar] [CrossRef] [PubMed]
- Iida, K.; Tada, T.; Iida, H. Molecular cloning in yeast by in vivo homologous recombinant of the yeast putative α1 subunit of the voltage-gated calcium channel. FEBS Lett. 2004, 576, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yan, Z.; Li, Z.; Qian, X.; Lu, S.; Dong, M.; Zhou, Q.; Yan, N. Structure of the voltage-gated calcium channel Cav1.1 at 3.6 Å resolution. Nature 2016, 537, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yan, Z.; Li, Z.; Yan, C.; Lu, S.; Dong, M.; Yan, N. Structure of the voltage-gated calcium channel Cav1.1 complex. Science 2015, 350, 1492. [Google Scholar] [CrossRef] [PubMed]
- Beyl, S.; Depil, K.; Hohaus, A.; Stary-Weinzinger, A.; Linder, T.; Timin, E.; Hering, S. Neutralisation of a single voltage sensor affects gating determinants in all four pore-forming S6 segments of Cav1.2: A cooperative gating model. Pflügers Arch. Eur. J. Physiol. 2012, 464, 391–401. [Google Scholar] [CrossRef]
- Xie, C.; Zhen, X.; Yang, J. Localization of the activation gate of a voltage-gated Ca2+ channel. J. Gen. Physiol. 2005, 126, 205–212. [Google Scholar] [CrossRef]
- Iida, K.; Teng, J.; Tada, T.; Saka, A.; Tamai, M.; Izumi-Nakaseko, H.; Adachi-Akahane, S.; Iida, H. Essential, completely conserved glycine residue in the domain III S2-S3 linker of voltage-gated calcium channel alpha1 subunits in yeast and mammals. J. Biol. Chem. 2007, 282, 25659–25667. [Google Scholar] [CrossRef]
- Hayashi, T.; Oishi, K.; Kimura, M.; Iida, K.; Iida, H. Highly conserved extracellular residues mediate interactions between pore-forming and regulatory subunits of the yeast Ca2+ channel related to the animal VGCC/NALCN family. J. Biol. Chem. 2020, 295, 13008–13022. [Google Scholar] [CrossRef]
- Tanifuji, M.; Sato, M.; Wada, Y.; Anraku, Y.; Kasai, M. Gating behaviors of a voltage-dependent and Ca2+-activated cation channel of yeast vacuolar membrane incorporated into planar lipid bilayer. J. Membr. Biol. 1988, 106, 47–55. [Google Scholar] [CrossRef]
- Zhou, X.L.; Batiza, A.F.; Loukin, S.H.; Palmer, C.P.; Kung, C.; Saimi, Y. The transient receptor potential channel on the yeast vacuole is mechanosensitive. Proc. Natl. Acad. Sci. USA 2003, 100, 7105–7110. [Google Scholar] [CrossRef]
- Hamill, O.P.; Martinac, B. Molecular basis of mechanotransduction in living cells. Physiol. Rev. 2001, 81, 685–740. [Google Scholar] [CrossRef]
- Viladevall, L.; Serrano, R.; Ruiz, A.; Domenech, G.; Giraldo, J.; Barcelo, A.; Arino, J. Characterization of the calcium-mediated response to alkaline stress in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 43614–43624. [Google Scholar] [CrossRef]
- Liu, M.; Du, P.; Heinrich, G.; Cox, G.M.; Gelli, A. Cch1 mediates calcium entry in Cryptococcus neoformans and is essential in low-calcium environments. Eukaryot. Cell 2006, 5, 1788–1796. [Google Scholar] [CrossRef]
- Peiter, E.; Fischer, M.; Sidaway, K.; Roberts, S.K.; Sanders, D. The Saccharomyces cerevisiae Ca2+ channel Cch1pMid1p is essential for tolerance to cold stress and iron toxicity. FEBS Lett. 2005, 579, 5697–5703. [Google Scholar] [CrossRef]
- Maresova, L.; Muend, S.; Zhang, Y.Q.; Sychrova, H.; Rao, R. Membrane hyperpolarization drives cation influx and fungicidal activit of amiodarone. J. Biol. Chem. 2009, 284, 2795–2802. [Google Scholar] [CrossRef]
- Muend, S.; Rao, R. Fungicidal activity of amiodarone is tightly coupled to calcium influx. FEMS Yeast Res. 2008, 8, 425–431. [Google Scholar] [CrossRef]
- Pereira, R.S. The use of calcium blockers to study biochemical behavior of Saccharomyces cerevisiae cells. Mol. Cell. Biochem. 2001, 228, 1–7. [Google Scholar] [CrossRef]
- Haynes, W.J.; Zhou, X.L.; Su, Z.W.; Loukin, S.H.; Saimi, Y.; Kung, C. Indole and other aromatic compounds activate the yeast TRPY1 channel. FEBS Lett. 2008, 582, 1514–1518. [Google Scholar] [CrossRef]
- Lee, J.H.; Wendisch, V.F. Biotechnological production of aromatic compounds of the extended shikimate pathway from renewable biomass. J. Biotechnol. 2017, 257, 211–221. [Google Scholar] [CrossRef]
- Monk, B.C.; Goffeau, A. Outwitting multidrug resistance to antifungals. Science 2008, 321, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.K.; McAinsh, M.; Cantopher, H.; Sandison, S. Calcium dependence of eugenol tolerance and toxicity in Saccharomyces cerevisiae. PLoS ONE 2014, 9, e102712. [Google Scholar] [CrossRef] [PubMed]
- Bogeski, I.; Kummerow, C.; Al-Ansary, D.; Schwarz, E.C.; Koehler, R.; Kozai, D.; Takahashi, N.; Peinelt, C.; Griesemer, D.; Bozem, M.; et al. Differential redox regulation of ORAI ion channels: A mechanism to tune cellular calcium signaling. Sci. Signal. 2010, 3, ra24. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Y.; Yuan, X.; Wang, R.J. Interaction of air cold plasma with Saccharomyces cerevisiae in the multi-scale microenvironment for improved ethanol yield. Bioresour. Technol. 2021, 323, 124621. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, S.; Mori, Y.; Yabe, I.; Uozumi, N. In vitro and in vivo characterization of modulation of the vacuolar cation channel TRPY1 from Saccharomyces cerevisiae. FEBS J. 2018, 285, 1146–1161. [Google Scholar] [CrossRef]
- Popa, C.V.; Dumitru, I.; Ruta, L.L.; Danet, A.F.; Farcasanu, I.C. Exogenous oxidative stress induces Ca2+ release in the yeast Saccharomyces cerevisiae. FEBS J. 2010, 277, 4027–4038. [Google Scholar] [CrossRef]
- Courchesne, W.E.; Vlasek, C.; Klukovich, R.; Coffee, S. Ethanol induces calcium influx via the Cch1-Mid1 transporter in Saccharomyces cerevisiae. Arch. Microbiol. 2011, 193, 323–334. [Google Scholar] [CrossRef]
- Groppi, S.; Belotti, F.; Brandao, R.L.; Martegani, E.; Tisi, R. Glucose-induced calcium influx in budding yeast involves a novel calcium transport system and can activate calcineurin. Cell Calcium 2011, 49, 376–386. [Google Scholar] [CrossRef]
- Cui, J.; Kaandorp, J.A.; Sloot, P.M.A.; Lloyd, C.M.; Filatov, M.V. Calcium homeostasis and signaling in yeast cells and cardiac myocytes. FEMS Yeast Res. 2009, 9, 1137–1147. [Google Scholar] [CrossRef]
- Imaizumi, K.; Katayama, T.; Tohyma, M. Presenilin and the UPR. Nat. Cell Biol. 2001, 3, E104. [Google Scholar] [CrossRef]
- Paschen, W.; Frandsen, A. Endoplasmic reticulum dysfunction-a common denominator for cell injury in acute and degenerative diseases of the brain? J. Neurochem. 2002, 79, 719–725. [Google Scholar] [CrossRef]
- Tan, Y.Z.; Fei, D.D.; He, X.N.; Dai, J.M.; Xu, R.C.; Xu, X.Y.; Wu, J.J.; Li, B. L-type voltage-gated calcium channels in stem cells and tissue engineering. Cell Proliferat 2019, 52, e12623. [Google Scholar] [CrossRef]
- Piacentini, R.; Ripoli, C.; Mezzogori, D.; Azzena, G.B.; Grassi, C. Extremely low-frequency electromagnetic fields promote in vitro neurogenesis via upregulation of Cav1-channel activity. J. Cell. Physiol. 2008, 215, 129–139. [Google Scholar] [CrossRef]
- Liu, L.; Yu, B.; Sun, W.; Liang, C.; Ying, H.; Zhou, S.; Niu, H.; Wang, Y.; Liu, D.; Chen, Y. Calcineurin signaling pathway influences Aspergillus niger biofilm formation by affecting hydrophobicity and cell wall integrity. Biotechnol. Biofuels 2020, 13, 54. [Google Scholar] [CrossRef]
- Vu, K.; Bautos, J.; Hong, M.P.; Gelli, A. The functional expression of toxic genes: Lessons learned from molecular cloning of CCH1, a high-affinity Ca2+ channel. Anal. Biochem. 2009, 393, 234–241. [Google Scholar] [CrossRef]
- Dong, X.Y. Preparation and identification of monoclonal antibodies of calcium channel membrane proteins in Saccharomyces cerevisiae. Chem. Ind. Eng. Prog. 2021, 40, 334–343. [Google Scholar]
Figure 1.
Calcium ion (Ca2+) channels in Saccharomyces cerevisiae. (A) Channel localization. (B–D) Predicted topologies of Ca2+ channels Cch1, Mid1, and Yvc1; the plus sign indicates the positively charged amino acids; solid circle indicates the cysteine (Cys) residues (Cch1, Yvc1) or Cys-rich regions (Mid1); hollow circle indicates the EF-hand-like motif of helix–loop–helix structure; solid triangle indicates the CKII phosphorylation motif; hollow triangle indicates the sheet–turn–sheet structure; solid square indicates the hydrophobic patch between the amino acid residues 509 and 518; hollow square indicates the negatively charged cluster 573DDDD576.
Figure 1.
Calcium ion (Ca2+) channels in Saccharomyces cerevisiae. (A) Channel localization. (B–D) Predicted topologies of Ca2+ channels Cch1, Mid1, and Yvc1; the plus sign indicates the positively charged amino acids; solid circle indicates the cysteine (Cys) residues (Cch1, Yvc1) or Cys-rich regions (Mid1); hollow circle indicates the EF-hand-like motif of helix–loop–helix structure; solid triangle indicates the CKII phosphorylation motif; hollow triangle indicates the sheet–turn–sheet structure; solid square indicates the hydrophobic patch between the amino acid residues 509 and 518; hollow square indicates the negatively charged cluster 573DDDD576.

Figure 2.
Structure of the α1 subunit in Cav1.1 [25,26]. (A) Overall structure of α1 subunit; CTD, C- terminal domain; red spheres indicate the tentatively assigned calcium ions (Ca2+) in the selectivity filter vestibule. (B) Structural elements of the selectivity filter. (C) Four-fold pseudo-symmetry of the pore domain; loops between S5 and P1 helices and between P2 and S6 helices are shown as L5 and L6 loops, respectively. (D) Structures of the four voltage-sensing domains (VSDs). Gating charges on helix S4 and the occluding phenylalanine residue on S2 are shown as sticks.
Figure 2.
Structure of the α1 subunit in Cav1.1 [25,26]. (A) Overall structure of α1 subunit; CTD, C- terminal domain; red spheres indicate the tentatively assigned calcium ions (Ca2+) in the selectivity filter vestibule. (B) Structural elements of the selectivity filter. (C) Four-fold pseudo-symmetry of the pore domain; loops between S5 and P1 helices and between P2 and S6 helices are shown as L5 and L6 loops, respectively. (D) Structures of the four voltage-sensing domains (VSDs). Gating charges on helix S4 and the occluding phenylalanine residue on S2 are shown as sticks.

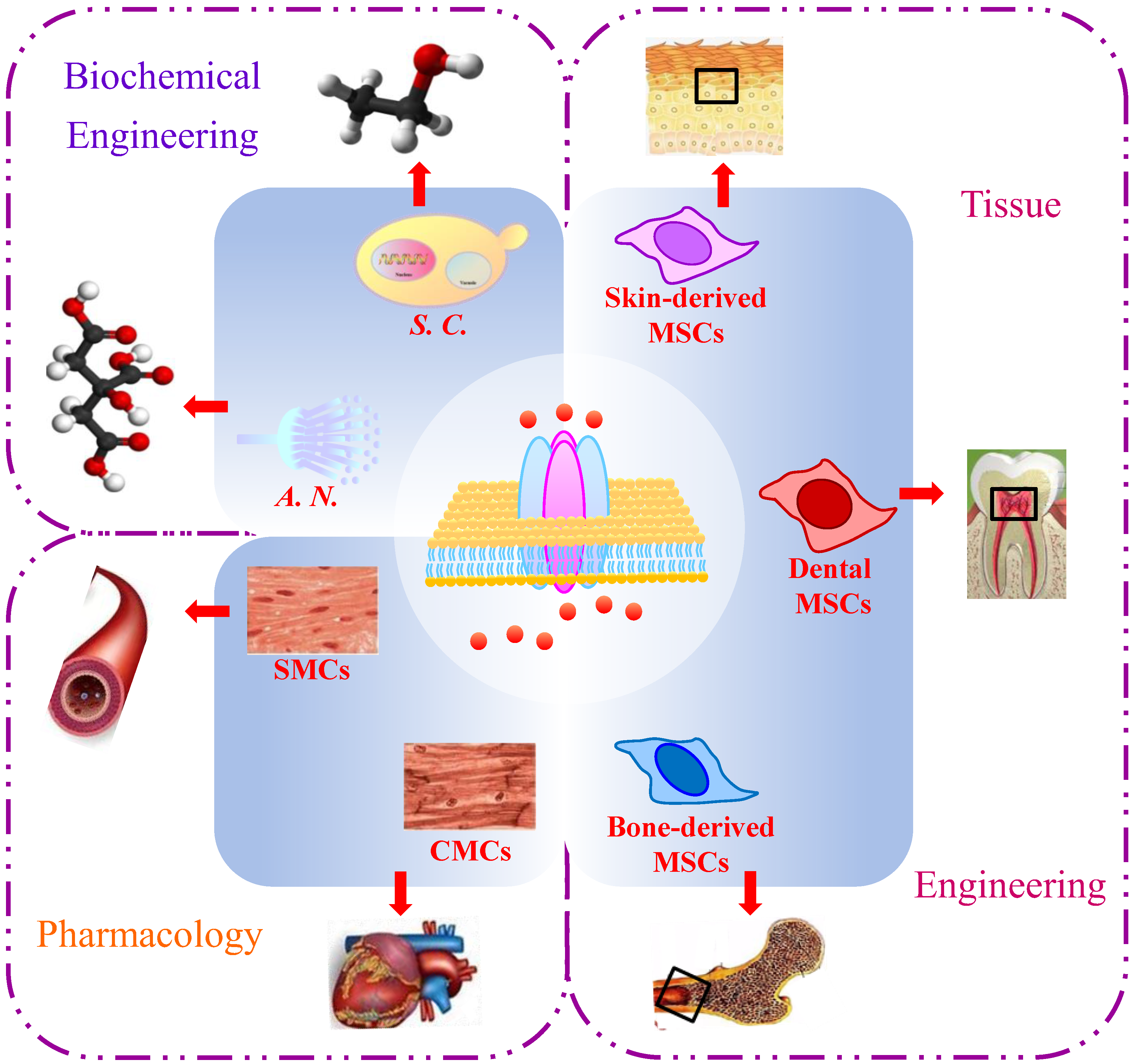
Figure 3.
Applications of calcium ion (Ca2+) channels in Saccharomyces cerevisiae. SMCs, smooth muscle cells; CMCs, cardiac muscle cells; S. C., Saccharomyces cerevisiae; A. N., Aspergillus niger; MSCs, mesenchymal stem cells.
Figure 3.
Applications of calcium ion (Ca2+) channels in Saccharomyces cerevisiae. SMCs, smooth muscle cells; CMCs, cardiac muscle cells; S. C., Saccharomyces cerevisiae; A. N., Aspergillus niger; MSCs, mesenchymal stem cells.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Environmental factors affecting calcium ion (Ca2+) channel gating.
| Affecting Factors | Reagents | Calcium Ion Channels | Effects | Gating Mechanism | Applications | References |
|---|---|---|---|---|---|---|
| Calcium ion (Ca2+) channel antagonists/blockers | Amiodarone, amlodipine, diltiazem, nicardipine, nifedipine, nimodipine, nitrendipine, verapamil | Cch1, Cch1-Mid1, or Mid1 | Blocking calcium channels or stimulating channel opening | Binding to the special sites on Ca2+ channels for blockage | Clinical treatment of hypertension, coronary heart disease, and cardiac arrhythmia | [7,37,38,39] |
| Aromatic compounds | Carvacrol, eugenol, indole, methylated, propylparaben, quinolone, thymol | Cch1 or Yvc1 | Improving cell tolerance or mediating the Ca2+ increase in the cytoplasm | Competing with the aromatic residues of the channel proteins for lipid anchors or generating force profile in the bilayer for open conformation | Antifungal drugs | [40,41,42,43] |
| Oxidative stress | Chloramine T, dithiothreitol, hydrogen peroxide, 2-mercaptoethanol, mitochondrion, N-ethylmaleimide, plasma discharge, reduced glutathione, β-phenylethylamine, tert-butylhydroperoxide (tBOOH) | Cch1, Mid1, or Yvc1 | Affecting activity, expression, open-time probability, or trafficking | Specific glutathionylation of cysteine residues in the pore forming region; oxidation of special amino acids; change in the channel conformation | Experiments on channel functions or metabolic regulation | [10,44,45,46,47] |
| Osmotic pressure | Ethanol, hexose (glucose and galactose), LiCl, NaCl, sorbitol | Cch1, Mid1, or Yvc1 | Channel opening or improving ethanol tolerance | Membrane perturbation, vacuolar shrinkage and deformation, or activation of the PKC1 pathway | Industrial microbial fermentation | [32,48,49] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dong, X.-Y. Calcium Ion Channels in Saccharomyces cerevisiae. J. Fungi 2023, 9, 524. https://doi.org/10.3390/jof9050524
AMA Style
Dong X-Y. Calcium Ion Channels in Saccharomyces cerevisiae. Journal of Fungi. 2023; 9(5):524. https://doi.org/10.3390/jof9050524
Chicago/Turabian StyleDong, Xiao-Yu. 2023. "Calcium Ion Channels in Saccharomyces cerevisiae" Journal of Fungi 9, no. 5: 524. https://doi.org/10.3390/jof9050524
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.