
The Genus Leccinum (Boletaceae, Boletales) from China Based on Morphological and Molecular Data
by
Xin Meng
1,2,3, 
Geng-Shen Wang
1,2,3,
Gang Wu
1,2,
Pan-Meng Wang
1,2,3,
Zhu L. Yang
1,2,* and
Yan-Chun Li
1,2,* 1
Key Laboratory for Plant Diversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
2
Yunnan Key Laboratory for Fungal Diversity and Green Development, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
3
College of Life Sciences, University of Chinese Academy of Sciences, Beijing 100049, China
*
Authors to whom correspondence should be addressed.
J. Fungi 2021, 7(9), 732; https://doi.org/10.3390/jof7090732
Submission received: 28 May 2021
/
Revised: 20 July 2021
/
Accepted: 31 August 2021
/
Published: 6 September 2021
(This article belongs to the Special Issue Biodiversity, Distribution and Conservation of Plants and Fungi; Effects of Global Warming and Environmental Stress)
Abstract
:Leccinum is one of the most important groups of boletes. Most species in this genus are ectomycorrhizal symbionts of various plants, and some of them are well-known edible mushrooms, making it an exceptionally important group ecologically and economically. The scientific problems related to this genus include that the identification of species in this genus from China need to be verified, especially those referring to European or North American species, and knowledge of the phylogeny and diversity of the species from China is limited. In this study, we conducted multi-locus (nrLSU, tef1-α, rpb2) and single-locus (ITS) phylogenetic investigations and morphological observisions of Leccinum from China, Europe and North America. Nine Leccinum species from China, including three new species, namely L. album, L. parascabrum and L. pseudoborneense, were revealed and described. Leccinum album is morphologically characterized by the white basidioma, the white hymenophore staining indistinct greenish blue when injured, and the white context not changing color in pileus but staining distinct greenish blue in the base of the stipe when injured. Leccinum parascabrum is characterized by the initially reddish brown to chestnut-brown and then pale brownish to brown pileus, the white to pallid and then light brown hymenophore lacking color change when injured, and the white context lacking color change in pileus but staining greenish blue in the base of the stipe when injured. Leccinum pseudoborneense is characterized by the pale brown to dark brown pileus, the initially white and then brown hymenophore lacking color change when injured, and the white context in pileus and stipe lacking color change in pileus but staining blue in stipe when bruised. Color photos of fresh basidiomata, line drawings of microscopic features and detailed descriptions of the new species are presented.
1. Introduction
The genus Leccinum Gray is a species-rich genus of Boletaceae and is characterized by a whitish or yellow hymenophore, a white to cream context unchanging or staining blue or red when injured, a brown to blackish scabrous to dotted squamules on the surface of the stipe, and comparatively long and smooth basidiospores. Generally, most species of the genus are widely spread in the subarctic, boreal, temperate and Mediterranean regions, with a few secondary expansions to the neotropics [1,2,3,4,5,6,7,8,9,10,11,12]. Species in Leccinum are both ecologically and economically important. Most species of this genus exhibit mycorrhizal host specificity. Species of Leccinum sect. Scabra Smith & Thiers are associated with plants of Betula, while species of L. sect. Fumosa (A.H. Smith, Thiers & Watling) Gelardi are associated with plants of Populus. In L. sect. Leccinum, species are found exclusively associated with plants of Populus (e.g., L. albostipitatum den Bakker & Noordel. and L. insigne A.H. Sm., Thiers & Watling), Betula (e.g., L. atrostipitatum A.H. Sm., Thiers & Watling), Pinaceae (e.g., L. vulpinum Watling and L. piceinum Pilát & Dermek) and Ericaceae that form arbutoidmycorrhizas (e.g., L. manzanitae Thiers and L. monticola Halling & G.M. Muell.). However, there are species in section Leccinum that are not host specific, i.e., L. aurantiacum (Bull.) Gray. This species is associated with plants of Betula, Populus, Quercus, Salix and sometimes with Tilia [13,14]. Some species of this genus are well-known edible mushrooms, such as L. quercinum (Pilát) E.E. Green & Watling, L. scabrum (Bull.) Gray and L. versipelle (Fr. & Hök) Snell, which are collected in China during the mushroom season.
The genus Leccinum was established by Gray in 1821 [13], based on the type species L. aurantiacum. Subsequently, more and more mycologists noticed the morphological distinctness and described many new species of this genus. As currently circumscribed, the genus comprises roughly 150 species [1,2,3,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56]. North America is the species diversity center of this genus, and in total 118 species have been recorded from this area [19]. Some of the most important works are the serial works of Smith and Thiers [1,15,16,17], in which three sections of this genus were proposed (L. sect. Leccinum Smith & Thiers, L. sect. Luteoscabra Smith & Thiers and L. sect. Scabra), with 68 species described from Michigan. Twelve species from Central America were described: one species from Belize, eight species from Costa Rica and three species from Colombia [20,21,22,23,24]. In Europe, Singer divided species of this genus into four sections, including two known sections, L. sect. Luteoscabra and L. sect. Leccinum, and two newly proposed sections, L. sect. Roseoscabra and L. sect. Eximia [3]. In Singer’s infrageneric classification, L. sect. Scabra, established by Smith and Thiers, was merged to L. sect. Leccinum. Recent molecular phylogenetic evidence has revealed that species of L. sect. Luteoscabra, L. sect. Roseoscabra and L. sect. Eximia belong to divergent clades of Boletaceae and represent many new genera (32,52–54). Thus, the genus Leccinum is restricted to the section Leccinum (Singer’s infrageneric classification) [3]. den Bakker and Noordelos revised the European Leccinum species based on morphology and nrLSU sequences and documented sixteen species [14]. In their subsequent study, they treated the three subclades revealed by den Bakker et al. in L. section Leccinum [33,57] as three subsections (viz. L. subsect. Leccinum, L. subsect. Fumosa A.H. Sm., Thiers & Watling and L. subsect. Scabra Pilat & Dermek) [14]. This infrageneric subdivision was followed in the treatment of the genus in this study. In the Southern Hemisphere, four species have been reported, including one from New Zealand and three from Australia [27,28,29].
In Asia, six species of Leccinum have been reported from Malaysia [6]; ten species from Japan [7,8,9,10]; and a total of 31 species have been reported from China based on an extensive literature review [34,35,36,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,56]. Among these Chinese species, twelve species, viz. L. albellum (Peck) Singer, L. chromapes (Frost) Singer, L. crocipodium (Letell.) Watling, L. eximium (Peck) Singer, L. extremiorientale (Lar. N. Vassiljeva) Singer, L. griseum (Quél.) Singer, L. hortonii (A.H. Sm. & Thiers) Hongo & Nagas., L. nigrescens (Richon & Roze) Singer, L. rubropunctum (Peck) Singer, L. rubrum M. Zang, L. rugosiceps (Peck) Singer and L. subglabripes (Peck) Singer have been transferred to other genera [5,11,35,52,53,54,55]; eight species, viz. L. duriusculum (Schulzer ex Fr.) Singer, L. intusrubens (Corner) Høil., L. oxydabile (Singer) Singer, L. quercinum, L. rufum (Schaeff.) Kreisel, L. subleucophaeum E.A. Dick & Snell, L. subradicatum Hongo and L. variicolor Watling were reported without specimen support [39,40,41,42,43,49,51]; and eleven species, viz. L. ambiguum A.H. Sm. & Thiers, L. atrostipitatum A.H. Sm., Thiers & Watling, L. aurantiacum, L. holopus (Rostk.) Watling, L. olivaceopallidum A.H. Sm., Thiers & Watling, L. potteri A.H. Sm., Thiers & Watling, L. roseofractum Watling, L. scabrum, L. subgranulosum A.H. Sm. & Thiers, L. subleucophaeum var. minimum C.S. Bi and L. versipelle were reported with specimen citations [34,38,44,45,46,47,48]. Among these eleven species reported with specimen citations, only L. subleucophaeum var. minimum was originally described from China, and the remaining species were identified as species originally described from Europe and North America based on general morphological similarities. Indeed, a few species described from Europe and North America do occur in China, especially in northeastern and northwestern China. However, most species found in China have evolved independently in the southern part of China. Thus, identification of the Chinese Leccinum species needs to be reconfirmed.
In this study, we used both morphological data and molecular sequences from the nuclear ribosomal internal transcribed spacer (ITS), the large subunit of the nuclear ribosomal RNA (nrLSU), the translation elongation factor 1-alpha (tef1-α) and the RNA polymerase II second largest subunit (rpb2), together with ecological data to (1) elucidate species diversity of Leccinum in China; (2) evaluate the phylogenetic relationships of species within Leccinum; (3) make morphological and ecological comparisons between closely related species.
2. Materials and Methods
2.1. Taxon Sampling
Nineteen specimens of the genus Leccinum from China were examined. For each collection, a part of the basidioma was dried with silica gel for DNA extraction. The remaining materials were then air-dried at 45–50 °C using an electric food dehydrator. Specimens studied in this work were deposited in the Herbarium of the Kunming Institute of Botany, Chinese Academy of Sciences (KUN). Genera are abbreviated as follows: L. for Leccinum, Le. for Leccinellum, O. for Octaviania, R. for Rossbeevera, Ru. for Rugiboletus, T. for Turmalinea, Ca. for Castanopsis, Li. for Lithocarpus, P. for Pinus and Q. for Quercus.
2.2. Morphological Observation
The macroscopic descriptions are based on the detailed field notes and photographs of fresh basidiomata. Color codes of the form “4B2” indicate the plate, row, and color block from Kornerup and Wanscher [58]. For microscopic studies, the microscopic features of each part of the basidioma were observed under microscope (Leica DM2000, Leica Microsystems, Wetzlar, Germany), including basidiospores, basidia, cheilocystidia, pleurocystidia and pileipellis, using 5% KOH as a mounting medium to revive the dried materials. Microscopic studies follow Zhou et al. [59]. In the description of Basidiospores, the abbreviation n/m/p means n basidiospores measured from m basidomata of p collections in 5% KOH solution. The notation of the form (a) b–c (d) stands for the dimensions of the basidiospores; the range b–c contains a minimum of 90% of the measured values, a or d given in parentheses stands for extreme values. Q is used to mean “length/width ratio” of a basidiospore in a side view; Qm means average Q of all basidiospores ± sample standard deviation. Measurements of basidospores, cystidia, basidia and terminal cells in pileipellis are presented as length × width. All microscopic structures were drawn freehand from rehydrated material under the microscope with 10× eyepiece and 100× objective (the total magnification is 1000×).
2.3. Molecular Procedures
Genomic DNA was extracted from silica gel dried materials or herbarium specimens using the CTAB (Cetyltrimethyl ammonium bromide) method [60]. Polymerase chain reactions (PCRs) were performed to amplify partial sequences of nrLSU, tef1-α, rpb2 and ITS using the extracted DNA. The nrLSU region was amplified with primers LROR/LR5 and LROR/LR3 [61]; tef1-α was amplified with primer pair EF1-983F and EF1-1567R [62]; rpb2 was amplified with primers bRPB2-6F and bRPB2-7.1R [63] and ITS was amplified with primer pair ITS1 and ITS4 [64]. Protocols for the polymerase chain reactions (PCRs) and sequencing followed those in Wu et al. [65] and the references therein.
2.4. Sequence Alignments and Phylogenetic Analyses
The newly generated sequences of each locus were blasted in GenBank, and the most closely related sequences (nucleotide identities >95%) were downloaded for further alignment. Sequences were aligned separately for each of the loci using MAFFT v7.130b with the E-INS-I strategy and manually optimized on BioEdit v7.0.9 [66,67]. Two datasets, the ITS dataset and the multi-locus (nrLSU + tef1-α + rpb2) dataset, were analyzed using RAxML and Bayesian methods, respectively. For the multi-locus dataset, single-gene analyses were conducted to assess incongruence among individual genes using the ML method (results not shown). Because no well-supported bootstrap value (BS > 70%) [55] conflict was detected among the topologies of the three genes, their sequences were then concatenated together for further multi-locus analyses.
For ML analyses, the multi-locus and ITS datasets were analyzed using RAxML (https://www.phylo.org/, accessed on 26 August 2021) under the model GTRGAMMA [68]. Statistical supports for the phylogenetic analyses were determined using nonparametric bootstrapping with 1000 replicates. For BI analyses, the parameter model was selected by the Akaike information criterion (AIC) as the best-fit likelihood model with Modeltest 3.7 (Free Software Foundation, Boston, MA, USA) [69]. The models employed for each of the four loci were GTR + I + G for ITS, nrLSU and tef1-α, and SYM + I + G for rpb2. Posterior probabilities (PP) were determined twice by running one cold and three heated chains in parallel mode, saving trees every 1000th generation. Other parameters were kept at their default settings. Runs were terminated once the average standard deviation of split frequencies went below 0.01 [70]. Chain convergence was determined using Tracer v1.5 (http://tree.bio.ed.ac.uk/software/tracer/, accessed on 26 August 2021) to confirm sufficiently large ESS values (>200). Subsequently, the sampled trees were summarized after omitting the first 25% of trees as burn-in using the ‘sump’ and ‘sumt’ commands implemented in MrBayes.
3. Results
3.1. Molecular Phylogenetic Analysis
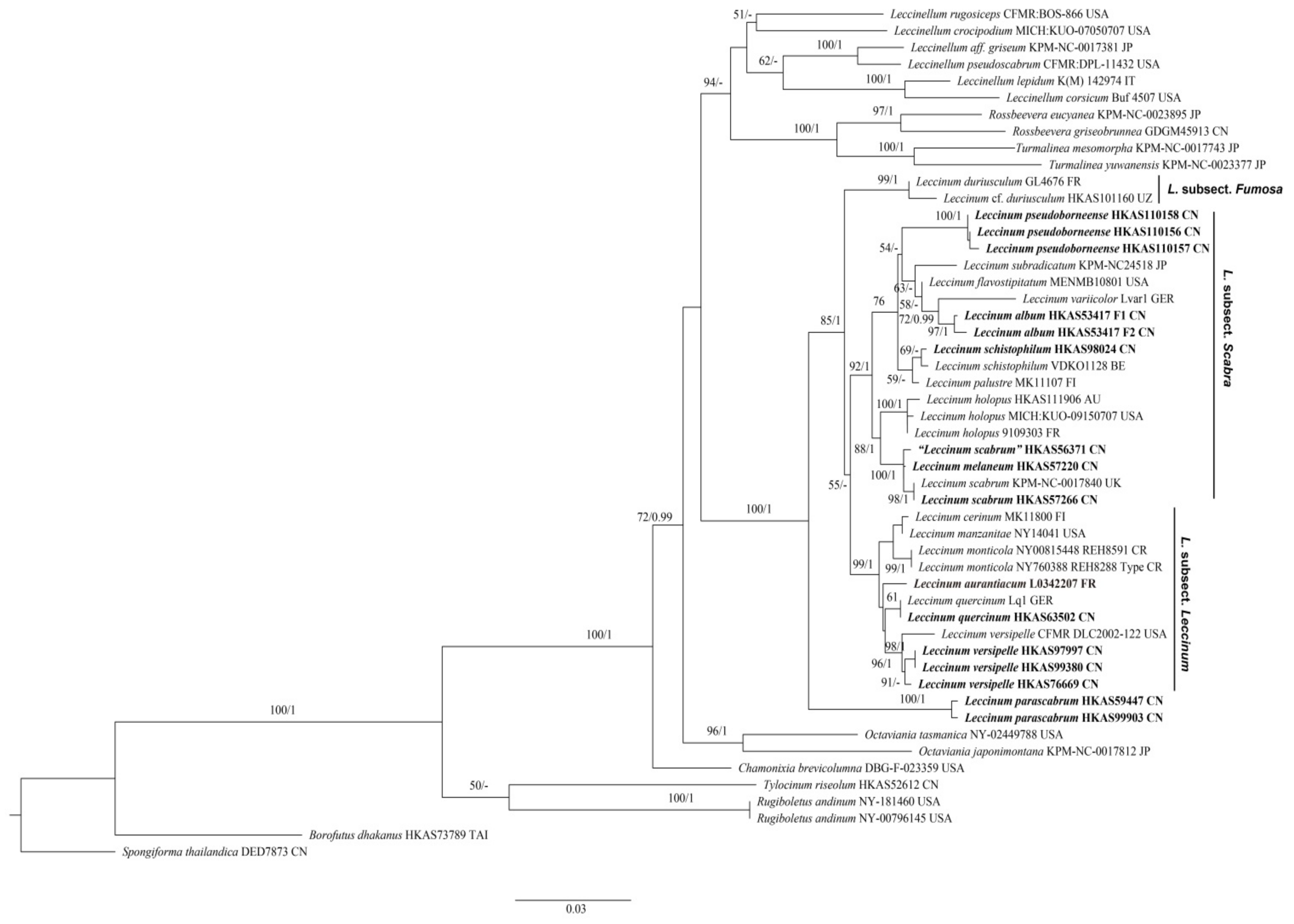
A total of 57 sequences, including fifteen for nrLSU, fifteen for tef1-α, fourteen for rpb2 and thirteen for ITS, were newly generated in this study and aligned with sequences downloaded from GenBank. Sequences retrieved from GenBank and obtained in this study for the multi-locus phylogenetic analyses are listed in Table 1. The multi-locus dataset (Supplementary File S1) contained 122 sequences (49 for nrLSU, 41 for tef1-α, 32 for rpb2), representing 51 samples, and the alignment contained 2195 nucleotide sites, of which 530 were parsimony informative. Borofutus dhakanus Hosen & Zhu L. Yang and Spongiforma thailandica Desjardin, Manfr. Binder, Roekring & Flegel were chosen as the outgroup [71,72]. ML and Bayesian analyses produced very similar estimates of tree topologies, and thus only the tree inferred from ML analysis is displayed (Figure 1). The monophyly of Leccinum was highly supported (BS = 100% and PP = 1) in our analyses. Four main clades were recovered, and three of them correspond to the three known subsections, viz. L. subsect. Leccinum, L. subsect. Fumosa and L. subsect. Scabra of L. sect. Leccinum [14]. Three new species, namely L. album, L. parascabrum and L. pseudoborneense, were revealed in our multi-locus phylogenetic analyses. Leccinum parascabrum formed the remaining clade with BS = 100% and PP = 1, while L. pseudoborneense and L. album nested in L. subsect. Scabra and clustered together with L. flavostipitatum E.A. Dick & Snell, L. subradicatum and L. variicolor with low supported lineage (BS = 54%).
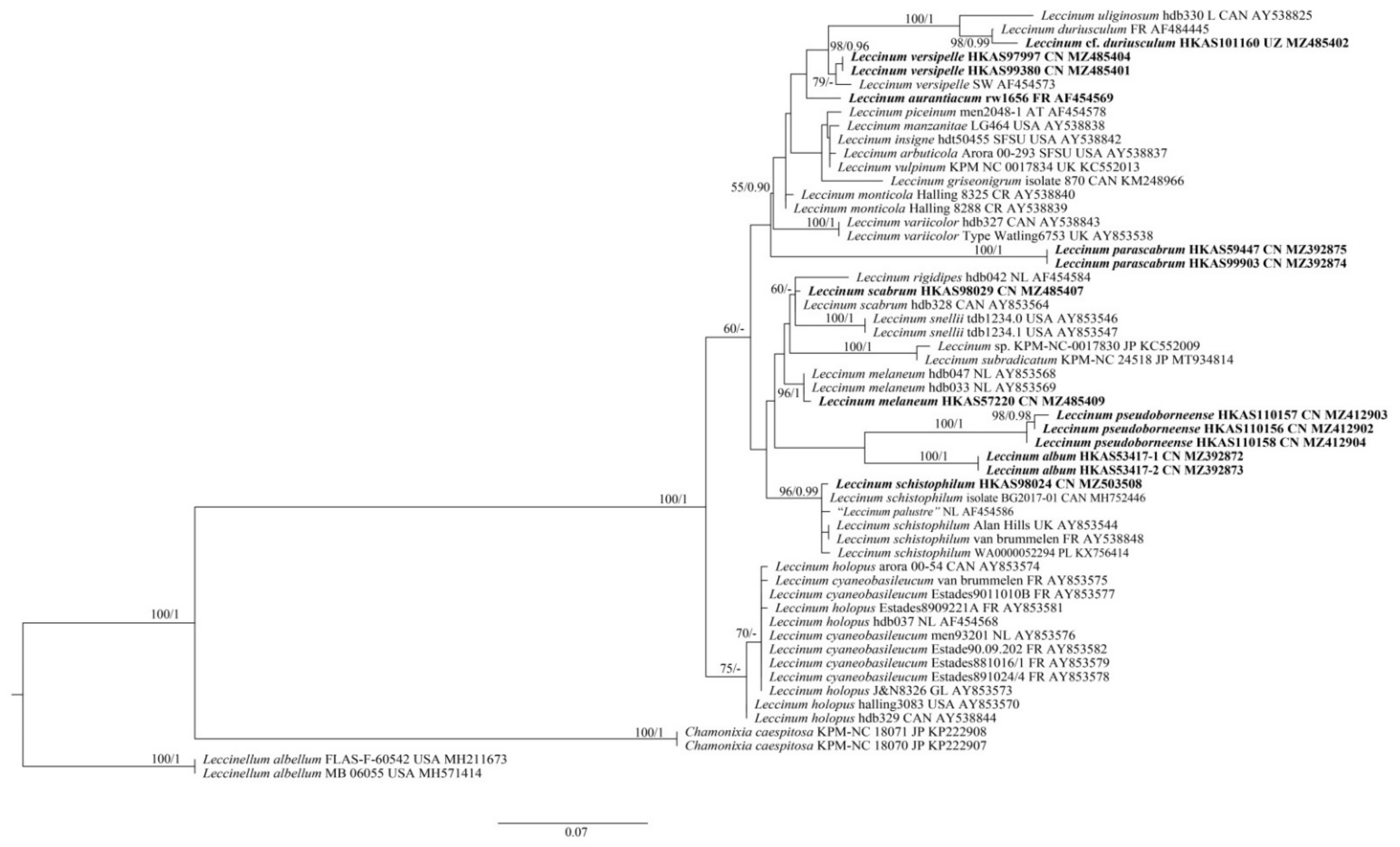
For the ITS dataset, as revealed by den Bakker et al. [57] and our primary analysis, the ITS1 region contains a minisatellite, which is characterized by the repeated presence of CTATTGAAAAG and CTAATAGAAAG core sequences and mutational derivatives. Moreover, some species contain a minisatellite in the ITS2 region, e.g., the newly described species L. album (GenBank Acc. No.: MZ392872 for clone 1 and MZ392873 for clone 2), with a region of 212 bp that consists of tandem repeats (see Supplementary Material for details). Though there is length variation in either the ITS1 or ITS2 spacers, it can also provide some phylogenetic signals. We performed phylogenetic analyses of the ITS dataset. In this dataset (Supplementary File S2), 51 samples were included. The length of the dataset was 1416 bp, of which 377 were parsimony informative. Leccinellum albellum (Peck) Bresinsky & Manfr. Binder was chosen as outgroup. ML and Bayesian analyses also produced very similar estimates of tree topologies, and only the tree inferred from ML analysis is displayed (Figure 2). The monophyly of Leccinum was also well supported (BS = 100% and PP = 1) in our analyses. Three new species viz. L. album, L. parascabrum and L. pseudoborneense) were revealed. Leccinum album is closely related to L. pseudoborneense yet without statistical support, while L. parascabrum forms an independent lineage. Species to which L. parascabrum is phylogenetically related remain as yet unknown.
Our ML and Bayesian analyses of ITS and multi-locus datasets revealed the existence of eight Leccinum species from China, including five known species viz. L. melaneum (Smotl.) Pilát & Dermek, L. quercinum, L. scabrum, L. schistophilum and L. versipelle and three new species viz. L. album, L. parascabrum, and L. pseudoborneense. The final alignments of both datasets were deposited in TreeBASE (S27490).
3.2. Taxonomy
MycoBank: MB 838917.
Diagnosis: This species differs from other species in Leccinum in the combination of the entirely white pileus, the white pileal context not changing color when injured, the white hymenophore staining indistinct greenish blue when hurt, the white stipe coarsely covered with initially white and then darkened verrucose squamules, and the white stipe context always staining greenish blue at the base when injured.
Holotype: CHINA. Hunan Province: Chenzhou, Zhanghua County, Mangshan National Forest Park, E 112°92′, N 24°94′, alt. 850 m, associated with Castanopsis fissa, Cyclobalanopsis glauca, Lithocarpus glabra and Pinus kwangtungensis, 3 September 2007, Y.C. Li1072 (KUN-HKAS53417, GenBank Acc. No.: MZ392872 and MZ392873 for ITS, MW413907 and HQ326880 for nrLSU, MW439267 and HQ326861 for tef1-α, and MW439259 and MW439260 for rpb2).
Etymology: Latin “album” means white, referring to the color of the basidiomata.
Basidiomata small to medium-sized. Pileus 3–5.5 cm in diam., hemispherical when young, subhemispherical to convex or plano-convex when mature, white (1A1) when young, white to cream (2B2–3) when mature; surface covered with concolorous farinose to pubescent squamules; context 5–10 mm thick in the center of pileus, taste mild, white (1A1) to pallid, not changing color when bruised; Hymenophore adnate when young, adnate to slightly depressed around apex of stipe; surface white (1A1), staining indistinct greenish blue (25B5–7) when injured; pores subangular to roundish, 0.3–1.5 mm wide; tubes up to 5 mm long, white to dirty pinkish (13A2), not changing color when bruised. Stipe 8–10 × 0.8–1.2 cm, clavate to subcylindrical, always enlarged downwards; surface white (1A1), densely covered with white (1A1) verrucose squamules, staining light greenish blue at base when injured; context whitish (1A1), staining blue at base when injured; basal mycelium white (1A1), lacking color change when injured.
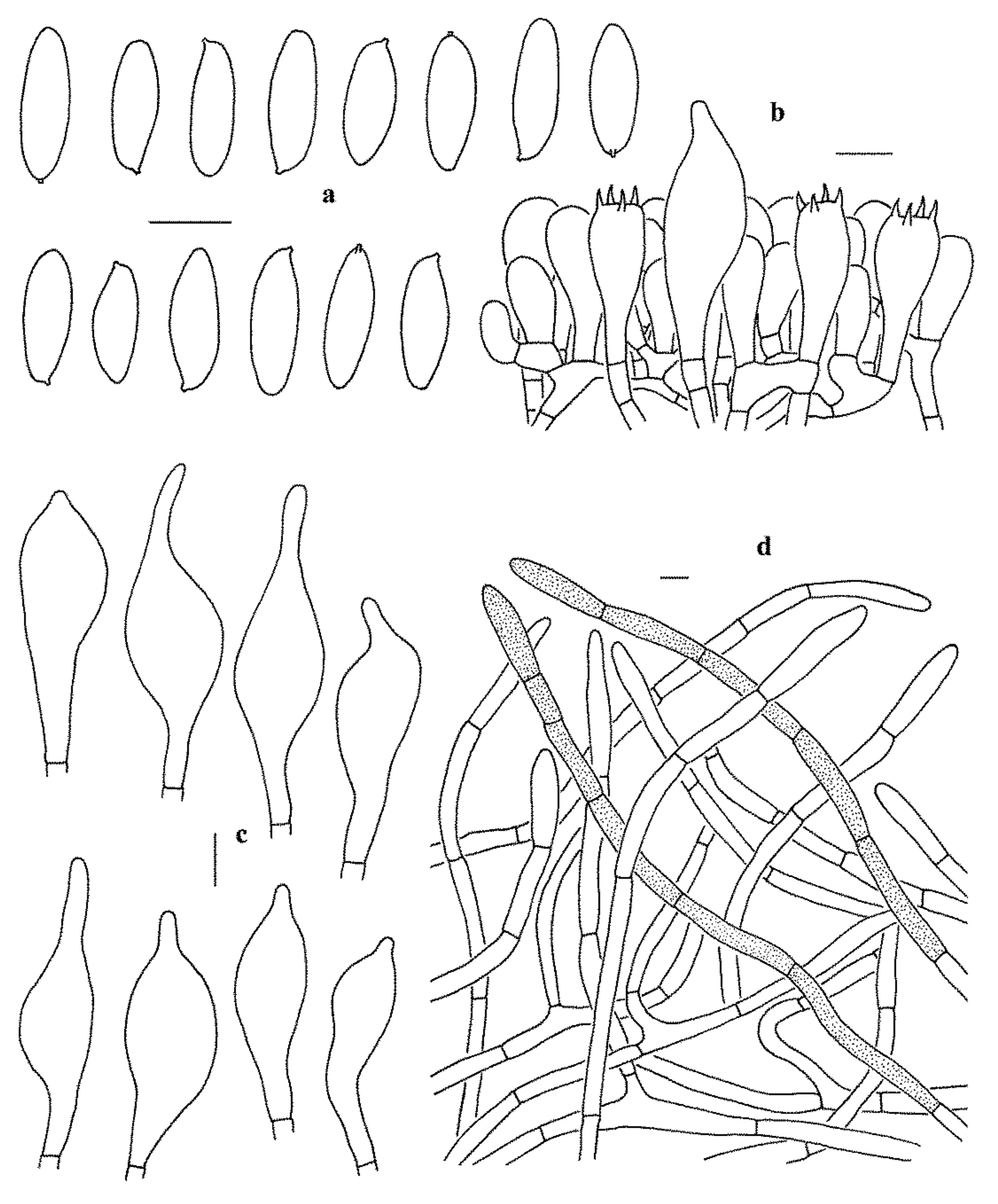
Basidiospores (40/2/1) 15–19 × 5–7 µm, Q = 2.5–3, Qm = 2.75 ± 0.15, subfusiform to narrowly ellipsoid in side view with slight suprahilar depression, subcylindrical to fusiform in ventral view, smooth, somewhat slightly thick-walled (up to 0.5 μm thick), hyaline to yellowish in KOH, brownish yellow to olivaceous brown in Melzer’s Reagent. Basidia 23–33 × 10–13 µm, clavate, 4-spored, hyaline to yellowish in KOH, yellowish to brownish yellow in Melzer’s Reagent. Hymenophoral trama boletoid, hyphae subcylindrical, 4–10 µm wide, hyaline to yellowish in KOH, yellowish to yellow in Melzer’s Reagent. Cheilo- and pleurocystidia 42–60 × 11–17.5 µm, abundant, subfusiform to fusiform, thin-walled, yellowish in KOH, yellowish to brownish yellow in Melzer’s Reagent. Pileipellis a trichoderm, composed of more or less vertically arranged 5–10 µm wide hyphae, hyaline to yellowish in KOH, yellowish to yellow in Melzer’s Reagent. Pileal trama made up of 6–12 µm wide filamentous hyphae, thin-walled, yellowish in KOH, yellowish to brownish yellow in Melzer’s Reagent. Clamp connections absent in all tissues.
Habitat and distribution: Solitary or scattered in tropical forests dominated by plants of the families Fagaceae (Castanopsis fissa, Cyclobalanopsis glauca and Lithocarpus glabra) and Pinaceae (Pinus kwangtungensis or P. armandii); on acidic, humid and loamy soils; distribution insufficiently known, rather rare in China and currently found in central and southeastern China (Hunan and Fujian Provinces).
Additional Specimen examined: CHINA. Fujian Province: Jianning County, E 116°84′, N 26°83′, alt. 900 m, associated with Castanopsis fissa, Cyclobalanopsis glauca and Pinus armandii, 16 July 1971, N.L. Huang 716 (KUN-HKAS39522).
Commentary: Leccinum album is characterized by the white pileus, the white hymenophore staining indistinct greenish blue when hurt, the white stipe densely covered with initially white and then darkened scabrous squamules, the white context in pileus not changing color when injured, and the white context in stipe unchanging or only staining distinct greenish blue at base when injured. Morphologically, L. album is close to L. holopus, L. cyaneobasileucum Lannoy & Estadès and Le. albellum (Peck) Bresinsky & Manfr. Binder in similar pileus colors. However, L. holopus, originally described from Europe (Germany), differs from L. album in its medium to large basidiomata (pileus 4–10 cm wide), becoming more viscid pileus with age, pure white or dirty white to pale buff or pale pallid pileus always with a glaucous green tinge, long hymenophoral tubes measuring 9–15 mm long, narrow and subcylindrical hymenial cystidia measuring 30–50 × 7.5–12.5 µm, narrow pileipellis hyphae measuring 3.5–5 µm wide, and association with trees of the genus Betula (Betulaceae) [80,81,82]. Leccinum cyaneobasileucum, originally described from France, is different from L. album in its white or greyish brown to light brown pielus, woolly stipe surface, slender basidiospores with Qm ≥ 3, relatively narrow hymenial cystidia measuring 32–44 × 5.5–7.5 µm, narrow pileipellis hyphae measuring 2–6.5 µm wide, and association with trees of the genus Betula [83]. Leccinellum albellum, originally described from New York, is characterized by its basidiomata not changing color when bruised and narrow basidiospores measuring 13–20 × 4–6 µm [16,17,30].
Phylogenetically, L. album is related to L. variicolor and L. pseudoborneense in the analyses of the multi-locus and ITS datasets, respectively (Figure 1 and Figure 2). However, L. variicolor differs from L. album in its white to grey or cream pileal context staining vinaceous to brown when bruised, white stipe context staining pink to coral red in the upper part and green-blue in the lower part when bruised and association with plants of Betula [81]. Leccinum pseudoborneense is different from L. album in its pale brown to dark brown pileus, white context in pileus and stipe staining blue when bruised, narrow basidiospores measuring (11) 12–19 (20) × 4–5 (6) µm, narrow hymenial cystidia measuring 28–40 × 4–10 µm, and distribution in southwestern China.
MycoBank: MB 838916.
Diagnosis: This species differs from other species in Leccinum by its initially reddish brown to chestnut-brown and then brown to pale brownish or even dirty white pileus, white pileal context lacking color change when injured, white to pallid and then light brown hymenophore lacking color change when injured, and the white stipe context staining greenish blue at the base when injured.
Holotype: CHINA. Hunan Province: Chenzhou, Zhanghua County, Mangshan National Forest Park, E 112°92′, N 24°94′, alt. 1100 m, associated with Castanopsis fissa, Lithocarpus glabra and Pinus kwangtudgensis, 12 September 2016, G. Wu 1784 (KUN-HKAS99903, GenBank Acc. No.: MZ392874 for ITS, MW413911 for nrLSU, MW439271 for tef1-α, and MW439264 for rpb2).
Etymology: Latin “parascabrum” refers to its similarity to L. scabrum.
Basidiomata small to medium-sized. Pileus 2.5–12.5 cm in diam., hemispherical when young, subhemispherical to convex or applanate when mature, reddish brown (12E8) to chestnut-brown (8C7–8) when young, brown (6C6) to pale brownish (7D7–8) or even dirty white (6A2) when mature; surface tomentose; context 6–13 mm thick in the center, white (1A1), not changing color when bruised; Hymenophore adnate when young, adnate to slightly depressed around apex of stipe; surface white to pallid (1A1) when young, and becoming light brown (6B4) when mature, not changing color when injured; tubes 6–14 mm long, 0.5–1.5 mm wide, creamy white (1A1), not changing color when bruised. Stipe 12–14 × 1.1–2.2 cm, clavate, swollen downwards, always staining greenish blue at base when injured; surface white (1A1), covered with initially white (1A1) to light beige (5A4) and then brownish (7D8) squamules; context white (1A1), staining greenish blue (25B6–7) at base when injured; basal mycelium white (1A1).
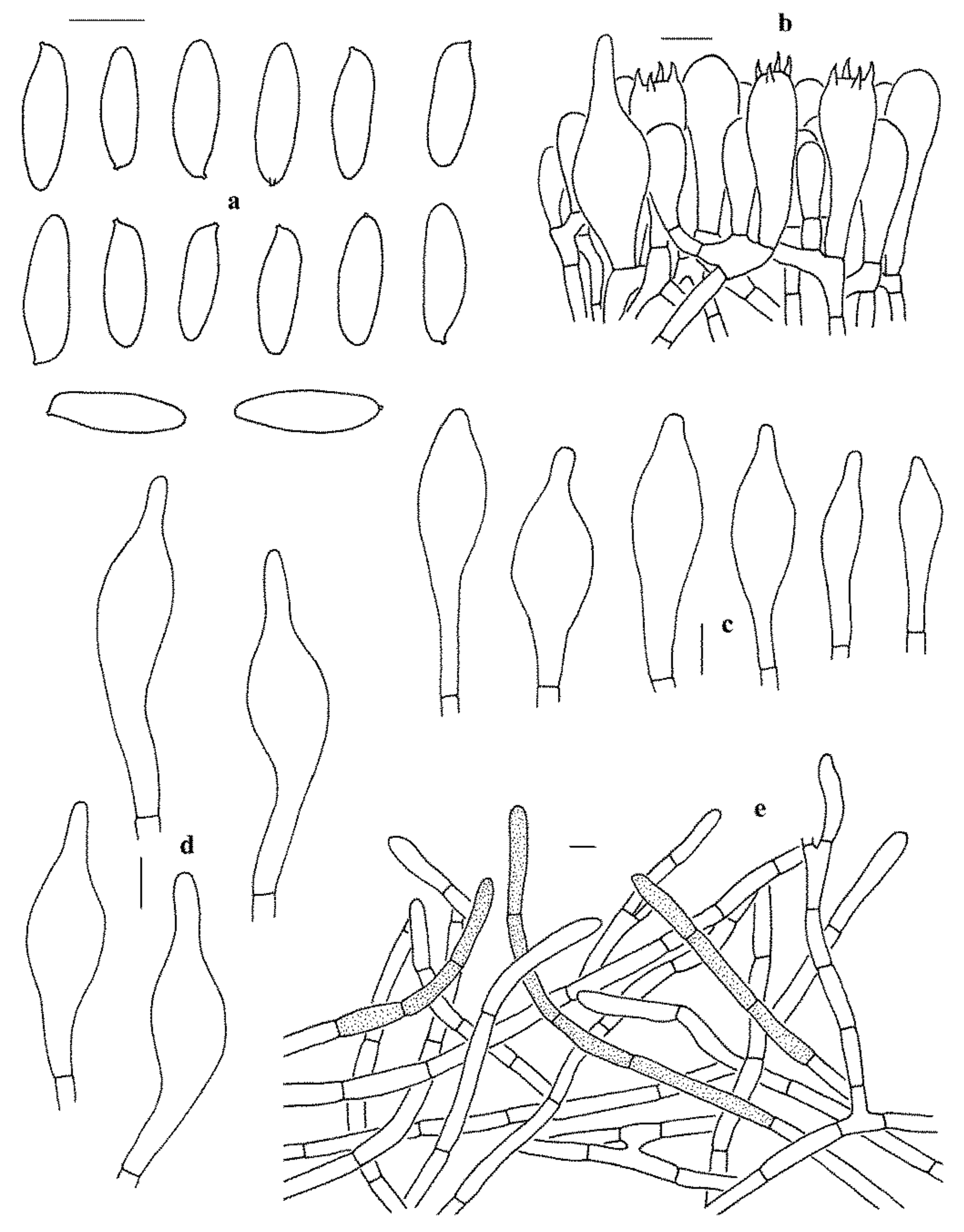
Basidiospores (80/2/2) 16–20 (–21) × 5–6 µm, Q = 3.2–3.8, Qm = 3.43 ± 0.18, subfusiform to fusiform, slightly thick-walled (up to 0.5 μm thick), yellowish to yellowish brown in KOH, yellow to yellow-brown in Melzer’s Reagent. Basidia 24–33 × 8–12 µm, clavate, 4-spored, hyaline to yellowish in KOH, yellowish to yellow in Melzer’s Reagent. Hymenophoral trama boletoid, hyphae cylindrical, 3–7 µm wide, hyaline to yellowish in KOH, yellowish to yellow in Melzer’s Reagent. Cheilo- and pleurocystidia 34–68 × 7.5–16 µm, abundant, subfusiform to fusiform, thin-walled, yellowish to pale yellowish brown in KOH, yellowish brown to brown in Melzer’s Reagent. Pileipellis a trichoderm, composed of 5–9 µm wide filamentous hyphae, yellowish to pale brownish in KOH. Pileal trama made up of 5–10 µm wide filamentous hyphae, thin-walled, hyaline to yellowish in KOH, yellowish to brownish yellow in Melzer’s Reagent. Clamp connections absent in all tissues.
Habitat and distribution: Solitary or scattered in tropical forests dominated by plants of the families Fagaceae (Lithocarpus glabra, Castanopsis fissa and Ca. hystrix) and Pinaceae (Pinus kwangtudgensis or P. yunnanensis.); on acidic or slightly alkaline, loamy soils; distribution insufficiently known, rather rare in China, currently known from central and southwestern China (Hunan and Yunnan Provinces).
Additional Specimen examined: CHINA. Yunnan Province: on the way from Tengchong County to Longling County, E 98°59′, N 24°81′, alt. 2010 m, associated with Lithocarpus glabra, Castanopsis hystrix and Pinus yunnanensis, 19 July 2009, Y.C. Li 1700 (KUN-HKAS59447, GenBank Acc. No.: MZ392875 for ITS, MW413912 for nrLSU, MW439272 for tef1-α, and MW439265 for rpb2).
Commentary: Leccinum parascabrum is characterized by the initially reddish brown to chestnut-brown and later brown to pale brownish or even dirty white pileus, the white pileal context not changing color when injured, the white to pallid and then light brown hymenophore not changing color when injured, the white stipe context with greenish blue color change at the base when injured, and the relatively large basidiospores measuring 16–20 (–21) × 5–6 µm, Q = 3.2–3.8. Leccinum parascabrum generally shares the similar colors of pileus and hymenophore, and the similar slender stems with L. duriusculum, L. griseonigrum A.H. Sm., Thiers & Watling, L. scabrum, L. uliginosum A.H. Sm. & Thiers and Le. pseudoscabrum (Kallenb.) Mikšík. However, L. duriusculum, originally described from Europe, can be distinguished from L. parascabrum by its pale grey-brown to dark greyish or reddish brown pileus, white context staining violaceous pink when bruised but yellow-green to blue-green in the base of stipe, relatively small basidiospores measuring 11.5–15.5 × 4.5–6 µm [84]. Leccinum griseonigrum, originally described from North America, differs from L. parascabrum in its avellaneous to dingy cinnamon-buff pileus, white pileal context staining blue when bruised, relatively small basidiospores measuring 13–16 × 4–5.5 µm, and association with trees of the genus Populus [16]. Leccinum scabrum differs from L. parascabrum in its wrinkled pileus, pale white hymenophore, pinkish discoloration when injured, and never bluish color change at the base of stipe [13,14,81]. Leccinum uliginosum, originally described from North America, is different from L. parascabrum in its dark fuscous to drab-grey pileus, white context in pileus becoming reddish and then fuscous when bruised, relatively small basidiospores measuring 14–18 × 3.5–5 µm, and small and inconspicuous hymenial cystidia [17]. Leccinellum pseudoscabrum differs from L. parascabrum in its initially red to purplish brown and then blackish brown context color change when injured, and the palisadoderm pileipellis composed of subglobose cells [14]. Leccinum parascabrum also shares the similar colors of pileus and hymenophore and the bluish color change at the base of stipe with L. variicolor. However, L. variicolor is different from L. parascabrum in its white to grey or cream pileal context staining vinaceous to brown when bruised, white stipe context staining pink to coral red in the upper part and green-blue in the lower part when bruised, relatively small basidiospores measuring (10) 13.5–17.5 (–20.0) × 5.0–6.5 (7.0) μm with Q = 2.4–3.1, and association with plants of Betula sp. [81]. In our phylogenetic analysis of the multi-locus and ITS datasets (Figure 1 and Figure 2), L. parascabrum formed independent clades within Leccinum. It might represent a distinct section or subsection. However, formal change of the infrageneric division of this clade should await more molecular and morphological data from additional taxa. Species to which it is phylogenetically related remain as yet unknown.
MycoBank: MB 838915.
Diagnosis: This species differs from other species in Leccinum in its nearly glabrous and pale brown to dark brown pileus, white context in pileus lacking color change when injured, white context in stipe staining blue when bruised, initially white and then brown hymenophore not changing color when injured, white stipe covered with ochraceous to dark brown squamules, and trichodermal pileipellis composed of 3–6 μm wide interwoven hyphae.
Holotype: CHINA. Yunnan Province: Xishuangbanna, Menghai County, Bada Town, E 100°12′, N 21°83′, alt. 1900 m, associated with Castanopsis calathiformis, Castanopsis indica and Lithocarpus truncatus, 22 June 2020, G.S. Wang 947 (KUN-HKAS110156, GenBank Acc. No.: MZ412902 for ITS, MW413908 for nrLSU, MW439268 for tef1-α, and MW439261 for rpb2)
Etymology: Latin “pseudo” = false, “borneense” = L. borneense, “pseudoborneense” is proposed because this species is similar to the species L. borneense originally described from Malaysia.
Basidiomata small to medium-sized. Pileus 4–10 cm diam, subhemispherical to convex or plano-convex; surface nearly glabrous, viscid when wet, pale brown (6D6–C5) to dark brown (6F6–E5); context 5–10 mm thick in the center, white (1A1), not changing color when bruised; Hymenophore adnate to depressed around apex of stipe; white (1A1) to pallid when young and becoming brown (6B5) when mature, not changing color when injured. Tubes 4–10 mm long, creamy white (1A1) when young, and becoming brownish yellow (5C7–8) when mature, not changing color when bruised; pores fine, no more than 1 mm wide. Stipe 10–15 × 2.1–2.9 cm, clavate, always swollen downwards; surface white (1A1), covered with ochraceous (2B3–5) to dark brown (6E7) squamules, staining asymmetric blue (23E7) when injured; context white (1A1), staining blue (23E7) when injured; basal mycelium white (1A1).
Basidiospores (100/5/5) (11–) 12–19 (–20) × 4–5 (–6) µm, Q = (2.75–) 3–3.58 (–3.6), Qm = 3.31 ± 0.16, subfusiform to ellipsoid, slightly thick-walled (up to 0.5 μm thick), yellowish brown to olive brown in KOH, yellow-brown to dark olive-brown in Melzer’s Reagent. Basidia 18–30 × 8–9 µm, clavate, 4-spored, hyaline to yellowish in KOH, yellowish to brownish yellow in Melzer’s Reagent. Hymenophoral trama boletoid, hyphae cylindrical, 3–6 µm wide, hyaline to yellowish in KOH, yellowish to brownish yellow in Melzer’s Reagent. Cheilo- and pleurocystidia 28–40 × 4–10 µm, abundant, subfusiform to fusiform, thin-walled, yellowish to brownish yellow in KOH, brownish to yellow-brown in Melzer’s Reagent. Pileipellis a trichoderm, composed of more or less vertically arranged 5–12 µm wide filamentous hyphae, yellowish brown to brownish in KOH, brown to dark brown in Melzer’s Reagent. Pileal trama made up of 6–12 µm wide filamentous hyphae, thin-walled, hyaline to yellowish in KOH, yellowish to yellow in Melzer’s Reagent. Clamp connections absent in all tissues.
Habitat and Distribution: Scattered in tropical forests dominated by plants of the families Fagaceae (Castanopsis calathiformis, Ca. orthacantha, Ca. indica, Lithocarpus truncatus, Li. mairei and Quercus griffithii); on acidic, loamy or mossy, humid soils; moderately common in southwestern China (Yunnan Province).
Additional specimens examined: CHINA. Yunnan Province: Xishuangbanna, Menghai County, Bada Township, E 100°13′, N 21°84′, alt. 1900 m, associated with Castanopsis calathiformis, Ca. indica and Lithocarpus truncatus, 22 June 2020, G.S. Wang 960 (KUN-HKAS110157, GenBank Acc. No.: MZ412903 for ITS, MW413909 for nrLSU, MW439269 for tef1-α, and MW439262 for rpb2), the same location, 22 June 2020, G.S. Wang 965 (KUN-HKAS110158, GenBank Acc. No.: MZ412904 for ITS, MW413910 for nrLSU, MW439270 for tef1-α, and MW439263 for rpb2); Nanjian County, Gonglang Town, Huangcaoping, E 100°30′, N 24°54′, alt. 1200 m, associated with Castanopsis orthacantha, Lithocarpus mairei and Quercus griffithii, 30 June 2015, K. Zhao 773 (KUN-HKAS92401, GenBank Acc. No.: MZ536632 for nrLSU, MZ543307 for tef1-α, and MZ543309 for rpb2); Jinghong County, Dadugang Town, E 100°25′, N 21°26′, alt. 600 m, associated with Castanopsis indica and Lithocarpus truncatus, 30 June 2014, K. Zhao 476 (KUN-HKAS89139, GenBank Acc. No.: MZ536631 for nrLSU, MZ543306 for tef1-α, and MZ543308 for rpb2).
Commentary: Leccinum pseudoborneense is characterized by the nearly glabrous and pale brown to dark brown pileus, the white context in pileus not changing color when injured, the white context in stipe staining blue when bruised, the initially white and then brown hymenophore not changing color when injured, the white stipe covered with ochraceous to dark brown squamules, and the trichodermal pileipellis composed of 3–6 μm wide interwoven hyphae. Leccinum pseudoborneense is similar to L. borneense (Corner) E. Horak, originally described from Malaysia, in that they share a brown pileus, bluish color change of the context in stipe when bruised, and similar size of basidiospores. However, L. borneense differs from L. pseudoborneense in its yellow to olive yellow hymenophore staining blue when hurt, pale yellow to yellow pileal context staining blue when hurt, and deep yellow context in stipe staining blue but sometimes with reddish tint at base when injured [6,84]. Leccinum pseudoborneense is phylogenetically close to L. album in our phylogenetica analyses (Figure 1 and Figure 2). However, L. album has a white basidioma, white hymenophore staining indistinctly greenish blue when hurt, white context in pileus not changing color when injured, white context in stipe unchanging or only staining distinctly greenish blue at base when injured, and relatively broad basidiospores measuring 15–19 × 5–7 µm.
4. Discussion
The genus Leccinum was defined and recognized variously by different mycologists. In an early molecular study, Leccinum was shown to be polyphyletic and proposed to be restricted to the sections Leccinum and Scabra by Binder and Besl [4]. Subsequently, Bresinsky and Besl [32] erected a genus Leccinellum Bresinsky & Manfr. Binder, to accommodate L. section Luteoscabra, including species with yellow hymenophores and/or context. In this study, the phylogenetic inferences based on the multi-locus dataset of nrLSU, tef1-α and rpb2 largely coincide with those of Binder and Besl [4], Bresinsky and Besl [32] and den Bakker et al. [33]. Thus, we adopt the treatment of Bakker et al. [33] and treat Leccinum in a strict circumscription, which only includes species of L. sect. Leccinum (Singer’s infrageneric classification with L. sect. Scabra merged to this section). Species in Leccinum are characterized by the white context lacking color changes or staining blue, gray or reddish tints when injured and the cutis-like pileipellis composed of interwoven filamentous hyphae
Eleven Leccinum species with specimen citations have been reported from China before this study, of which five species (L. ambiguum, L. atrostipitatum, L. olivaceopallidum, L. potteri and L. subgranulosum) were originally described from North America, five species (L. aurantiacum, L. holopus, L. roseofractum, L. scabrum and L. versipelle) were originally described from Europe, and only one taxon (L. subleucophaeum var. minimum) was originally described from China. Our molecular phylogenetic analyses along with morphological studies identified the existence of L. quercinum, L. scabrum, L. subleucophaeum var. minimum and L. versipelle in China. The distribution of other reported species have not yet been found, based on morphological and/or molecular data. In addition, three species new to science (L. album, L. parascabrum and L. pseudoborneense) and two species new to China (L. melaneum and L. schistophilum) were revealed in our study, based on molecular and morphology evidence. In conclusion, there are nine species of Leccinum in China.
Most species of Leccinum exhibit strong mycorrhizal host specificity. The host specificity along with climate type and edaphic factors appear to be important factors determining the distribution of different species. In China, L. melaneum, L. scabrum, L. schistophilum and L. versipelle are found in temperate forests and associated with plants of Betula platyphylla on acidic soils. Leccinum album, L. parascabrum, L. pseudoborneense and L. subleucophaeum var. minimum are found in tropical forests and associated with plants of Fagaceae (Castanopsis calathiformis, Ca. hystrix, Ca. indica, Ca. orthacantha, Cyclobalanopsis glauca, Lithocarpus mairei, Li. truncatus and Quercus griffithii) and/or Pinaceae (Pinus kwangtudgensis and P. yunnanensis) on acidic soils. It is noteworthy that L. parascabrum can be found in acidic or slightly alkaline habitats. Leccinum versipelle is found in subtropical forests and is associated with plants of Populus yunnanensis on acidic soils. The combination of the color of basidioma, the morphology of pileal surface, the size of basidiospores, the morphology of stipe, the color changes when injured, the climate type, the edaphic factors and the host preferences is very important in distinguishing species in this genus.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/jof7090732/s1, Alignment S1: alignment of multi-locus dataset; Alignment S2: alignment of ITS dataset.
Author Contributions
Conceptualization: Z.L.Y., Y.-C.L. and X.M.; field sampling: Z.L.Y., Y.-C.L., G.W. and G.-S.W.; molecular experiments and data analysis: X.M. and P.-M.W.; original draft—writing: X.M.; original draft—review and editing: Y.-C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Nos. 31872618, 31750001, 32070024, 31970015), the Natural Science Foundation of Yunnan Province (2018FB027), the Ten Thousand Talents Program of Yunnan (YNWR-QNBJ-2018-125), and the Key Research Program of Frontier Sciences, CAS (QYZDY-SSW-SMC029).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: https://www.ncbi.nlm.nih.gov/; http://www.mycobank.org/; https://www.treebase.org/treebase-web/home.html, accessed on 26 August 2021.
Acknowledgments
The authors thank X. H. Wang, Z. W. Ge, B. Feng and Q. Cai, Kunming Institute of Botany, CAS, for providing samples and/or related literatures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, A.H.; Thiers, H.D.; Watling, R. Notes on species of Leccinum. I. Additions to section Leccinum. Lloydia 1968, 31, 252–267. [Google Scholar]
- Engel, H.; Krieglsteiner, G.J. Rauhstielröhrlinge–die Gattung Leccinum in Europa; Hilmar Schneider: Coburg, Germany, 1978; p. 76. [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986; pp. 1–981. [Google Scholar]
- Binder, M.; Besl, H. 28S rDNA sequence data and chemotaxonomical analyses on the generic concept of Leccinum (Boletales). In Micologia; Associazone Micologica Bresadola, Ed.; Grafica Sette: Brescia, Italy, 2000; pp. 75–86. [Google Scholar] [CrossRef]
- Kuo, M.; Ortiz-Santana, B. Revision of leccinoid fungi, with emphasis on North American taxa, based on molecular and morphological data. Mycologia 2020, 112, 1–15. [Google Scholar] [CrossRef]
- Corner, E.J.H. Boletus in Malaysia; Government Printer: Singapore, 1972; p. 263.
- Nagasawa, E. A preliminary checklist of the Japanese agaricales, 1: The boletineae. Rep. Tottori. Mycol. Inst. 1997, 35, 39–78. [Google Scholar]
- Takahashi, H. Five new species of the Boletaceae from Japan. Mycoscience 2007, 48, 90–99. [Google Scholar] [CrossRef]
- Terashima, Y.; Takahashi, H.; Taneyama, Y. The Fungal Flora in Southwestern Japan: Agarics and Boletes; Tokai University Press: Tokyo, Japan, 2016; pp. 1–303. [Google Scholar]
- Katumoto, K. List of Fungi Recorded in Japan; Kanto Branch of the Mycological Society of Japan: Kyoto, Japan, 2010; pp. 1–954. [Google Scholar]
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Li, T.H.; Song, B. Chinese boletes: A comparison of boreal and tropical elements. In Proceedings of the Tropical Mycology 2000, the Millennium Meeting on Tropical Mycology (Main Meeting 2000), British Mycological Society & Liverpool John Moores University, Liverpool, UK, 25–29 April 2000. [Google Scholar]
- Gray, S.F.; Gray, J.E.; Shury, J. A Natural Arrangement of British Plants; Cradock and Joy: London, UK, 1821; pp. 1–647. [Google Scholar]
- Den Bakker, H.C.; Noordeloos, M.E. A revision of European species of Leccinum Gray and notes on extralimital species. Persoonia 2005, 18, 511–587. [Google Scholar]
- Smith, A.H.; Thiers, H.D.; Watling, R. A preliminary account of the North American species of Leccinum, Section Leccinum. Mich. Bot. 1966, 5, 131–178. [Google Scholar]
- Smith, A.H.; Thiers, H.D.; Watling, R. A Preliminary account of the North American species of Leccinum, Sections Luteoscabra and Scabra. Mich. Bot. 1967, 6, 107–179. [Google Scholar]
- Smith, A.H.; Thiers, H.D. The Boletes of Michigan; University of Michigan Press: Ann Arbor, MI, USA, 1971; p. 428. [Google Scholar]
- Thiers, H.D. California Boletes. IV. The Genus Leccinum. Mycologia 1971, 63, 261–276. [Google Scholar] [CrossRef]
- Both, E. The Boletes of North America, a Compendium; Buffalo Museum of Science: Buffalo, NY, USA, 1993; pp. 1–436. [Google Scholar]
- Ortiz-Santana, B.; Halling, R.E. A new species of Leccinum (Basidiomycota, Boletales) from Belize. Brittonia 2009, 61, 172–174. [Google Scholar] [CrossRef]
- Halling, R.E.; Mueller, G.M. Leccinum (Boletaceae) in Costa Rica. Mycologia 2003, 95, 488–499. [Google Scholar] [CrossRef]
- Halling, R.E.; Mueller, G.M. Common Mushrooms of the Talamanca Mountains, Costa Rica; New York Botanical Garden Press: New York, NY, USA, 2005; pp. 1–197. [Google Scholar]
- Halling, R.E. New Leccinum from Costa Rica. Kew Bull. 1999, 54, 747–753. [Google Scholar] [CrossRef]
- Halling, R.E. A synopsis of Colombian boletes. Mycotaxon 1989, 34, 93–113. [Google Scholar]
- Assyov, B.; Denchev, C.M. Preliminary checklist of Boletales s. str. in Bulgaria. Mycol. Balc. 2004, 1, 195–208. [Google Scholar]
- Segedin, B.P.; Pennycook, S.R. A nomenclatural checklist of agarics, boletes, and related secotioid and gasteromycetous fungi recorded from New Zealand. N. Z. J. Bot. 2001, 39, 285–348. [Google Scholar] [CrossRef]
- Watling, R. Australian boletes: Their diversity and possible origins. Aus. Syst. Bot. 2001, 14, 407–416. [Google Scholar] [CrossRef]
- Segedin, P.B. An annotated checklist of Agarics and Boleti recorded from New Zealand. N. Z. J. Bot. 1987, 25, 185–215. [Google Scholar] [CrossRef]
- Mcnabb, R.F.R. The Boletaceae of New Zealand. N. Z. J. Bot. 1968, 6, 137–176. [Google Scholar] [CrossRef]
- Peck, C.H. Report of the botanist. Ann. Rep. N. Y. St. Mus. Nat. Hist. 1888, 41, 51–122. [Google Scholar]
- Šutara, J. The delimitation of the genus Leccinum. Czech. Mycol. 1989, 43, 1–12. [Google Scholar] [CrossRef]
- Bresinsky, A.; Besl, H. Beiträge zu einer Mykoflora Deutschlands: Schlüssel zur Gattungsbestimmung der Blätter-, Leisten- und Röhrenpilze mit Literaturhinweisen zur Artbestimmung. Regensb. Mykol. Schr. 2003, 11, 1–236. [Google Scholar]
- Den Bakker, H.C.; Zuccarello, G.C.; Kuyper, T.W.; Noordeloos, M.E. Evolution and host specificity in the ectomycorrhizal genus Leccinum. New Phytol. 2004, 163, 201–215. [Google Scholar] [CrossRef]
- Zang, M. Notes on the boletales from Eastern Himalayas and adjacent areas of China. Acta Bot. Yunnanica 1986, 8, 1–22. [Google Scholar]
- Wu, G.; Zhao, K.; Li, Y.C.; Zeng, N.K.; Feng, B.; Halling, R.E.; Yang, Z.L. Four new genera of the fungal family Boletaceae. Fungal Divers. 2016, 81, 1–24. [Google Scholar] [CrossRef]
- Wu, K.; Wu, G.; Yang, Z.L. A taxonomic revision of Leccinum rubrum in subalpine coniferous forests, Southwestern China. Acta Edulis Fungi 2020, 72, 92–100. [Google Scholar]
- Halling, R.E.; Fechner, N.; Nuhn, M.; Osmundson, T.; Soytong, K.; Arora, D.; Binder, M.; Hibbett, D. Evolutionary relationships of Heimioporus and Boletellus (Boletales) with an emphasis on Australian taxa including new species and new combinations in Aureoboletus, Hemileccinum, and Xerocomus. Aust. Syst. Bot. 2015, 28, 1–22. [Google Scholar] [CrossRef]
- Chiu, W.F. The boletes of Yunnan. Mycologia 1948, 40, 199–231. [Google Scholar] [CrossRef]
- Mao, X.L.; Jiang, C.P.; Ouzhu, C.W. Economic Macrofungi of Tibet; Beijing Science and Technology Press: Beijing, China, 1993; pp. 1–651. (In Chinese) [Google Scholar]
- Mao, X.L. The Economic Fungi in China; Science Press: Beijing, China, 1998; pp. 1–762. (In Chinese) [Google Scholar]
- Mao, X.L. The Macrofungi of China; Henan Science and Technology Press: Zhengzhou, China, 2000; pp. 1–719. (In Chinese) [Google Scholar]
- Wu, X.L.; Zang, M.; Xia, T.Y. Coloured Illustrations of the Ganodermataceae and other Fungi; Guizhou Science & Technology Press: Guiyang, China, 1997; pp. 1–358. (In Chinese) [Google Scholar]
- Ying, J.Z.; Wen, H.A.; Zong, Y.C. The Economic Macromycetes from Western Sichuan; Science Press: Beijing, China, 1994; pp. 1–399. (In Chinese) [Google Scholar]
- Bi, Z.S.; Zheng, G.Y.; Li, T.H.; Wang, Y.Z. Macrofungus Flora of the Mountainous District of North. Guangdong; Guangdong Science & Technology Press: Guangzhou, China, 1990; pp. 1–450. (In Chinese) [Google Scholar]
- Yuan, M.S.; Sun, P.Q. Sichuan Mushrooms; Sichuan Science and Technology Press: Chengdu, China, 1995; pp. 1–737. (In Chinese) [Google Scholar]
- Bi, Z.S.; Li, T.H.; Zhang, W.M.; Song, B. A Preliminary Agaric Flora of Hainan Province; Guangdong Higher Education Press: Guangzhou, China, 1997; pp. 1–388. (In Chinese) [Google Scholar]
- Yeh, K.W.; Chen, Z.C. The boletes of Taiwan, I. Taiwania 1980, 25, 166–184. [Google Scholar]
- Bi, Z.S.; Li, T.H.; Zheng, G.Y.; Li, C. Basidiomycetes from Dinghu Mountain of China. III. Some species of Boletaceae (2). Acta Myco. Sinica 1984, 3, 199–206. (In Chinese) [Google Scholar]
- Song, B.; Li, T.H.; Wu, X.L.; Shen, Y.H. A preliminary review on Boletales resources in Yunnan, Guizhou and Guangxi, China. Guizhou Sci. 2004, 22, 91–97. [Google Scholar]
- Fu, S.Z.; Wang, Q.B.; Yao, Y.J. An annotated checklist of Leccinum in china. Mycotaxon 2006, 96, 47–50. [Google Scholar]
- Li, T.H.; Song, B. Bolete species known from China. Guizhou Sci. 2003, 21, 78–86. [Google Scholar]
- Li, Y.C.; Feng, B.; Yang, Z.L. Zangia, a new genus of Boletaceae supported by molecular and morphological evidence. Fungal Divers. 2011, 49, 125–143. [Google Scholar] [CrossRef]
- Halling, R.E.; Nuhn, M.E.; Fechner, N.; Osmundson, T.; Soytong, K.; Arora, D.; Hibbet, D.S.; Binder, M. Sutorius: A new genus for Boletus eximius (Boletaceae). Mycologia 2012, 104, 951–961. [Google Scholar]
- Halling, R.E.; Nuhn, M.E.; Osmundson, T.; Fechner, N.; Trappe, J.M.; Soytong, K.; Arora, D.; Hibbett, D.S.; Binder, M. Affinities of the Boletus chromapes group to Royoungia and the description of two new genera, Harrya and Australopilus. Austral. Syst. Bot. 2012, 25, 418–431. [Google Scholar] [CrossRef]
- Nuhn, M.E.; Binder, M.; Taylor, A.F.S.; Halling, R.E.; Hibbett, D.S. Phylogenetic overview of the boletineae. Fungal Biol. 2013, 117, 479–511. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Den Bakker, H.C.; Gravendeel, B.; Kuyper, T.W. An ITS Phylogeny of Leccinum and an Analysis of the Evolution of Minisatellite-like Sequences within ITS1. Mycologia 2004, 96, 102–118. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1983; pp. 1–252. [Google Scholar]
- Zhou, M.; Dai, Y.C.; Vlasák, J.; Yuan, Y. Molecular Phylogeny and Global Diversity of the Genus Haploporus (Polyporales, Basidiomycota). J. Fungi 2021, 7, 96. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bullet. 1987, 19, 11–15. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bact. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with rpb1 and rpb2 nucleotide sequences (Inocybe, Agaricales). Mol. Phylogenet. Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols. A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Wu, G.; Feng, B.; Xu, J.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analyses program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web-servers. Syst. Biol. 2008, 75, 758–771. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of the AIC and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. Bayesian analysis of molecular evolution using MrBayes. In Statistical Methods in Molecular Evolution; Nielsen, R., Ed.; Springer: New York, NY, USA, 2005; pp. 183–232. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Binder, M.; Roekring, S.; Flegel, T. Spongiforma, a new genus of gasteroid boletes from Thailand. Fungal Divers. 2009, 37, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hosen, M.I.; Feng, B.; Wu, G.; Zhu, X.T.; Li, Y.C.; Yang, Z.L. Borofutus, a new genus of Boletaceae from tropical Asia: Phylogeny, morphology and taxonomy. Fungal Divers. 2013, 58, 215–226. [Google Scholar] [CrossRef]
- Orihara, T.; Smith, M.E.; Shimomura, N.; Iwase, K.; Maekawa, N. Diversity and systematics of the sequestrate genus Octaviania in Japan: Two new subgenera and eleven new species. Persoonia 2012, 28, 85–112. [Google Scholar] [CrossRef] [PubMed]
- Binder, M.; Hibbett, D. Molecular systematics and biological diversification of Boletales. Mycologia 2006, 98, 71–81. [Google Scholar] [CrossRef]
- Raspé, O.; Vadthanarat, S.; Kesel, A.D.; Degreef, J.; Hyde, K.D.; Lumyong, S. Pulveroboletus fragrans, a new Boletaceae species from Northern Thailand, with a remarkable aromatic odor. Mycol. Progr. 2016, 15, 38. [Google Scholar] [CrossRef]
- Orihara, T.; Healy, R.; Corrales, A.; Smith, M.E. Multi-locus phylogenies reveal three new truffle-like taxa and the traces of interspecific hybridization in Octaviania (Boletales). IMA Fungus 2021, 12, 1–22. [Google Scholar] [CrossRef]
- Orihara, T.; Lebel, T.; Ge, Z.W.; Smith, M.E.; Maekawa, N. Evolutionary history of the sequestrate genus Rossbeevera (Boletaceae) reveals a new genus Turmalinea and highlights the utility of ITS minisatellite-like insertions for molecular identification. Persoonia 2016, 37, 173–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Bakker, H.C.; Zuccarello, G.C.; Kuyper, T.W.; Noordeloos, M.E. Phylogeographic patterns in Leccinum sect. Scabra and the status of the arctic-alpine species L. rotundifoliae. Mycol. Res. 2007, 111, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Hosen, M.I.; Zhong, X.J.; Gates, G.; Orihara, T.; Li, T.H. Type studies of Rossbeevera bispora, and a new species of Rossbeevera from south China. MycoKeys 2019, 51, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Watling, R. British Fungus Flora—Agarics and Boleti. I. Boletaceae, Gomphidiaceae, Paxillaceae; Her Majesty’s Stationery Office: Edinburgh, UK, 1970. [Google Scholar]
- Rostkovius, F.W.T. Die Pilze Deutschlands. In Deutschlands Flora in Abbildungen nach der Natur mit Beschreibungen; Sturm, J., Ed.; Gedruckt auf Kosten des Verfassers: Nürnberg, Germany, 1832; Part III; Volume 5, p. 132. [Google Scholar] [CrossRef]
- Watling, R. British records. Trans. Br. mycol. Soc. 1960, 43, 691–694. [Google Scholar]
- Noordeloos, M.E.; Kuyper, T.W.; Somhorst, I.; Vellinga, E.C. Flora Agaricina Neerlandica; Lidia Carla Candusso: Varese, Italia, 2018; Volume 7, pp. 178–181. [Google Scholar]
- Horak, E. Revision of Malaysian species of Boletales s.l. (Basidiomycota) described by E.J.H. Corner (1972, 1974). Malay. For. Rec. 2011, 51, 1–283. [Google Scholar]
Figure 1.
Maximum-likelihood phylogenetic tree generated from a three-locus (nrLSU + tef1-α + rpb2) dataset. BS > 50% in ML analysis and PP > 0.95 in Bayesian analysis are indicated as RAxML BS/PP above or below supported branches. Species of this genus from China and type species of this genus (L. aurantiacum) are indicated in bold. Voucher specimens and localities where the specimens were collected are provided behind the species names. AU = Austria, BE = Belgium, CN = China, CR = Costa Rica, FI = Finland, FR = France, GER = Germany, IT = Italy, JP = Japan, TAI = Thailand, UK =United Kingdom, USA = United States of America and UZ = Uzbekistan.
Figure 1.
Maximum-likelihood phylogenetic tree generated from a three-locus (nrLSU + tef1-α + rpb2) dataset. BS > 50% in ML analysis and PP > 0.95 in Bayesian analysis are indicated as RAxML BS/PP above or below supported branches. Species of this genus from China and type species of this genus (L. aurantiacum) are indicated in bold. Voucher specimens and localities where the specimens were collected are provided behind the species names. AU = Austria, BE = Belgium, CN = China, CR = Costa Rica, FI = Finland, FR = France, GER = Germany, IT = Italy, JP = Japan, TAI = Thailand, UK =United Kingdom, USA = United States of America and UZ = Uzbekistan.

Figure 2.
Maximum-likelihood phylogenetic tree generated from ITS dataset. BS > 50% in ML analysis is indicated above or below supported branches. Species of this genus from China and type species of this genus (L. aurantiacum) are indicated in bold. Voucher specimens, localities and GenBank numbers are provided behind the species names. AT = Austria, CAN = Canada, CN = China, CR = Costa Rica, FR = France, GL = Greenland, JP = Japan, NL = The Netherlands, PL = Poland, SW = Sweden, UK =United Kingdom, USA = United States of America and UZ = Uzbekistan.
Figure 2.
Maximum-likelihood phylogenetic tree generated from ITS dataset. BS > 50% in ML analysis is indicated above or below supported branches. Species of this genus from China and type species of this genus (L. aurantiacum) are indicated in bold. Voucher specimens, localities and GenBank numbers are provided behind the species names. AT = Austria, CAN = Canada, CN = China, CR = Costa Rica, FR = France, GL = Greenland, JP = Japan, NL = The Netherlands, PL = Poland, SW = Sweden, UK =United Kingdom, USA = United States of America and UZ = Uzbekistan.

Figure 3.
Basidiomata of Leccinum species. (a–c) Leccinum parascabrum (KUN-HKAS99903, holotype); (d–f) Leccinum pseudoborneense ((d) from KUN-HKAS110157; (e,f) from KUN-HKAS110156, holotype); (g,h) Leccinum album (KUN-HKAS53417, holotype).
Figure 3.
Basidiomata of Leccinum species. (a–c) Leccinum parascabrum (KUN-HKAS99903, holotype); (d–f) Leccinum pseudoborneense ((d) from KUN-HKAS110157; (e,f) from KUN-HKAS110156, holotype); (g,h) Leccinum album (KUN-HKAS53417, holotype).

Figure 4.
Microscopic features of Leccinum album (KUN-HKAS53417, holotype). (a) Basidiospores. (b) Basidia and pleurocystidium. (c) Cheilocystidia and pleurocystidia. (d) Pileipellis. Bars = 10 µm. Drawings by Y.-C. Li.
Figure 4.
Microscopic features of Leccinum album (KUN-HKAS53417, holotype). (a) Basidiospores. (b) Basidia and pleurocystidium. (c) Cheilocystidia and pleurocystidia. (d) Pileipellis. Bars = 10 µm. Drawings by Y.-C. Li.

Figure 5.
Microscopic features of Leccinum parascabrum (KUN-HKAS99903, holotype). (a) Basidiospores. (b) Basidia and pleurocystidium. (c) Cheilocystidia. (d) Pleurocystidia. (e) Pileipellis. Bars = 10 µm. Drawings by Y.-C. Li.
Figure 5.
Microscopic features of Leccinum parascabrum (KUN-HKAS99903, holotype). (a) Basidiospores. (b) Basidia and pleurocystidium. (c) Cheilocystidia. (d) Pleurocystidia. (e) Pileipellis. Bars = 10 µm. Drawings by Y.-C. Li.

Figure 6.
Microscopic features of Leccinum pseudoborneense (KUN-HKAS110156, holotype). (a) Basidiospores. (b) Basidia and pleurocystidia. (c) Pleurocystidia. (d) Cheilocystidia. (e) Pileipellis. Bars = 10 µm. Drawings by Y.-C. Li.
Figure 6.
Microscopic features of Leccinum pseudoborneense (KUN-HKAS110156, holotype). (a) Basidiospores. (b) Basidia and pleurocystidia. (c) Pleurocystidia. (d) Cheilocystidia. (e) Pileipellis. Bars = 10 µm. Drawings by Y.-C. Li.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Information on specimens used in multi-locus phylogenetic analyses and their GenBank accession numbers. Sequences newly generated in this study are indicated in bold.
Table 1.
Information on specimens used in multi-locus phylogenetic analyses and their GenBank accession numbers. Sequences newly generated in this study are indicated in bold.
| Species | Voucher | Locality | GenBank Number | Reference | |||
|---|---|---|---|---|---|---|---|
| ITS | nrLSU | tef1-α | rpb2 | ||||
| Leccinellum corsicum | Buf 4507 | USA | - | KF030347 | KF030435 | - | [55] |
| Le. crocipodium | MICH:KUO-07050707 | USA | - | MK601749 | MK721103 | MK766311 | [5] |
| Le. aff. griseum | KPM-NC-0017381 | Japan | - | JN378508 | JN378449 | - | [73] |
| Le. lepidum | K(M)-142974 | Italy | - | MK601751 | MK721105 | MK766312 | [5] |
| Le. pseudoscabrum | CFMR:DPL-11432 | USA | - | MK601752 | MK721106 | MK766313 | [5] |
| Le. rugosiceps | CFMR:BOS-866 | USA | - | MK601770 | MK721124 | MK766329 | [5] |
| Leccinum album | KUN-HKAS53417 | China | MZ392872 MZ392873 | MW413907 HQ326880 | MW439267 HQ326861 | MW439259 MW439260 | This study |
| L. aurantiacum | L-0342207 | France | - | MK601759 | MK721113 | MK766318 | [5] |
| L. cerinum | MK11800 | Finland | - | AF139692 | - | - | [4] |
| L. duriusculum | KUN-HKAS101160 | Uzbekistan | MZ485402 | MZ675541 | MZ707785 | MZ707779 | This study |
| L. duriusculum | GL4676 | France | - | AF139699 | - | - | [4] |
| L. flavostipitatum | MENMB10801 | USA | - | MH620342 | - | - | GenBank |
| L. holopus | MICH:KUO09150707 | USA | - | MK601763 | MK721117 | MK766322 | [5] |
| L. holopus | 9109303 | France | - | AF139700 | [4] | ||
| L. holopus | KUN-HKAS111906 | Austria | - | MW413906 | MW439266 | MW439258 | This study |
| L. manzanitae | NY-14041 | USA | - | MK601765 | MK721119 | MK766324 | [5] |
| L. melaneum | KUN-HKAS57220 | China | MZ485409 | MZ675542 | MZ707786 | MZ707780 | This study |
| L. monticola | NY-00815448 | Costa Rica | - | MK601767 | MK721121 | MK766326 | [5] |
| L. monticola | NY-760388 | Costa Rica | - | MK601766 | MK721120 | MK766325 | [5] |
| L. palustre | MK11107 | Germany | - | AF139701 | - | - | [4] |
| “L. palustre” | hdb030 | Netherlands | AF454586 | - | - | - | [57] |
| L. parascabrum | KUN-HKAS99903 | China | MZ392874 | MW413911 | MW439271 | MW439264 | This study |
| L. parascabrum | KUN-HKAS59447 | China | MZ392875 | MW413912 | MW439272 | MW439265 | This study |
| L. pseudoborneense | KUN-HKAS110156 | China | MZ412902 | MW413908 | MW439268 | MW439261 | This study |
| L. pseudoborneense | KUN-HKAS110157 | China | MZ412903 | MW413909 | MW439269 | MW439262 | This study |
| L. pseudoborneense | KUN-HKAS110158 | China | MZ412904 | MW413910 | MW439270 | MW439263 | This study |
| L. pseudoborneense | KUN-HKAS89139 | China | - | MZ536631 | MZ543306 | MZ543308 | This study |
| L. pseudoborneense | KUN-HKAS92401 | China | - | MZ536632 | MZ543307 | MZ543309 | This study |
| L. quercinum | KUN-HKAS63502 | China | - | KF112724 | KF112250 | KF112724 | [65] |
| L. quercinum | Lq1 | Germany | - | DQ534612 | - | - | [74] |
| “L. scabrum” | KUN-HKAS56371 | China | - | KT990587 | KT990782 | KT990423 | [11] |
| L. scabrum | KUN-HKAS57266 | China | - | KF112442 | KF112248 | KF112722 | [65] |
| L. scabrum | KUN-HKAS98029 | China | MZ485407 | MZ675543 | MZ707787 | - | This study |
| L. scabrum | KPM-NC-0017840 | UK | - | JN378515 | JN378455 | - | [73] |
| L. schistophilum | KUN-HKAS98024 | China | MZ503508 | MZ675544 | MZ707788 | - | This study |
| L. schistophilum | VDKO1128 | Belgium | - | - | KT824055 | KT824022 | [75] |
| L. subradicatum | KPM-NC-24518 | Japan | MT934814 | MT812736 | MT874822 | - | [76] |
| L. sp. | KPM-NC-0017830 | Japan | KC552009 | [77] | |||
| L. variicolor | Lvar1 | Germany | - | AF139706 | - | - | [4] |
| L. variicolor | Hdb327 | Canada | AY538843 | - | - | - | [33] |
| L. variicolor | Watling6753 | UK | AY853538 | - | - | - | [78] |
| L. versipelle | KUN-HKAS76669 | China | - | KF112443 | KF112249 | KF112723 | [65] |
| L. versipelle | CFMR DLC2002-122 | USA | - | MK601778 | - | - | [5] |
| L. versipelle | - | Sweden | AF454573 | - | - | - | [57] |
| L. versipelle | KUN-HKAS97997 | China | MZ485404 | MZ675545 | MZ707789 | MZ707781 | This study |
| L. versipelle | KUN-HKAS99380 | China | MZ485401 | MZ675546 | MZ707790 | MZ707782 | This study |
| L. violaceotinctum | CFMR:BZ-1676 | Belize | - | MK601780 | MK721133 | MK766337 | [5] |
| L. violaceotinctum | CFMR:BZ-3169 | Belize | - | - | MK721134 | MK766338 | [5] |
| Borofutus dhakanus | KUN-HKAS73789 | Bengal | - | JQ928616 | JQ928576 | JQ928597 | [72] |
| Chamonixia brevicolumna | DBG:F023359 | USA | - | MK601728 | MK721082 | MK766290 | [5] |
| Octaviania japonimontana | KPM-NC-0017812 | Japan | - | JN378486 | JN378428 | - | [73] |
| O. tasmanica | NY-02449788 | USA | - | MK601798 | MK721152 | MK766355 | [5] |
| Rossbeevera griseobrunnea | GDGM45913 | China | - | MH537793 | - | - | [79] |
| R. eucyanea | KPM-NC-0023895 | Japan | - | KP222896 | KP222915 | - | [76] |
| Rugiboletus andinus | NY-00796145 | USA | - | MK601758 | MK721112 | MK766317 | [5] |
| Ru. andinus | NY-181460 | USA | - | MK601757 | MK721111 | MK766316 | [5] |
| Spongiforma thailandica | DED7873 | Thailand | - | NG_042464 | KF030436 | MG212648 | [71] |
| Turmalinea mesomorpha | KPM-NC-0017743 | Japan | - | KC552050 | - | - | [77] |
| T. yuwanensis | KPM-NC0023377 | Japan | - | KJ001098 | KJ001083 | - | [77] |
| Tylocinum griseolum | KUN-HKAS52612 | China | - | KT990631 | KT990825 | - | [11] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Meng, X.; Wang, G.-S.; Wu, G.; Wang, P.-M.; Yang, Z.L.; Li, Y.-C. The Genus Leccinum (Boletaceae, Boletales) from China Based on Morphological and Molecular Data. J. Fungi 2021, 7, 732. https://doi.org/10.3390/jof7090732
AMA Style
Meng X, Wang G-S, Wu G, Wang P-M, Yang ZL, Li Y-C. The Genus Leccinum (Boletaceae, Boletales) from China Based on Morphological and Molecular Data. Journal of Fungi. 2021; 7(9):732. https://doi.org/10.3390/jof7090732
Chicago/Turabian StyleMeng, Xin, Geng-Shen Wang, Gang Wu, Pan-Meng Wang, Zhu L. Yang, and Yan-Chun Li. 2021. "The Genus Leccinum (Boletaceae, Boletales) from China Based on Morphological and Molecular Data" Journal of Fungi 7, no. 9: 732. https://doi.org/10.3390/jof7090732
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.