
Morphology, Molecular Identification, and Pathogenicity of Two Novel Fusarium Species Associated with Postharvest Fruit Rot of Cucurbits in Northern Thailand

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Fungal Isolation
2.3. Fungal Identification
2.3.1. Morphological Study
2.3.2. DNA Extraction and PCR Amplification and Sequencing
2.3.3. Sequencing
2.3.4. Sequence Alignment and Phylogenetic Analyses
2.4. Pathogenicity Tests
3. Results
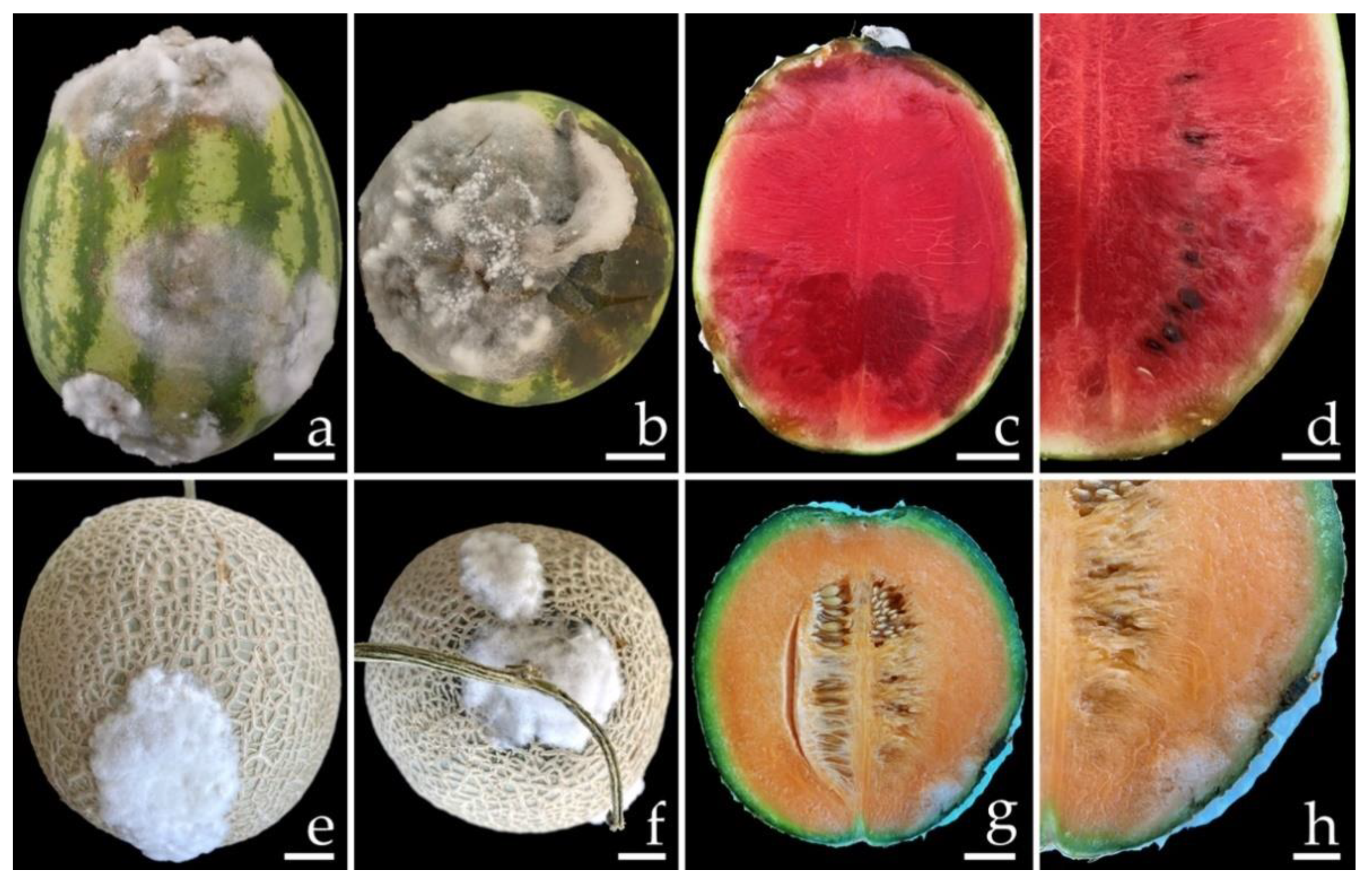
3.1. Sample Collection and Disease Symptoms
3.2. Fungal Isolation
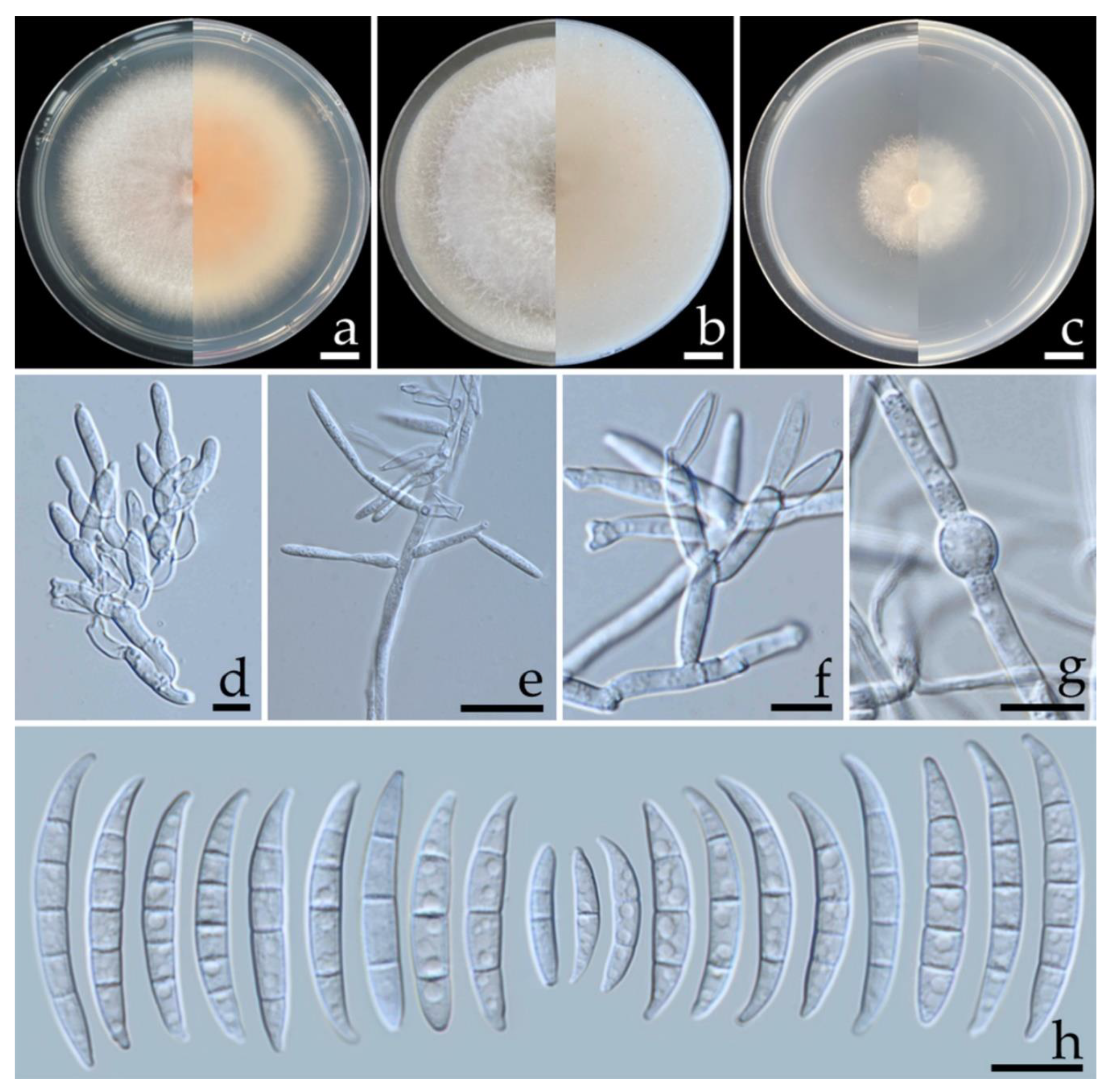
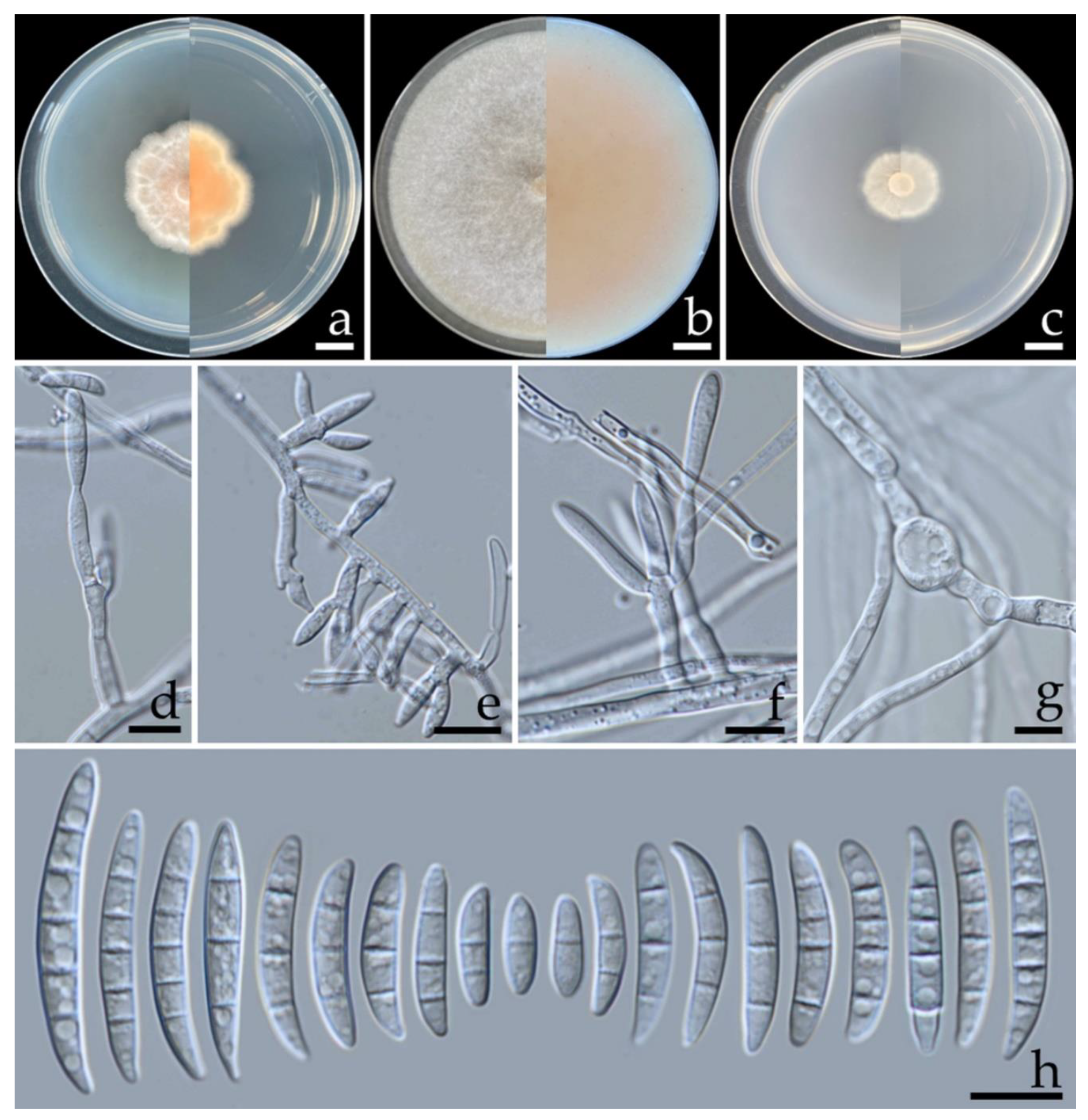
3.3. Morphological Study
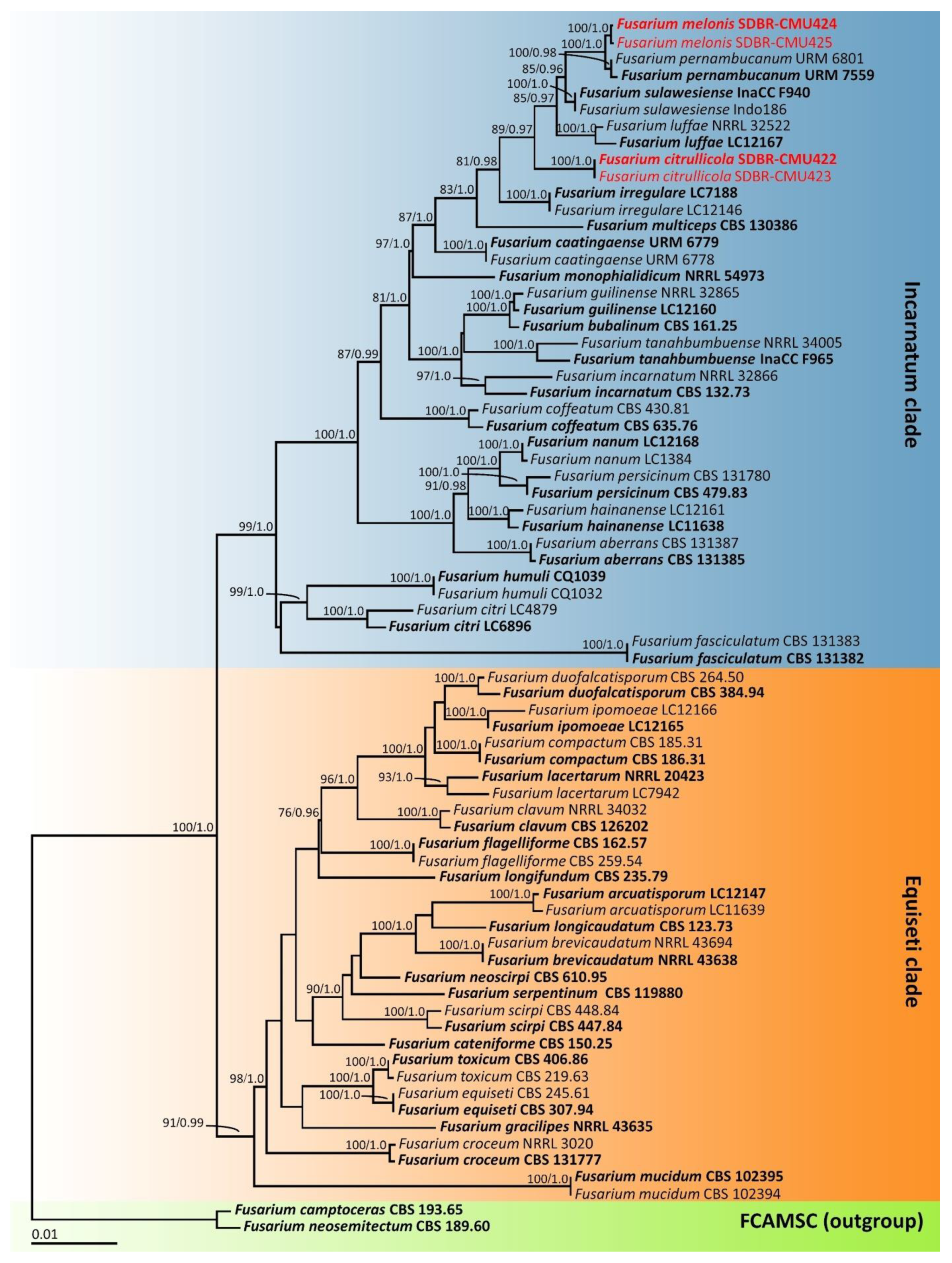
3.4. Phylogenetic Results
3.5. Taxonomic Description


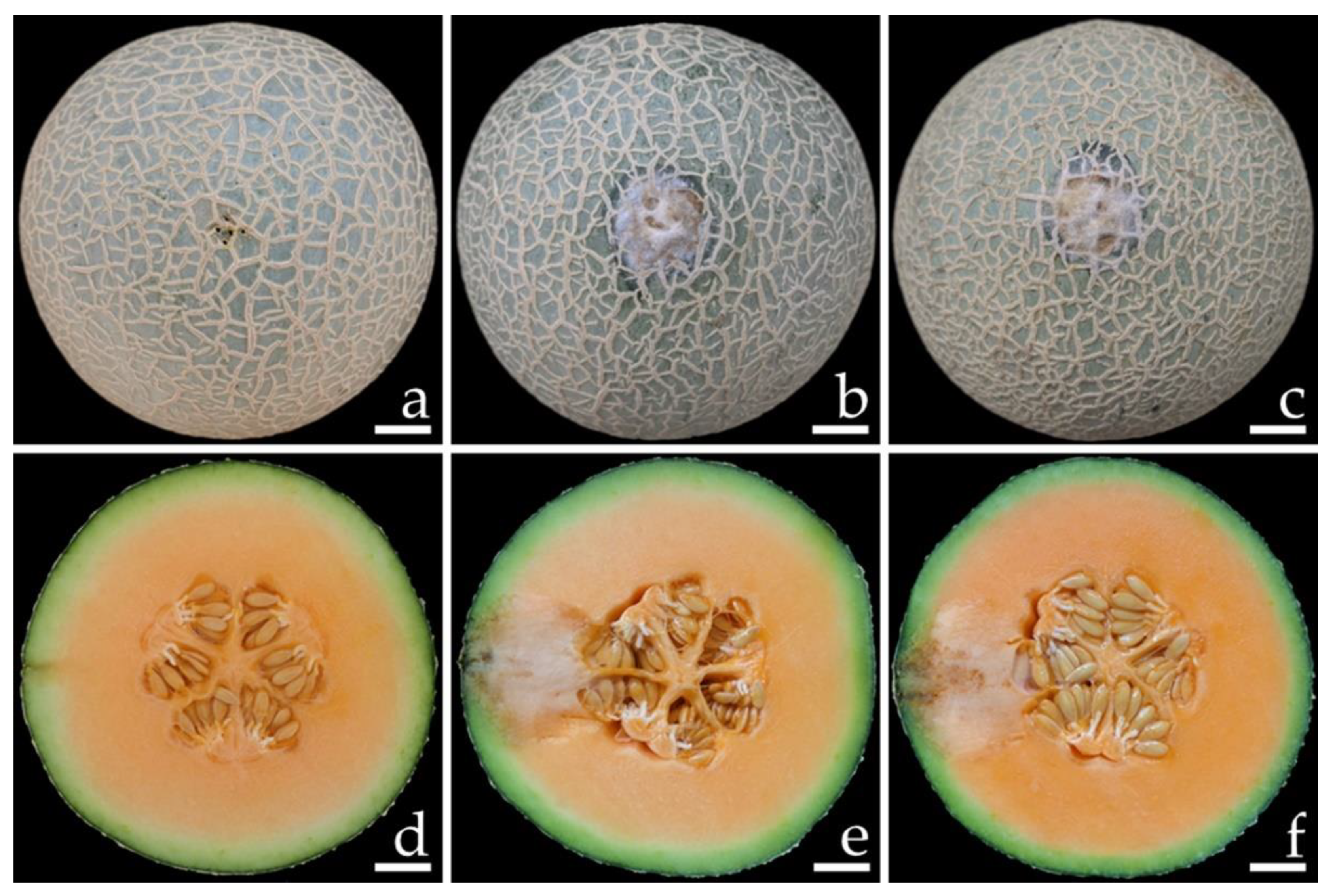
3.6. Pathogenicity Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saediman, H.; Alwi, L.O.; Rianse, I.S.; Taridala, S.A.A.; Salahuddin, S.; Indarsyih, Y.; Astuti, R.W. Comparative profitability of melon and watermelon production in South Konawe District of Southeast Sulawesi. WSEAS Trans. Bus. Econ. 2020, 17, 933–939. [Google Scholar] [CrossRef]
- Assefa, A.D.; Hur, O.S.; Ro, N.Y.; Lee, J.E.; Hwang, A.J.; Kim, B.S.; Rhee, J.H.; Yi, J.Y.; Kim, J.H.; Lee, H.S.; et al. Fruit morphology, citrulline, and arginine levels in diverse watermelon (Citrullus lanatus) germplasm collections. Plants 2020, 9, 1054. [Google Scholar] [CrossRef] [PubMed]
- Kesh, H.; Kaushik, P. Advances in melon (Cucumis melo L.) breeding: An update. Sci. Hortic. 2021, 282, 110045. [Google Scholar] [CrossRef]
- Nuangmek, W.; Aiduang, W.; Suwannarach, N.; Kumla, J.; Kiatsiriroat, T.; Lumyong, S. First report of fruit rot on cantaloupe caused by Fusarium equiseti in Thailand. J. Gen. Plant Pathol. 2019, 85, 295–300. [Google Scholar] [CrossRef]
- Manivannan, A.; Lee, E.S.; Han, K.; Lee, H.E.; Kim, D.S. Versatile nutraceutical potentials of watermelon—A modest fruit loaded with pharmaceutically valuable phytochemicals. Molecules 2020, 25, 5258. [Google Scholar] [CrossRef] [PubMed]
- Perkins-Veazie, P.; Davis, A.; Collins, J.K. Watermelon: From dessert to functional food. Isr. J. Plant Sci. 2013, 60, 395–402. [Google Scholar]
- Lester, G.E.; Hodges, D.M. Antioxidants associated with fruit senescence and human health: Novel orange-fleshed non-netted honey dew melon genotype comparisons following different seasonal productions and cold storage durations. Postharv. Biol. Technol. 2008, 48, 347–354. [Google Scholar] [CrossRef]
- Parle, M.; Singh, K. Musk melon is eat-must melon. Int. Res. J. Pharm. 2011, 2, 52–57. [Google Scholar]
- Maoto, M.M.; Beswa, D.; Jideani, A.I.O. Watermelon as a potential fruit snack. Int. J. Food Prop. 2019, 22, 355–370. [Google Scholar] [CrossRef] [Green Version]
- Vella, F.M.; Cautela, D.; Laratta, B. Characterization of polyphenolic compounds in cantaloupe melon by-products. Foods 2019, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#home (accessed on 23 August 2022).
- Keinath, A.P. From native plants in Central Europe to cultivated crops worldwide: The emergence of Didymella bryoniae as a cucurbit pathogen. HortScience 2011, 46, 532–535. [Google Scholar] [CrossRef] [Green Version]
- Li, P.F.; Ren, R.S.; Yao, X.F.; Xu, J.H.; Babu, B.; Paret, M.L.; Yang, X.P. Identification and characterization of the causal agent of gummy stem blight from muskmelon and watermelon in East China. J. Phytopathol. 2015, 163, 314–319. [Google Scholar] [CrossRef]
- Nuangmek, W.; Aiduang, W.; Suwannarach, N.; Kumla, J.; Lumyong, S. First report of gummy stem blight caused by Stagonosporopsis cucurbitacearum on cantaloupe in Thailand. Can. J. Plant Pathol. 2018, 40, 306–311. [Google Scholar] [CrossRef]
- Babadoost, M.; Zitter, T.A. Fruit rots of pumpkin: A serious threat to the pumpkin industry. Plant Dis. 2009, 93, 772–782. [Google Scholar] [CrossRef] [Green Version]
- Ezrari, S.; Lahlali, R.; Radouane, N.; Tahiri, A.; Lazraq, A. First report of Fusarium equiseti causing pre- and postharvest fruit rot on zucchini in Morocco. J. Plant Pathol. 2020, 102, 251. [Google Scholar] [CrossRef] [Green Version]
- García-Estrada, R.S.; Márquez-Zequera, I.; Tovar-Pedraza, J.M.; Cruz-Lachica, I. First report of cucumber fruit rot caused by Fusarium incarnatum in Mexico. Plant Dis. 2020, 105, 497. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Ahmad, K.; Siddiqui, Y.; Saad, N.; Hun, T.G.; Hata, E.M.; Rashed, O.; Hossain, M.I. First report of Fusarium equiseti, causing fruit rot disease of watermelon in Malaysia. Plant Dis. 2022, 106, 326. [Google Scholar] [CrossRef]
- Wonglom, P.; Sunpapao, A. Fusarium incarnatum is associated with postharvest fruit rot of muskmelon (Cucumis melo). J. Phytopathol. 2020, 168, 204–210. [Google Scholar] [CrossRef]
- Li, Y.G.; Zhang, R.; Meng, L.; Ali, E.; Ji, P.; Zhang, Q.F.; Cui, G.W. Occurrence of fruit rot of cantaloupe caused by Fusarium equiseti in China. Plant Dis. 2019, 103, 2683. [Google Scholar] [CrossRef]
- Lima, E.N.; Oster, A.H.; Bordallo, P.N.; Araújo, A.A.C.; Silva, D.E.M.; Lima, C.S. A novel lineage in the Fusarium incarnatum-equiseti species complex is one of the causal agents of fusarium rot on melon fruits in Northeast Brazil. Plant Pathol. 2021, 70, 133–143. [Google Scholar] [CrossRef]
- Oyedeji, E.O.; Arogundade, O.; Tairu, F.M.; Elum, C.G. Identification and characterization of fungi pathogen causing fruit rot disease of watermelon (Citrullus lanatus). Arch. Phytopathol. Plant Prot. 2022, 55, 344–354. [Google Scholar] [CrossRef]
- Tuttle McGrath, M. Diseases of Cucurbits and their Management. In Diseases of Fruits and Vegetables; Naqvi, S.A.M.H., Ed.; Springer: Dordrecht, The Netherlands, 2004; Volume 1, pp. 455–510. [Google Scholar]
- Li, Y.; Ji, P. First report of fruit rot of watermelon caused by Fusarium equiseti in Georgia in the United States. Plant Dis. 2015, 99, 1272. [Google Scholar] [CrossRef]
- Rivedal, H.M.; Stone, A.G.; Johnson, K.B. First report of Fusarium culmorum causing fruit rot of winter squash (Cucurbita maxima) in Oregon. Plant Dis. 2018, 102, 2659. [Google Scholar] [CrossRef]
- Suwannarach, N.; Khuna, S.; Kumla, J.; Tanruean, K.; Lumyong, S. First report of Lasiodiplodia theobromae causing fruit rot on melon (Cucumis melo) in Thailand. Plant Dis. 2019, 104, 280. [Google Scholar] [CrossRef]
- Wilkinson, K.; Grant, W.P.; Green, L.E.; Hunter, S.; Jeger, M.J.; Lowe, P.; Medley, G.F.; Mills, P.; Phillipson, J.; Poppy, G.M.; et al. Infectious diseases of animals and plants: An interdisciplinary approach. Philos. Trans. R. Soc. B 2011, 366, 1933–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suwannarach, N.; Khuna, S.; Kumla, J.; Cheewangkoon, R.; Suttiprapan, P.; Lumyong, S. Morphology characterization, molecular identification, and pathogenicity of fungal pathogen causing kaffir lime leaf blight in northern Thailand. Plants 2022, 11, 273. [Google Scholar] [CrossRef]
- Choi, Y.W.; Hyde, K.D.; Ho, W.H. Single spore isolation of fungi. Fungal Divers. 1999, 3, 29–38. [Google Scholar]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.-J.; Chaverri, P.; Gené, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Wang, M.M.; Chen, Q.; Diao, Y.Z.; Duan, W.J.; Cai, L. Fusarium incarnatum-equiseti complex from China. Persoonia 2019, 43, 70–89. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.M.; Crous, P.W.; Sandoval-Denis, M.; Han, S.L.; Liu, F.; Liang, J.M.; Duan, W.J.; Cai, L. Fusarium and allied genera from China: Species diversity and distribution. Persoonia 2022, 48, 1–53. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; 252p. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Sarver, B.A.J.; Balajee, S.A.; Schroers, H.-J.; Summerbell, R.C.; Robert, V.A.R.G.; Crous, P.W.; Zhang, N.; et al. Internet-accessible DNA sequence database for identifying fusaria from human and animal infections. J. Clin. Microbiol. 2010, 48, 3708–3718. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. Bioedit Version 6.0.7. 2004. Available online: http://www.mbio.ncsu.edu/bioedit/bioedit.html (accessed on 20 August 2022).
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the cipres science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Manhattan, NY, USA; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131; Institute of Evolutionary 623 Biology, University of Edinburgh: Edinburgh, Scotland, 2019; Available online: http://treebioedacuk/software/figtree/ (accessed on 10 August 2022).
- Xia, J.W.; Sandoval-Denis, M.; Crous, P.W.; Zhang, X.G.; Lombard, L. Numbers to names—Restyling the Fusarium incarnatum-equiseti species complex. Persoonia 2019, 43, 186–221. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Gueidan, C.; Crous, P.W.; Geiser, D.M. Novel multilocus sequence typing scheme reveals high genetic diversity of human pathogenic members of the Fusarium incarnatum-F. equiseti and F. chlamydosporum species complexes within the United States. J. Clin. Microbiol. 2009, 47, 3851–3861. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.C.S.; Trindade, J.V.C.; Lima, C.S.; Barbosa, R.N.; Costa, A.F.; Tiago, P.V.; Oliveira, N.T. Morphology, phylogeny, and sexual stage of Fusarium caatingaense and Fusarium pernambucanum, new species of the Fusarium incarnatum-equiseti species complex associated with insects in Brazil. Mycologia 2019, 111, 244–259. [Google Scholar] [CrossRef]
- Lombard, L.; van Doorn, R.; Crous, P.W. Neotypification of Fusarium chlamydosporum—A reappraisal of a clinically important species complex. Fungal Syst. Evol. 2019, 4, 183–200. [Google Scholar] [CrossRef]
- Maryani, N.; Sandoval-Denis, M.; Lombard, L.; Crous, P.W.; Kema, G.H.J. New endemic Fusarium species hitch-hiking with pathogenic Fusarium strains causing Panama disease in small-holder banana plots in Indonesia. Persoonia 2019, 43, 48–69. [Google Scholar] [CrossRef]
- De Oliveira, M.J.; Laranjeira, D.; Câmara, M.P.S.; Laranjeira, F.F.; Armengol, J.; Michereff, S.J. Effects of wounding, humidity, temperature, and inoculum concentrations on the severity of corky dry rot caused by Fusarium semitectum in melon fruits. Acta Sci. Agron. 2014, 36, 281–289. [Google Scholar] [CrossRef]
- Safari, Z.S.; Ding, P.; Nakasha, J.J.; Yuso, S.F. Combining chitosan and vanillin to retain postharvest quality of tomato fruit during ambient temperature storage. Coatings 2020, 10, 1222. [Google Scholar] [CrossRef]
- Bika, R.; Baysal-Gurel, F. Identification of Fusarium commune, the causal agent of postharvest zinnia meltdown disease in Tennessee. HortTechnology 2021, 31, 432–439. [Google Scholar] [CrossRef]
- Yi, R.H.; Lian, T.; Su, J.J.; Chen, J. First report of internal black rot on Carica papaya fruit caused by Fusarium sulawesiense in China. Plant Dis. 2022, 106, 319. [Google Scholar] [CrossRef]
- Zhang, X.P.; Xia, J.W.; Liu, J.K.; Zhao, D.; Kong, L.G.; Zhu, X.P. First report of Fusarium pernambucanum causing fruit rot of muskmelon in China. Plant Dis. 2022, 106, 1997. [Google Scholar] [CrossRef]
- Araújo, M.B.; Moreira, G.M.; Nascimento, L.V.; Nogueira, G.A.; Nascimento, S.R.C.; Pfenning, L.H.; Ambrósio, M.M.Q. Fusarium rot of melon is caused by several Fusarium species. Plant Pathol. 2021, 70, 712–721. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, Y.; Li, Q.; Huang, S.; Tang, L.; Chen, X.; Guo, T.; Mo, J.; Ma, L. First report of leaf blight caused by Fusarium pernambucanum and Fusarium sulawesiense on plum in Sichuan, China. Plant Dis. 2022, 106, 2759. [Google Scholar] [CrossRef]
- Laraba, I.; McCormick, S.P.; Vaughan, M.M.; Geiser, D.M.; O’Donnell, K. Phylogenetic diversity, trichothecene potential, and pathogenicity within Fusarium sambucinum species complex. PLoS ONE 2021, 16, e0245037. [Google Scholar]
- Pavlou, G.C.; Vakalounakis, D.J.; Ligoxigakis, E.K. Control of root and stem rot of cucumber, caused by Fusarium oxysporum f. sp. radicis-cucumerinum, by grafting onto resistant rootstocks. Plant Dis. 2002, 86, 379–382. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, V.; Veena, K.H.; Jain, S.; Tripathi, M.; Aggarwal, R.; Singh, A.K. First report of seedling blight caused by Fusarium solani on cucumber from India. J. Plant Pathol. 2016, 98, 677–697. [Google Scholar]
- Gao, X.; Wang, Y.; Liu, Y.; Zhang, M.; Zhang, W.; Li, Y. First report of leaf spot on cucumber caused by Fusarium incarnatum in China. Plant Dis. 2020, 104, 973. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Ames, I.A., Ed.; Blackwell Publishing Professional: New York, NY, USA, 2006; pp. 8–240. [Google Scholar]
- Rahjoo, V.; Zad, J.; Javan-Nikkhah, M.; Mirzadi Gohari, A.; Okhovvat, S.M.; Bihamta, M.R.; Razzaghian, J.; Klemsdal, S.S. Morphological and molecular identification of Fusarium isolated from maize ears in Iran. J. Plant Pathol. 2008, 90, 463–468. [Google Scholar]
- Geiser, D.M.; Jiménez-Gasco, M.M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O’Donnell, K. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Nitschke, E.; Nihlgard, M.; Varrelmann, M. Differentiation of eleven Fusarium spp. isolated from sugar beet, using restriction fragment analysis of a polymerase chain reaction-amplified translation elongation factor 1α gene fragment. Phytopathology 2009, 99, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Jedidi, I.; Jurado, M.; Cruz, A.; Trabelsi, M.M.; Said, S.; González-Jaén, M.T. Phylogenetic analysis and growth profiles of Fusarium incarnatum-equiseti species complex strains isolated from Tunisian cereals. Int. J. Food Microbiol. 2021, 353, 109297. [Google Scholar] [CrossRef]
- O’Donnell, K.; Ward, T.J.; Robert, V.A.R.G.; Crous, P.W.; Geiser, D.M.; Kang, S. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 2015, 43, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Balajee, S.A.; Borman, A.M.; Brandt, M.E.; Cano, J.; Cuenca-Estrella, M.; Dannaoui, E.; Guarro, J.; Haase, G.; Kibbler, C.C.; Meyer, W.; et al. Sequence-based identification of Aspergillus, Fusarium, and Mucorales species in the clinical mycology laboratory: Where are we and where should we go from here? J. Clin. Microbiol. 2009, 47, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Akram, W.; Ahmad, A.; Luo, W.; Yasin, N.A.; Wu, T.; Guo, J.; Wang, Q.; Li, G. First report of stem and root rot of Chinese kale caused by Fusarium incarnatum-equiseti species complex in China. Plant Dis. 2019, 103, 1781. [Google Scholar] [CrossRef]
- Ismail, S.I.; Noor Asha, N.A.; Zulperi, D. First report of Fusarium incarnatum-equiseti species complex causing leaf spot on rockmelon (Cucumis melo) in Malaysia. Plant Dis. 2021, 105, 1197. [Google Scholar] [CrossRef]
- Villani, A.; Moretti, A.; Saeger, S.D.; Han, Z.; Mavungu, J.D.D.; Soares, C.M.G.; Proctor, R.H.; Venâncio, A.; Lima, N.; Stea, G.; et al. A polyphasic approach for characterization of a collection of cereal isolates of the Fusarium incarnatum-equiseti species complex. Int. J. Food Microbiol. 2016, 234, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic am-biguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Li, Y.G.; Song, X.L.; Wang, X.Q.; Zhang, H.; Tian, S.; Ji, P. First report of fruit rot of watermelon caused by Fusarium equiseti in China. Plant Dis. 2018, 102, 1852. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, H.J. Fusarium fruit rot of posthavest oriental melon (Cucumis melo L. var. makuwa Mak.) caused by Fusarium spp. Res. Plant Dis. 2004, 10, 260–267. [Google Scholar] [CrossRef]
- Ikediugwu, F.E.O.; Ogieva, W.O. Fruit rot of Citrullus lanatus in Nigeria caused by Fusarium solani. Trans. Br. Mycol. Soc. 1978, 71, 209–213. [Google Scholar] [CrossRef]
- Rampersad, S.N. First report of Fusarium solani fruit rot of pumpkin (Cucurbita pepo) in Trinidad. Plant Dis. 2009, 93, 547. [Google Scholar] [CrossRef]
- González, V.; Armengol, J.; Garcés-Claver, A. First report of Fusarium petroliphilum causing fruit rot of butternut squash in Spain. Plant Dis. 2018, 102, 1662. [Google Scholar] [CrossRef]
- Li, Y.G.; Jiang, W.Y.; Jiang, D.; Wang, R.T.; Tian, S.; Ji, P.; Jiang, B.W. First report of fruit rot on postharvest pumpkin caused by Fusarium acuminatum in China. Plant Dis. 2019, 103, 1035. [Google Scholar] [CrossRef]
- Hao, F.; Zang, Q.; Ding, W.; Ma, E.; Huang, Y.; Wang, Y. First report of fruit rot of melon caused by Fusarium asiaticum in China. Plant Dis. 2021, 105, 1225. [Google Scholar] [CrossRef]
- Parra, M.Á.; Gómez, J.; Aguilar, F.W.; Martínez, J.A. Fusarium annulatum causes Fusarium rot of cantaloupe melons in Spain. Phytopathol. Mediterr. 2022, 61, 269–277. [Google Scholar] [CrossRef]
- Nuangmek, W.; Aiduang, W.; Kumla, J.; Lumyong, S.; Suwannarach, N. Evaluation of a newly identified endophytic fungus, Trichoderma phayaoense for plant growth promotion and biological control of gummy stem blight and wilt of muskmelon. Front. Microbiol. 2021, 12, 634772. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Primer Sequence | The Obtained Length (bp) | |||
|---|---|---|---|---|---|---|
| SDBR-CMU422 | SDBR-CMU423 | SDBR-CMU424 | SDBR-CMU425 | |||
| tef-1 | EF1 EF2 | 5′-ATGGGTAAGGARGACAAGAC-3′ 5′-GGARGTACCAGTSATCATG-3′ | 692 | 691 | 691 | 686 |
| cam | CAL-228F CAL-2Rd | 5′-GAGTTCAAGGAGGCCTTCTCCC-3′ 5′-TGRTCNGCCTCDCGGATCATCTC-3′ | 606 | 603 | 601 | 597 |
| rpb2 | RPB2-5F2 RPB2-7cR | 5′-GGGGWGAYCAGAAGAAGGC-3′ 5′-CCCATRGCTTGYTTRCCCAT-3′ | 1152 | 1148 | 1141 | 1131 |
| Fungal Taxa | Strain/Isolate | GenBank Accession Number | Reference | ||
|---|---|---|---|---|---|
| tef-1 | cam | rpb2 | |||
| Fusarium aberrans | CBS 131385 T | MN170445 | MN170311 | MN170378 | [45] |
| F. aberrans | CBS 131387 | MN170446 | MN170312 | MN170379 | [45] |
| F. arcuatisporum | LC12147 T | MK289584 | MK289697 | MK289739 | [31] |
| F. arcuatisporum | LC11639 | MK289586 | MK289658 | MK289736 | [31] |
| F. brevicaudatum | NRRL 43638 T | GQ505665 | GQ505576 | GQ505843 | [46] |
| F. brevicaudatum | NRRL 43694 | GQ505668 | GQ505579 | GQ505846 | [46] |
| F. bubalinum | CBS 161.25 T | MN170448 | MN170314 | MN170381 | [45] |
| F. caatingaense | URM 6779 T | LS398466 | – | LS398495 | [47] |
| F. caatingaense | URM 6778 | LS398465 | – | LS398494 | [47] |
| F. cateniforme | CBS 150.25 T | MN170451 | MN170317 | MN170384 | [45] |
| F. citri | LC6896 T | MK289617 | MK289668 | MK289771 | [31] |
| F. citri | LC4879 | MK289615 | MK289665 | MK289768 | [31] |
| F. citrullicola | SDBR-CMU422 T | OP020920 | OP020924 | OP020928 | This study |
| F. citrullicola | SDBR-CMU423 | OP020921 | OP020925 | OP020929 | This study |
| F. clavum | CBS 126202 T | MN170456 | MN170322 | MN170389 | [45] |
| F. clavum | NRRL 34032 | GQ505635 | GQ505547 | GQ505813 | [46] |
| F. coffeatum | CBS 635.76 T | MN120755 | MN120696 | MN120736 | [48] |
| F. coffeatum | CBS 430.81 | MN120756 | MN120697 | MN120737 | [48] |
| F. compactum | CBS 186.31 ET | GQ505648 | GQ505560 | GQ505826 | [46] |
| F. compactum | CBS 185.31 | GQ505646 | GQ505558 | GQ505824 | [46] |
| F. croceum | CBS 131777 T | MN170463 | MN170329 | MN170396 | [45] |
| F. croceum | NRRL 3020 | GQ505586 | GQ505498 | GQ505764 | [46] |
| F. duofalcatisporum | CBS 384.94 T | GQ505652 | GQ505564 | GQ505830 | [46] |
| F. duofalcatisporum | CBS 264.50 | GQ505651 | GQ505563 | GQ505829 | [46] |
| F. equiseti | CBS 307.94 NT | GQ505599 | GQ505511 | GQ505777 | [46] |
| F. equiseti | CBS 245.61 | GQ505594 | GQ505506 | GQ505772 | [46] |
| F. fasciculatum | CBS 131382 T | MN170473 | MN170339 | MN170406 | [45] |
| F. fasciculatum | CBS 131383 | MN170474 | MN170340 | MN170407 | [45] |
| F. flagelliforme | CBS 162.57 T | GQ505645 | GQ505557 | GQ505823 | [46] |
| F. flagelliforme | CBS 259.54 | GQ505650 | GQ505562 | GQ505828 | [46] |
| F. gracilipes | NRRL 43635 T | GQ505662 | GQ505573 | GQ505840 | [46] |
| F. guilinense | LC12160 T | MK289594 | MK289652 | MK289747 | [31] |
| F. guilinense | NRRL 32865 | GQ505614 | GQ505526 | GQ505792 | [46] |
| F. hainanense | LC11638 T | MK289581 | MK289657 | MK289735 | [31] |
| F. hainanense | LC12161 | MK289595 | MK289648 | MK289748 | [31] |
| F. humuli | CQ1039 T | MK289570 | MK289712 | MK289724 | [31] |
| F. humuli | CQ1032 | MK289568 | MK289710 | MK289722 | [31] |
| F. incarnatum | CBS 132.73 NT | MN170476 | MN170342 | MN170409 | [45] |
| F. incarnatum | NRRL 32866 | GQ505615 | GQ505527 | GQ505793 | [46] |
| F. ipomoeae | LC12165 T | MK289599 | MK289704 | MK289752 | [31] |
| F. ipomoeae | LC12166 | MK289600 | MK289706 | MK289753 | [31] |
| F. irregulare | LC7188 T | MK289629 | MK289680 | MK289783 | [31] |
| F. irregulare | LC12146 | MK289583 | MK289682 | MK289738 | [31] |
| F. lacertarum | NRRL 20423 T | GQ505593 | GQ505505 | GQ505771 | [46] |
| F. lacertarum | LC7942 | MK289643 | MK289696 | MK289797 | [31] |
| F. longicaudatum | CBS 123.73 T | MN170481 | MN170347 | MN170414 | [45] |
| F. longifundum | CBS 235.79 T | GQ505649 | GQ505561 | GQ505827 | [46] |
| F. luffae | LC12167 T | MK289601 | MK289698 | MK289754 | [31] |
| F. luffae | NRRL 32522 | GQ505612 | GQ505524 | GQ505790 | [46] |
| F. melonis | SDBR-CMU424 T | OP020922 | OP020926 | OP020930 | This study |
| F. melonis | SDBR-CMU425 | OP020923 | OP020927 | OP020931 | This study |
| F. monophialidicum | NRRL 54973 T | MN170483 | MN170349 | MN170416 | [45] |
| F. mucidum | CBS 102395 T | MN170485 | MN170351 | MN170418 | [45] |
| F. mucidum | CBS 102394 | MN170484 | MN170350 | MN170417 | [45] |
| F. multiceps | CBS 130386 T | GQ505666 | GQ505577 | GQ505844 | [46] |
| F. nanum | LC12168 T | MK289602 | MK289651 | MK289755 | [31] |
| F. nanum | LC1384 | MK289611 | MK289661 | MK289764 | [31] |
| F. neoscirpi | CBS 610.95 T | GQ505601 | GQ505513 | GQ505779 | [46] |
| F. pernambucanum | URM 7559 T | LS398489 | – | LS398519 | [47] |
| F. pernambucanum | URM 6801 | LS398483 | – | LS398513 | [47] |
| F. persicinum | CBS 479.83 T | MN170495 | MN170361 | MN170428 | [45] |
| F. persicinum | CBS 131780 | MN170496 | MN170362 | MN170429 | [45] |
| F. scirpi | CBS 447.84 NT | GQ505654 | GQ505566 | GQ505832 | [46] |
| F. scirpi | CBS 448.84 | GQ505592 | GQ505504 | GQ505770 | [46] |
| F. serpentinum | CBS 119880 T | MN170499 | MN170365 | MN170432 | [45] |
| F. sulawesiense | InaCC F940 T | LS479443 | LS479422 | LS479855 | [49] |
| F. sulawesiense | Indo186 | LS479449 | LS479426 | LS479864 | [49] |
| F. tanahbumbuense | InaCC F965 T | LS479448 | LS479432 | LS479863 | [49] |
| F. tanahbumbuense | NRRL 34005 | GQ505629 | GQ505541 | GQ505807 | [46] |
| F. toxicum | CBS 406.86 T | MN170508 | MN170374 | MN170441 | [45] |
| F. toxicum | CBS 219.63 | MN170507 | MN170373 | MN170440 | [45] |
| F. camptoceras | CBS 193.65 ET | MN170450 | MN170316 | MN170383 | [45] |
| F. neosemitectum | CBS 189.60 T | MN170489 | MN170355 | MN170422 | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khuna, S.; Kumla, J.; Thitla, T.; Nuangmek, W.; Lumyong, S.; Suwannarach, N. Morphology, Molecular Identification, and Pathogenicity of Two Novel Fusarium Species Associated with Postharvest Fruit Rot of Cucurbits in Northern Thailand. J. Fungi 2022, 8, 1135. https://doi.org/10.3390/jof8111135
Khuna S, Kumla J, Thitla T, Nuangmek W, Lumyong S, Suwannarach N. Morphology, Molecular Identification, and Pathogenicity of Two Novel Fusarium Species Associated with Postharvest Fruit Rot of Cucurbits in Northern Thailand. Journal of Fungi. 2022; 8(11):1135. https://doi.org/10.3390/jof8111135
Chicago/Turabian StyleKhuna, Surapong, Jaturong Kumla, Tanapol Thitla, Wipornpan Nuangmek, Saisamorn Lumyong, and Nakarin Suwannarach. 2022. "Morphology, Molecular Identification, and Pathogenicity of Two Novel Fusarium Species Associated with Postharvest Fruit Rot of Cucurbits in Northern Thailand" Journal of Fungi 8, no. 11: 1135. https://doi.org/10.3390/jof8111135