
Pan-Echinocandin Resistant C. parapsilosis Harboring an F652S Fks1 Alteration in a Patient with Prolonged Echinocandin Therapy
 , , , ,
, , , , 
Abstract
:1. Introduction
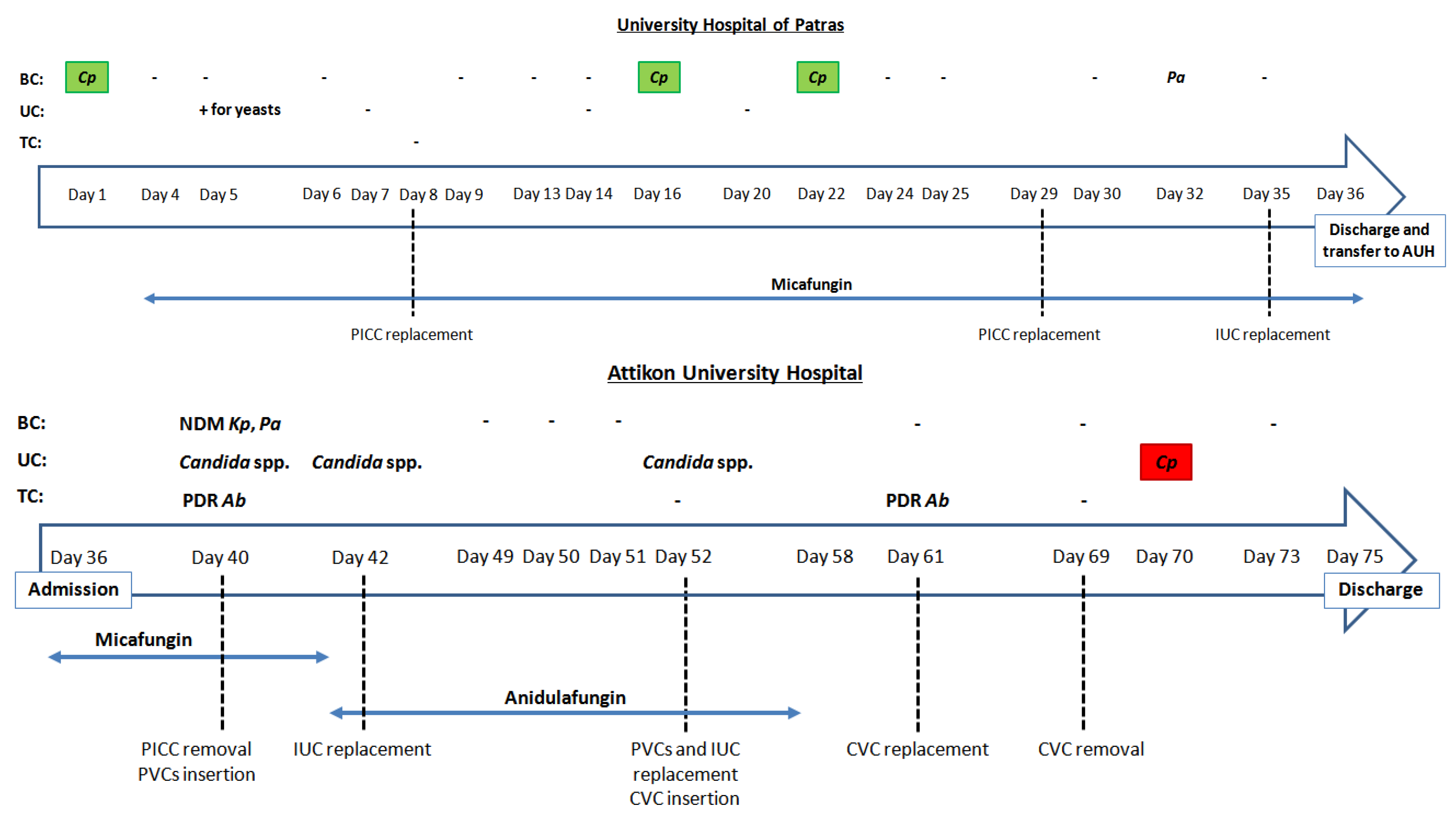
2. Case Report
2.1. Clinical Case
2.2. Mycological Workup
3. Discussions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tóth, R.; Nosek, J.; Mora-Montes, H.M.; Gabaldon, T.; Bliss, J.M.; Nosanchuk, J.D.; Turner, S.A.; Butler, G.; Vágvölgyi, C.; Gácser, A. Candida parapsilosis: From Genes to the Bedside. Clin. Microbiol. Rev. 2019, 32, e00111-18. [Google Scholar] [CrossRef] [PubMed]
- Mamali, V.; Siopi, M.; Charpantidis, S.; Samonis, G.; Tsakris, A.; Vrioni, G. Increasing Incidence and Shifting Epidemiology of Candidemia in Greece: Results from the First Nationwide 10-Year Survey. J. Fungi 2022, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Govender, N.P.; Patel, J.; Magobo, R.E.; Naicker, S.; Wadula, J.; Whitelaw, A.; Coovadia, Y.; Kularatne, R.; Govind, C.; Lockhart, S.R.; et al. Emergence of azole-resistant Candida parapsilosis causing bloodstream infection: Results from laboratory-based sentinel surveillance in South Africa. J. Antimicrob. Chemother. 2016, 71, 1994–2004. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.K.B.; Bonfietti, L.X.; Garcia, R.A.; Araujo, M.R.; Rodrigues, J.S.; Gimenes, V.M.F.; Melhem, M.S.C. Antifungal susceptibility profile of Candida clinical isolates from 22 hospitals of São Paulo State, Brazil. Braz. J. Med. Biol. Res. 2021, 54, e10928. [Google Scholar] [CrossRef]
- Bustamante, B.; Martins, M.A.; Bonfietti, L.X.; Szeszs, M.W.; Jacobs, J.; Garcia, C.; Melhem, M.S.C. Species distribution and antifungal susceptibility profile of Candida isolates from bloodstream infections in Lima, Peru. J. Med. Microbiol. 2014, 63, 855–860. [Google Scholar] [CrossRef]
- Nucci, M.; Queiroz-Telles, F.; Alvarado-Matute, T.; Tiraboschi, I.N.; Cortes, J.; Zurita, J.; Guzman-Blanco, M.; Santolaya, M.E.; Thompson, L.; Sifuentes-Osornio, J.; et al. Epidemiology of candidemia in Latin America: A laboratory-based survey. PLoS ONE 2013, 8, e59373. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef]
- Cornely, O.A.; Bassetti, M.; Calandra, T.; Garbino, J.; Kullberg, B.J.; Lortholary, O.; Meersseman, W.; Akova, M.; Arendrup, M.C.; Arikan-Akdagli, S.; et al. ESCMID* guideline for the diagnosis and management of Candida diseases 2012: Non-neutropenic adult patients. Clin. Microbiol. Infect. 2012, 18 (Suppl. S7), 19–37. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Katiyar, S.K.; Park, S.; Edlind, T.D.; Perlin, D.S. A Naturally Occurring Proline-to-Alanine Amino Acid Change in Fks1p in Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis Accounts for Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2008, 52, 2305. [Google Scholar] [CrossRef]
- Pfeiffer, C.D.; Garcia-Effron, G.; Zaas, A.K.; Perfect, J.R.; Perlin, D.S.; Alexander, B.D. Breakthrough Invasive Candidiasis in Patients on Micafungin. J. Clin. Microbiol. 2010, 48, 2373. [Google Scholar] [CrossRef] [Green Version]
- Kabbara, N.; Lacroix, C.; De Latour, R.P.; Socié, G.; Ghannoum, M.; Ribaud, P. Breakthrough, C. parapsilosis and C. guilliermondii blood stream infections in allogeneic hematopoietic stem cell transplant recipients receiving long-term caspofungin therapy. Haematologica 2008, 93, 639–640. [Google Scholar] [CrossRef] [PubMed]
- Chiotos, K.; Vendetti, N.; Zaoutis, T.E.; Baddley, J.; Ostrosky-Zeichner, L.; Pappas, P.; Fisher, B.T. Comparative effectiveness of echinocandins versus fluconazole therapy for the treatment of adult candidaemia due to Candida parapsilosis: A retrospective observational cohort study of the Mycoses Study Group (MSG-12). J. Antimicrob. Chemother. 2016, 71, 3536–3539. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.; Horn, T.; Frimodt-Møller, N. In Vivo Pathogenicity of Eight Medically Relevant Candida Species in an Animal Model. Infection 2002, 30, 286–291. [Google Scholar] [CrossRef]
- Papp, C.; Kocsis, K.; Tóth, R.; Bodai, L.; Willis, J.R.; Ksiezopolska, E.; Lozoya-Pérez, N.E.; Vágvölgyi, C.; Montes, H.M.; Gabaldón, T.; et al. Echinocandin-Induced Microevolution of Candida parapsilosis Influences Virulence and Abiotic Stress Tolerance. mSphere 2018, 3, e00547-18. [Google Scholar] [CrossRef]
- Chassot, F.; Venturini, T.P.; Piasentin, F.B.; Rossato, L.; Fiorini, A.; Svidzinski, T.I.E.; Alves, S.H. Exploring the In Vitro Resistance of Candida parapsilosis to Echinocandins. Mycopathologia 2016, 181, 663–670. [Google Scholar] [CrossRef]
- Moudgal, V.; Little, T.; Boikov, D.; Vazquez, J.A. Multiechinocandin- and Multiazole-Resistant Candida parapsilosis Isolates Serially Obtained during Therapy for Prosthetic Valve Endocarditis. Antimicrob. Agents Chemother. 2005, 49, 767. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Daneshnia, F.; Hilmioglu-Polat, S.; Ilkit, M.; Yasar, M.; Polat, F.; Metin, D.Y.; Dokumcu, Ü.Z.; Pan, W.; Hagen, F.; et al. Genetically related micafungin-resistant Candida parapsilosis blood isolates harbouring novel mutation R658G in hotspot 1 of Fks1p: A new challenge? J. Antimicrob. Chemother. 2021, 76, 418–422. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Hamal, P.; Guinea, J. EUCAST DEFINITIVE DOCUMENT E.DEF 7.3.2. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts. Available online: http://www.eucast.org (accessed on 25 September 2021).
- Siopi, M.; Tarpatzi, A.; Kalogeropoulou, E.; Damianidou, S.; Vasilakopoulou, A.; Vourli, S.; Pournaras, S.; Meletiadis, J. Epidemiological trends of fungemia in Greece with a focus on candidemia during the recent financial crisis: A 10-year survey in a tertiary care academic hospital and review of literature. Antimicrob. Agents Chemother. 2020, 64, e01516-19. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Najafzadeh, M.J.; Hagen, F.; Mahmoudi, S.; Salehi, M.; Zarrinfar, H.; Namvar, Z.; Zareshahrabadi, Z.; Khodavaisy, S.; et al. Evaluation of Molecular Epidemiology, Clinical Characteristics, Antifungal Susceptibility Profiles, and Molecular Mechanisms of Antifungal Resistance of Iranian Candida parapsilosis Species Complex Blood Isolates. Front. Cell. Infect. Microbiol. 2020, 10, 206. [Google Scholar] [CrossRef]
- Hare, R.K.; Arastehfar, A.; Rosendahl, S.; Charsizadeh, A.; Daneshnia, F.; Eshaghi, H.; Mirhendi, H.; Boekhout, T.; Hagen, F.; Arendrup, M.C. Candidemia among Hospitalized Pediatric Patients Caused by Several Clonal Lineages of Candida parapsilosis. J. Fungi 2022, 8, 183. [Google Scholar] [CrossRef]
- McCarty, T.P.; White, C.M.; Pappas, P.G. Candidemia and Invasive Candidiasis. Infect. Dis. Clin. 2021, 35, 389–413. [Google Scholar] [CrossRef] [PubMed]
- Odabasi, Z.; Mert, A. Candida urinary tract infections in adults. World J. Urol. 2019, 38, 2699–2707. [Google Scholar] [CrossRef]
- Shields, R.K.; Nguyen, M.H.; Clancy, C.J. Clinical perspectives on echinocandin resistance among Candida species. Curr. Opin. Infect. Dis. 2015, 28, 514–522. [Google Scholar] [CrossRef]
- Healey, K.R.; Nagasaki, Y.; Zimmerman, M.; Kordalewska, M.; Park, S.; Zhao, Y.; Perlin, D.S. The Gastrointestinal Tract Is a Major Source of Echinocandin Drug Resistance in a Murine Model of Candida glabrata Colonization and Systemic Dissemination. Antimicrob. Agents Chemother. 2017, 61, e01412-17. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.H.; Johansen, H.K.; Søes, L.M.; Lemming, L.E.; Rosenvinge, F.S.; Nielsen, L.; Olesen, B.; Kristensen, L.; Dzajic, E.; Astvad, K.M.T.; et al. Posttreatment Antifungal Resistance among Colonizing Candida Isolates in Candidemia Patients: Results from a Systematic Multicenter Study. Antimicrob. Agents Chemother. 2016, 60, 1500. [Google Scholar] [CrossRef]
- Perlin, D.S. Echinocandin Resistance in Candida. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S612–S617. [Google Scholar] [CrossRef] [PubMed]
- Naicker, S.D.; Magobo, R.E.; Zulu, T.G.; Maphanga, T.G.; Luthuli, N.; Lowman, W.; Govender, N.P. Two echinocandin-resistant Candida glabrata FKS mutants from South Africa. Med. Mycol. Case Rep. 2016, 11, 24–26. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Perlin, D.S. Echinocandin resistance: An emerging clinical problem? Curr. Opin. Infect. Dis. 2014, 27, 484–492. [Google Scholar] [CrossRef]
- Jamiu, A.T.; Albertyn, J.; Sebolai, O.M.; Pohl, C.H. Update on Candida krusei, a potential multidrug-resistant pathogen. Med. Mycol. 2021, 59, 14–30. [Google Scholar] [CrossRef]
- Arastehfar, A.; Lass-Flörl, C.; Garcia-Rubio, R.; Daneshnia, F.; Ilkit, M.; Boekhout, T.; Gabaldon, T.; Perlin, D.S. The Quiet and Underappreciated Rise of Drug-Resistant Invasive Fungal Pathogens. J. Fungi 2020, 6, 138. [Google Scholar] [CrossRef]
- Mesquida, A.; Díaz-García, J.; Sánchez-Carrillo, C.; Martín-Rabadán, P.; Alcalá, L.; Muñoz, P.; Escribano, P.; Guinea, J. ΔF659 and F659S substitutions at the HS1 of FKS2 gene, along with E655A and W715L upstream and downstream substitutions, correlate with high ibrexafungerp MICs against Candida glabrata. Clin. Microbiol. Infect. 2022, 28, 1154.e5–1154.e8. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Macesic, N.; Blakeway, L. Comment on: Genetically related micafungin-resistant Candida parapsilosis blood isolates harbouring novel mutation R658G in hotspot 1 of Fks1p: A new challenge? J. Antimicrob. Chemother. 2022, 77, 1790. [Google Scholar] [CrossRef]
- Davari, A.; Haghani, I.; Hassanmoghadam, F.; Nabili, M.; Shokohi, T.; Hedayati, M.T.; Shabanzadeh, S.; Moazeni, M. Echinocandin resistance in Candida parapsilosis sensu stricto: Role of alterations in CHS3, FKS1 and Rho gene expression. J. Glob. Antimicrob. Resist. 2020, 22, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Martí-Carrizosa, M.; Sánchez-Reus, F.; March, F.; Cantón, E.; Coll, P. Implication of Candida parapsilosis FKS1 and FKS2 Mutations in Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 2015, 59, 3570. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Healey, K.R.; Shor, E.; Kordalewska, M.; Ortigosa, C.J.; Paderu, P.; Xiao, M.; Wang, H.; Zhao, Y.; Lin, L.Y.; et al. Novel FKS1 and FKS2 modifications in a high-level echinocandin resistant clinical isolate of Candida glabrata. Emerg. Microbes Infect. 2019, 8, 1619. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, M.; Qiao, D.; Shen, H.; Wang, L.; Wang, D.; Li, L.; Liu, Y.; Lu, H.; Wang, C.; et al. Prevalence and Antifungal Susceptibility of Candida parapsilosis Species Complex in Eastern China: A 15-Year Retrospective Study by ECIFIG. Front. Microbiol. 2021, 12, 644000. [Google Scholar] [CrossRef]


{kind=link}
| Isolate | Clinical Specimen | Antifungal Treatment, Duration in Days | EUCAST MIC (mg/L) | Fks1Alteration | Genotype | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMB | FLC | VRC | ITC | POS | ISA | AFG | CAS | MFG | RZF | HS1 | HS2 | ||||
| N293 | Blood | None | 0.5 | 2 | 0.03 | 0.03 | 0.016 | 0.016 | 2 | 1 | 1 | ND | None | None | Same a |
| N308 | Blood | MFG, 13 days | 0.5 | 2 | 0.03 | 0.06 | 0.03 | 0.016 | 1 | 1 | 1 | ND | None | None | Same a |
| N315 | Blood | MFG, 19 days | 0.25 | 2 | 0.03 | 0.06 | 0.016 | 0.016 | 1 | 1 | 1 | ND | None | None | Same a |
| AUH1957 | Urine | MFG, 38 days + AFG, 16 days | 0.25 | 2 | 0.03 | 0.06 | 0.03 | 0.016 | >8 | >8 | >8 | >8 | F652S | None | Same a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siopi, M.; Papadopoulos, A.; Spiliopoulou, A.; Paliogianni, F.; Abou-Chakra, N.; Arendrup, M.C.; Damoulari, C.; Tsioulos, G.; Giannitsioti, E.; Frantzeskaki, F.; et al. Pan-Echinocandin Resistant C. parapsilosis Harboring an F652S Fks1 Alteration in a Patient with Prolonged Echinocandin Therapy. J. Fungi 2022, 8, 931. https://doi.org/10.3390/jof8090931
Siopi M, Papadopoulos A, Spiliopoulou A, Paliogianni F, Abou-Chakra N, Arendrup MC, Damoulari C, Tsioulos G, Giannitsioti E, Frantzeskaki F, et al. Pan-Echinocandin Resistant C. parapsilosis Harboring an F652S Fks1 Alteration in a Patient with Prolonged Echinocandin Therapy. Journal of Fungi. 2022; 8(9):931. https://doi.org/10.3390/jof8090931
Chicago/Turabian StyleSiopi, Maria, Antonios Papadopoulos, Anastasia Spiliopoulou, Fotini Paliogianni, Nissrine Abou-Chakra, Maiken Cavling Arendrup, Christina Damoulari, Georgios Tsioulos, Efthymia Giannitsioti, Frantzeska Frantzeskaki, and et al. 2022. "Pan-Echinocandin Resistant C. parapsilosis Harboring an F652S Fks1 Alteration in a Patient with Prolonged Echinocandin Therapy" Journal of Fungi 8, no. 9: 931. https://doi.org/10.3390/jof8090931