
Comparison of Two Schizophyllum commune Strains in Production of Acetylcholinesterase Inhibitors and Antioxidants from Submerged Cultivation


 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Submerged Cultivation and Preparation of Extracts
2.3. Determination of AChE Activity
2.4. Determination of Antioxidant Activity
2.4.1. ABTS Assay
2.4.2. FRAP Assay
2.4.3. Superoxide Anion Radical Scavenging (SOA) Assay
2.4.4. OH Assay
2.4.5. NO Assay
2.5. Antibacterial Activity
2.6. Determination of Total Phenolic Content
2.7. Fourier Transform Infrared Spectroscopy Analysis (FTIR)
2.8. Hydrolysis
2.9. LC–MS/MS Analysis
2.10. Statistical Analysis
3. Results
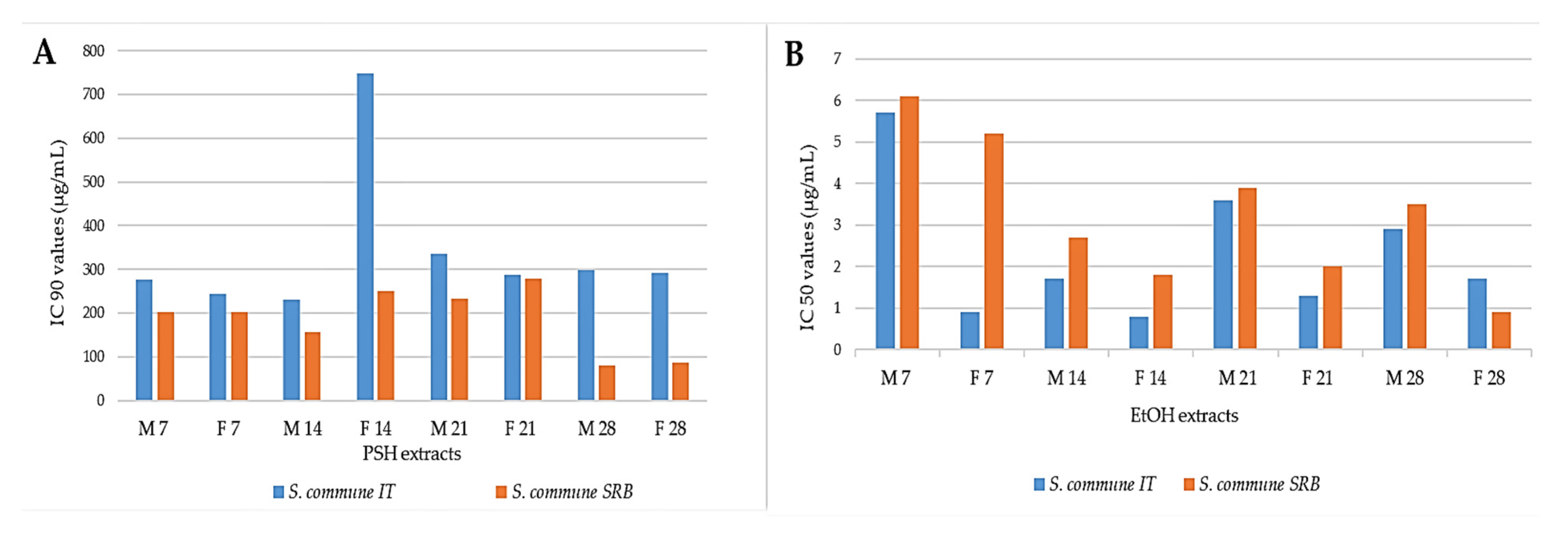
3.1. Inhibition of AChEEnzyme
3.2. Determination of Antioxidant Activity
3.2.1. DPPH Assay
3.2.2. ABTS Assay
3.2.3. FRAP Assay
3.2.4. SOA Assay
3.2.5. OH Assay
3.2.6. NO Assay
3.3. Determination of Antibacterial Activity
3.4. Total Phenol (TP) Content and Correlation Analysis
3.5. Chemical Characterization of Extracts
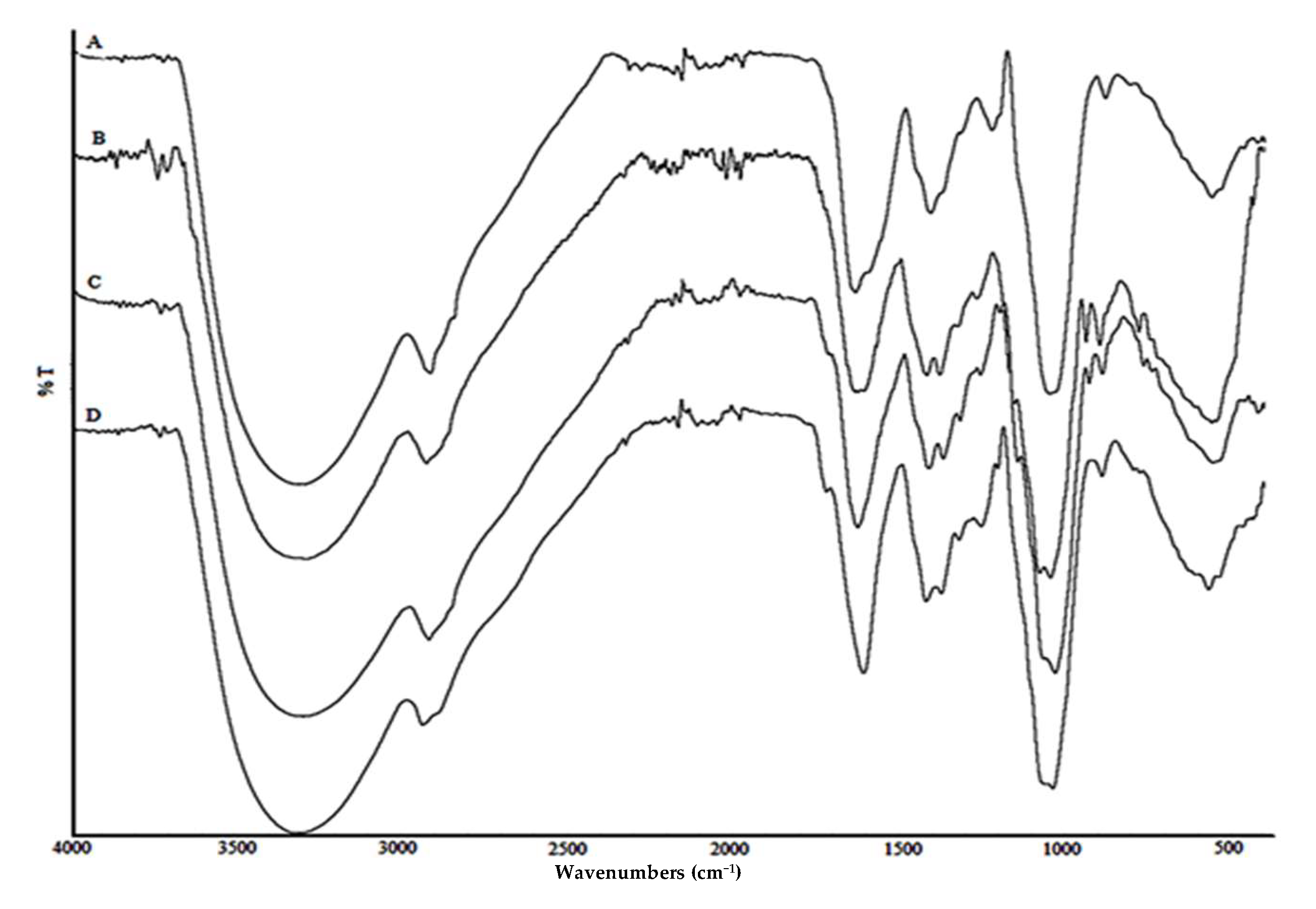
3.5.1. FTIR Analysis
3.5.2. Monosaccharide Composition
3.5.3. LC–MS/MS Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dulay, R. Antioxidant activity and total phenolic content of Volvariellavolvacea and Schizophyllum commune mycelia cultured in indigenous liquid media. Mycosphere 2016, 7, 131–138. [Google Scholar] [CrossRef]
- Veurink, G.; Perry, G.; Singh, S.K. Role of antioxidants and a nutrient rich diet in Alzheimer’s disease: AD and Nutraceuticals. Open Biol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Moneim, A.E.A. Oxidant/Antioxidant Imbalance and the Risk of Alzheimer’s Disease. Curr. Alzheimer Res. 2015, 12, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Nunomura, A.; Perry, G.; Aliev, G.; Hirai, K.; Takeda, A.; Balraj, E.K.; Jones, P.K.; Ghanbari, H.; Wataya, T.; Shimohama, S.; et al. Oxidative Damage Is the Earliest Event in Alzheimer Disease. J. Neuropathol. Exp. Neurol. 2001, 60, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Jomova, K.; Vondrakova, D.; Lawson, M.; Valko, M. Metals, oxidative stress and neurodegenerative disorders. Mol. Cell. Biochem. 2010, 345, 91–104. [Google Scholar] [CrossRef]
- Chopra, K.; Misra, S.; Kuhad, A. Current perspectives on pharmacotherapy of Alzheimer’s disease. Expert Opin. Pharmacother. 2011, 12, 335–350. [Google Scholar] [CrossRef]
- Pinho, B.R.; Ferreres, F.; Valentão, P.; Andrade, P.B. Nature as a source of metabolites with cholinesterase-inhibitory activity: An approach to Alzheimer’s disease treatment. J. Pharm. Pharmacol. 2013, 65, 1681–1700. [Google Scholar] [CrossRef]
- Hostettmann, K.; Borloz, A.; Urbain, A.; Marston, A. Natural Product Inhibitors of Acetylcholinesterase. Curr. Org. Chem. 2006, 10, 825–847. [Google Scholar] [CrossRef]
- Moniruzzaman, A.; Hossain, S.; Sarker, J.; Rahman, S.M.A.; Rashid, M.; Rahman, M. In vitro antioxidant and cholinesterase inhibitory activities of methanolic fruit extract of Phyllanthus acidus. BMC Complement. Altern. Med. 2015, 15, 403. [Google Scholar] [CrossRef] [Green Version]
- Janjušević, L.; Karaman, M.; Šibul, F.; Tommonaro, G.; Iodice, C.; Jakovljević, D.; Pejin, B. The lignicolous fungus Trametes versicolor (L.) Lloyd (1920): A promising natural source of antiradical and AChE inhibitory agents. J. Enzym. Inhib. Med. Chem. 2017, 32, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Liu, H.; Guo, K.; Chen, L.; Yang, M.; Chen, Q. Research Advances and Detection Methodologies for Microbe-Derived Acetylcholinesterase Inhibitors: A Systemic Review. Molecules 2017, 22, 176. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Niksic, M.; Jakovljevic, D.; Helsper, J.P.; Van Griensven, L.J. Antioxidative and immunomodulating activities of polysaccharide extracts of the medicinal mushrooms Agaricusbisporus, Agaricusbrasiliensis, Ganoderma lucidum and Phellinus linteus. Food Chem. 2011, 129, 1667–1675. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, S.; Cheung, P.; Wang, Q. Antitumor polysaccharides from mushrooms: A review on their isolation process, structural characteristics and antitumor activity. Trends Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- Mahmoud, M.G.; Ibrahim, A.Y.; Asker, M.M.S.; El Sayed, O.H. Therapeutic potential and structural elucidation of a water-soluble polysaccharide of a wild edible mushroom Agaricusbisporus against neurodegenerative disease, Alzheimer. World J. Pharm. Sci. 2014, 2, 1136–1145. [Google Scholar]
- Phan, C.-W.; David, P.; Vikineswary, S. Edible and Medicinal Mushrooms: Emerging Brain Food for the Mitigation of Neurodegenerative Diseases. J. Med. Food. 2017, 20, 1–10. [Google Scholar] [CrossRef]
- Karaman, M.; Stahl, M.; Vulić, J.; Vesić, M.; Čanadanović-Brunet, J. Wild-growing lignicolous mushroom species as sources of novel agents with antioxidative and antibacterial potentials. Int. J. Food Sci. Nutr. 2014, 65, 311–319. [Google Scholar] [CrossRef]
- Umeo, S.; Souza, G.; Rapachi, P.; Garcia, D.; Paccola-Meirelles, L.; Valle, J.S.D.; Colauto, N.; Linde, G. Short Communication Screening of basidiomycetes in submerged cultivation based on antioxidant activity. Genet. Mol. Res. 2015, 14, 9907–9914. [Google Scholar] [CrossRef]
- Ohm, R.A.; De Jong, J.F.; Lugones, L.G.; Aerts, A.; Kothe, E.; Stajich, J.E.; De Vries, R.P.; Record, E.; Levasseur, A.; Baker, S.E.; et al. Genome sequence of the model mushroom Schizophyllum commune. Nat. Biotechnol. 2010, 28, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, R.; Smith, J.E.; Rowan, N.J. Medicinal Mushrooms and Cancer Therapy: Translating a traditional practice into Western medicine. Perspect. Biol. Med. 2006, 49, 159–170. [Google Scholar] [CrossRef]
- Du, B.; Yang, Y.; Bian, Z.; Xu, B. Characterization and Anti-Inflammatory Potential of an Exopolysaccharide from Submerged Mycelial Culture of Schizophyllum commune. Front. Pharmacol. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Hermann, J. Fungi on Trees: Saprophytes and Parasites That Grow on Wood; Patzer: Blunder, Germany, 2005. [Google Scholar]
- Ren, L.; Hemar, Y.; Perera, C.O.; Lewis, G.D.; Krissansen, G.W.; Buchanan, P.K. Antibacterial and antioxidant activities of aqueous extracts of eight edible mushrooms. Bioact. Carbohydr. Diet. Fibre. 2014, 3, 41–51. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Espín, J.C.; Soler-Rivas, C.; Wichers, H.J. Characterization of the Total Free Radical Scavenger Capacity of Vegetable Oils and Oil Fractions Using 2,2-Diphenyl-1-picrylhydrazyl Radical. J. Agric. Food Chem. 2000, 48, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Link. Biomacromol. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.; Aruoma, O.I. The deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Green, L.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Limbago, B. M100-S11, Performance standards for antimicrobial susceptibility testing. Clin. Microbiol. Newsl. 2001, 23, 49. [Google Scholar] [CrossRef]
- Weinstein, M.P.; Limbago, B.; Patel, J.B.; Mathers, A.J.; Burnham, C.; Mazzulli, T.; Campeau, S.; Munro, S.D.; Conville, P.S.; Doer, C.; et al. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 9th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012; Volume 32. [Google Scholar]
- Karaman, M.; Kaisarevic, S.; Somborski, J.; Kebert, M.; Matavulj, M. Biological activities of the lignicolous fungus Meripilus giganteus (Pers.: Pers.) Karst. Arch. Biol. Sci. 2009, 61, 853–861. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M.; Lester, P. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Chaplin, M.F.; Kennedy, J.F. Carbohydrate Analysis: A Practical Approach, 2nd ed.; IRL Press: Oxford, UK, 1986; pp. 1–36. [Google Scholar]
- Orčić, D.; Francišković, M.; Bekvalac, K.; Svirčev, E.; Beara, I.; Lesjak, M.; Mimica-Dukic, N. Quantitative determination of plant phenolics in Urticadioica extracts by high-performance liquid chromatography coupled with tandem mass spectrometric detection. Food Chem. 2014, 143, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Mousaviasl, S.; Saleh, T.; Shojaosadati, S.A.; Boddohi, S. Synthesis and characterization of schizophyllan nanogels via inverse emulsion using biobased materials. Int. J. Biol. Macromol. 2018, 120, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wu, L.-X.; Qiao, Z.-R.; Cai, W.-D.; Ma, H. Effect of different drying methods on the product quality and bioactive polysaccharides of bitter gourd (Momordica charantia L.) slices. Food Chem. 2019, 271, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Niksic, M.; Vrvić, M.M.; Todorovic, N.; JakovljeviĆ, D.; Van Griensven, L.J. Antioxidative activities and chemical characterization of polysaccharide extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinusedodes and Trametes versicolor. J. Food Compos. Anal. 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Šandula, J.; Kogan, G.; Kačuráková, M.; Machová, E. Microbial (1→3)-β-d-glucans, their preparation, physico-chemical characterization and immunomodulatory activity. Carbohydr. Polym. 1999, 38, 247–253. [Google Scholar] [CrossRef]
- Chen, Z.; Yin, C.; Fan, X.; Ma, K.; Yao, F.; Zhou, R.; Shi, D.; Cheng, W.; Gao, H. Characterization of physicochemical and biological properties of Schizophyllum commune polysaccharide extracted with different methods. Int. J. Biol. Macromol. 2020, 156, 1425–1434. [Google Scholar] [CrossRef]
- Mathlouthi, M.; Koenig, J.L. Vibrational Spectra of Carbohydrates. Adv. Carbohydr. Chem. Biochem. 1987, 44, 7–89. [Google Scholar] [CrossRef]
- Jamshidian, H.; Shojaosadati, S.A.; Vilaplana, F.; Mousavi, S.M.; Soudi, M.R. Characterization and optimization of schizophyllan production from date syrup. Int. J. Biol. Macromol. 2016, 92, 484–493. [Google Scholar] [CrossRef]
- Orhan, I.E.; Kartal, M.; Tosun, F.; Şener, B. Screening of Various Phenolic Acids and Flavonoid Derivatives for their Anticholinesterase Potential. Z. Nat. C 2007, 62, 829–832. [Google Scholar] [CrossRef]
- Szwajgier, D. Anticholinesterase Activity of Phenolic Acids and their Derivatives. Z. Nat. C 2013, 68, 125–132. [Google Scholar] [CrossRef]
- Tešanović, K.; Pejin, B.; Šibul, F.; Matavulj, M.; Rašeta, M.; Janjušević, L.; Karaman, M. A comparative overview of antioxidative properties and phenolic profiles of different fungal origins: Fruiting bodies and submerged cultures of Coprinus comatus and Coprinellus truncorum. J. Food Sci. Technol. 2017, 54, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, Y.; Jannat, S.; Jung, H.A.; Choi, R.J.; Roy, A.; Choi, J.S. Anti-Alzheimer’s disease potential of coumarins from Angelica decursiva and Artemisia capillaris and structure-activity analysis. Asian Pac. J. Trop. Med. 2016, 9, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, X.; Jin, G.; Yang, X.; Zhang, Y. Polysaccharides from Pleurotus ostreatus alleviate cognitive impairment in a rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 92, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.; Ismail, S.M.; Aminudin, N.; Shuib, A.S.; Lau, B.F. Evaluation of Selected Culinary-Medicinal Mushrooms for Antioxidant and ACE Inhibitory Activities. Evid. Based Complement. Altern. Med. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, M.; Tešanović, K.; Novaković, A.; Šibul, F.; Živić, M.; Stevanović, K.; Pejin, B. Fatty Acids Predominantly Affect Anti-Hydroxyl Radical Activity and FRAP Value: The Case Study of Two Edible Mushrooms. Antioxidants 2019, 8, 480. [Google Scholar] [CrossRef] [Green Version]
- Chandrawanshi, N.K.; Tandia, D.K.; Jadhav, S.K. Nutraceutical Properties Evaluation of Schizophyllum commune. Indian J. Sci. Res. 2017, 13, 57–62. [Google Scholar]
- Tripathi, A.M.; Tiwary, B.N. Biochemical constituents of a wild strain of Schizophyllum commune isolated from Achanakmar-Amarkantak Biosphere Reserve (ABR), India. World J. Microbiol. Biotechnol. 2013, 29, 1431–1442. [Google Scholar] [CrossRef]
- Emsen, B.; Kaya, A.K.A.; Cinar, S.; Aasim, M.; Sadi, G. In vitro cytotoxicity, antibacterial and antioxidant properties of various extracts from Schizophyllum commune Fr. Fresenius Environ. Bull. 2017, 26, 1144–1153. [Google Scholar]
- Razak, D.L.A.; Fadzil, N.H.M.; Jamaluddin, A.; Rashid, N.Y.A.; Sani, N.A.; Manan, M.A. Effects of different extracting conditions on anti-tyrosinase and antioxidant activities of Schizophyllum commune fruit bodies. Biocatal. Agric. Biotechnol. 2019, 19, 101116. [Google Scholar] [CrossRef]
- Basso, V.; Schiavenin, C.; Mendonça, S.; De Siqueira, F.G.; Salvador, M.; Camassola, M. Chemical features and antioxidant profile by Schizophyllum commune produced on different agroindustrial wastes and byproducts of biodiesel production. Food Chem. 2020, 329, 127089. [Google Scholar] [CrossRef] [PubMed]
- Jiamworanunkul, S. Effective antioxidant production through submerged fermentation of edible mushrooms. Thai J. Pharm. Sci. 2019, 43, 213–218. [Google Scholar]
- Mayakrishnan, V.; Abdullah, N.; Abidin, M.H.Z.; Fadzil, N.H.M.; Johari, N.M.K.; Aminudin, N.; Abidin, N.Z. Investigation of the Antioxidative Potential of Various Solvent Fractions from Fruiting Bodies of Schizophyllum commune (Fr.) Mushrooms and Characterization of Phytoconstituents. J. Agric. Sci. 2013, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Velderrain-Rodríguez, G.R.; Torres-Moreno, H.; Villegas-Ochoa, M.A.; Ayala-Zavala, J.F.; Robles-Zepeda, R.E.; Wall-Medrano, A.; González-Aguilar, G.A. Gallic Acid Content and an Antioxidant Mechanism Are Responsible for the Antiproliferative Activity of ‘Ataulfo’ Mango Peel on LS180 Cells. Molecules 2018, 23, 695. [Google Scholar] [CrossRef] [Green Version]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Jakovljevic, D.; Todorovic, N.; Vunduk, J.; Petrovic, P.; Niksic, M.; Vrvic, M.; Van Griensven, L.J. Antioxidants of Edible Mushrooms. Molecules 2015, 20, 19489–19525. [Google Scholar] [CrossRef] [Green Version]
- Duncan, S.H.; Leitch, E.C.M.; Stanley, K.N.; Richardson, A.J.; Laven, R.; Flint, H.J.; Stewart, C.S. Effects of esculin and esculetin on the survival of Escherichia coli O157 in human faecal slurries, continuous-flow simulations of the rumen and colon and in calves. Br. J. Nutr. 2004, 91, 749–755. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.J.; Ferreira, I.C.F.R.; Froufe, H.J.C.; Abreu, R.M.V.; Martins, A.; Pintado, M. Antimicrobial activity of phenolic compounds identified in wild mushrooms, SAR analysis and docking studies. J. Appl. Microbiol. 2013, 115, 346–357. [Google Scholar] [CrossRef]
- Díaz-Gómez, R.; Toledo-Araya, H.; López-Solís, R.; Obreque-Slier, E. Combined effect of gallic acid and catechin against Escherichia coli. LWTFood Sci. Technol. 2014, 59, 896–900. [Google Scholar] [CrossRef]
- Arun, G.; Eyini, M.; Gunasekaran, P. Characterization and biological activities of extracellular melanin produced by Schizophyllum commune (Fries). Indian J. Exp. Biol. 2015, 53, 380–387. [Google Scholar]



{kind=link}
{kind=link}
| Extracts | DPPH 1 (IC50) (µg/mL) | ABTS 2(mg TE/g d.w.) | FRAP 3(mg AAE/g d.w.) | ||||
|---|---|---|---|---|---|---|---|
| S. commune IT | PSH | EtOH | PSH | EtOH | PSH | EtOH | |
| F | 7 days | 16.03 ± 4.30 a | 72.65 ± 9.15 a | 6.64 ± 0.66 a | 5.17 ± 0.03 a | 65.11 ± 6.27 a | 42.58 ± 3.47 a |
| 14 days | 107.94 ± 2.36 a | 63.67 ± 2.65 a | 8.20 ± 0.94 a | 4.95 ± 0.41 a | 66.73 ± 12.11 a | 99.00 ± 2.73 a | |
| 21 days | 74.76 ± 19.70 a | 55.43 ± 1.89 a | 5.69 ± 0.52 a | 5.27 ± 0.22 a | 82.96 ± 0.94 a | 91.65 ± 0.96 a | |
| 28 days | 189.81 ± 7.96 a | 69.15 ± 3.98 a | 2.86 ± 1.30 a | 3.38 ± 0.06 a | 28.44 ± 7.72 a | 47.82 ± 5.28 a | |
| M | 7 days | 94.89 ± 8.41 a | 82.69 ± 1.93 a | 5.16 ± 1.63 a | 5.29 ± 0.05 a | 40.03 ± 3.41 a | 66.39 ± 3.84 a |
| 14 days | 14.45 ± 6.83 a | 49.34 ± 0.65 a | 1.63 ± 1.71 a | 5.20 ± 0.08 a | 6.49 ± 7.40 a | 81.30 ± 1.98 a | |
| 21 days | 85.39 ± 1.61 a | 120.63 ± 3.29 a | 7.49 ± 0.67 a | 5.84 ± 0.06 a | 79.12 ± 10.77 a | 14.22 ± 1.32 a | |
| 28 days | 93.48 ± 0.01 a | 105.39 ± 1.66 a | 5.12 ± 0.39 a | 4.85 ± 0.19 a | 33.67 ± 5.35 a | 32.46 ± 1.27 a | |
| S. commune SRB | |||||||
| F | 7 days | 15.76 ± 0.63 a | 70.86 ± 2.52 b | 2.60 ± 0.14 a | 3.80 ± 0.08 a | 0.28 ± 0.09 b | 61.81 ± 4.78 a,c |
| 14 days | 32.42 ± 3.09 a | 53.61 ± 2.32 b | 7.40 ± 0.19 a | 3.26 ± 0.96 a | 9.69 ± 3.67 b | 107.86 ± 12.8 a,c | |
| 21 days | 30.18 ± 2.25 a | 55.96 ± 1.31 b | 7.62 ± 0.62 a | 4.65 ± 0.31 a | 57.10 ± 2.35 b | 89.97 ± 12.81 a,c | |
| 28 days | 69.69 ± 3.93 a | 67.31 ± 1.03 b | 3.25 ± 1.98 a | 3.40 ± 0.09 a | 4.36 ± 1.36 b | 80.28 ± 3.21 a,c | |
| M | 7 days | 18.92 ± 6.12 a | 377.71 ± 14.85 b | 7.78 ± 0.41 a | 6.01 ± 0.07 a | 20.89 ± 1.67 b | 11.44 ± 0.84 a,d |
| 14 days | 71.55 ± 3.43 a | 74.65 ± 1.74 b | 3.17 ± 0.44 a | 3.02 ± 0.10 a | 9.55 ± 0.51 b | 5.99 ± 0.20 a,d | |
| 21 days | 69.13 ± 4.24 a | 164.28 ± 2.52 b | 2.30 ± 0.31 a | 5.43 ± 0.53 a | 5.61 ± 1.83 b | 10.77 ± 1.05 a,d | |
| 28 days | 51.41 ± 11.27 a | 137.5 ± 5.87 b | 2.14 ± 0.35 a | 5.49 ± 0.23 a | 0.90 ± 1.91 b | 10.91 ± 0.46 a,d | |
| Extracts | SOA (IC50) (µg/mL) | OH (IC25) (µg/mL) | NO (IC25) (µg/mL) | TP (mgGAE/g.d.w.) | |
|---|---|---|---|---|---|
| S. commune IT | |||||
| F | 7 days | 526.68 ± 152.45 a | 103.02 ± 12.23 a | 614.02 ± 14.79 a | 69.48 ± 1.00 a |
| 14 days | 652.14 ± 6.56 a | 124.27 ± 7.00 a | 965.43 ± 129.75 a | 75.23 ± 1.04 a | |
| 21 days | 90% 1 | 110.85 ± 2.31 a | 707.05 ± 34.84 a | 77.52 ± 0.97 a | |
| 28 days | 90% 1 | 10.80 ± 0.54 a | 1161.41 ± 48.90 a | 68.70 ± 0.30 a | |
| M | 7 days | 602.41 ± 0.01 a | 20.20 ± 1.36 a | 788.80 ± 36.36 a | 53.49 ± 1.97 a |
| 14 days | 216.98 ± 45.84 a | 228.36 ± 3.25 a | 1182.78 ± 36.35 a | 76.65 ± 1.30 a | |
| 21 days | 634.14 ± 38.25 a | 44.29 ± 4.20 a | n.a. 2 | 42.74 ± 1.40 a | |
| 28 days | 492.92 ± 125.11 a | 84.74 ± 2.36 a | n.a. 2 | 70.01 ± 0.65 a | |
| S. commune SRB | |||||
| F | 7 days | 635.74 ± 8.99 a | 494.34 ± 59.08 a | 1794.48 ± 13.21 a | 84.60 ± 1.64 a |
| 14 days | 90% 1 | 257.92 ± 0.01 a | <IC25 3 | 81.93 ± 0.81 a | |
| 21 days | 90% 1 | 261.38 ± 0.01 a | <IC25 3 | 78.27 ± 1.75 a | |
| 28 days | 90% 1 | 89.69 ± 0.38 a | <IC25 3 | 76.14 ± 1.42 a | |
| M | 7 days | 161.60 ± 2.06 a | 170.57 ± 0.35 a | 737.12 ± 32.91 a | 40.22 ± 0.82 a |
| 14 days | 253.14 ± 17.33 a | 163.94 ± 19.22 a | 145.88 ± 2.83 a | 82.62 ± 0.99 a | |
| 21 days | 429.60 ± 44.01 a | 89.45 ± 0.01 a | 785.34 ± 39.03 a | 63.28 ± 1.28 a | |
| 28 days | 218.01 ± 13.59 a | 85.57 ± 0.12 a | 112.19 ± 19.73 a | 2.09 ± 0.06 a | |
| Extracts | Class of Analyzed Compounds | |||
|---|---|---|---|---|
| Hydroxybenzoic Acids | Cyclohexane Carboxylic Acid | |||
| p-Hydroxybenzoic Acid | Protocatechuic Acid | Gallic Acid | Quinic Acid | |
| PSH | ||||
| IT F 7 | 5.50 | 3.17 | <12.2 * | <3.05 * |
| IT M 7 | 1.87 | <1.525 * | <12.2 * | <3.05 * |
| IT M 14 | 8.28 | <1.525 * | <12.2 * | <3.05 * |
| SRB F 7 | <1.525 * | <1.525 * | <12.2 * | 4.29 |
| EtOH | ||||
| IT F 14 | 3.81 | <1.525 * | <12.2 * | 13.24 |
| IT F 21 | 9.51 | <1.525 * | <12.2 * | <3.05 * |
| IT M 14 | 4.10 | <1.525 * | <12.2 * | 8.57 |
| IT M 21 | 9.66 | <1.525 * | <12.2 * | <3.05 * |
| IT M 28 | 22.19 | 5.41 | <12.2 * | 4.82 |
| SRB F 7 | 12.14 | <1.525 * | <12.2 * | 15.51 |
| SRB F 14 | 3.77 | <1.525 * | 75.77 | <3.05 * |
| SRB F 21 | 11.25 | 1.90 | <12.2 * | <3.05 * |
| SRB F 28 | 7.69 | <1.525 * | <12.2 * | <3.05 * |
| SRB M 7 | 5.93 | <1.525 * | <12.2 * | 19.52 |
| SRB M 14 | 5.94 | <1.525 * | <12.2 * | 20.06 |
| SRB M 21 | 6.52 | <1.525 * | <12.2 * | 5.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mišković, J.; Karaman, M.; Rašeta, M.; Krsmanović, N.; Berežni, S.; Jakovljević, D.; Piattoni, F.; Zambonelli, A.; Gargano, M.L.; Venturella, G. Comparison of Two Schizophyllum commune Strains in Production of Acetylcholinesterase Inhibitors and Antioxidants from Submerged Cultivation. J. Fungi 2021, 7, 115. https://doi.org/10.3390/jof7020115
Mišković J, Karaman M, Rašeta M, Krsmanović N, Berežni S, Jakovljević D, Piattoni F, Zambonelli A, Gargano ML, Venturella G. Comparison of Two Schizophyllum commune Strains in Production of Acetylcholinesterase Inhibitors and Antioxidants from Submerged Cultivation. Journal of Fungi. 2021; 7(2):115. https://doi.org/10.3390/jof7020115
Chicago/Turabian StyleMišković, Jovana, Maja Karaman, Milena Rašeta, Nenad Krsmanović, Sanja Berežni, Dragica Jakovljević, Federica Piattoni, Alessandra Zambonelli, Maria Letizia Gargano, and Giuseppe Venturella. 2021. "Comparison of Two Schizophyllum commune Strains in Production of Acetylcholinesterase Inhibitors and Antioxidants from Submerged Cultivation" Journal of Fungi 7, no. 2: 115. https://doi.org/10.3390/jof7020115