
The Negative Effect of Protein Phosphatase Z1 Deletion on the Oxidative Stress Tolerance of Candida albicans Is Synergistic with Betamethasone Exposure

 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Fungal Strains
2.2. Stress Treatments
2.3. RNA Sequencing (RNASeq)
2.4. Reverse Transcription Quantitative Real-Time PCR (RT-qPCR) Assay
2.5. Gene Set Enrichment Analysis
- (i)
- antioxidant enzyme genes—this gene group includes genes encoding functionally verified and/or putative antioxidant enzymes including the catalase (GOID: 4096), superoxide dismutase (GOID: 4784), glutaredoxin (GOID: 6749), thioredoxin (GOIDs: 8379 and 51920) and peroxidase (GOID: 4601) GO terms;
- (ii)
- iron metabolism-related genes—genes involved in iron acquisition by C. albicans were collected according to Fourie et al. [22];
- (iii)
- zinc and copper homeostasis genes—genes involved in zinc and copper acquisition by C. albicans were collected according to Gerwien et al. [23];
- (iv)
- metabolic pathway-related genes—this group contains all genes related to the TCA, ethanol fermentation, glycogen metabolism, and ergosterol biosynthesis biochemical pathways according to the pathway database (http://pathway.candidagenome.org, accessed on 5 July 2021).
2.6. Determination of Virulence Attributes
Biofilm Development and Metabolic Activity-Based Susceptibility Testing
2.7. Assays of Redox Homeostasis and Antioxidant Enzyme Activities
2.8. Assay of Glucose Consumption, Ethanol Production, and Iron, Zinc and Copper Contents of C. albicans Cells
2.9. Statistical Analysis
3. Results
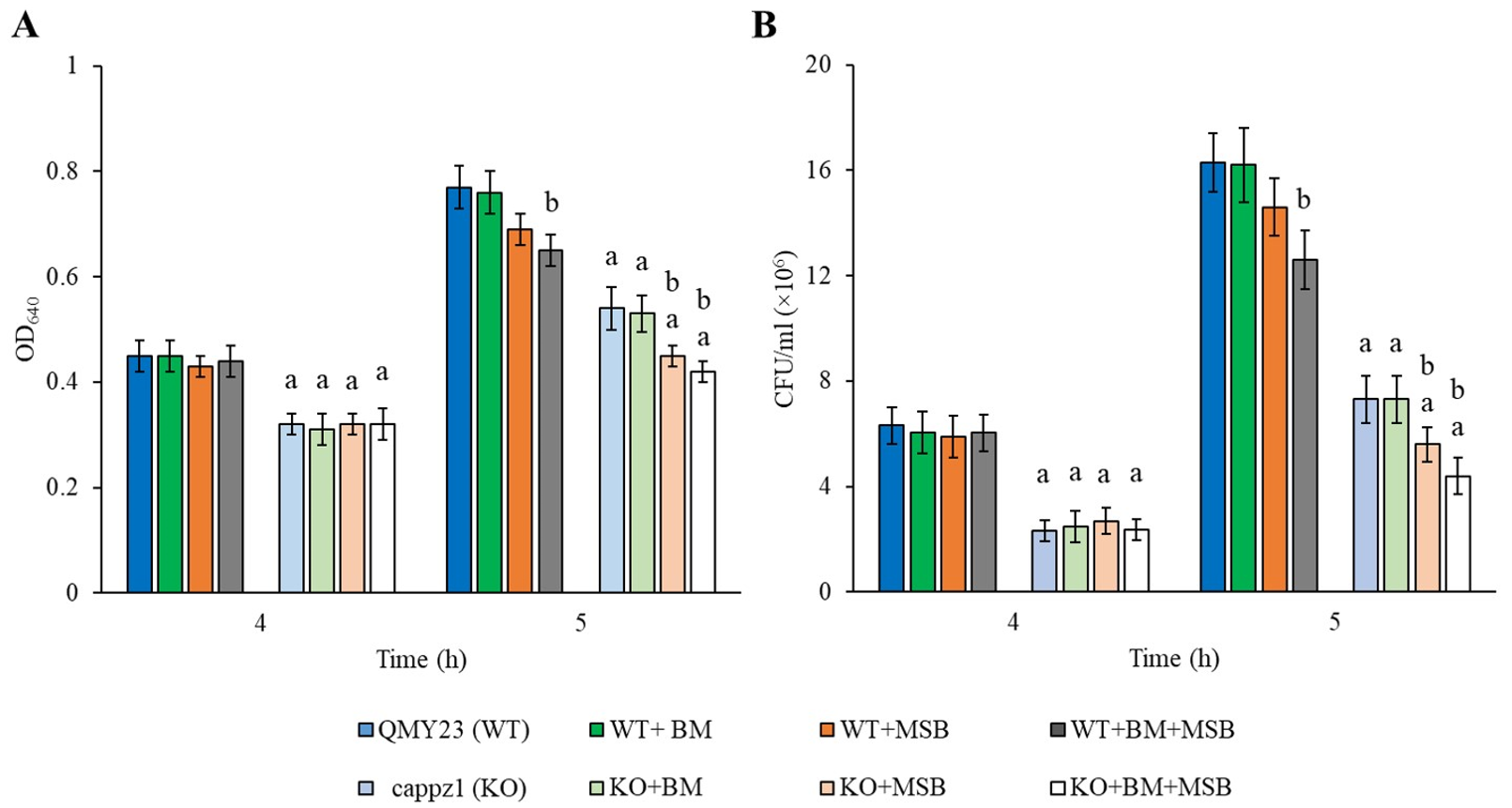
3.1. BM and MSB Suppressed Growth and Metabolic Activity of Biofilm Synergistically
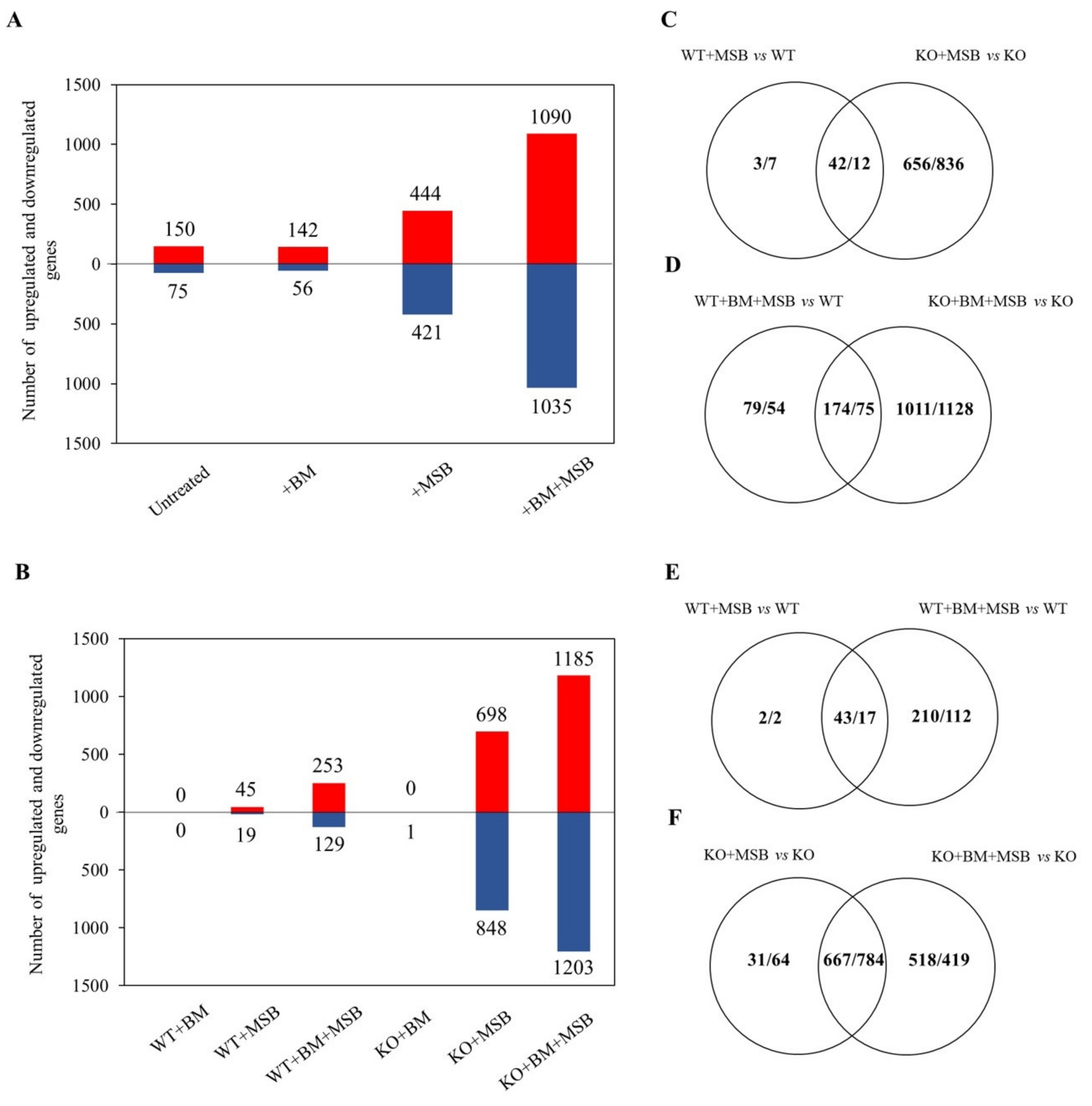
3.2. Genome-Wide Transcriptional Changes Confirmed the Interaction between BM and MSB Treatments
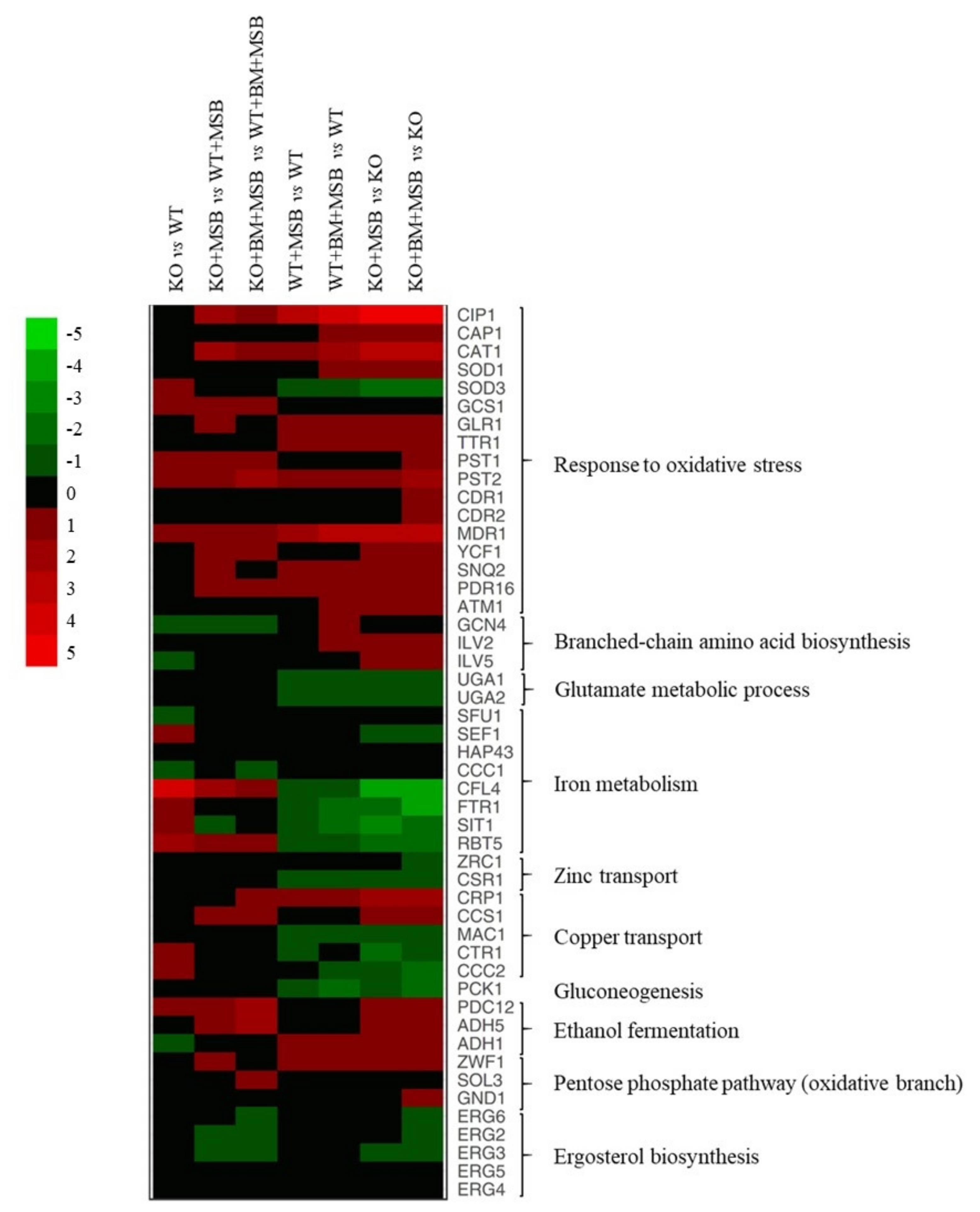
3.3. Gene Ontology Analysis of the Transcriptome Changes
3.3.1. The CaPPZ1 Gene Deletion Has a Moderate Transcriptomic Effect
3.3.2. The Cellular Response to the Oxidative Stress Was More Extensive in the Cappz1 Knock Out Strain
3.3.3. Combination of CaPPZ1 Gene Deletion and MSB Stress Led to a Global Reprogramming of Gene Expression in BM Pre-Treated Cells
3.4. Testing of the Predicted Consequences of Transcriptome Changes
3.4.1. BM Did Not Influence the Growth, Extracellular Phospholipase and Proteinase Production and Hypha Formation of C. albicans
3.4.2. BM Pre-Treatment Enhanced the Oxidative Stress and the Stress Response Induced by MSB
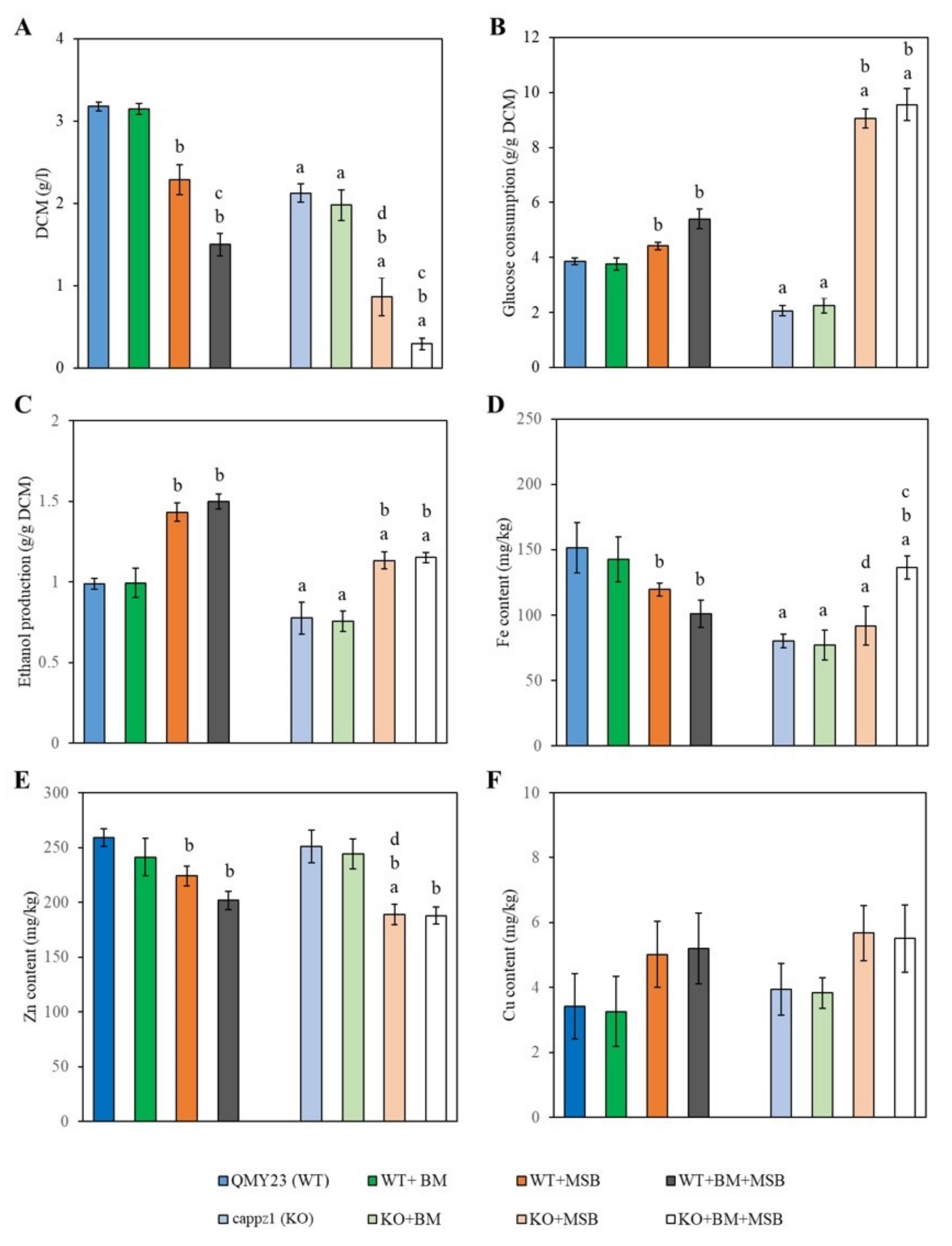
3.4.3. Deletion of CaPPZ1 and MSB-Induced Oxidative Stress Reduced Glucose Consumption, Ethanol Production and Influenced Metal Ion Content of C. albicans
4. Discussion
4.1. Phenotypes and Transcriptional Changes Attributed to CaPPZ1 Gene Deletion
4.2. Transcriptional Changes Related to MSB Exposures
4.3. Combined Effects of CaPPZ1 Deletion and MSB Treatment
4.4. Synergistic Effects of BM Exposure and MSB Treatment on the Phosphatase Deletion Mutant
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lionakis, M.S.; Kontoyiannis, D.P. Glucocorticoids and invasive fungal infections. Lancet 2003, 362, 1828–1838. [Google Scholar] [CrossRef]
- Yasir, M.; Goyal, A.; Bansal, P.; Sonthalia, S. Corticosteroid Adverse Effects; StatPearls [Internet]: Treasure Island, FL, USA, 2021. [Google Scholar]
- Pfaller, M.A.; Andes, D.R.; Diekema, D.J.; Horn, D.L.; Reboli, A.C.; Rotstein, C.; Franks, B.; Azie, N.E. Epidemiology and outcomes of invasive candidiasis due to non-albicans species of Candida in 2,496 patients: Data from the Prospective Antifungal Therapy (PATH) registry 2004–2008. PLoS ONE 2014, 9, e101510. [Google Scholar] [CrossRef] [Green Version]
- Gani, F.; Caminati, M.; Bellavia, F.; Baroso, A.; Faccioni, P.; Pancera, P.; Batani, V.; Senna, G. Oral health in asthmatic patients: A review: Asthma and its therapy may impact on oral health. Clin. Mol. Allergy 2020, 18, 22. [Google Scholar] [CrossRef]
- Neofytos, D.; Fishman, J.A.; Horn, D.; Anaissie, E.; Chang, C.H.; Olyaei, A.; Pfaller, M.; Steinbach, W.J.; Webster, K.M.; Marr, K.A. Epidemiology and outcome of invasive fungal infections in solid organ transplant recipients. Transpl. Infect Dis. 2010, 12, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Danza, A.; Ruiz-Irastorza, G. Infection risk in systemic lupus erythematosus patients: Susceptibility factors and preventive strategies. Lupus 2013, 22, 1286–1294. [Google Scholar] [CrossRef]
- Gyetvai, A.; Emri, T.; Fekete, A.; Varga, Z.; Gazdag, Z.; Pesti, M.; Belágyi, J.; Emõdy, L.; Pócsi, I.; Lenkey, B. High-dose methylprednisolone influences the physiology and virulence of Candida albicans ambiguously and enhances the candidacidal activity of the polyene antibiotic amphotericin B and the superoxide-generating agent menadione. FEMS Yeast Res. 2007, 7, 265–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakab, Á.; Emri, T.; Sipos, L.; Kiss, Á.; Kovács, R.; Dombrádi, V.; Kemény-Beke, Á.; Balla, J.; Majoros, L.; Pócsi, I. Betamethasone augments the antifungal effect of menadione—Towards a novel anti-Candida albicans combination therapy. J. Basic Microbiol. 2015, 55, 973–981. [Google Scholar] [CrossRef]
- Jakab, Á.; Mogavero, S.; Förster, T.M.; Pekmezovic, M.; Jablonowski, N.; Dombrádi, V.; Pócsi, I.; Hube, B. Effects of the glucocorticoid betamethasone on the interaction of Candida albicans with human epithelial cells. Microbiology 2016, 162, 2116–2125. [Google Scholar] [CrossRef]
- Kobayashi-Sakamoto, M.; Tamai, R.; Isogai, E.; Kiyoura, Y. Gastrointestinal colonisation and systemic spread of Candida albicans in mice treated with antibiotics and prednisolone. Microb. Pathog. 2018, 117, 191–199. [Google Scholar] [CrossRef]
- Rigopoulos, D.; Ioannides, D.; Kalogeromitros, D.; Gregoriou, S.; Katsambas, A. Pimecrolimus cream 1% vs. betamethasone 17-valerate 0.1% cream in the treatment of seborrhoeic dermatitis. A randomized open-label clinical trial. Br. J. Dermatol. 2004, 15, 1071–1075. [Google Scholar] [CrossRef]
- Sticherling, M.; Eicke, C.; Anger, T. 2013. Practicability of combined treatment with calcipotriol/betamethasone gel (Daivobet1 Gel) and improvement of quality of life in patients with psoriasis. J. Dtsch. Dermatol. Ges. 2013, 11, 420–427. [Google Scholar] [PubMed]
- Szabó, K.; Miskei, M.; Farkas, I.; Dombrádi, V. The phosphatome of opportunistic pathogen Candida species. Fungal Biol. Rev. 2021, 35, 40–51. [Google Scholar] [CrossRef]
- Ádám, C.; Erdei, É.; Casado, C.; Kovács, L.; González, A.; Majoros, L.; Petrényi, K.; Bagossi, P.; Farkas, I.; Molnar, M.; et al. Protein phosphatase CaPpz1 is involved in cation homeostasis, cell wall integrity and virulence of Candida albicans. Microbiology 2012, 158, 1258–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiter, É.; González, A.; Erdei, É.; Casado, C.; Kovács, L.; Ádám, C.; Oláh, J.; Miskei, M.; Molnar, M.; Farkas, I.; et al. Protein phosphatase Z modulates oxidative stress response in fungi. Fungal Genet. Biol. 2012, 49, 708–716. [Google Scholar] [CrossRef]
- Muszkieta, L.; de Jesus Carrion, S.; Robinet, P.; Beau, R.; Elbim, C.; Pearlman, E.; Latgé, J.P. The protein phosphatase PhzA of A. fumigatus is involved in oxidative stress tolerance and fungal virulence. Fungal Genet. Biol. 2014, 66, 79–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, K.; Jakab, Á.; Póliska, S.; Petrényi, K.; Kovács, K.; Issa, L.H.B.; Emri, T.; Pócsi, I.; Dombrádi, V. Deletion of the fungus specific protein phosphatase Z1 exaggerates the oxidative stress response in Candida albicans. BMC Genom. 2019, 20, 873. [Google Scholar] [CrossRef]
- Mitrovich, Q.M.; Tuch, B.B.; Guthrie, C.; Johnson, A.D. Computational and experimental approaches double the number of known introns in the pathogenic yeast Candida albicans. Genome Res. 2007, 17, 492–502. [Google Scholar] [CrossRef] [Green Version]
- Jakab, Á.; Tóth, Z.; Nagy, F.; Nemes, D.; Bácskay, I.; Kardos, G.; Emri, T.; Pócsi, I.; Majoros, L.; Kovács, R. Physiological and transcriptional responses of Candida parapsilosis to exogenous tyrosol. Appl. Environ. Microbiol. 2019, 85, e01388-19. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus, NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Fourie, R.; Kuloyo, O.O.; Mochochoko, B.M.; Albertyn, J.; Pohl, C.H. Iron at the Centre of Candida albicans Interactions. Front. Cell Infect. Microbiol. 2018, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Gerwien, F.; Skrahina, V.; Kasper, L.; Hube, B.; Brunke, S. Metals in fungal virulence. FEMS Microbiol. Rev. 2018, 42, fux050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, F.; Tóth, Z.; Daróczi, L.; Székely, A.; Borman, A.M.; Majoros, L.; Kovács, R. Farnesol increases the activity of echinocandins against Candida auris biofilms. Med. Mycol. 2020, 58, 404–407. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard—3rd ed.; Document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008; Volume 28, pp. 1–25. [Google Scholar]
- Meletiadis, J.; Verweij, P.E.; TeDorsthorst, D.T.; Meis, J.F.; Mouton, J.W. Assessing in vitro combinations of antifungal drugs against yeasts and filamentous fungi, comparison of different drug interaction models. Med. Mycol. 2005, 43, 133–152. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Boczonádi, I.; Török, Z.; Jakab, Á.; Kónya, G.; Gyurcsó, K.; Baranyai, E.; Szoboszlai, Z.; Döncző, B.; Fábián, I.; Leiter, É.; et al. Increased Cd2+ biosorption capability of Aspergillus nidulans elicited by crpA deletion. J. Basic. Microbiol. 2020, 60, 574–584. [Google Scholar] [CrossRef]
- Leary, N.O.; Pembroke, A.; Duggan, P.F. Improving accuracy of glucose oxidase procedure for glucose determinations on discrete analyzers. Clin. Chem. 1992, 38, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Boczonádi, I.; Jakab, Á.; Baranyai, E.; Tóth, C.N.; Daróczi, L.; Csernoch, L.; Kis, G.; Antal, M.; Pusztahelyi, T.; Grawunder, A.; et al. Rare earth element sequestration by Aspergillus oryzae biomass. Environ. Technol. 2020, 16, 1–11. [Google Scholar] [CrossRef]
- Tiscione, N.B.; Alford, I.; Yeatman, D.T.; Shan, X. Ethanol analysis by headspace gas chromatography with simultaneous flame-ionization and mass spectrometry detection. J. Anal. Toxicol. 2011, 35, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Diekmann, J.A., 3rd; Cochran, J.; Hodgson, J.A.; Smuts, D.J. Quantitation and identification of ethanol and inhalant compounds in whole blood using static headspace gas chromatography vacuum ultraviolet spectroscopy. J. Chromatogr. A 2020, 1611, 460607. [Google Scholar] [CrossRef] [PubMed]
- Pócsi, I.; Prade, R.A.; Penninckx, M.J. Glutathione, altruistic metabolite in fungi. Adv Microb. Physiol. 2004, 49, 1–76. [Google Scholar]
- Li, L.; Naseem, S.; Sharma, S.; Konopka, J.B. Flavodoxin-Like Proteins Protect Candida albicans from Oxidative Stress and Promote Virulence. PLoS Pathog. 2015, 11, e1005147. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, D.; Albacar, M.; Zhang, C.; Calafí, C.; López-Malo, M.; Torres-Torronteras, J.; Martí, R.; Kovalchuk, S.I.; Pinson, B.; Jensen, O.N.; et al. Yeast Ppz1 protein phosphatase toxicity involves the alteration of multiple cellular targets. Sci. Rep. 2020, 10, 15613. [Google Scholar] [CrossRef] [PubMed]
- Mattiasson, G. Analysis of mitochondrial generation and release of reactive oxygen species. Cytom. Part A 2004, 62, 89–96. [Google Scholar] [CrossRef]
- Pereira, C.; Silva, R.D.; Saraiva, L.; Johansson, B.; Sousa, M.J.; Côrte-Real, M. Mitochondria-dependent apoptosis in yeast. Biochim. Biophys. Acta 2008, 1783, 1286–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, B.; Cheng, S.; Clancy, C.J.; Nguyen, M.H. Caspofungin kills Candida albicans by causing both cellular apoptosis and necrosis. Antimicrob. Agents Chemother. 2013, 57, 326–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiter, É.; Park, H.S.; Kwon, N.J.; Han, K.H.; Emri, T.; Oláh, V.; Mészáros, I.; Dienes, B.; Vincze, J.; Csernoch, L.; et al. Characterization of the aodA, dnmA, mnSOD and pimA genes in Aspergillus nidulans. Sci. Rep. 2016, 6, 20523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saibabu, V.; Singh, S.; Ansari, M.A.; Fatima, Z.; Hameed, S. Insights into the intracellular mechanisms of citronellal in Candida albicans: Implications for reactive oxygen species-mediated necrosis, mitochondrial dysfunction, and DNA damage. Rev. Soc. Bras. Med. Trop. 2017, 50, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Winkelströter, L.K.; Dolan, S.K.; Fernanda Dos Reis, T.; Bom, V.L.; Alves de Castro, P.; Hagiwara, D.; Alowni, R.; Jones, G.W.; Doyle, S.; Brown, N.A.; et al. Systematic Global Analysis of Genes Encoding Protein Phosphatases in Aspergillus fumigatus. G3 2015, 5, 1525–1539. [Google Scholar] [CrossRef] [Green Version]
- Manfiolli, A.O.; de Castro, P.A.; Dos Reis, T.F.; Dolan, S.; Doyle, S.; Jones, G.; Riaño Pachón, D.M.; Ulaş, M.; Noble, L.M.; Mattern, D.J.; et al. Aspergillus fumigatus protein phosphatase PpzA is involved in iron assimilation, secondary metabolite production, and virulence. Cell Microbiol. 2017, 19, e12770. [Google Scholar] [CrossRef] [Green Version]
- Skrahina, V.; Brock, M.; Hube, B.; Brunke, S. Candida albicans Hap43 Domains Are Required under Iron Starvation but Not Excess. Front. Microbiol. 2017, 8, 2388. [Google Scholar] [CrossRef]
- Ariño, J.; Velázquez, D.; Casamayor, A. Ser/Thr protein phosphatases in fungi: Structure, regulation and function. Microb. Cell 2019, 6, 217–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, G.; Hennig, G.; Petrényi, K.; Kovács, L.; Pócsi, I.; Dombrádi, V.; Bánfalvi, G. Time-lapse video microscopy and image analysis of adherence and growth patterns of Candida albicans strains. Appl. Microbiol. Biotechnol. 2014, 98, 5185–5194. [Google Scholar] [CrossRef]
- Chen, D.; Wilkinson, C.R.; Watt, S.; Penkett, C.J.; Toone, W.M.; Jones, N.; Bähler, J. Multiple pathways differentially regulate global oxidative stress responses in fission yeast. Mol. Biol. Cell. 2008, 19, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.L.; Tseng, S.F.; Huang, Y.C.; Shen, Z.J.; Hsu, P.H.; Hsieh, M.H.; Yang, C.W.; Tognetti, S.; Canal, B.; Subirana, L.; et al. Yeast Cip1 is activated by environmental stress to inhibit Cdk1–G1 cyclins via Mcm1 and Msn2/4. Nat. Commun. 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Kusch, H.; Biswas, K.; Schwanfelder, S.; Engelmann, S.; Rogers, P.D.; Hecker, M.; Morschhäuser, J. A proteomic approach to understanding the development of multidrug-resistant Candida albicans strains. Mol. Genet. Genom. 2004, 271, 554–565. [Google Scholar] [CrossRef]
- Pócsi, I.; Miskei, M.; Karányi, Z.; Emri, T.; Ayoubi, P.; Pusztahelyi, T.; Balla, G.; Prade, R.A. Comparison of gene expression signatures of diamide, H2O2 and menadione exposed Aspergillus nidulans cultures-linking genome-wide transcriptional changes to cellular physiology. BMC Genom. 2005, 6, 182. [Google Scholar] [CrossRef] [Green Version]
- Orosz, E.; Antal, K.; Gazdag, Z.; Szabó, Z.; Han, K.H.; Yu, J.H.; Pócsi, I.; Emri, T. Transcriptome-Based Modeling Reveals that Oxidative Stress Induces Modulation of the AtfA-Dependent Signaling Networks in Aspergillus nidulans. Int. J. Genom. 2017, 2017, 6923849. [Google Scholar]
- Dantas Ada, S.; Day, A.; Ikeh, M.; Kos, I.; Achan, B.; Quinn, J. Oxidative stress responses in the human fungal pathogen, Candida albicans. Biomolecules 2015, 5, 142–165. [Google Scholar] [CrossRef] [Green Version]
- Dixon, S.J.; Stockwell, B.R. The role of iron and reactive oxygen species in cell death. Nat. Chem. Biol. 2014, 10, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kurucz, V.; Krüger, T.; Antal, K.; Dietl, A.M.; Haas, H.; Pócsi, I.; Kniemeyer, O.; Emri, T. Additional oxidative stress reroutes the global response of Aspergillus fumigatus to iron depletion. BMC Genom. 2018, 19, 357. [Google Scholar] [CrossRef]
- Gasch, A.P.; Werner-Washburne, M. The genomics of yeast responses to environmental stress and starvation. Funct. Integr. Genom. 2002, 2, 181–192. [Google Scholar] [CrossRef]
- Buechel, E.R.; Pinkett, H.W. Transcription factors and ABC transporters: From pleiotropic drug resistance to cellular signaling in yeast. FEBS Lett. 2020, 594, 3943–3964. [Google Scholar] [CrossRef]
- Sethiya, P.; Rai, M.N.; Rai, R.; Parsania, C.; Tan, K.; Wong, K.H. Transcriptomic analysis reveals global and temporal transcription changes during Candida glabrata adaptation to an oxidative environment. Fungal Biol. 2020, 124, 427–439. [Google Scholar] [CrossRef]
- Saidane, S.; Weber, S.; De Deken, X.; St-Germain, G.; Raymond, M. PDR16-mediated azole resistance in Candida albicans. Mol. Microbiol. 2006, 60, 1546–1562. [Google Scholar] [CrossRef]
- Askew, C.; Sellam, A.; Epp, E.; Hogues, H.; Mullick, A.; Nantel, A.; Whiteway, M. Transcriptional regulation of carbohydrate metabolism in the human pathogen Candida albicans. PLoS Pathog. 2009, 5, e1000612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, W.A.; Roach, P.J.; Montero, M.; Baroja-Fernández, E.; Muñoz, F.J.; Eydallin, G.; Viale, A.M.; Pozueta-Romero, J. Regulation of glycogen metabolism in yeast and bacteria. FEMS Microbiol. Rev. 2010, 34, 952–985. [Google Scholar] [CrossRef] [Green Version]
- Riggle, P.J.; Kumamoto, C.A. Role of a Candida albicans P1-type ATPase in resistance to copper and silver ion toxicity. J. Bacteriol. 2000, 182, 4899–4905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissman, Z.; Berdicevsky, I.; Cavari, B.Z.; Kornitzer, D. The high copper tolerance of Candida albicans is mediated by a P-type ATPase. Proc. Natl. Acad. Sci. USA 2000, 97, 3520–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marvin, M.E.; Williams, P.H.; Cashmore, A.M. The Candida albicans CTR1 gene encodes a functional copper transporter. Microbiology 2003, 149, 1461–1474. [Google Scholar] [CrossRef] [Green Version]
- Marvin, M.E.; Mason, R.P.; Cashmore, A.M. The CaCTR1 gene is required for high-affinity iron uptake and is transcriptionally controlled by a copper-sensing transactivator encoded by CaMAC1. Microbiology 2004, 150, 2197–2208. [Google Scholar] [CrossRef] [PubMed]
- Gleason, J.E.; Li, C.X.; Odeh, H.M.; Culotta, V.C. Species-specific activation of Cu/Zn SOD by its CCS copper chaperone in the pathogenic yeast Candida albicans. J. Biol. Inorg. Chem. 2014, 19, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.X.; Gleason, J.E.; Zhang, S.X.; Bruno, V.M.; Cormack, B.P.; Culotta, V.C. Candida albicans adapts to host copper during infection by swapping metal cofactors for superoxide dismutase. Proc. Natl. Acad. Sci. USA 2015, 112, E5336–E5342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, C.; Festa, R.A.; Chen, Y.L.; Espart, A.; Palacios, Ò.; Espín, J.; Capdevila, M.; Atrian, S.; Heitman, J.; Thiele, D.J. Cryptococcus neoformans copper detoxification machinery is critical for fungal virulence. Cell Host Microbe 2013, 13, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackie, J.; Szabo, E.K.; Urgast, D.S.; Ballou, E.R.; Childers, D.S.; MacCallum, D.M.; Feldmann, J.; Brown, A.J. Host-Imposed Copper Poisoning Impacts Fungal Micronutrient Acquisition during Systemic Candida albicans Infections. PLoS ONE 2016, 11, e0158683. [Google Scholar] [CrossRef] [Green Version]
- Wiemann, P.; Perevitsky, A.; Lim, F.Y.; Shadkchan, Y.; Knox, B.P.; Landero Figueora, J.A.; Choera, T.; Niu, M.; Steinberger, A.J.; Wüthrich, M.; et al. Aspergillus fumigatus Copper Export Machinery and Reactive Oxygen Intermediate Defense Counter Host Copper-Mediated Oxidative Antimicrobial Offense. Cell Rep. 2017, 19, 1008–1021. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, P.; Bidaud, A.L.; Dannaoui, E. In vitro synergy of isavuconazole in combination with colistin against Candida auris. Sci. Rep. 2020, 10, 21448. [Google Scholar] [CrossRef] [PubMed]
- Badave, K.D.; Khan, A.A.; Rane, S.Y. Anticancer Vitamin K3 Analogs: A Review. Anticancer Agents Med. Chem. 2016, 16, 1017–1030. [Google Scholar] [CrossRef]
- Shertzer, H.G.; Låstbom, L.; Sainsbury, M.; Moldéus, P. Menadione-mediated membrane fluidity alterations and oxidative damage in rat hepatocytes. Biochem. Pharmacol. 1992, 43, 2135–2141. [Google Scholar] [CrossRef]
- Xu, C.J.; Zhang, Y.; Wang, J.; Zhang, T.M. Menadione reduced doxorubicin resistance in Ehrlich ascites carcinoma cells in vitro. Zhongguo Yao Li Xue Bao. 1998, 19, 273–276. [Google Scholar]
- Cho, Y.S.; Kim, M.J.; Lee, J.Y.; Chung, J.H. The role of thiols in protecting against simultaneous toxicity of menadione to platelet plasma and intracellular membranes. J. Pharmacol. Exp. Ther. 1997, 280, 1335–1340. [Google Scholar] [PubMed]
- Sitcheran, R.; Emter, R.; Kralli, A.; Yamamoto, K.R. A genetic analysis of glucocorticoid receptor signaling. Identification and characterization of ligand-effect modulators in Saccharomyces cerevisiae. Genetics 2000, 156, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Emter, R.; Heese-Peck, A.; Kralli, A. ERG6 and PDR5 regulate small lipophilic drug accumulation in yeast cells via distinct mechanisms. FEBS. Lett. 2002, 521, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Jensen-Pergakes, K.L.; Kennedy, M.A.; Lees, N.D.; Barbuch, R.; Koegel, C.; Bard, M. Sequencing, disruption, and characterization of the Candida albicans sterol methyltransferase (ERG6) gene: Drug susceptibility studies in erg6 mutants. Antimicrob. Agents Chemother. 1998, 42, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, F.F.M.; Paes, H.C.; Peconick, L.D.F.; Fonseca, F.L.; Marina, C.L.F.; Bocca, A.L.; Homem-de-Mello, M.; Rodrigues, M.L.; Albuquerque, P.; Nicola, A.M.; et al. Erg6 affects membrane composition and virulence of the human fungal pathogen Cryptococcus neoformans. Fungal Genet. Biol. 2020, 140, 103368. [Google Scholar] [CrossRef]
- Monk, B.C.; Mason, A.B.; Abramochkin, G.; Haber, J.E.; Seto-Young, D.; Perlin, D.S. The yeast plasma membrane proton pumping ATPase is a viable antifungal target. I. Effects of the cysteine-modifying reagent omeprazole. Biochim. Biophys. Acta 1995, 1239, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S.; Seto-Young, D.; Monk, B.C. The plasma membrane H(+)-ATPase of fungi. A candidate drug target? Ann. N. Y. Acad. Sci. 1997, 834, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.; Morsomme, P. The plasma membrane H+ -ATPase, a simple polypeptide with a long history. Yeast 2019, 36, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Fradin, C.; Kretschmar, M.; Nichterlein, T.; Gaillardin, C.; d’Enfert, C.; Hube, B. Stage-specific gene expression of Candida albicans in human blood. Mol. Microbiol. 2003, 47, 1523–1543. [Google Scholar] [CrossRef]
- Kolaczkowski, M.; van der Rest, M.; Cybularz-Kolaczkowska, A.; Soumillion, J.P.; Konings, W.N.; Goffeau, A. Anticancer drugs, ionophoric peptides, and steroids as substrates of the yeast multidrug transporter Pdr5p. J. Biol. Chem. 1996, 271, 31543–31548. [Google Scholar] [CrossRef] [Green Version]
- Juhnke, H.; Krems, B.; Kötter, P.; Entian, K.D. Mutants that show increased sensitivity to hydrogen peroxide reveal an important role for the pentose phosphate pathway in protection of yeast against oxidative stress. Mol. Gen. Genet. 1996, 252, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.M. Role of the glutathione/glutaredoxin and thioredoxin systems in yeast growth and response to stress conditions. Mol. Microbiol. 2001, 39, 533–541. [Google Scholar] [CrossRef]
- Chen, E.; Choy, M.S.; Petrényi, K.; Kónya, Z.; Erdődi, F.; Dombrádi, V.; Peti, W.; Page, R. Molecular Insights into the Fungus-Specific Serine/Threonine Protein Phosphatase Z1 in Candida albicans. mBio 2016, 7, e00872-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, K.; Kónya, Z.; Erdődi, F.; Farkas, I.; Dombrádi, V. Dissection of the regulatory role for the N-terminal domain in Candida albicans protein phosphatase Z1. PLoS ONE 2019, 14, e0211426. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Median (Range) MIC Values | Interaction | ||||
|---|---|---|---|---|---|---|
| MIC Alone | MIC in Combination | Median (Range) FICI | Type of Interaction | |||
| BM (mM) | MSB (mM) | BM (mM) | MSB (mM) | |||
| QMY23 (WT) | >4 a | 0.25 | 2 (2–4) | 0.06 (0.03–0.06) | 0.49 (0.49–1) | Synergy |
| cappz1 (KO) | >4 a | 0.25 | 0.125 (0.125–0.5) | 0.06 (0.008–0.06) | 0.49 (0.302–0.49) | Synergy |
| Significantly Enriched Gene Groups (Number of Genes Associated with the Group) | Number of Up- and Down-Regulated Genes (with Corrected p-Value) a | ||||||
|---|---|---|---|---|---|---|---|
| KO vs. WT | KO + MSB vs. WT + MSB | KO + BM + MSB vs. WT + BM + MSB | WT + MSB vs. WT | WT + BM + MSB vs. WT | KO + MSB vs. KO | KO + BM + MSB vs. KO | |
| Transmembrane transport b (365) | 46(3 × 10−2) | 103(3 × 10−5) | 27(9.9 × 10−7) | ||||
| Plasma membrane b (509) | 32 (1 × 10−5) | 132 (1.3 × 10−5) | 28 (6.6 × 10−5) | ||||
| Carbohydrate transport b (34) | 11(9 × 10−3) | 20(7.4 × 10−5) | 6(2.6 × 10−2) | ||||
| Oxidoreductase activity b (429) | 27 (2 × 10−4) | 64(2.9 × 10−7) | 127 (3.9 × 10−9) | 21 (5.3 × 10−12) 8 (5 × 10−3) | 60(7.24 × 10−17) | 110 (5.6 × 10−17) | 165(9.7 × 10−22) |
| Response to oxidative stress b (153) | 27(4 × 10−3) | 12 (7.2 × 10−8) | 26(1.1 × 10−7) | 43 (2.5 × 10−6) | 54 (1 × 10−3) | ||
| Antioxidant activity c (58) | 13(1.1 × 10−4) | 17 (1.3 × 10−2) | 6(2.5 × 10−6) | 16 (3.4 × 10−10) | 23 (8.9 × 10−9) | 26(2.8 × 10−6) | |
| Peroxisome b (70) | 18(1 × 10−4) | 38 (2.6 × 10−10) | 23 (2.0 × 10−4) | 35 (5.0 × 10−7) | |||
| RNA metabolic process b (776) | 92 (2.5 × 10−6) | 231 (5.4 × 10−22) | 252 (7 × 10−49) | 325 (4.1 × 10−56) | |||
| Ribosome b (194) | 76 (4.6 × 10−39) | 131 (7.8 × 10−58) | 73(4.7 × 10−16) | 95(6.3 × 10−20) | |||
| RNA transport b (114) | 47 (1.4 × 10−7) | 37 (8.4 × 10−5) | 51 (1.6 × 10−7) | ||||
| Alpha-amino acid metabolism b (132) | 9 (8 × 10−3) | 21(2.3 × 10−5) | 41(2.1 × 10−7) | 51 (5.2 × 10−5) | |||
| Branched-chain amino acid biosynthesis b (12) | 4 (3 × 10−3) | 6(2 × 10−3) | 9(1 × 10−3) | 10(4 × 10−3) | |||
| Glutamate metabolic process b (11) | 2(3 × 10−2) | ||||||
| Mitochondrion b (649) | 145 (1.5 × 10−9) | 201 (2.2 × 10−13) | |||||
| Iron homeostasis-related genes c (48) | 11 (7.8 × 10−9) | 7(4 × 10−2) | 5(2 × 10−7) | 9 (2.4 × 10−8) | 19(8 × 10−7) | 18(4 × 10−3) | |
| Glycolytic process b (17) | 7(1.6 × 10−7) | 14 (2.1 × 10−8) | 16 (4.4 × 10−8) | ||||
| Gluconeogenesis b (9) | 5 (9.3 × 10−6) | ||||||
| Ethanol fermentation pathway c (8) | 4 (3 × 10−2) | 4(7 × 10−3) | 7 (4.6 × 10−5) | ||||
| Glycogen biosynthesis pathway c (7) | 6(2.6 × 10−5) | 4(7 × 10−3) | 5(7 × 10−3) | ||||
| Fatty acid catabolic process b (18) | 15 (2.3 × 10−6) | 12(2 × 10−2) | |||||
| Ergosterol biosynthesis pathway c (20) | 8 (1.9 × 10−5) | 7 (3 × 10−2) | |||||
| Virulence Attributes | QMY23 (WT) | cappz1 (KO) | ||
|---|---|---|---|---|
| Untreated | +BM | Untreated | +BM | |
| Extracellular proteinase activity (Pz values) a | 0.72 ± 0.05 | 0.71 ± 0.06 | 0.74 ± 0.04 | 0.73 ± 0.05 |
| Secreted phospholipase activity (Pz values) a | 0.48 ± 0.06 | 0.44 ± 0.07 | 0.45 ± 0.06 | 0.43± 0.08 |
| Hypha formation (%) b | ||||
| 7 days | 15.0 ± 3.1 | 17.5 ± 1.8 | 9.7 ± 2.4 c | 11.1 ± 2.1 c |
| 10 days | 31.3 ± 3.3 | 33.2 ± 2.5 | 24.5 ± 2.1 c | 25.1 ± 2.9 c |
| Oxidative Stress Related Parameters | QMY23 (WT) | cappz1 (KO) | ||||||
|---|---|---|---|---|---|---|---|---|
| Untreated | +BM | +MSB | +BM + MSB | Untreated | +BM | +MSB | +BM + MSB | |
| glutathione reductase (mkat/kg protein) | 1.8 ± 0.4 | 1.8 ± 0.45 | 2.5 ± 0.2 c | 3.2 ± 0.6 c | 2.3 ± 0.15 b | 2.1 ± 0.15 | 3.3 ± 0.25 b,c | 5.0 ±0.85 b,c,d |
| glutathione peroxidase (mkat/kg protein) | 0.17 ± 0.02 | 0.19 ± 0.01 | 0.22 ± 0.02 c | 0.26 ± 0.03 c | 0.22 ± 0.04 b | 0.21 ± 0.02 | 0.34 ± 0.07 b,c,e | 0.44 ± 0.05 b,c,d |
| catalase (kat/kg protein) | 0.44 ± 0.07 | 0.46 ± 0.06 | 0.65 ± 0.08 c | 0.83 ± 0.17 c | 0.6 ± 0.11 | 0.59 ± 0.10 | 0.9 ± 0.17 b,c | 1.7 ± 0.47 b,c,d |
| superoxide dismutase (mU/mg protein) | 52 ± 2.3 | 50.8 ± 1.4 | 56.9 ± 3.3 | 59.4 ± 3.1 c | 56.6 ± 2.3 | 54.5 ± 2.4 | 65.3 ± 4.5 c | 69.5 ± 7.3 c |
| DCF (nmol DCF/OD640) a | 13.4 ± 3.1 | 12.5 ± 3.3 | 21.5 ± 2.0 c | 26.5 ± 2.6 c,d | 33.1 ± 2.3 b | 32.8 ± 3.0 b | 60.3 ± 8.7 b,c,e | 95.2 ± 11.9 b,c,d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakab, Á.; Emri, T.; Csillag, K.; Szabó, A.; Nagy, F.; Baranyai, E.; Sajtos, Z.; Géczi, D.; Antal, K.; Kovács, R.; et al. The Negative Effect of Protein Phosphatase Z1 Deletion on the Oxidative Stress Tolerance of Candida albicans Is Synergistic with Betamethasone Exposure. J. Fungi 2021, 7, 540. https://doi.org/10.3390/jof7070540
Jakab Á, Emri T, Csillag K, Szabó A, Nagy F, Baranyai E, Sajtos Z, Géczi D, Antal K, Kovács R, et al. The Negative Effect of Protein Phosphatase Z1 Deletion on the Oxidative Stress Tolerance of Candida albicans Is Synergistic with Betamethasone Exposure. Journal of Fungi. 2021; 7(7):540. https://doi.org/10.3390/jof7070540
Chicago/Turabian StyleJakab, Ágnes, Tamás Emri, Kinga Csillag, Anita Szabó, Fruzsina Nagy, Edina Baranyai, Zsófi Sajtos, Dóra Géczi, Károly Antal, Renátó Kovács, and et al. 2021. "The Negative Effect of Protein Phosphatase Z1 Deletion on the Oxidative Stress Tolerance of Candida albicans Is Synergistic with Betamethasone Exposure" Journal of Fungi 7, no. 7: 540. https://doi.org/10.3390/jof7070540