
Overcome Double Trouble: Baloxavir Marboxil Suppresses Influenza Thereby Mitigating Secondary Invasive Pulmonary Aspergillosis

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Influenza Virus and Aspergillus Strain
2.2. Mouse Model
2.3. Micro-Computed Tomography (µCT)
2.4. Bioluminescence Imaging (BLI)
2.5. Quantification of Virus and Fungal Titers
2.6. Histopathology
2.7. Statistical Analysis
3. Results
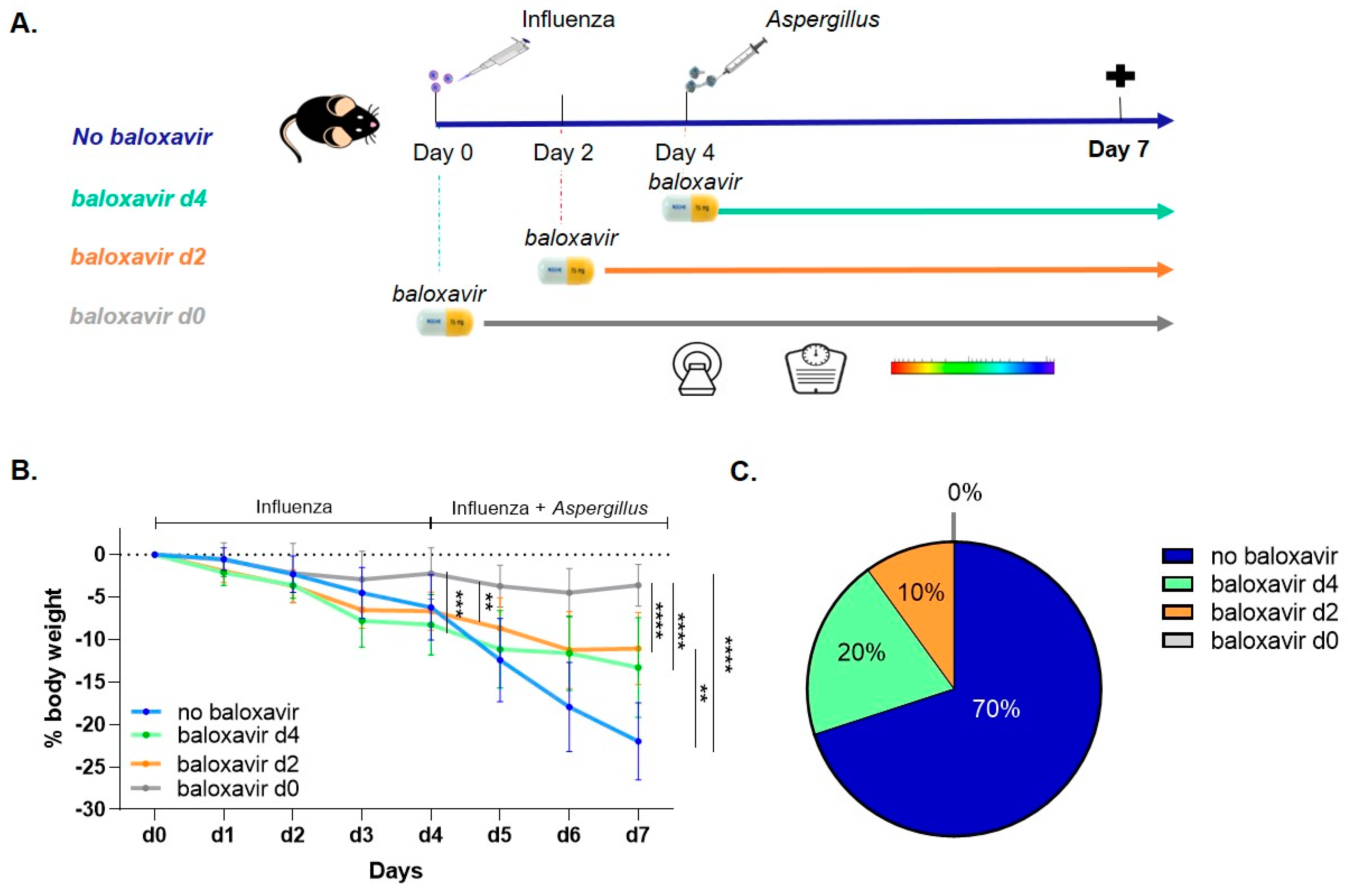
3.1. Baloxavir Marboxil Treatment Improves Survival and Limits Body Weight Loss
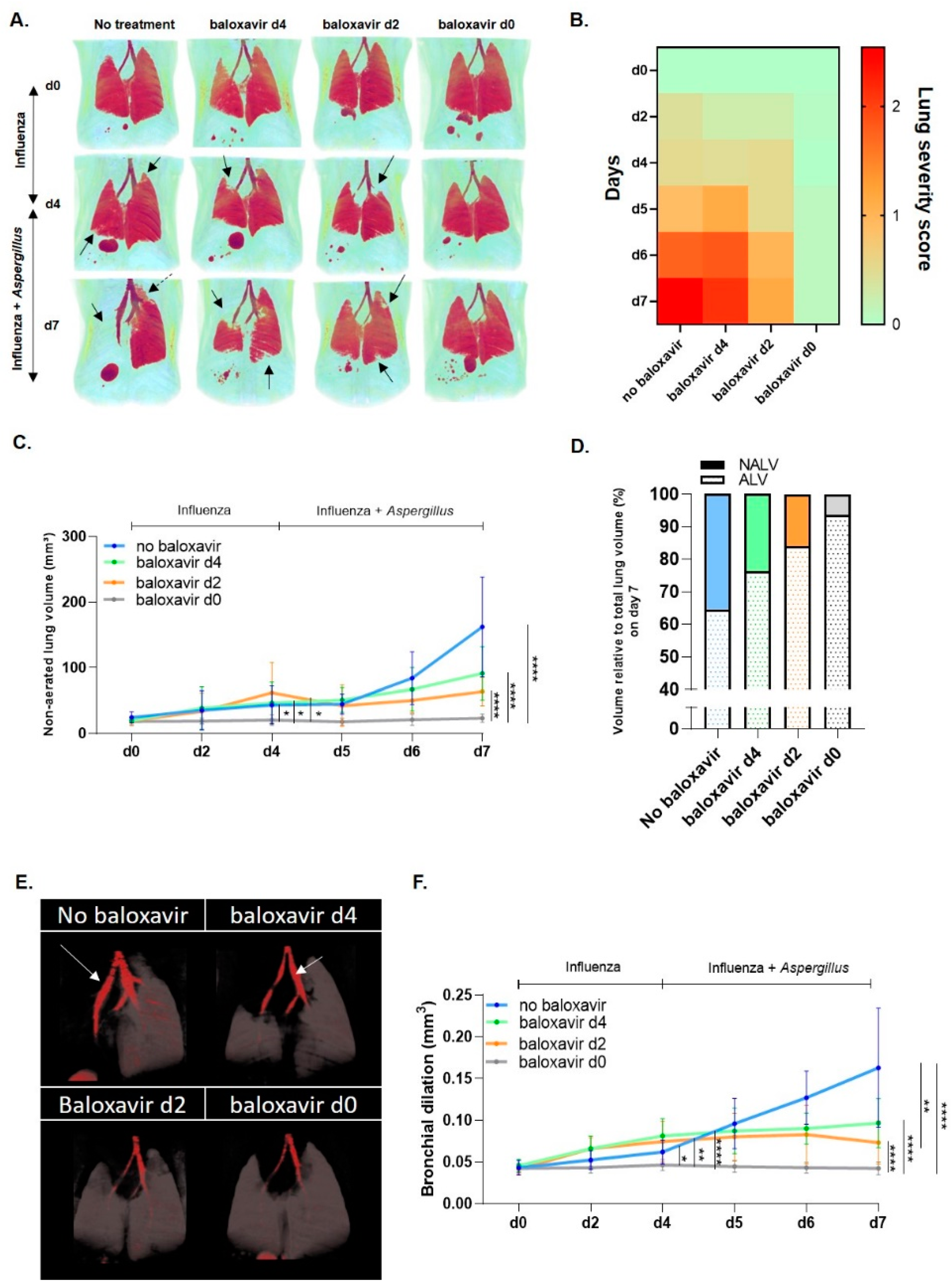
3.2. Baloxavir Marboxil Reduces Infiltrates and Bronchial Dilation on Lung µCT after Influenza and Aspergillus Co-Infection
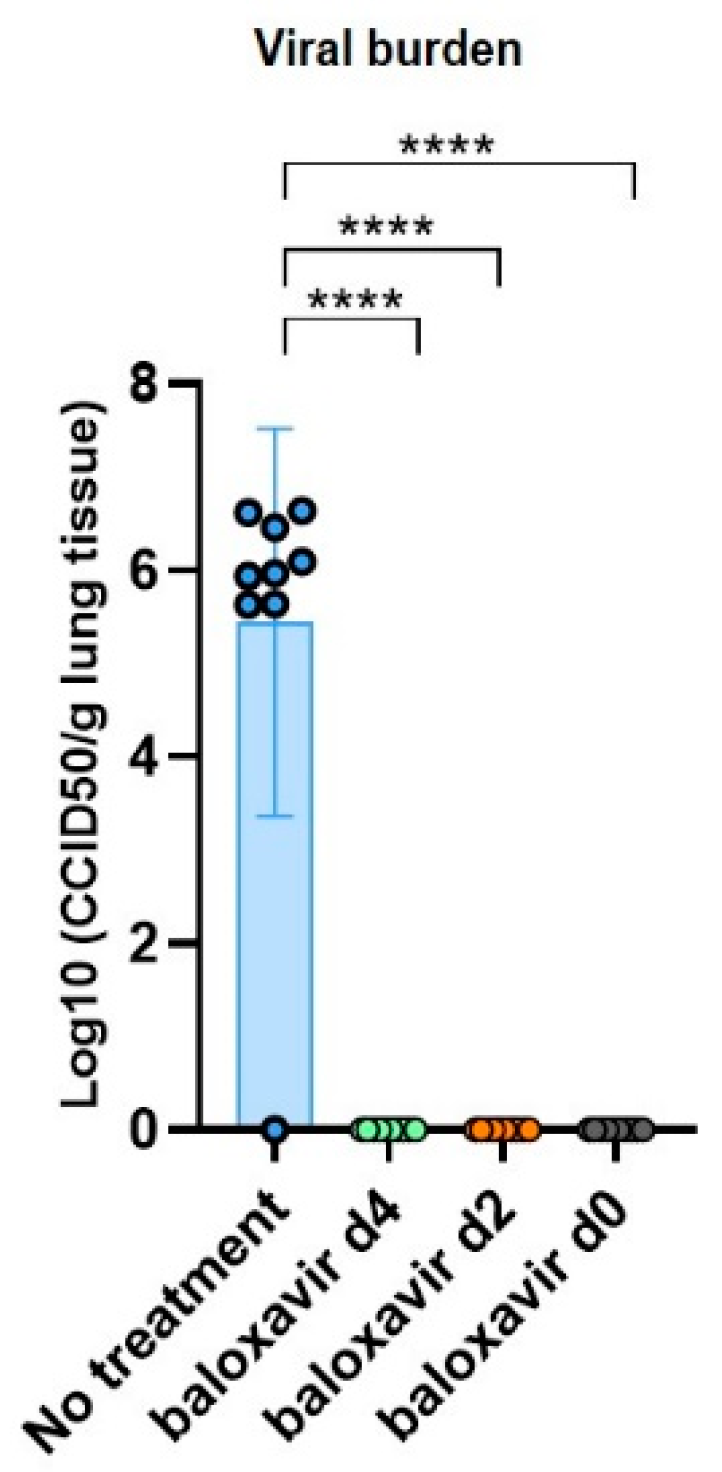
3.3. Baloxavir Marboxil Effectively Clears Influenza
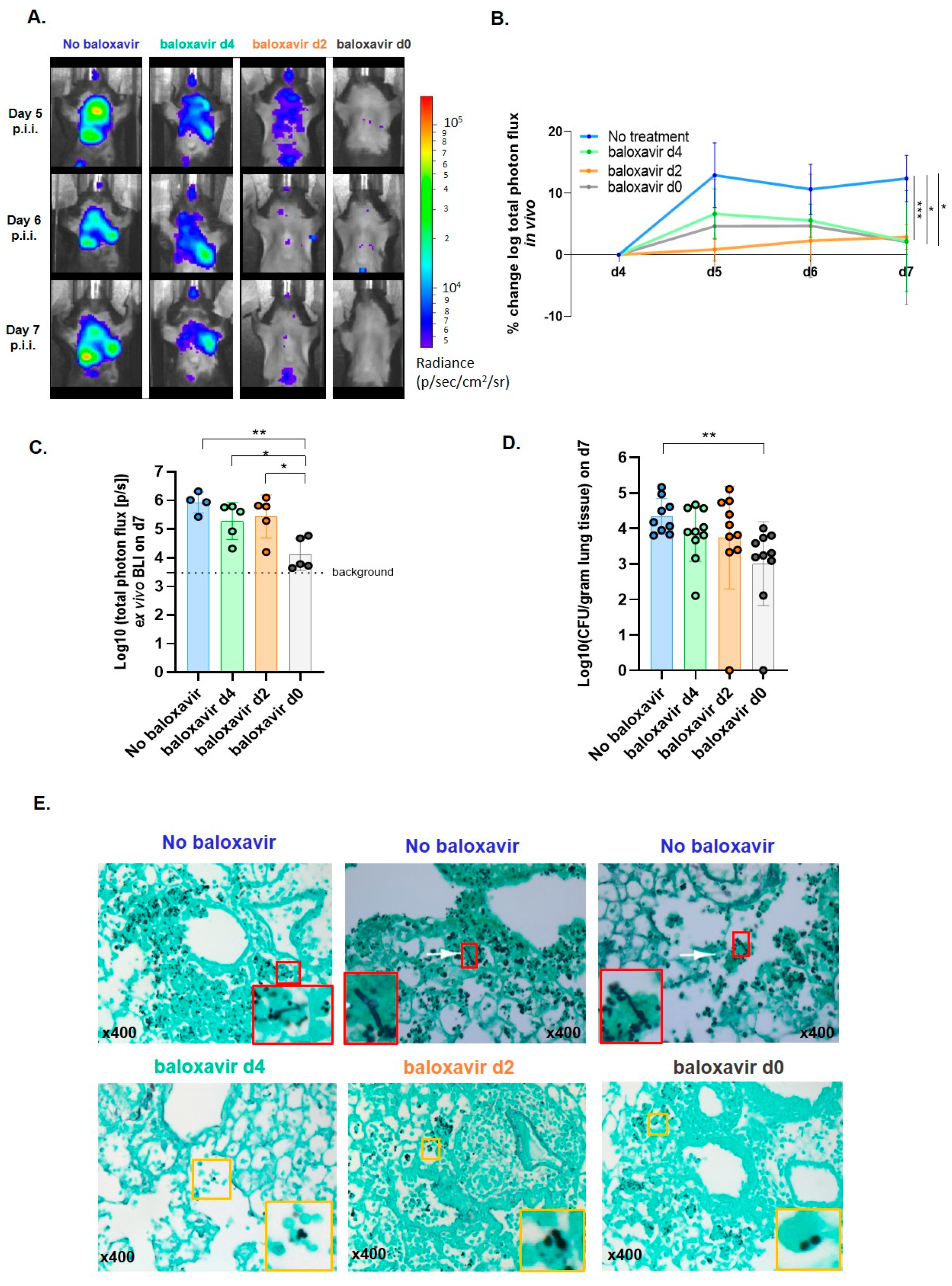
3.4. Baloxavir Marboxil Reduces the Risk for a Secondary Invasive Pulmonary Aspergillosis
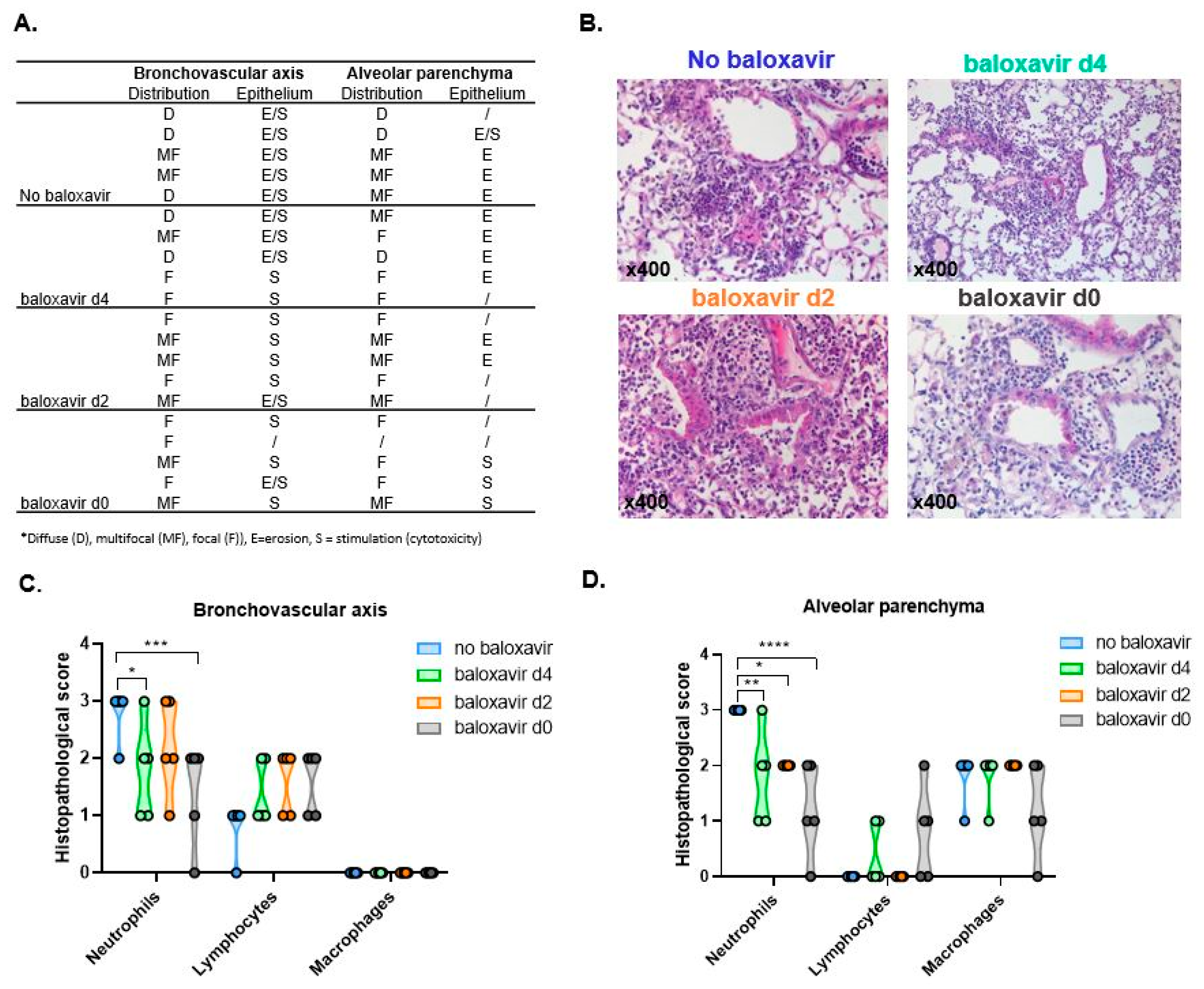
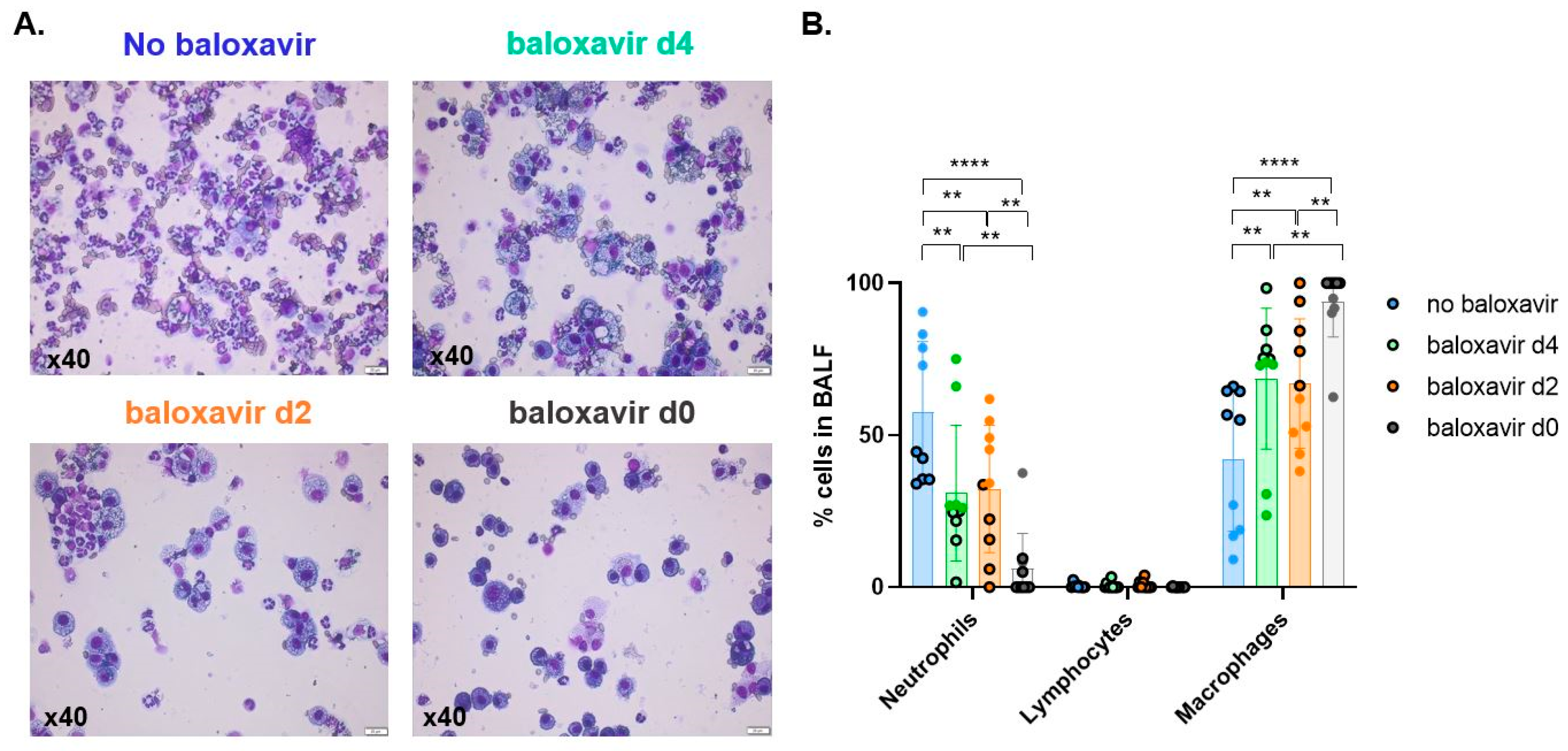
3.5. Baloxavir Marboxil Treatment Onset-Dependent Reduction in Lung Pathology and Local Immune Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fischer, J.J.; Walker, D.H. Invasive Pulmonary Aspergillosis Associated with Influenza. JAMA J. Am. Med. Assoc. 1979, 241, 1493–1494. [Google Scholar] [CrossRef]
- Vanderbeke, L.; Spriet, I.; Breynaert, C.; Rijnders, B.J.A.; Verweij, P.E.; Wauters, J. Invasive pulmonary aspergillosis complicating severe influenza: Epidemiology, diagnosis and treatment. Curr. Opin. Infect. Dis. 2018, 31, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Rijnders, B.J.A.; Brüggemann, R.J.M.; Azoulay, E.; Bassetti, M.; Blot, S.; Calandra, T.; Clancy, C.J.; Cornely, O.A.; Chiller, T.; et al. Review of influenza-associated pulmonary aspergillosis in ICU patients and proposal for a case definition: An expert opinion. Intensive Care Med. 2020, 46, 1524–1535. [Google Scholar] [CrossRef]
- Schauwvlieghe, A.F.A.D.; Rijnders, B.J.A.; Philips, N.; Verwijs, R.; Vanderbeke, L.; Van Tienen, C.; Lagrou, K.; Verweij, P.E.; Van De Veerdonk, F.L.; Gommers, D.; et al. Invasive aspergillosis in patients admitted to the intensive care unit with severe influenza: A retrospective cohort study. Lancet Respir. Med. 2018, 6, 782–792. [Google Scholar] [CrossRef]
- Van de Veerdonk, F.L.; Kolwijck, E.; Lestrade, P.P.A.; Hodiamont, C.J.; Rijnders, B.A.; van Paassen, J.; Haas, P.-J.; dos Santos, C.O.; Kampinga, G.A.; Bergmans, D.C.J.J.; et al. Influenza-associated aspergillosis in critically ill patients. Am. J. Respir. Crit. Care Med. 2017, 196, 524–527. [Google Scholar] [CrossRef]
- Wauters, J.; Baar, I.; Meersseman, P.; Meersseman, W.; Dams, K.; De Paep, R.; Lagrou, K.; Wilmer, A.; Jorens, P.; Hermans, G. Invasive pulmonary aspergillosis is a frequent complication of critically ill H1N1 patients: A retrospective study. Intensive Care Med. 2012, 38, 1761–1768. [Google Scholar] [CrossRef]
- Liu, W.-L.; Yu, W.-L.; Chan, K.-S.; Yang, C.-C.; Wauters, J.; Verweij, P. Aspergillosis related to severe influenza: A worldwide phenomenon? Clin. Respir. J. 2019, 13, 540–542. [Google Scholar] [CrossRef]
- Donnelly, J.P.; Chen, S.C.; Kauffman, C.A.; Steinbach, W.J.; Baddley, J.W.; Verweij, P.E.; Clancy, C.J.; Wingard, J.R.; Lockhart, S.R.; Groll, A.H.; et al. Revision and update of the consensus definitions of invasive fungal disease from the european organization for research and treatment of cancer and the mycoses study group education and research consortium. Clin. Infect. Dis. 2020, 71, 1367–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feys, S.; Almyroudi, M.P.; Braspenning, R.; Lagrou, K.; Spriet, I.; Dimopoulos, G.; Wauters, J. A Visual and Comprehensive Review on COVID-19-Associated. J. Fungi 2021, 7, 1067. [Google Scholar] [CrossRef]
- Segala, F.V.; Bavaro, D.F.; Di Gennaro, F.; Salvati, F.; Marrota, C.; Saracino, A.; Murri, R.; Fantoni, M. Impact of SARS-CoV-2 epidemic on antimicrobial resistance: A literature review. Viruses 2021, 13, 2110. [Google Scholar] [CrossRef]
- Chen, L.; Han, X.; Li, Y.; Zhang, C.; Xing, X. Invasive pulmonary aspergillosis in immunocompetent patients hospitalised with influenza A-related pneumonia: A multicenter retrospective study. BMC Pulm. Med. 2020, 20, 239. [Google Scholar] [CrossRef] [PubMed]
- Seldeslachts, L.; Vanderbeke, L.; Fremau, A.; Reséndiz-Sharpe, A.; Jacobs, C.; Laeveren, B.; Ostyn, T.; Naesens, L.; Brock, M.; Van De Veerdonk, F.L.; et al. Early oseltamivir reduces risk for influenza-associated aspergillosis in a double-hit murine model. Virulence 2021, 12, 2493–2508. [Google Scholar] [CrossRef] [PubMed]
- Tobin, J.M.; Nickolich, K.L.; Ramanan, K.; Pilewski, M.J.; Lamens, K.D.; Alcorn, J.F.; Robinson, K.M. Influenza Suppresses Neutrophil Recruitment to the Lung and Exacerbates Secondary Invasive Pulmonary Aspergillosis. J. Immunol. 2020, 205, 480–488. [Google Scholar] [CrossRef]
- Jaiswal, S.R.; Bhagwati, G.; Soni, M.; Thatai, A.; Aiyer, H.; Chakrabarti, S. Prophylactic oseltamivir during major seasonal influenza H1N1 outbreak might reduce both H1N1 and associated pulmonary aspergillosis in children undergoing haploidentical transplantation. Transpl. Infect. Dis. 2020, 22, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Koseki, K.; Yamaguchi, K.; Moriya, S.; Suzuki, Y.; Yingsakmongkon, S.; Hirai, G.; Sodeoka, M.; von Itzstein, M.; Miyagi, T. Limited inhibitory effects of oseltamivir and zanamivir on human sialidases. Antimicrob. Agents Chemother. 2008, 52, 3484–3491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewi, I.M.W.; Cunha, C.; Jaeger, M.; Gresnigt, M.S.; Gkountzinopoulou, M.E.; Garishah, F.M.; Duarte-Oliveira, C.; Campos, C.F.; Vanderbeke, L.; Sharpe, A.R.; et al. Neuraminidase and SIGLEC15 modulate the host defense against pulmonary aspergillosis. Cell Rep. Med. 2021, 2, 100289. [Google Scholar] [CrossRef] [PubMed]
- Ison, M.G.; Hayden, F.G.; Hay, A.J.; Gubareva, L.V.; Govorkova, E.A.; Takashita, E.; McKimm-Breschkin, J.L. Influenza polymerase inhibitor resistance: Assessment of the current state of the art—A report of the isirv Antiviral group. Antivir. Res. 2021, 194, 105158. [Google Scholar] [CrossRef]
- Antiviral Drug Resistance among Influenza Viruses. 2016. Available online: https://www.cdc.gov/flu/professionals/antivirals/antiviral-drug-resistance.htm (accessed on 21 August 2021).
- Behillil, S.; May, F.; Fourati, S.; Luyt, C.-E.; Chicheportiche, T.; Sonneville, R.; Tandjaoui-Lambiotte, Y.; Roux, D.; Guérin, L.; Mayaux, J.; et al. Oseltamivir resistance in severe influenza a (H1N1)pdm09 pneumonia and acute respiratory distress syndrome: A french multicenter observational cohort study. Clin. Infect. Dis. 2020, 71, 1089–1091. [Google Scholar] [CrossRef]
- Hurt, A.C.; Hardie, K.; Wilson, N.J.; Deng, Y.-M.; Osbourn, M.; Leang, S.K.; Lee, R.T.C.; Iannello, P.; Gehrig, N.; Shaw, R.; et al. Characteristics of a widespread community cluster of H275Y oseltamivir-resistant a(H1N1)pdm09 influenza in Australia. J. Infect. Dis. 2012, 206, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Macesic, N.; Laplante, J.M.; Aaron, J.G.; DiMango, E.A.; Miko, B.A.; Pereira, M.R.; Reshef, R.; George, K.S. Baloxavir treatment of oseltamivir-resistant influenza A/H1pdm09 in two immunocompromised patients. Transpl. Infect. Dis. 2021, 23, 1–5. [Google Scholar] [CrossRef]
- FDA Approves New Drug to Treat Influenza. 2018. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-new-drug-treat-influenza (accessed on 21 August 2021).
- Xofluza (Baloxavir Marboxil). 2021. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/xofluza (accessed on 11 August 2021).
- Hayden, F.G.; Sugaya, N.; Hirotsu, N.; Lee, N.; De Jong, M.D.; Hurt, A.C.; Ishida, T.; Sekino, H.; Yamada, K.; Portsmouth, S.; et al. Baloxavir marboxil for uncomplicated influenza in adults and adolescents. N. Engl. J. Med. 2018, 379, 913–923. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Lai, C.C.; Wang, Y.H.; Chen, C.H.; Wang, C.Y. Clinical efficacy and safety of baloxavir marboxil in the treatment of influenza: A systematic review and meta-analysis of randomized controlled trials. J. Microbiol. Immunol. Infect. 2021, 54, 865–875. [Google Scholar] [CrossRef]
- Fukao, K.; Ando, Y.; Noshi, T.; Kitano, M.; Noda, T.; Kawai, M.; Yoshida, R.; Sato, A.; Shishido, T.; Naito, A. Baloxavir marboxil, a novel cap-dependent endonuclease inhibitor potently suppresses influenza virus replication and represents therapeutic effects in both immunocompetent and immunocompromised mouse models. PLoS ONE 2019, 14, e0217307. [Google Scholar] [CrossRef]
- Fukao, K.; Noshi, T.; Yamamoto, A.; Kitano, M.; Ando, Y.; Noda, T.; Baba, K.; Matsumoto, K.; Higuchi, N.; Ikeda, M.; et al. Combination treatment with the cap-dependent endonuclease inhibitor baloxavir marboxil and a neuraminidase inhibitor in a mouse model of influenza A virus infection. J. Antimicrob. Chemother. 2019, 74, 654–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resendiz-Sharpe, A.; Peres Da Silva, R.; Geib, E. Longitudinal multimodal imaging-compatible mouse model of triazole sensitive and resistant invasive pulmonary aspergillosis. Under Consideration.
- Luther, K.; Rohde, M.; Sturm, K.; Kotz, A.; Heesemann, J.; Ebel, F. Characterisation of the phagocytic uptake of Aspergillus fumigatus conidia by macrophages. Microbes Infect. 2008, 10, 175–184. [Google Scholar] [CrossRef]
- Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Am. Soc. Microbiol. Clin. Microbiol. Rev. 2020, 33, 1–75. [Google Scholar]
- Nakamura, S.; Davis, K.M.; Weiser, J.N. Synergistic stimulation of type I interferons during influenza virus coinfection promotes Streptococcus pneumoniae colonization in mice. J. Clin. Investig. 2011, 121, 3657–3665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, F.; Bignell, E.; Brown, G.D.; Cook, P.C.; Warris, A. Pathogenesis of Respiratory Viral and Fungal Coinfections. Clin. Microbiol. Rev. 2021. [Google Scholar] [CrossRef]
- Sun, K.; Metzger, D.W. Inhibition of pulmonary antibacterial defense by interferon-γ during recovery from influenza infection. Nat. Med. 2008, 14, 558–564. [Google Scholar] [CrossRef]
- Bigot, P.; Auffret, M.; Gautier, S.; Weinborn, M.; Ettahar, N.K.; Coupé, P. Unexpected platelets elevation in a patient with idiopathic thrombocytopenia treated with oseltamivir for influenza infection. Fundam. Clin. Pharmacol. 2016, 30, 483–485. [Google Scholar] [CrossRef]
- Groeneveld, G.H.; Marbus, S.D.; Ismail, N.; de Vries, J.J.; Schneeberger, P.; Oosterheert, J.J.; van Dissel, J.T.; de Boer, M.G. Effectiveness of oseltamivir in reduction of complications and 30-day mortality in severe seasonal influenza infection. Int. J. Antimicrob. Agents 2020, 56, 106155. [Google Scholar] [CrossRef] [PubMed]
- Muthuri, S.G.; Venkatesan, S.; Myles, P.; Leonardi-Bee, J.; Al Khuwaitir, T.S.A.; Al Mamun, A.; Anovadiya, A.; Azziz-Baumgartner, E.; Báez, C.; Bassetti, M.; et al. Effectiveness of neuraminidase inhibitors in reducing mortality in patients admitted to hospital with influenza A H1N1pdm09 virus infection: A meta-analysis of individual participant data. Lancet Respir. Med. 2014, 2, 395–404. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seldeslachts, L.; Jacobs, C.; Tielemans, B.; Vanhoffelen, E.; Van der Sloten, L.; Humblet-Baron, S.; Naesens, L.; Lagrou, K.; Verbeken, E.; Wauters, J.; et al. Overcome Double Trouble: Baloxavir Marboxil Suppresses Influenza Thereby Mitigating Secondary Invasive Pulmonary Aspergillosis. J. Fungi 2022, 8, 1. https://doi.org/10.3390/jof8010001
Seldeslachts L, Jacobs C, Tielemans B, Vanhoffelen E, Van der Sloten L, Humblet-Baron S, Naesens L, Lagrou K, Verbeken E, Wauters J, et al. Overcome Double Trouble: Baloxavir Marboxil Suppresses Influenza Thereby Mitigating Secondary Invasive Pulmonary Aspergillosis. Journal of Fungi. 2022; 8(1):1. https://doi.org/10.3390/jof8010001
Chicago/Turabian StyleSeldeslachts, Laura, Cato Jacobs, Birger Tielemans, Eliane Vanhoffelen, Lauren Van der Sloten, Stephanie Humblet-Baron, Lieve Naesens, Katrien Lagrou, Erik Verbeken, Joost Wauters, and et al. 2022. "Overcome Double Trouble: Baloxavir Marboxil Suppresses Influenza Thereby Mitigating Secondary Invasive Pulmonary Aspergillosis" Journal of Fungi 8, no. 1: 1. https://doi.org/10.3390/jof8010001