
The AMP-Activated Protein Kinase (AMPK) Positively Regulates Lysine Biosynthesis Induced by Citric Acid in Flammulina filiformis
 and
and 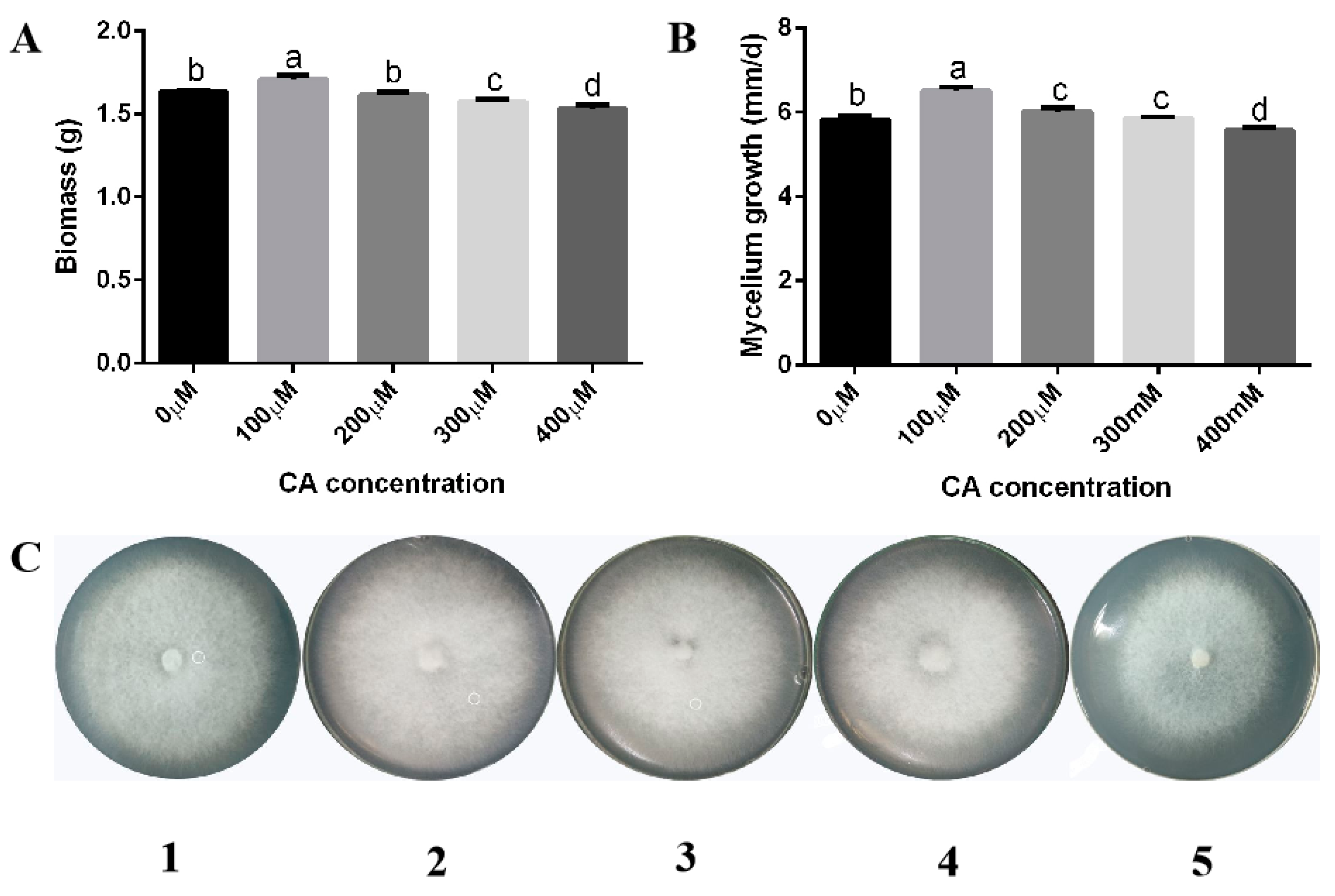{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Procedures
2.1. Strains and Culture Conditions
2.2. Mycelium Treatment Conditions
2.3. Fruiting Body Cultivation of F. filiformis Induced by Citric Acid
2.4. Measurement of Mycelium Biomass and Growth Rate
2.5. Determination of Lysine Content
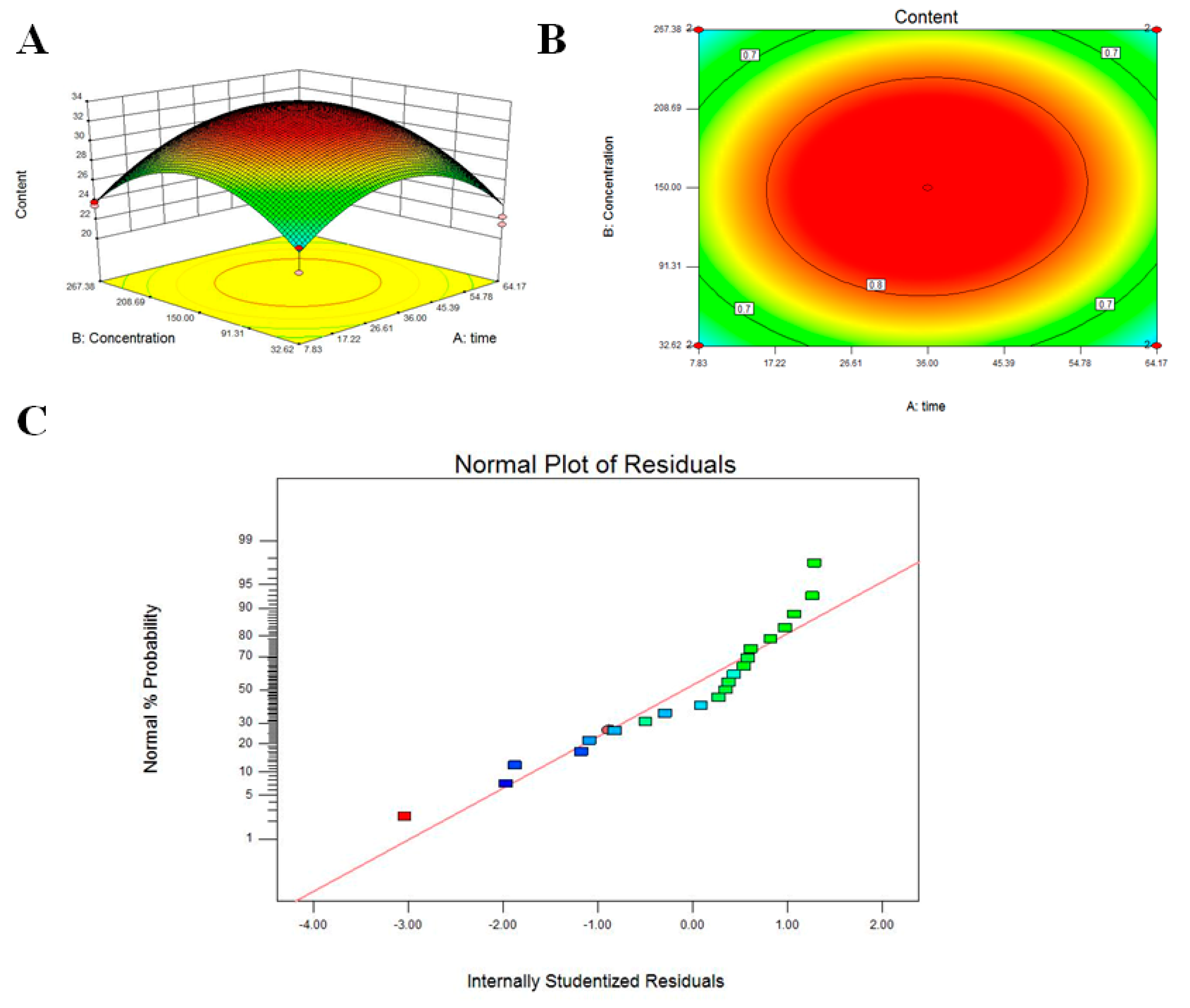
2.6. Optimization by Response Surface Methodology
2.7. RNA Extraction and Gene Expression Analysis
2.8. AMPK-Silencing Strain Construction
2.9. Western Blot for AMPK Total Protein and Phosphorylation Levels
2.10. Statistical Analysis
3. Results
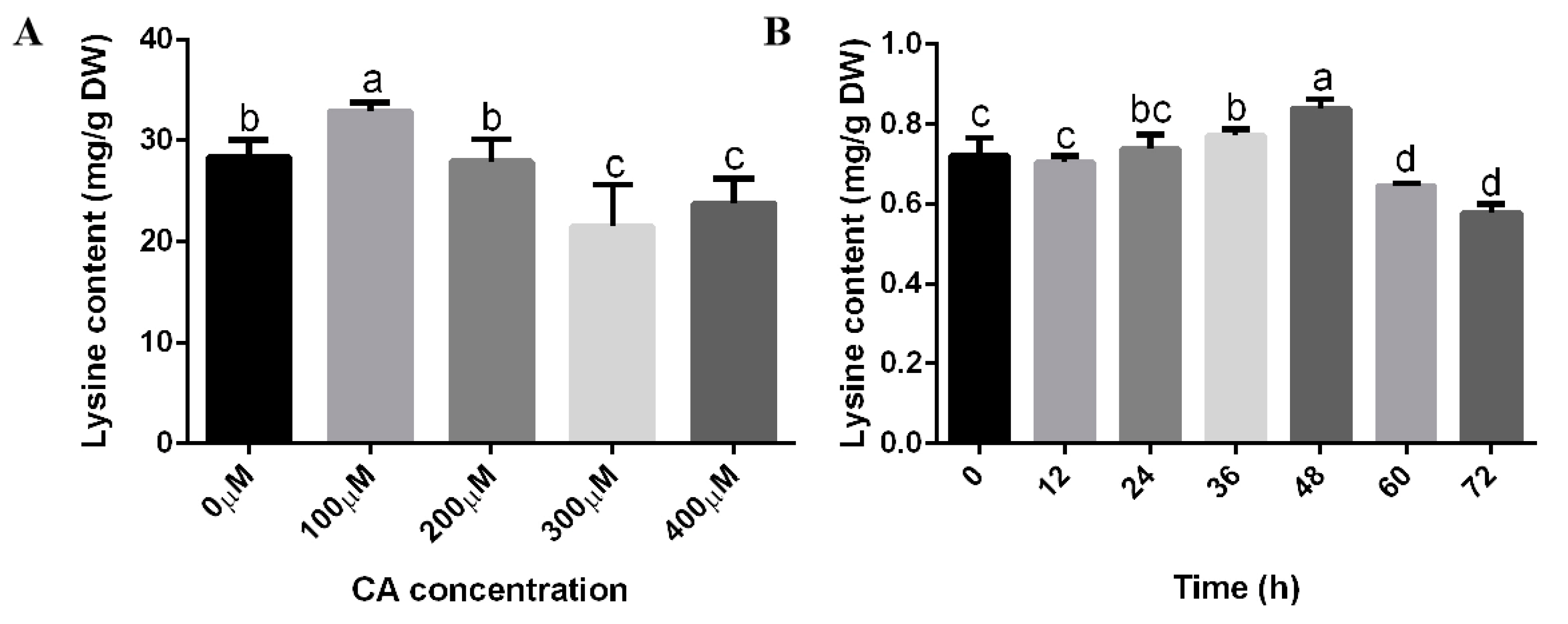
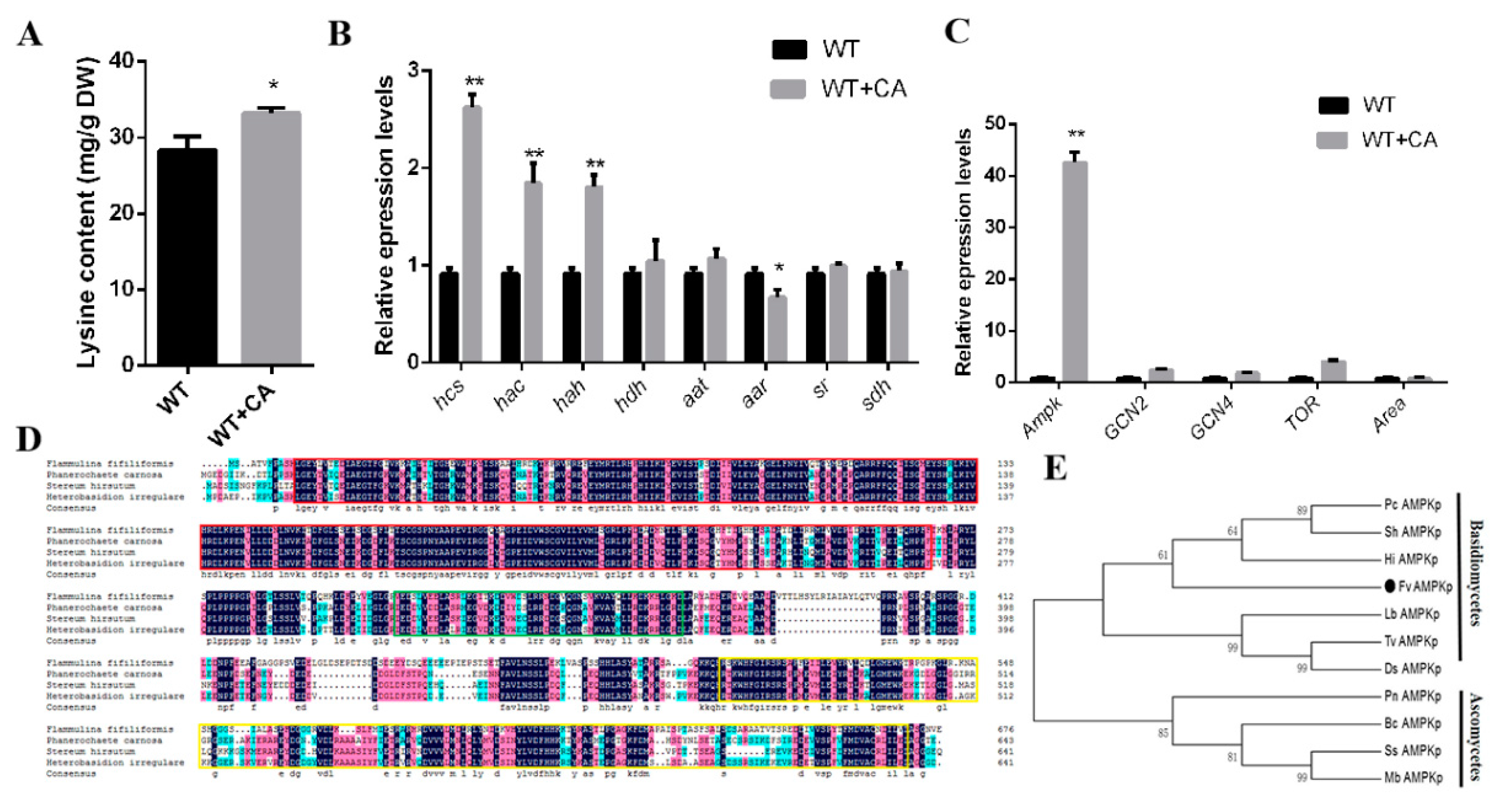
3.1. Citric Acid Induction Increased the Biomass, Growth Rate, and Lysine Content of F. filiformis
3.2. Response Surface Methodology to Optimize Citric Acid Induction Conditions
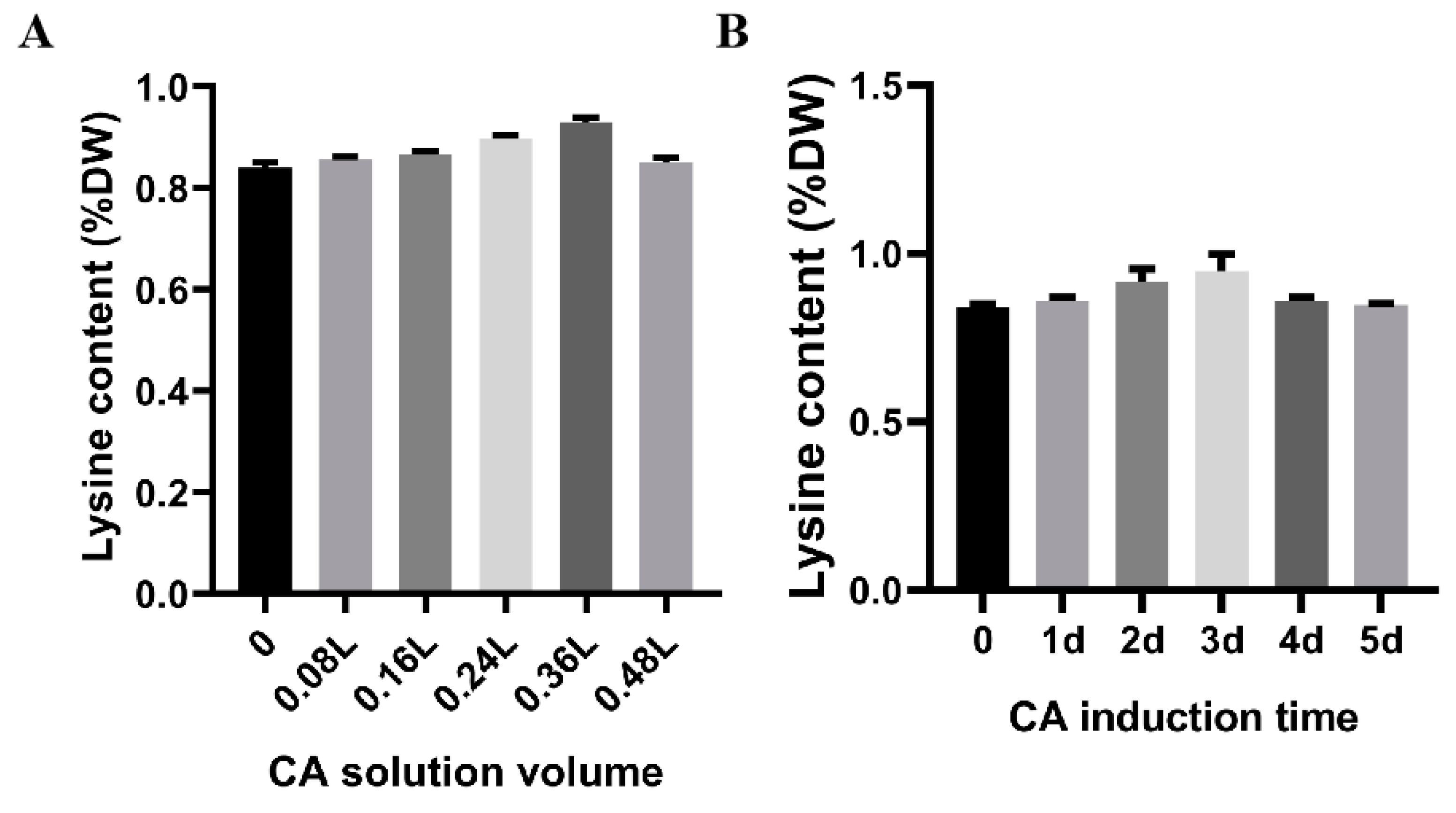
3.3. Citric Acid Induction Changes the Lysine Content of F. filiformis Fruiting Bodies
3.4. Changes in Lysine Content, Gene Transcription Level, and Key Regulators under Optimal Citric Acid Induction Conditions
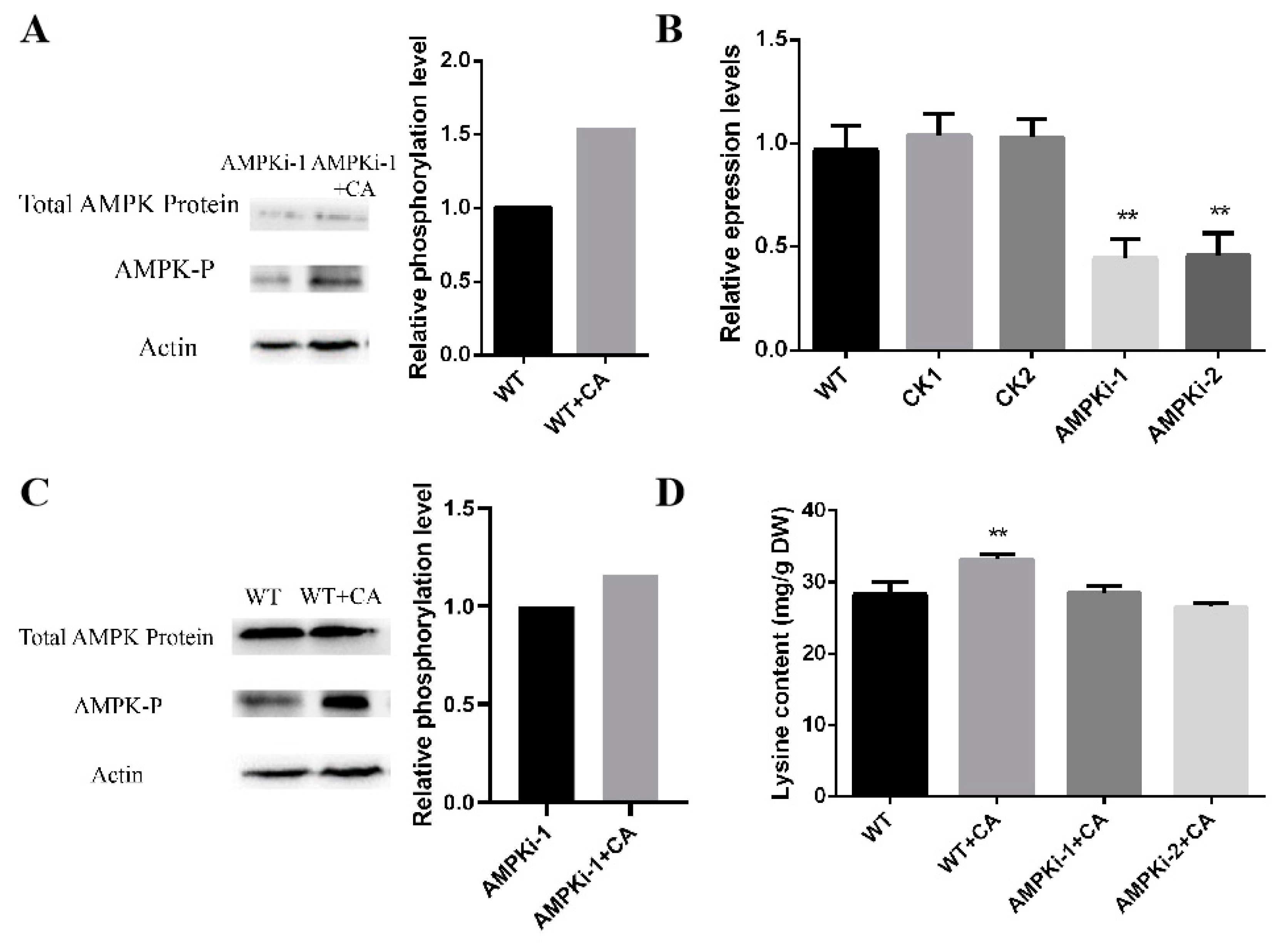
3.5. Changes in the Phosphorylation and Lysine Content of Ampk-Silenced Strains under Optimal Induction by Citric Acid
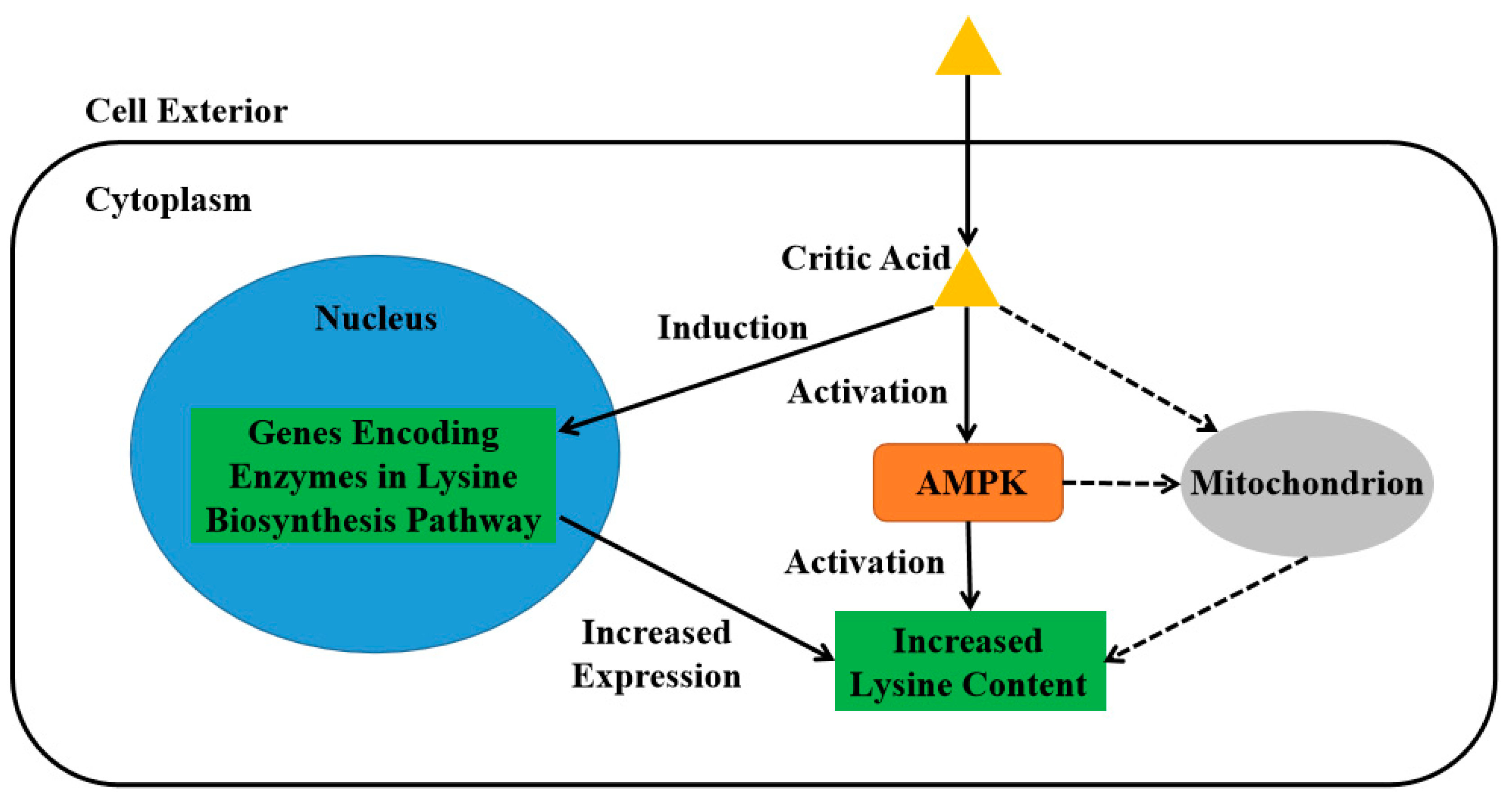
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar]
- Cong, W.R.; Liu, Y.; Li, Q.Z.; Zhou, X.W. Cloning and analysis of a functional promoter of fungal immunomodulatory protein from Flammulina velutipes. Mol. Biol. Rep. 2014, 41, 4381–4387. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wu, Y.Y.; Fang, D.L.; Pei, F.; Mariga, A.M.; Yang, W.J.; Hu, Q.H. Effect of nanocomposite packaging on postharvest senescence of Flammulina velutipes. Food Chem. 2018, 246, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shi, L.; Tang, W.; Xia, W.; Zhong, Y.; Xu, X.; Xie, B.; Tao, Y. Comprehensive Genetic Analysis of Monokaryon and Dikaryon Populations Provides Insight into Cross-Breeding of Flammulina filiformis. Front. Microbiol. 2022, 13, 887259. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Dong, J.; Liao, J.; Tian, L.; Qiu, H.; Wu, T.; Ge, F.; Zhu, J.; Shi, L.; Jiang, A.; et al. Establishment of CRISPR/Cas9 Genome-Editing System Based on Dual sgRNAs in Flammulina filiformis. J. Fungi 2022, 8, 693. [Google Scholar] [CrossRef]
- Ribeiro, B.; Andrade, P.B.; Silva, B.M.; Baptista, P.; Seabra, R.M.; Valentao, P. Comparative Study on Free Amino Acid Composition of Wild Edible Mushroom Species. J. Agric. Food Chem. 2008, 56, 10973–10979. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Carbonero, E.R.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Characterization of a heterogalactan: Some nutritional values of the edible mushroom Flammulina velutipes. Food Chem. 2008, 108, 329–333. [Google Scholar] [CrossRef]
- Cai, H.H.; Liu, X.M.; Chen, Z.Y.; Liao, S.T.; Zou, Y.X. Isolation, purification and identification of nine chemical compounds from Flammulina velutipes fruiting bodies. Food Chem. 2013, 141, 2873–2879. [Google Scholar] [CrossRef]
- Irla, M.; Wendisch, V.F. Efficient cell factories for the production of N-methylated amino acids and for methanol-based amino acid production. Microb. Biotechnol. 2022, 15, 2145–2159. [Google Scholar] [CrossRef]
- Lou, H.H.; Li, H.; Wei, T.Y.; Chen, Q.H. Stimulatory Effects of Oleci Acid and Fungal Elicitor on Betulinic Acid Production by Submerged Cultivation of Medicinal Mushroom Inonotus obliquus. J. Fungi 2021, 7, 266. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.C.; Liu, R.P.; Tong, X.Y.; Zou, L. New Insights into Methyl Jasmonate Regulation of Triterpenoid Biosynthesis in Medicinal Fungal Species Sanghuangporus baumii (Pilat) LW Zhou & YC Dai. J. Fungi 2022, 8, 889. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L. Notes on the nomenclature of five important edible fungi in China. Mycosystema 2018, 37, 1572–1577. [Google Scholar]
- Fu, Y.P.; Liang, Y.; Dai, Y.T.; Yang, C.T.; Duan, M.Z.; Zhang, Z.; Hu, S.N.; Zhang, Z.W.; Li, Y. De Novo Sequencing and Transcriptome Analysis of Pleurotus eryngii subsp. tuoliensis (Bailinggu) Mycelia in Response to Cold Stimulation. Molecules 2016, 21, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.N.; Xia, X.X.; Zhong, J.J. Induced effect of Na plus on ganoderic acid biosynthesis in static liquid culture of Ganoderma lucidum via calcineurin signal transduction. Biotechnol. Bioeng. 2013, 110, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.N.; Xia, X.X.; Zhong, J.J. Induction of Ganoderic Acid Biosynthesis by Mn2+ in Static Liquid Cultivation of Ganoderma Lucidum. Biotechnol. Bioeng. 2014, 111, 2358–2365. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Ma, Z.; Qiu, H.; Liu, X.; Wu, T.; Ge, F.; Liu, R.; Zhu, J.; Shi, L.; Jiang, A.; et al. Chitosan Increases Lysine Content through Amino Acid Transporters in Flammulina filiformis. Foods 2022, 11, 2163. [Google Scholar] [CrossRef]
- Xu, H.Y.; Andi, B.; Qian, J.H.; West, A.H.; Cook, P.F. The alpha-aminoadipate pathway for lysine biosynthesis in fungi. Cell Biochem. Biophys. 2006, 46, 43–64. [Google Scholar] [CrossRef]
- Zabriskie, T.M.; Jackson, M.D. Lysine biosynthesis and metabolism in fungi. Nat. Prod. Rep. 2000, 17, 85–97. [Google Scholar] [CrossRef]
- Lopez-Bucio, J.; Nieto-Jacobo, M.F.; Ramirez-Rodriguez, V.; Herrera-Estrella, L. Organic acid metabolism in plants: From adaptive physiology to transgenic varieties for cultivation in extreme soils. Plant Sci. 2000, 160, 1–13. [Google Scholar] [CrossRef]
- Gong, M.; Huang, T.Y.; Li, Y.; Li, J.X.; Tang, L.H.; Su, E.Z.; Zou, G.; Bao, D.P. Multi-Omics Analysis of Low-Temperature Fruiting Highlights the Promising Cultivation Application of the Nutrients Accumulation in Hypsizygus marmoreus. J. Fungi 2022, 8, 695. [Google Scholar] [CrossRef]
- Wu, T.J.; Hu, C.C.; Xie, B.G.; Zhang, L.; Yan, S.J.; Wang, W.; Tao, Y.X.; Li, S.J. A Single Transcription Factor (PDD1) Determines Development and Yield of Winter Mushroom (Flammulina velutipes). Appl. Environ. Microbiol. 2019, 85, e01735-19. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Xia, J.; Ge, F.; Qiu, H.; Tian, L.; Liu, X.; Liu, R.; Jiang, A.; Zhu, J.; Shi, L.; et al. Target of Rapamycin Mediated Ornithine Decarboxylase Antizyme Modulate Intracellular Putrescine and Ganoderic Acid Content in Ganoderma lucidum. Microbiol. Spectr. 2022, 10, e0163322. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, Z.H.; Ren, A.; Shi, L.; Shi, D.K.; Li, X.B.; Zhao, M.W. The mitogen-activated protein kinase GlSlt2 regulates fungal growth, fruiting body development, cell wall integrity, oxidative stress and ganoderic acid biosynthesis in Ganoderma lucidum. Fungal Genet. Biol. 2017, 104, 6–15. [Google Scholar] [CrossRef]
- Sattlegger, E.; Swanson, M.J.; Ashcraft, E.A.; Jennings, J.L.; Fekete, R.A.; Link, A.J.; Hinnebusch, A.G. YIH1 is an actin-binding protein that inhibits protein kinase GCN2 and impairs general amino acid control when overexpressed. J. Biol. Chem. 2004, 279, 29952–29962. [Google Scholar] [CrossRef] [Green Version]
- Gallinetti, J.; Harputlugil, E.; Mitchell, J.R. Amino acid sensing in dietary-restriction-mediated longevity: Roles of signal-transducing kinases GCN2 and TOR. Biochem. J. 2013, 449, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Horst, R.J.; Zeh, C.; Saur, A.; Sonnewald, S.; Sonnewald, U.; Voll, L.M. The Ustilago maydis Nit2 Homolog Regulates Nitrogen Utilization and Is Required for Efficient Induction of Filamentous Growth. Eukaryot. Cell 2012, 11, 368–380. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Y.; Chang, M.C.; Meng, J.L.; Feng, C.P.; Wang, Y. A Comparative Proteome Approach Reveals Metabolic Changes Associated with Flammulina velutipes Mycelia in Response to Cold and Light Stress. J. Agric. Food Chem. 2018, 66, 3716–3725. [Google Scholar] [CrossRef]
- Khatun, M.R.; Mukta, R.H.; Islam, M.A.; Huda, A. Insight into citric acid-induced chromium detoxification in rice (Oryza sativa. L). Int. J. Phytoremediation 2019, 21, 1234–1240. [Google Scholar] [CrossRef]
- Teves, F.; Lamas-Maceiras, M.; Garcia-Estrada, C.; Casqueiro, J.; Naranjo, L.; Ullan, R.V.; Scervino, J.M.; Wu, X.B.; Velasco-Conde, T.; Martin, J.F. Transcriptional upregulation of four genes of the lysine biosynthetic pathway by homocitrate accumulation in Penicillium chrysogenum: Homocitrate as a sensor of lysine-pathway distress. Microbiology-Sgm 2009, 155, 3881–3892. [Google Scholar] [CrossRef] [Green Version]
- O’Doherty, P.J.; Lyons, V.; Tun, N.M.; Rogers, P.J.; Bailey, T.D.; Wu, M.J. Transcriptomic and biochemical evidence for the role of lysine biosynthesis against linoleic acid hydroperoxide-induced stress in Saccharomyces cerevisiae. Free. Radic. Res. 2014, 48, 1454–1461. [Google Scholar] [CrossRef]
- Zhou, L.B.; Zeng, A.P. Exploring Lysine Riboswitch for Metabolic Flux Control and Improvement of L-Lysine Synthesis in Corynebacterium glutamicum. ACS Synth. Biol. 2015, 4, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Li, Q.Z.; Jiang, P.Y.; Xu, Z.; Zhang, D.; Zhang, L.J.; Zhang, M.Y.; Yu, H.L.; Song, C.Y.; Tan, Q.; et al. Overexpression of the saccharopine dehydrogenase gene improves lysine biosynthesis in Flammulina velutipes. J. Basic Microbiol. 2019, 59, 890–900. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yang, Z.Y.; Chen, X.; Han, D.; Han, J.; Wang, L.S.; Ren, A.; Yu, H.S.; Zhao, M.W. Lenthionine, a Key Flavor Substance in Lentinula edodes, Is Regulated by Cysteine under Drought Stress. J. Agric. Food Chem. 2021, 69, 12645–12653. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, Y.; Natarajan, K. Reconfiguration of Transcriptional Control of Lysine Biosynthesis in Candida albicans Involves a Central Role for the Gcn4 Transcriptional Activator. Msphere 2016, 1, e00016-15. [Google Scholar] [CrossRef] [Green Version]
- Woods, A.; Dickerson, K.; Heath, R.; Hong, S.P.; Momcilovic, M.; Johnstone, S.R.; Carlson, M.; Carling, D. Ca2+/calmodulin-dependent protein kinase kinase-beta acts upstream of AMP-activated protein kinase in mammalian cells. Cell Metab. 2005, 2, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, B.; Sanders, M.J.; Underwood, E.; Heath, R.; Mayer, F.V.; Carmena, D.; Jing, C.; Walker, P.A.; Eccleston, J.F.; Haire, L.F.; et al. Structure of mammalian AMPK and its regulation by ADP. Nature 2011, 472, 230–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baena-Gonzalez, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A central integrator of transcription networks in plant stress and energy signalling. Nature 2007, 448, 938–942. [Google Scholar] [CrossRef]
- Backhaus, K.; Rippert, D.; Heilmann, C.J.; Sorgo, A.G.; de Koster, C.G.; Klis, F.M.; Rodicio, R.; Heinisch, J.J. Mutations in SNF1 complex genes affect yeast cell wall strength. Eur. J. Cell Biol. 2013, 92, 383–395. [Google Scholar] [CrossRef]
- Dai, Y.C.; Cao, Y.; Zhou, L.W.; Wu, S.H. Notes on the nomenclature of the most widely cultivated Ganoderma species in China. Mycosystema 2013, 32, 947–952. [Google Scholar]
- Hu, Y.R.; Xu, W.Z.; Hu, S.S.; Lian, L.D.; Zhu, J.; Shi, L.; Ren, A.; Zhao, M.W. In Ganoderma lucidum, Glsnf1 regulates cellulose degradation by inhibiting GlCreA during the utilization of cellulose. Environ. Microbiol. 2020, 22, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.R.; Xu, W.Z.; Hu, S.S.; Lian, L.D.; Zhu, J.; Ren, A.; Shi, L.; Zhao, M.W. Glsnf1-mediated metabolic rearrangement participates in coping with heat stress and influencing secondary metabolism in Ganoderma lucidum. Free. Radic. Biol. Med. 2020, 147, 220–230. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, H.; Ge, F.; Wu, T.; Liu, Y.; Tian, L.; Liu, Y.; Xiang, T.; Yu, H.; Shi, L.; He, Q.; et al. The AMP-Activated Protein Kinase (AMPK) Positively Regulates Lysine Biosynthesis Induced by Citric Acid in Flammulina filiformis. J. Fungi 2023, 9, 340. https://doi.org/10.3390/jof9030340
Fan H, Ge F, Wu T, Liu Y, Tian L, Liu Y, Xiang T, Yu H, Shi L, He Q, et al. The AMP-Activated Protein Kinase (AMPK) Positively Regulates Lysine Biosynthesis Induced by Citric Acid in Flammulina filiformis. Journal of Fungi. 2023; 9(3):340. https://doi.org/10.3390/jof9030340
Chicago/Turabian StyleFan, Hao, Feng Ge, Tao Wu, Yongzhi Liu, Li Tian, Yueqian Liu, Taobo Xiang, Hanshou Yu, Liang Shi, Qin He, and et al. 2023. "The AMP-Activated Protein Kinase (AMPK) Positively Regulates Lysine Biosynthesis Induced by Citric Acid in Flammulina filiformis" Journal of Fungi 9, no. 3: 340. https://doi.org/10.3390/jof9030340