
Comparative Proteomic Analyses within Three Developmental Stages of the Mushroom White Hypsizygus marmoreus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions and Acquisition of the H. marmoreus Samples
2.2. Protein Extraction and Peptide Digestion
2.3. Label-Free LC-MS/MS Quantitative Proteomic Analysis
2.4. Peptide and Protein Identification
2.5. Bioinformatic Analysis
3. Results
3.1. Phenotype of H. marmoreus in Three Developmental Stages from Scratching to the Thirteenth Day after Scratching
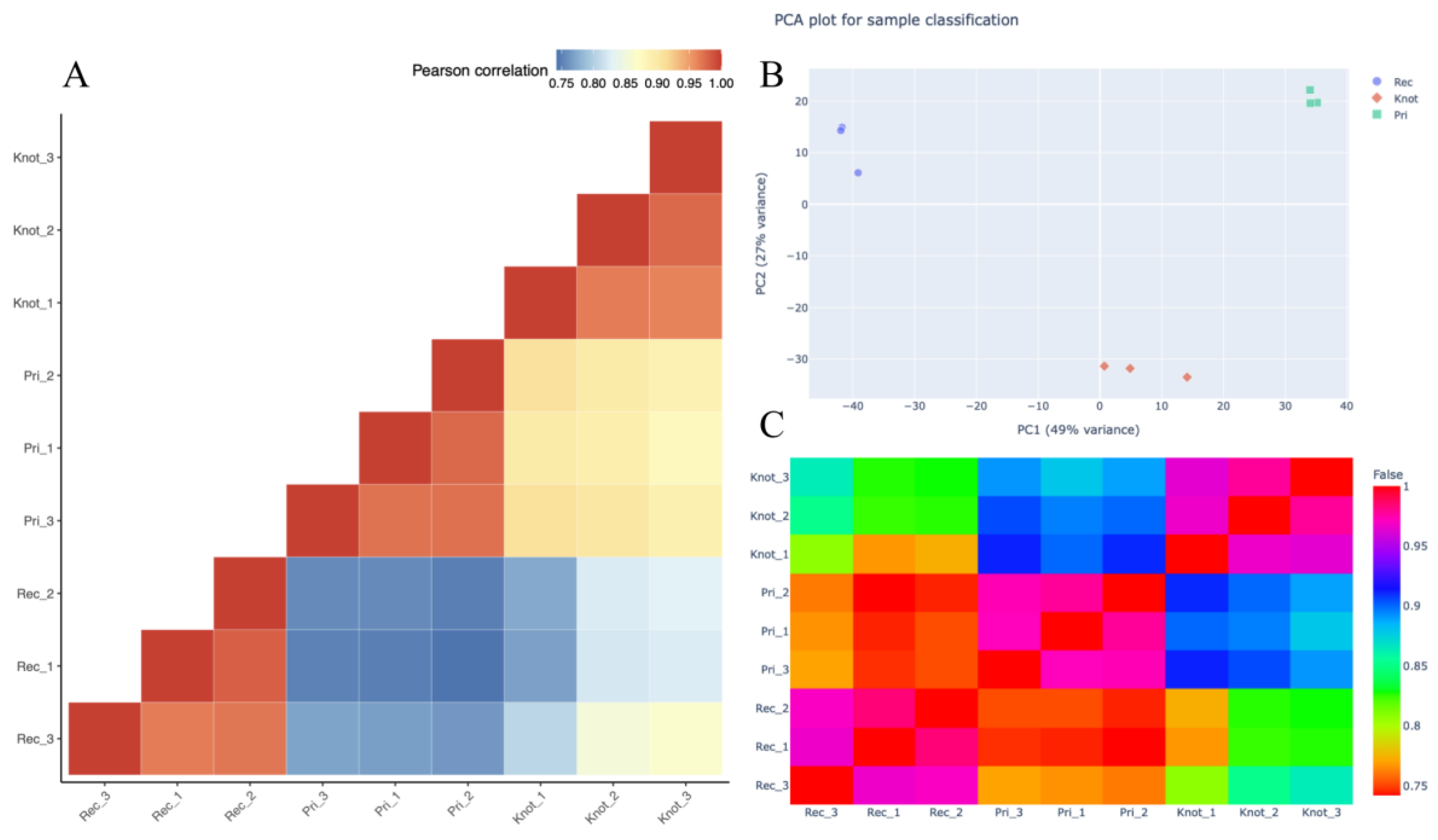
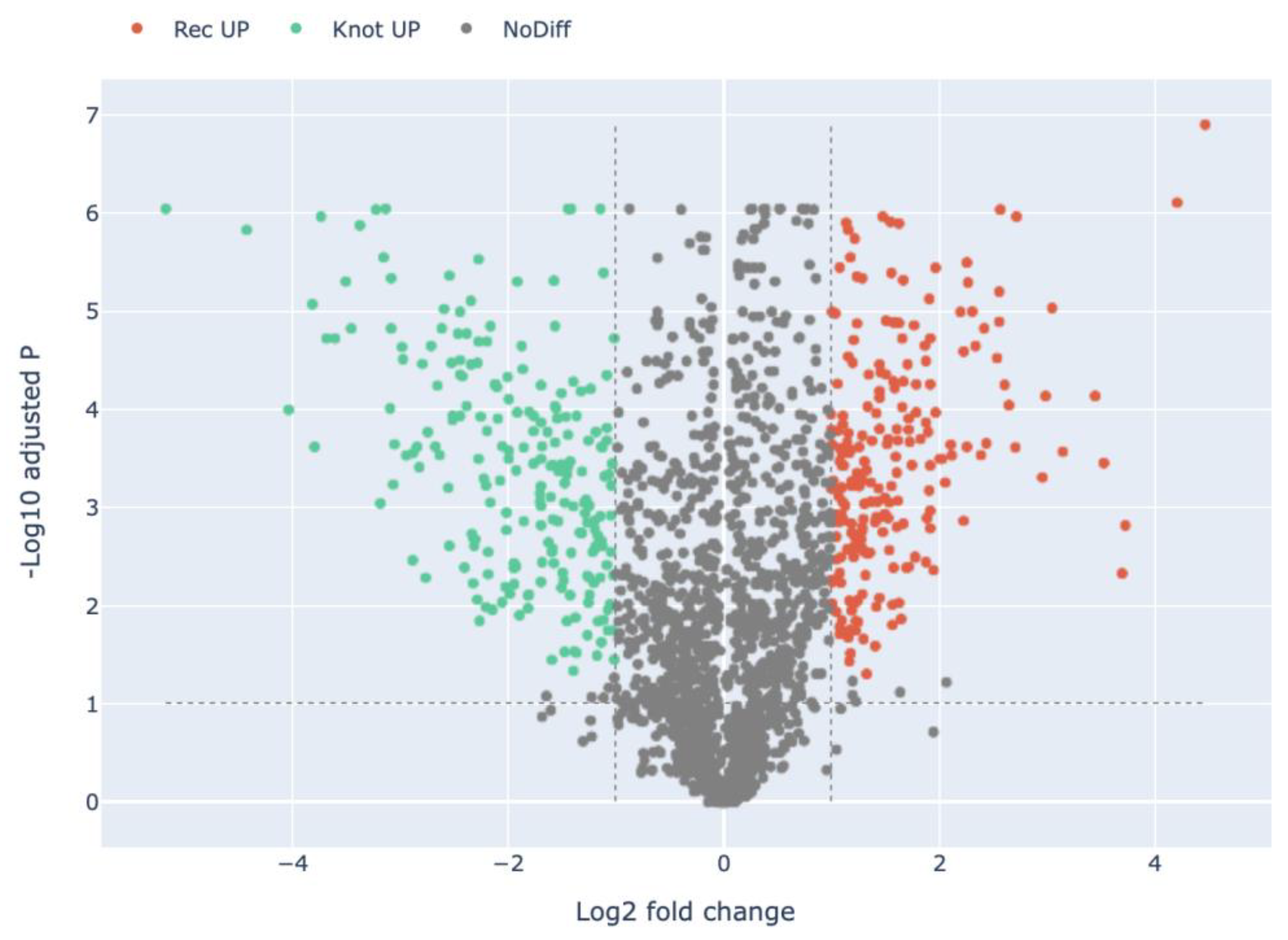
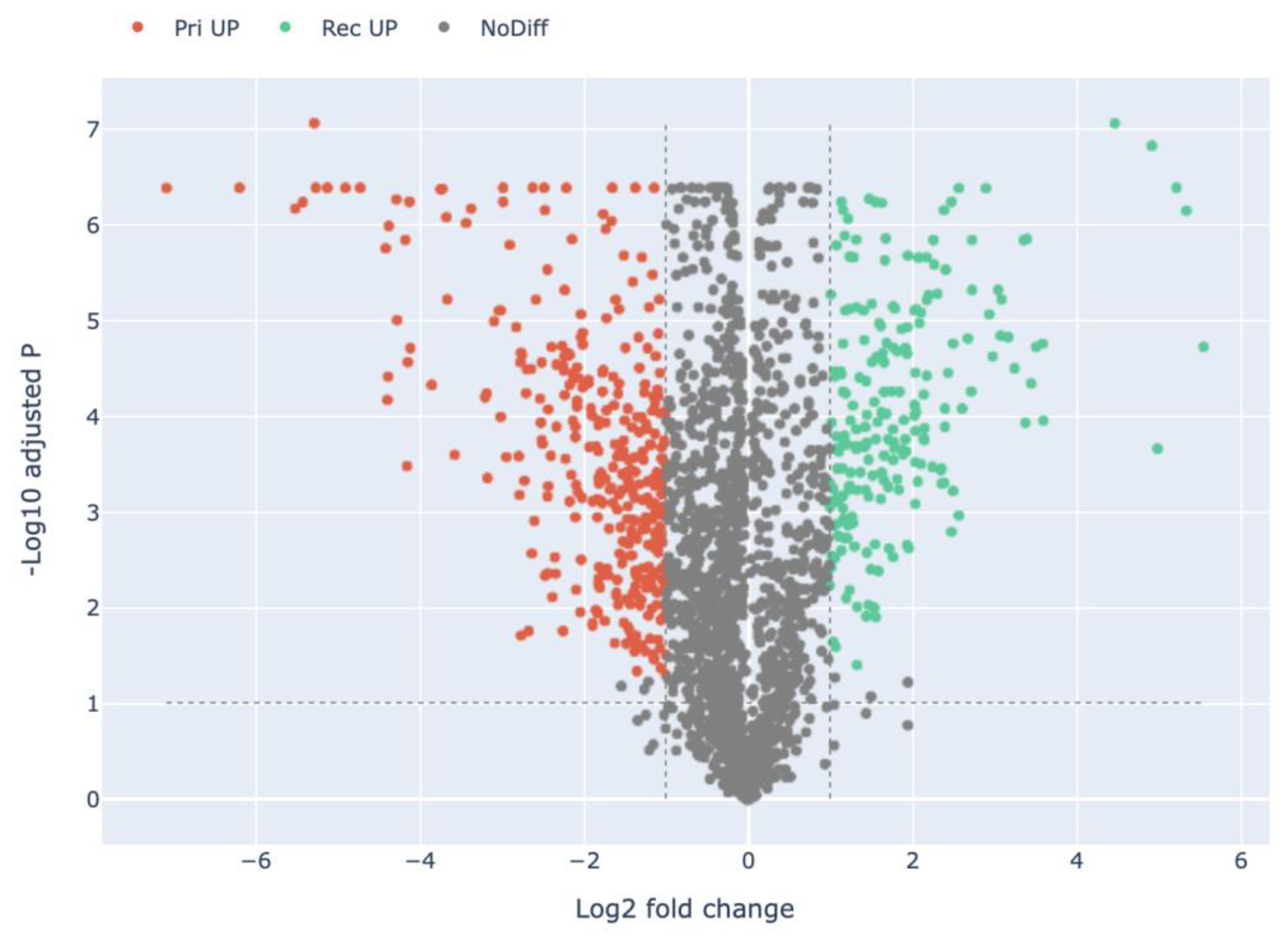
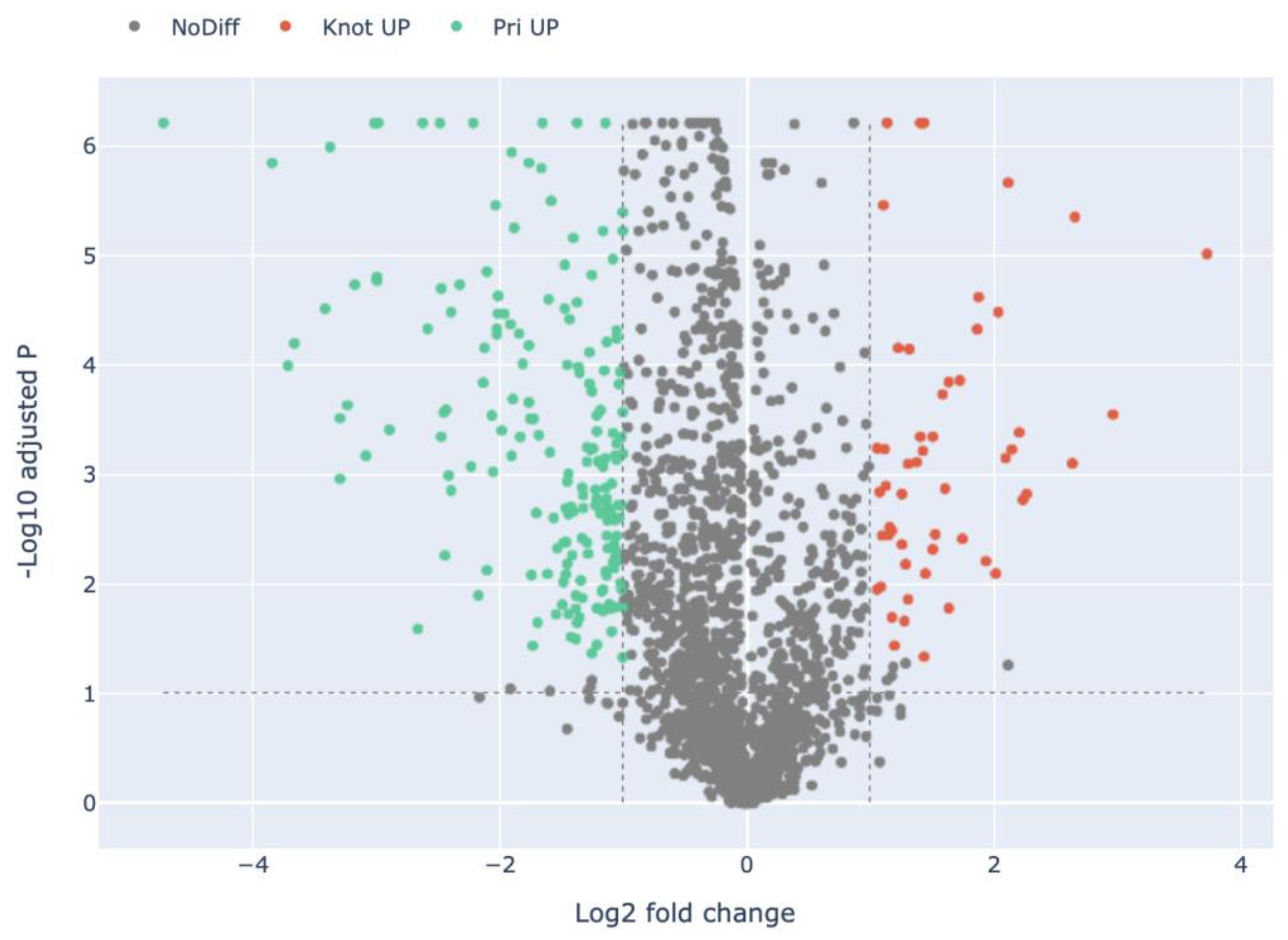
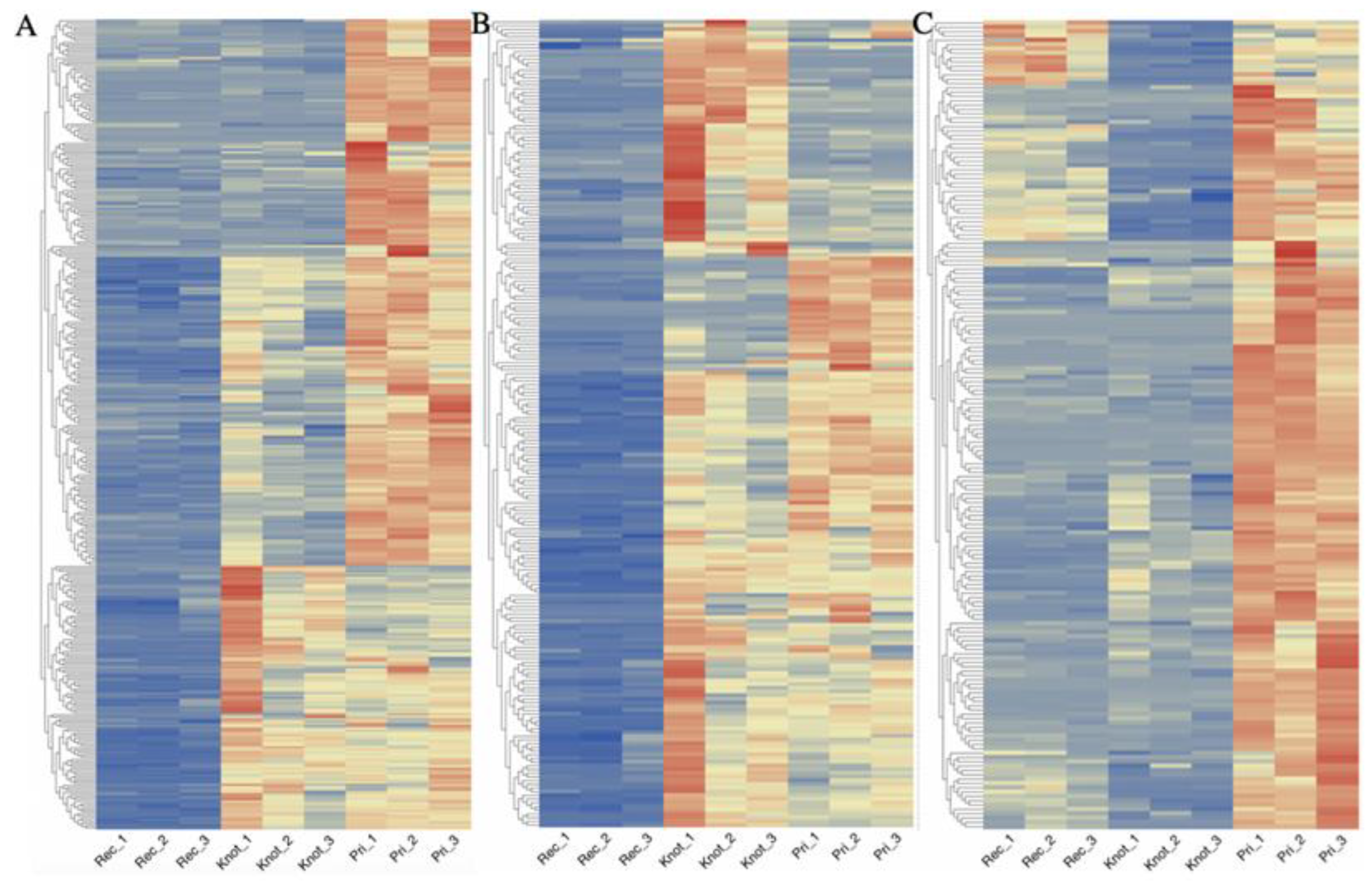
3.2. Quantitative Visualization of the Differential Expression of Proteins of H. marmoreus in Three Developmental Stages
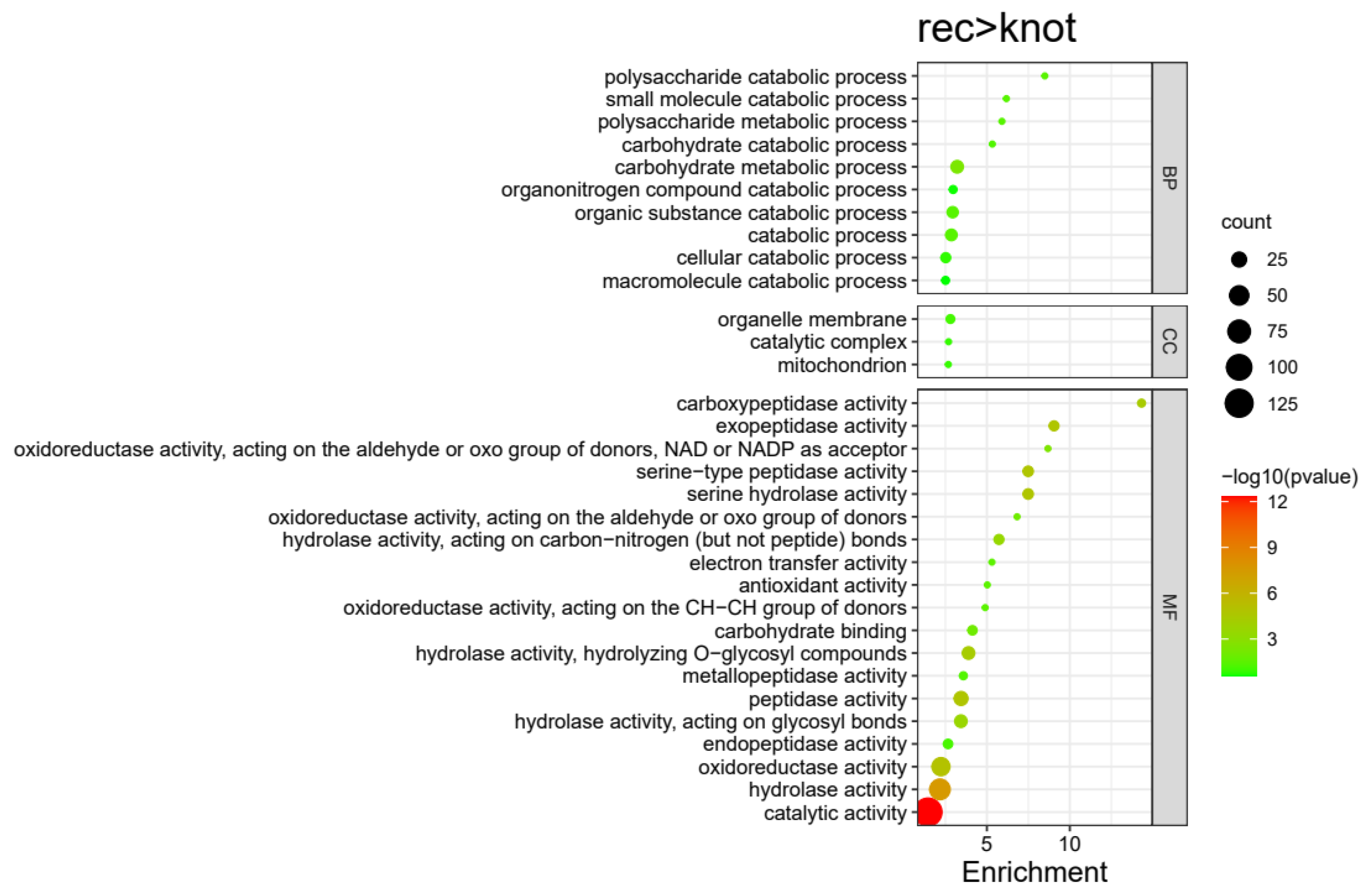
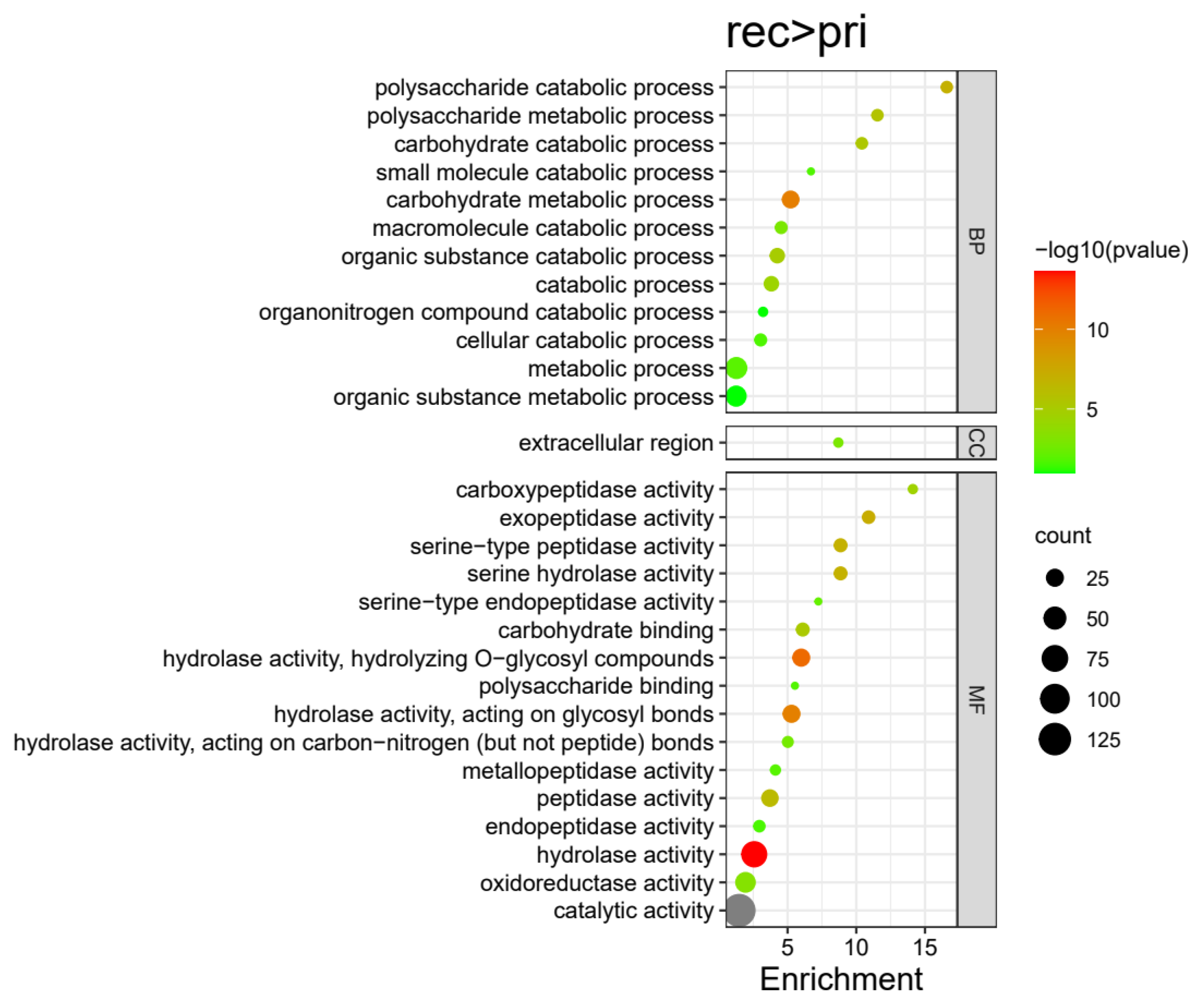
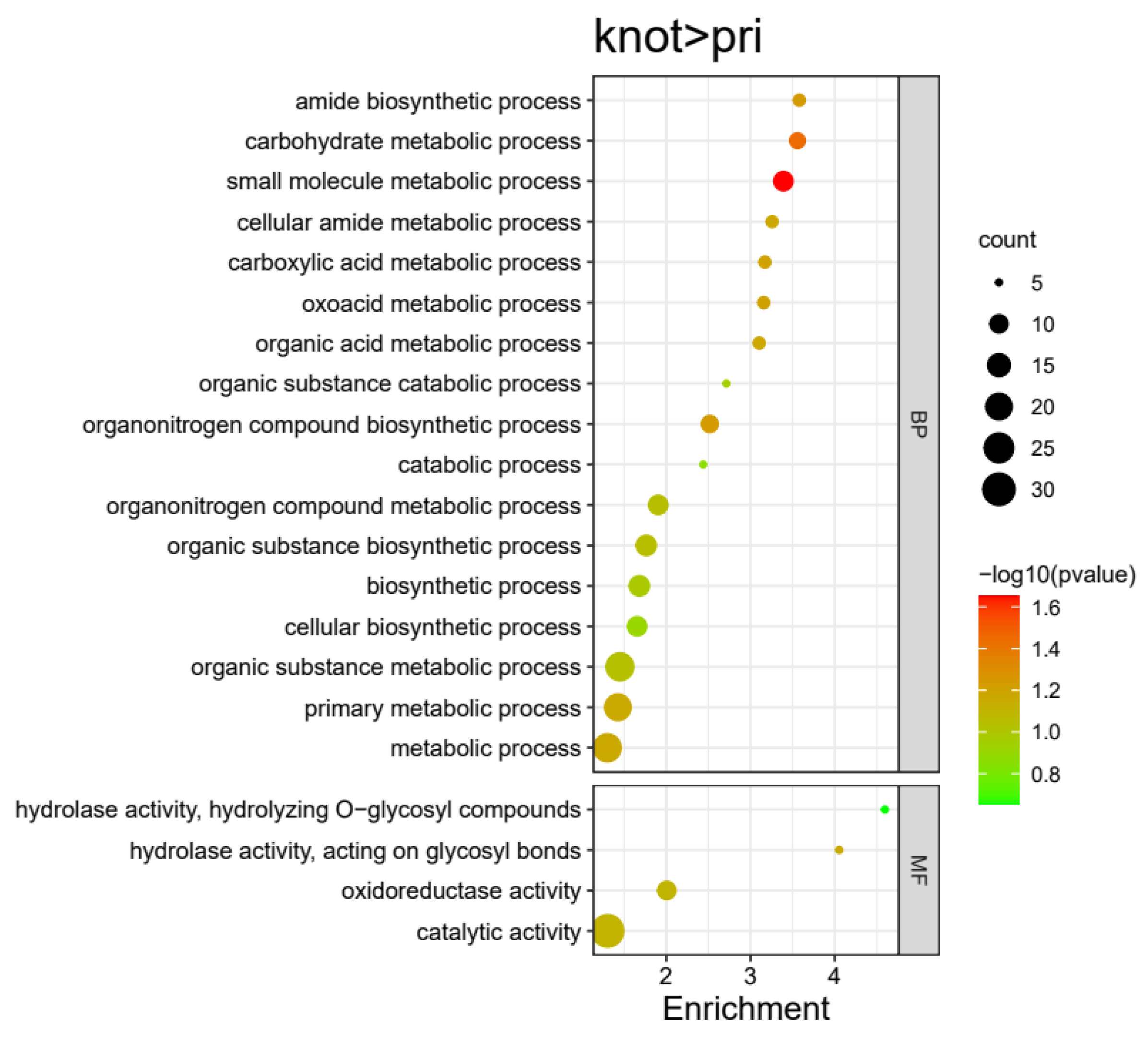
3.3. Gene Ontology (GO) Enrichment Analysis for DEPs
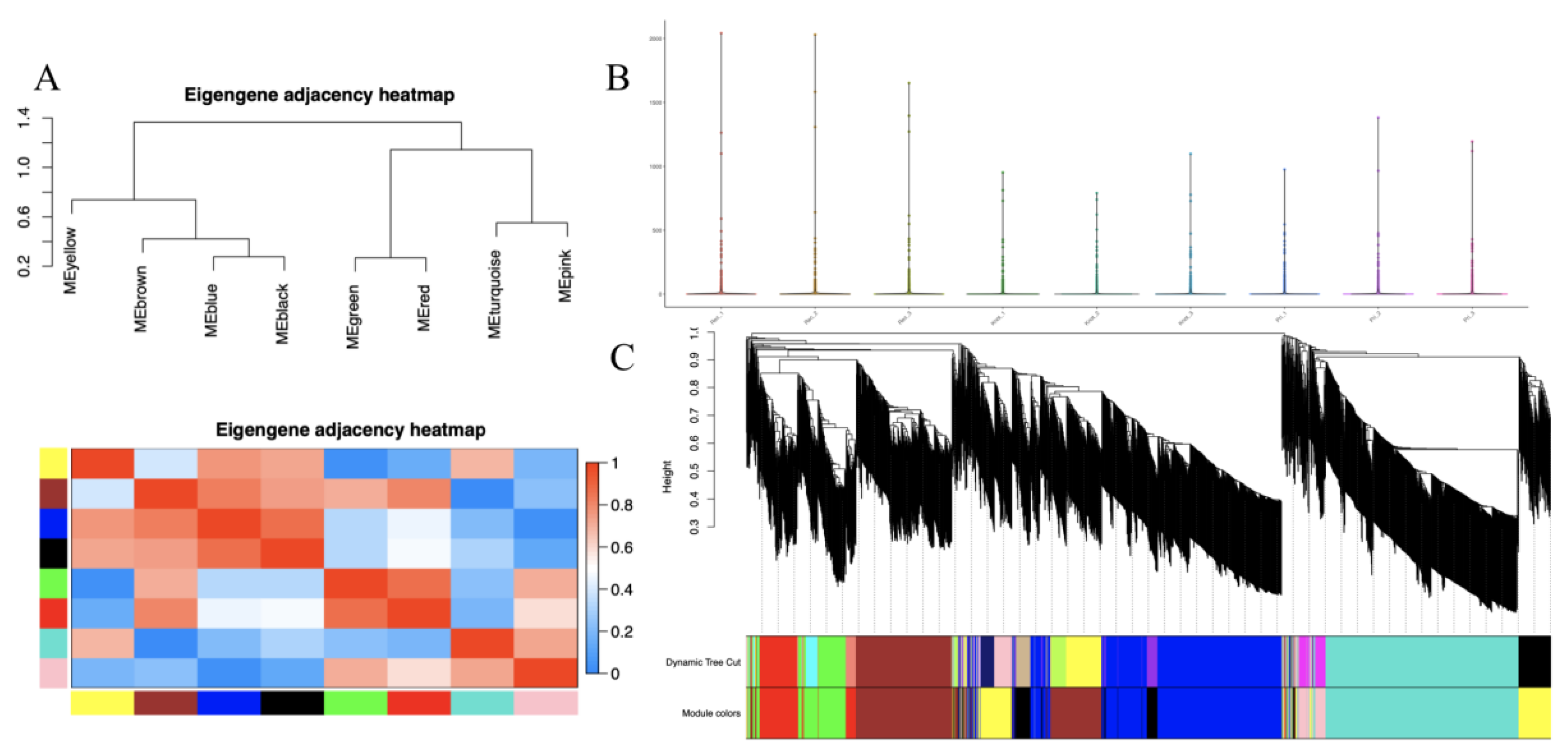
3.4. Weighted Gene Co-Expression Network Analysis (WGCNA) for DEPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Royse, D.J.; Baars, J.; Tan, Q. Current overview of mushroom production in the world. Edible Med. Mushrooms Technol. Appl. 2017, 5–13. [Google Scholar] [CrossRef]
- Kuees, U.; Navarro-Gonzalez, M. How do Agaricomycetes shape their fruiting bodies? 1. Morphological aspects of development. Fungal Biol. Rev. 2015, 29, 63–97. [Google Scholar] [CrossRef]
- Sakamoto, Y. Influences of environmental factors on fruiting body induction, development and maturation in mushroom-forming fungi. Fungal Biol. Rev. 2018, 32, 236–248. [Google Scholar] [CrossRef]
- Jang, K.-Y.; Jhune, C.-S.; Park, J.-S.; Cho, S.-M.; Weon, H.-Y.; Cheong, J.-C.; Choi, S.-G.; Sung, J.-M. Characterization of fruitbody morphology on various environmental conditions in H. marmoreus. Mycobiology 2003, 31, 145–150. [Google Scholar] [CrossRef]
- Scrase, R.J.; Elliott, T.J. Biology and technology of mushroom culture. In Microbiology of Fermented Foods; Springer: Berlin, Germany, 1998; pp. 543–584. [Google Scholar]
- Klengel, T.; Liang, W.-J.; Chaloupka, J.; Ruoff, C.; Schröppel, K.; Naglik, J.R.; Eckert, S.E.; Mogensen, E.G.; Haynes, K.; Tuite, M.F. Fungal adenylyl cyclase integrates CO2 sensing with cAMP signaling and virulence. Curr. Biol. 2005, 15, 2021–2026. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Lin, J.; Shi, Y.; Chang, X.; Wang, Y.; Guo, L.; Wang, W.; Dou, M.; Deng, Y.; Ming, R. Mitochondrial genome in Hypsizygus marmoreus and its evolution in Dikarya. BMC Genom. 2019, 20, 765. [Google Scholar] [CrossRef] [PubMed]
- Son, S.Y.; Park, Y.J.; Jung, E.S.; Singh, D.; Lee, Y.W.; Kim, J.G.; Lee, C.H. Integrated metabolomics and transcriptomics unravel the metabolic pathway variations for different sized beech mushrooms. Int. J. Mol. Sci. 2019, 20, 6007. [Google Scholar] [CrossRef]
- Mlezek, M.; Siwulski, M.; Rzymski, P.; Budka, A.; Kala, P.; Jasińska, A.; Gsecka, M.; Budzyńska, S.; Niedzielski, P. Comparison of elemental composition of mushroom Hypsizygus marmoreus originating from commercial production and experimental cultivation. Sci. Hortic. 2018, 236, 30–35. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, H.; Li, T.; Chen, L.; Liu, R. Comparison of phenolics, antioxidant, and antiproliferative activities of two Hypsizygus marmoreus varieties. J. Food Sci. 2020, 85, 2227–2235. [Google Scholar] [CrossRef]
- Liu, M.; Li, S.; Wang, X.; Zhu, Y.; Zhang, J. Characterization, anti-oxidation and anti-inflammation of polysaccharides by Hypsizygus marmoreus against LPS-induced toxicity on lung. Int. J. Biol. Macromol. 2018, 111, 121–128. [Google Scholar] [CrossRef]
- Oliveira, R.S.; Biscaia SM, P.; Bellan, D.L.; Viana SR, F.; Carbonero, E.R. Structure elucidation of a bioactive fucomannogalactan from the edible mushroom Hypsizygus marmoreus. Carbohydr. Polym. 2019, 225, 115203. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.F.; Ma, C.Y. Effect of a glycoprotein from mushroom Hypsizygus marmoreus (Peck) Bigelow on growth and differentiation of human leukemic U937 cells. Food Chem. Toxicol. 2013, 62, 782–785. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.H.; Tarbasa, M.; Chae, H.M.; Guan, Y.S. Molecular properties of water-unextractable proteoglycans from Hypsizygus marmoreus and their In Vitro immunomodulatory activities. Molecules 2012, 17, 207–226. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.G.; Kim, Y.H.; Bolormaa, Z.; Kim, M.K.; Lee, J.C. Characterization of an antihypertensive angiotensin I-converting enzyme inhibitory peptide from the edible mushroom Hypsizygus marmoreus. BioMed Res. Int. 2013, 2013, 283964. [Google Scholar] [CrossRef] [PubMed]
- Harada, A.; Yoneyama, S.; Doi, S.; Aoyama, M. Changes in contents of free amino acids and soluble carbohydrates during fruit-body development of Hypsizygus marmoreus. Food Chem. 2003, 83, 343–347. [Google Scholar] [CrossRef]
- Terashita, T.; Murao, R.; Yoshikawa, K.; Shishiyama, J. Changes in carbohydrase activities during vegetative growth and development of fruit-bodies of Hypsizygus marmoreus grown in sawdust-based culture. J. Wood Sci. 1998, 44, 234–236. [Google Scholar] [CrossRef]
- Terashita, T.; Nakaie, Y.; Inoue, T.; Yoshikawa, K.; Shishiyama, J. Role of metal proteinases in the fruit-body formation of Hypsizygus marmoreus. J. Wood Sci. 1998, 44, 379–384. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, H.; Chen, M. Cloning and functional analysis of a laccase gene during fruiting body formation in Hypsizygus marmoreus. Microbiol. Res. 2015, 179, 54–63. [Google Scholar] [CrossRef]
- Min, B.; Kim, S.; Oh, Y.L.; Kong, W.S.; Park, H.; Cho, H.; Jang, K.Y.; Kim, J.Y.; Choi, I.G. Genomic discovery of the hypsin gene and biosynthetic pathways for terpenoids in Hypsizygus marmoreus. BMC Genom. 2018, 19, 789. [Google Scholar] [CrossRef]
- Zhang, J.; Ren, A.; Chen, H.; Zhao, M.; Shi, L.; Chen, M.; Wang, H.; Feng, Z.; Minou, N. Transcriptome analysis and its application in identifying genes associated with fruiting body development in basidiomycete Hypsizygus marmoreus. PLoS ONE 2015, 10, e0123025. [Google Scholar] [CrossRef]
- Xu, L.; Guo, L.; Yu, H. Label-free comparative proteomics analysis revealed heat stress responsive mechanism in Hypsizygus marmoreus. Front. Microbiol. 2021, 11, 541967. [Google Scholar] [CrossRef]
- Yang, X.Q.; Lin, R.M.; Xu, K.; Guo, L.Z.; Yu, H. Comparative Proteomic Analysis within the Developmental Stages of the Mushroom White Hypsizygus marmoreus. J. Fungi 2021, 7, 1064. [Google Scholar] [CrossRef] [PubMed]
- Meijueiro, M.L.; Santoyo, F.; Ramírez, L.; Pisabarro, A.G. Transcriptome characteristics of filamentous fungi deduced using high-throughput analytical technologies. Brief Funct. Genom. 2014, 13, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Li, S.; Zhao, S.; Hu, C.; Mao, C.; Guo, L.; Yu, H.L.; Yu, H. Whole genome sequence of an edible mushroom Stropharia rugosoannulata (Daqiugaigu). J. Fungi 2021, 8, 99. [Google Scholar] [CrossRef]
- Riley, R.; Salamov, A.A.; Brown, D.W.; Nagy, L.G.; Floudas, D.; Held, B.W.; Levasseur, A.; Lombard, V.; Morin, E.; Otillar, R.; et al. Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc. Natl. Acad. Sci. USA 2014, 111, 9923–9928. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Van, P.; Jiang, W.; Gottardo, R.; Greg, F. ggCyto: Next generation open-source visualization software for cytometry. Bioinformatics 2018, 34, 3951–3953. [Google Scholar] [CrossRef]
- Perrot, T.; Schwartz, M.; Deroy, A.; Girardet, J.M.; Kohler, A.; Morel-Rouhier, M.; Favier, F.; Gelhaye, E.; Didierjean, C. Diversity of Omega Glutathione Transferases in mushroom-forming fungi revealed by phylogenetic, transcriptomic, biochemical and structural approaches. Fungal Genet Biol. 2021, 148, 103506. [Google Scholar] [CrossRef]
- Wu, D.; Zhu, H.; Chu, J.; Wu, J. N-acetyltransferase co-expression increases α-glucosidase expression level in Pichia pastoris. J. Biotechnol. 2019, 289, 26–30. [Google Scholar] [CrossRef]
- Betancor, D.; Nuñez-Borque, E.; Cuesta-Herranz, J.; Escudero, C.; Freundt, N.; Pastor-Vargas, C.; Ibañez, M.D. Porin: A New Button Mushroom (Agaricus bisporus) Allergen. J. Investig. Allergol. Clin. Immunol. 2020, 30, 135–136. [Google Scholar] [CrossRef]
- Ito, T.; Kobayashi, T.; Egusa, C.; Maeda, T.; Abe, N.; Okubo, Y.; Tsuboi, R.; Niitsuma, T. A case of food allergy due to three different mushroom species. Allergol. Int. 2020, 69, 152–153. [Google Scholar] [CrossRef]
- Zan, X.Y.; Wu, X.H.; Cui, F.J.; Zhu, H.A.; Sun, W.J.; Jiang, L.H.; Tao, T.L.; Zhao, X. UDP-glucose pyrophosphorylase gene affects mycelia growth and polysaccharide synthesis of Grifola frondosa. Int. J. Biol. Macromol. 2020, 161, 1161–1170. [Google Scholar] [CrossRef] [PubMed]
- Kristan, K.; Rižner, T.L. Steroid-transforming enzymes in fungi. J. Steroid Biochem. Mol. Biol. 2012, 129, 79–91. [Google Scholar] [CrossRef]
- Hoff, M.; Trüeb, R.M.; Ballmer-Weber, B.K.; Vieths, S.; Wuethrich, B. Immediate-type hypersensitivity reaction to ingestion of mycoprotein (Quorn) in a patient allergic to molds caused by acidic ribosomal protein P2. J. Allergy Clin. Immunol. 2003, 111, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Sun, L.; Yin, X.; Gao, M.; Zhao, Y.; Jia, P.; Yuan, X.; Fu, Y.; Li, Y. Pleurotus eryngii Genomes Reveal Evolution and Adaptation to the Gobi Desert Environment. Front. Microbiol. 2019, 10, 2024. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Chang, M.C.; Meng, J.L.; Feng, C.P.; Zhao, H.; Zhang, M.L. Comparative Proteome Reveals Metabolic Changes during the Fruiting Process in Flammulina velutipes. J. Agric. Food Chem. 2017, 65, 5091–5100. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Module | Rec_1 | Rec_2 | Rec_3 | Knot_1 | Knot_2 | Knot_3 | Pri_1 | Pri_2 | Pri_3 |
|---|---|---|---|---|---|---|---|---|---|
| MEyellow | 2.80 × 10−1 | 2.47 × 10−1 | 4.44 × 10−2 | −4.33 × 10−1 | −4.68 × 10−1 | −4.76 × 10−1 | 3.42 × 10−1 | 2.19 × 10−1 | 2.44 × 10−1 |
| MEbrown | −4.54 × 10−1 | −4.63 × 10−1 | −4.37 × 10−1 | 3.19 × 10−1 | 1.42 × 10−1 | 7.06 × 10−5 | 2.80 × 10−1 | 3.15 × 10−1 | 2.98 × 10−1 |
| MEblue | −2.34 × 10−1 | −2.41 × 10−1 | −2.74 × 10−1 | −1.81 × 10−1 | −2.26 × 10−1 | −2.49 × 10−1 | 5.12 × 10−1 | 5.00 × 10−1 | 3.93 × 10−1 |
| MEblack | −1.54 × 10−1 | −1.73 × 10−1 | −1.82 × 10−1 | −1.20 × 10−1 | −2.00 × 10−1 | −2.74 × 10−1 | 1.77 × 10−1 | 6.08 × 10−2 | 8.65 × 10−1 |
| MEgreen | −2.79 × 10−1 | −2.91 × 10−1 | −2.65 × 10−1 | 4.07 × 10−1 | 6.03 × 10−1 | 3.72 × 10−1 | −1.75 × 10−1 | −1.97 × 10−1 | −1.74 × 10−1 |
| MEred | −3.04 × 10−1 | −3.13 × 10−1 | −2.60 × 10−1 | 8.28 × 10−1 | 7.81 × 10−2 | 1.78 × 10−1 | −1.09 × 10−1 | −7.61 × 10−2 | −2.28 × 10−2 |
| MEturquoise | 5.18 × 10−1 | 4.85 × 10−1 | 4.03 × 10−1 | −2.39 × 10−1 | −1.99 × 10−1 | −1.99 × 10−1 | −2.53 × 10−1 | −2.69 × 10−1 | −2.48 × 10−1 |
| MEpink | 8.02 × 10−2 | 3.69 × 10−2 | 4.93 × 10−1 | 2.14 × 10−1 | 2.85 × 10−1 | 2.08 × 10−1 | −4.56 × 10−1 | −4.37 × 10−1 | −4.25 × 10−1 |
| MEgrey | −4.77 × 10−1 | 9.62 × 10−2 | 2.91 × 10−1 | −1.02 × 10−1 | −2.19 × 10−1 | 3.37 × 10−1 | 5.75 × 10−1 | −9.32 × 10−2 | −4.08 × 10−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Lin, R.; Li, X.; Zhang, C.; Yang, X.; Guo, L.; Yu, H.; Gao, X.; Hu, C. Comparative Proteomic Analyses within Three Developmental Stages of the Mushroom White Hypsizygus marmoreus. J. Fungi 2023, 9, 225. https://doi.org/10.3390/jof9020225
Xu L, Lin R, Li X, Zhang C, Yang X, Guo L, Yu H, Gao X, Hu C. Comparative Proteomic Analyses within Three Developmental Stages of the Mushroom White Hypsizygus marmoreus. Journal of Fungi. 2023; 9(2):225. https://doi.org/10.3390/jof9020225
Chicago/Turabian StyleXu, Lili, Rongmei Lin, Xiaohang Li, Chenxiao Zhang, Xiuqing Yang, Lizhong Guo, Hao Yu, Xia Gao, and Chunhui Hu. 2023. "Comparative Proteomic Analyses within Three Developmental Stages of the Mushroom White Hypsizygus marmoreus" Journal of Fungi 9, no. 2: 225. https://doi.org/10.3390/jof9020225