
The Relationship between Endophytic Fungi of Chimonanthus praecox and Volatile Metabolites under Different Circadian Rhythms and Blooming Stages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Flower Samples Collection
2.3. Floral Volatile Collection and Identification
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Data Processing and Statistical Analysis
3. Results
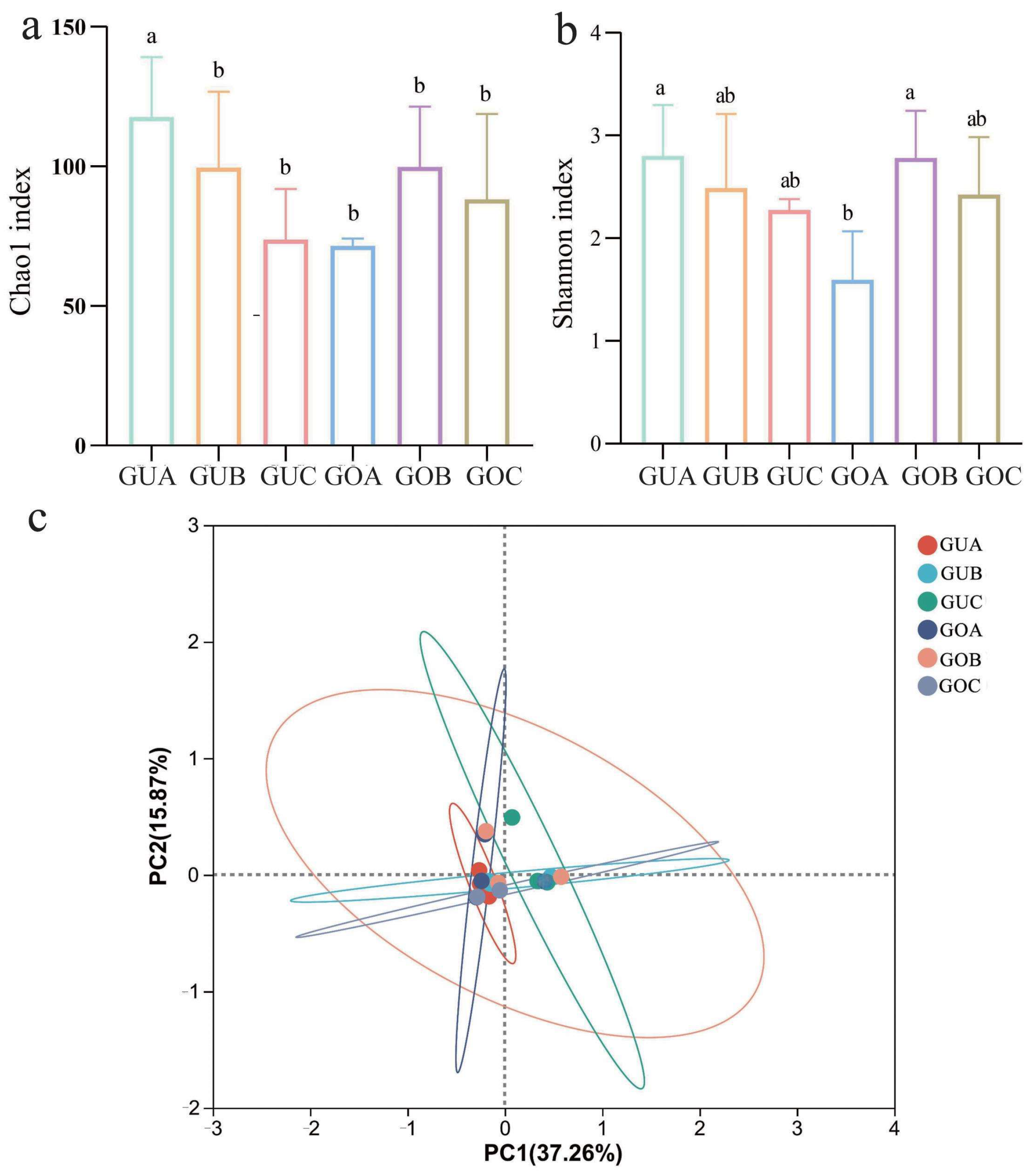
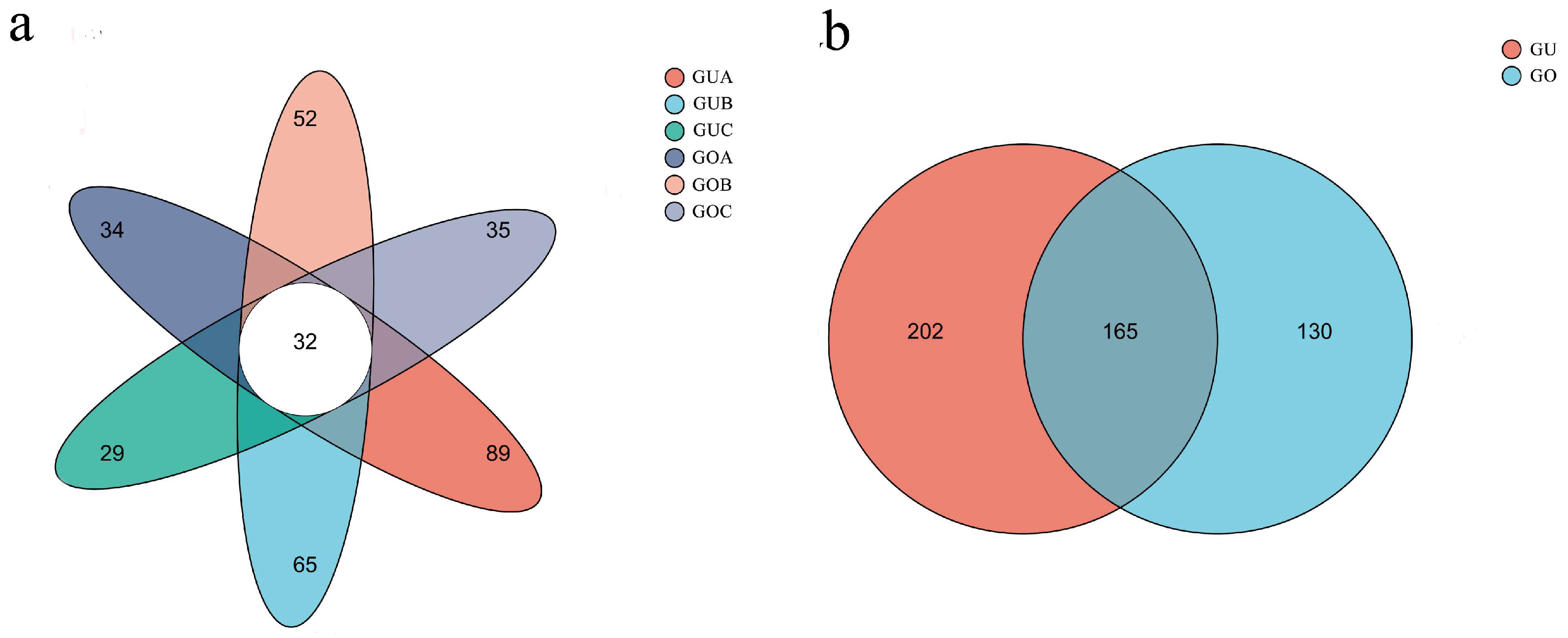
3.1. Alpha and Beta Diversity of Endophytic Fungi
3.2. Taxonomic Analysis of Endophytic Fungal Communities
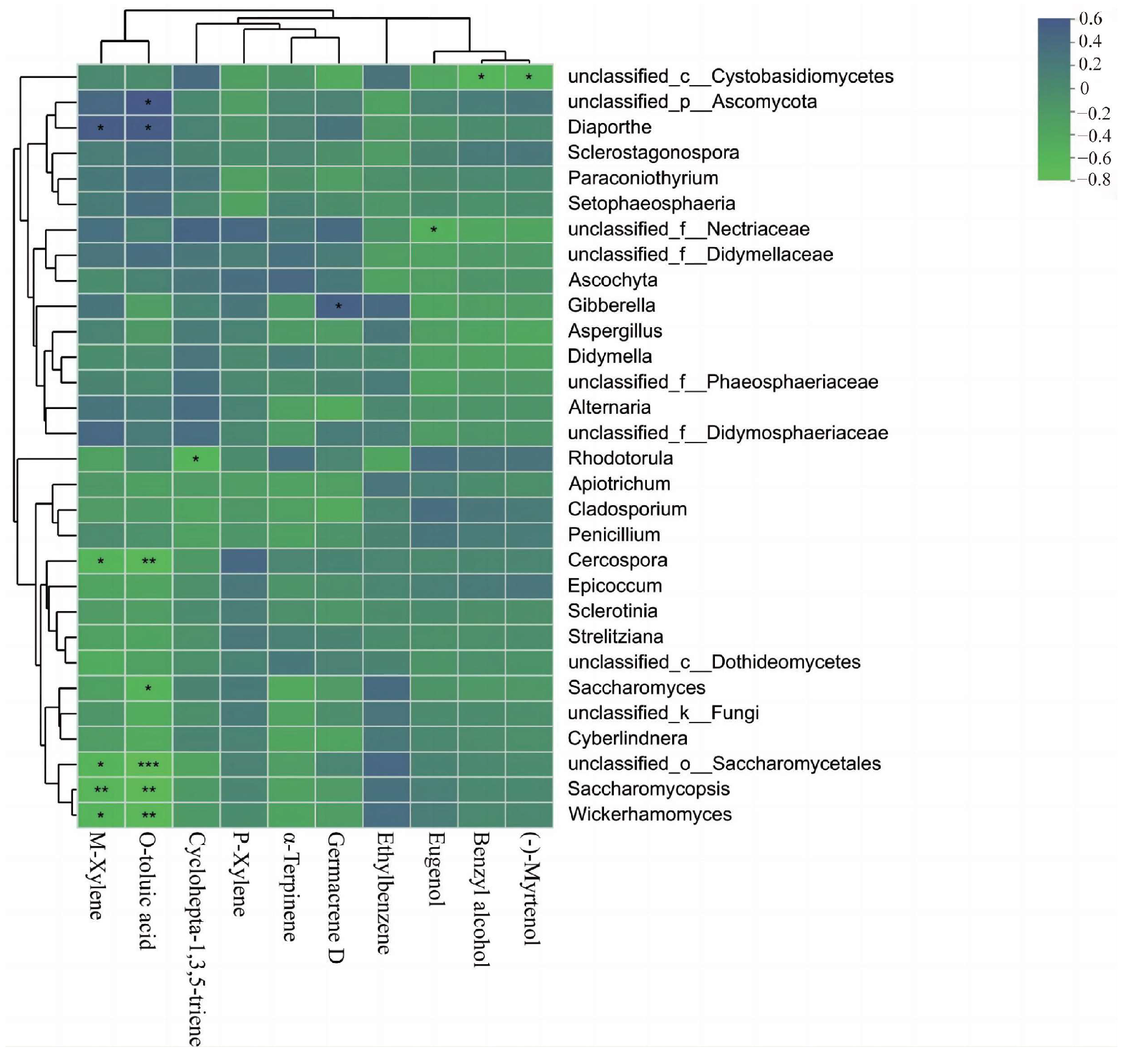
3.3. Correlation Analysis between Endophytic Fungi and Floral Volatiles in C. praecox
4. Discussion
4.1. Endophytic Fungal Diversity in C. praecox Is Altered by Circadian Rhythm and Blooming Stage
4.2. Dominant Fungal Communities under Different Circadian Rhythms and in Different Blooming Stages
4.3. Relationship between Endophytic Fungi and Floral Volatiles in C. praecox
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arnold, A.E. Understanding the diversity of foliar endophytic fungi: Progress, challenges, and frontiers. Fungal Biol. Rev. 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Li, P.Q.; Mou, Y.; Shan, T.J.; Xu, J.M.; Li, Y.; Lu, S.Q.; Zhou, L.G. Effects of polysaccharide elicitors from endophytic Fusarium oxysporium Dzf17 on growth and diosgenin production in cell suspension culture of Dioscorea zingiberensis. Molecules 2011, 16, 9003–9016. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Q.; Lu, X.D.; Okane, I.; Kakishima, M. Endophytic fungal community in stems and leaves of plants from desert areas in China. Mycol. Prog. 2012, 11, 781–790. [Google Scholar] [CrossRef]
- Muhammad, W.; Abdul, L.K.; Muhammad, H.; Raheem, S.; Sang, M.K.; Jong, G.K.; In, J.L. Endophytic fungi promote plant growth and mitigate the adverse effects of stem rot: An example of Penicillium citrinum and Aspergillus terreus. J. Plant Interact. 2015, 10, 280–287. [Google Scholar]
- Kornsakulkarn, J.; Choowong, W.; Rachtawee, P.; Boonyuen, N.; Kongthong, S.; Isaka, M.; Thongpanchang, C. Bioactive hydroanthraquinones from endophytic fungus Nigrospora sp. Phytochem. Lett. 2018, 24, 46–50. [Google Scholar] [CrossRef]
- Li, S.J.; Zhang, X.; Wang, X.H.; Zhao, C.Q. Novel natural compounds from endophytic fungi with anticancer activity. Eur. J. Med. Chem. 2018, 157, 960–961. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Liu, S.M.; Chu, Z.M. Isolation and Identification Endophytic Fungi from Lilium Siberia and Analysis of Volatile Components. Food Ind. 2013, 34, 203–205. [Google Scholar]
- Chukalo, C.E.; Krishna, C.R. Endophytic mycoflora and their bioactive compounds from azadirachta indica: A comprehensive review. J. Fungi 2018, 4, 42. [Google Scholar]
- Wu, X.M.; Ren, W.M.; Yang, X.D.; Wang, Y.H. Isolation of endophytic fungi and screening of salidrosid-producing strains from Rhodiola sachalinensis. Lishizhen Med. Mater. Medica 2014, 25, 2769–2772. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=SZGY201411087&DbName=CJFQ2014 (accessed on 6 June 2022).
- Li, H.; Yan, S.; Mi, Z.; Zuo, P.; Li, T.; Wei, Y.L.; Lin, L.B. Diversity and cold adaptation of endophytic fungi from five dominant plant species collected from the Baima Snow Mountain Southwest China. Fungal Divers. 2012, 54, 79–86. [Google Scholar] [CrossRef]
- Martin, U.; Almut, R.; Knut, F.; Peter, O.; Wilfried, M. Species richness and distribution patterns of leaf-inhabiting endophytic fungi in a temperate forest canopy. Mycol. Prog. 2007, 6, 201–212. [Google Scholar]
- Shi, Y.W.; Li, C.; Yang, H.M.; Zhang, T.; Gao, Y.; Zeng, J.; Lin, Q.; Mahemuti, O.; Li, Y.G.; Huo, X.D.; et al. Endophytic fungal diversity and space-time dynamics in sugar beet. Eur. J. Soil Biol. 2016, 77, 77–85. [Google Scholar] [CrossRef]
- Hoffman, M.T.; Arnold, A.E. Geographic locality and host identity shape fungal endophyte communities in cupressaceous trees. Mycol. Res. 2007, 112, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.Z.; Zhang, E.H.; Yin, X.; Cai, H.; Yuan, L.; Li, L.Q.; Zhao, K.T.; Lan, X.Z. Diversity of endophytic fungi and the rhizosphere soil fungi communities of the endangered plant Paeonia ludlowii in Tibe. J. Biol. 2020, 37, 5. [Google Scholar]
- Li, H.L.; Yu, L. Diversity of endophytic fungi in Zhanjiang honeysuckle. Guangdong Agric. Sci. 2014, 41, 5. [Google Scholar]
- Lupo, S.; Tiscornia, S.; Bettucci, L. Endophytic fungi from flowers, capsules and seeds of eucalyptus globulus. Rev. Iberoam. Micol. 2001, 18, 38–41. [Google Scholar]
- Rajeswari, S.; Umamaheswari, S.; Prasanth, D.A.; Rajamanikandan, K. Study of endophytic fungal community of Moringa oleifera from Omalur Region-Salem. Int. J. Pharm. Sci. Res. 2014, 5, 4887. [Google Scholar]
- Zeng, X.H.; Diao, H.X.; Ni, Z.Y.; Shao, L.; Jiang, K.; Hu, C.; Huang, Q.J.; Huang, W.C. Temporal Variation in Community Composition of Root Associated Endophytic Fungi and Carbon and Nitrogen Stable Isotope Abundance in Two Bletilla Species (Orchidaceae). Plants 2020, 10, 18. [Google Scholar] [CrossRef]
- Paul, N.C.; Jian, X.D.; Sang, H.K.; Choi, Y.P.; Yu, S.H. Distribution and Antifungal Activity of Endophytic Fungi in Different Growth Stages of Chili Pepper (Capsicum annuum L.) in Korea. Plant Pathol. J. 2012, 28, 10–19. [Google Scholar] [CrossRef]
- Jin, H.; Yan, Z.Q.; Liu, Q.; Yang, X.Y.; Chen, J.X.; Qin, B. Diversity and dynamics of fungal endophytes in leaves, stems and roots of Stellera chamaejasme L. in northwestern China. Antonie Van Leeuwenhoek 2013, 104, 949–963. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar ungal endophytes: Are tropical leaves biodiversity hotspots. Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Mishra, A.; Gond, S.K.; Kumar, A.; Sharma, V.K.; Verma, S.K.; Kharwar, R.N.; Sieber, T.N. Season and tissue type affect fungal endophyte communities of the Indian medicinal plant Tinospora cordifolia more strongly than geographic location. Microb. Ecol. 2012, 64, 388–398. [Google Scholar] [CrossRef]
- Lemfack, M.C.; Nickel, J.; Dunkel, M.; Preissner, R.; Piechulla, B. mVOC: A database of microbial volatiles. Nucleic Acids Res. 2014, 42, D744–D748. [Google Scholar] [CrossRef]
- Qian, C.D.; Fu, Y.H.; Jiang, F.S.; Xu, Z.H.; Cheng, D.Q.; Ding, B.; Gao, C.X.; Ding, Z.S. Lasiodiplodia sp. ME4-2, an endophytic fungus from the floral parts of Viscum coloratum, produces indole-3-carboxylic acid and other aromatic metabolites. BMC Microbiol. 2014, 14, 297. [Google Scholar] [CrossRef] [PubMed]
- Yakir, E.; Hilman, D.; Harir, Y.; Green, R.M. Regulation of output from the plant circadian clock. FEBS J. 2007, 274, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Mc-Tavish, H.S.; Davies, N.W.; Menary, R.C. Emission of Volatiles from brown boronia flowers: Some comparative observations. Ann. Bot. 2000, 86, 347–354. [Google Scholar] [CrossRef]
- Fan, J.J.; Zhang, W.X.; Zhang, D.L.; Wang, G.B.; Cao, F.L. Flowering Stage and Daytime Affect Scent Emission of Malus ioensis “Prairie Rose”. Molecules 2019, 24, 2356. [Google Scholar] [CrossRef] [PubMed]
- Kanani, M.; Chamani, E.; Shokouhian, A.A.; Torabi, G.M. Plant secondary metabolism and flower color changes in damask rose at different flowering development stages. Acta Physiol. Plant. 2021, 43, 55. [Google Scholar] [CrossRef]
- Chuang, Y.C.; Lee, M.C.; Chang, Y.L.; Chen, W.H.; Chen, H.H. Diurnal regulation of the floral scent emission by light and circadian rhythm in the Phalaenopsis orchids. Bot. Stud. 2017, 58, 50. [Google Scholar] [CrossRef] [PubMed]
- Cna’ani, A.; Muhlemann, J.K.; Ravid, J.; Masci, T.; Klempien, A.; Nguyen, T.T.H.; Dudareva, N.; Pichersky, E.; Vainstein, A. Petunia × hybrida floral scent production is negatively affected by high-temperature growth conditions. Plant Cell Environ. 2015, 38, 1333–1346. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, H.; Leng, P.; Zhao, J.; Wang, W.H.; Wang, S.D. The emission of floral scent from lilium ‘siberia’ in response to light intensity and temperature. Acta Physiol. Plant. 2013, 35, 1691–1700. [Google Scholar] [CrossRef]
- Rasulov, B.; Hüve, K.; Välbe, M.; Laisk, A.; Niinemets, Ü. Evidence that light, carbon dioxide, and oxygen dependencies of leaf isoprene emission are driven by energy status in hybrid aspen. Plant Physiol. 2009, 151, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Kawoosa, T.; Singh, H.; Kumar, A.; Sharma, S.K.; Devi, K.; Dutt, S.; Vats, S.K.; Sharma, M.; Ahuja, P.S.; Kumar, S. Light and temperature regulated terpene biosynthesis: Hepatoprotective monoterpene picroside accumulation in picrorhiza kurrooa. Funct. Integr. Genomic. 2010, 10, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.R.; Ni, D.J. Changes in Flower Aroma Compounds of Cultivars of Chimonanthus Praecox (L.) Link and at Different Stages Relative to Chimonanthus Tea Quality. Acta Hortic. Sin. 2010, 37, 1621–1628. [Google Scholar] [CrossRef]
- Li, Z.G.; Cao, H.; Lee, M.R.; De, L.S. Analysis of volatile compounds emitted from Chimonanthus praecox (L.) in different florescence and QSRR study of GC retention indices. Chromatographia 2009, 70, 1153–1162. [Google Scholar] [CrossRef]
- Li, Y.; Jia, W.J.; Wang, Q.; Wang, B.; Wang, S. Comparative analysis of floral scent profiles between two Chimonanthus praecox plants under different rhythms and blooming stages. Sci. Hortic. 2022, 301, 111129. [Google Scholar] [CrossRef]
- Yu, L. The Study of Violate Compounds and Flower Pigments in Chimonanthus praecox L. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2013. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1016190451.nh&DbName=CMFD2017 (accessed on 27 July 2022).
- Meng, L.B.; Shi, R.; Wang, Q.; Wang, S. Analysis of Floral Fragrance Compounds of Chimonanthus praecox with Different Floral Colors in Yunnan, China. Separations 2021, 8, 122. [Google Scholar] [CrossRef]
- Chen, S.Y.; Rui, R.; Wang, S.; He, X.H. Comparative Analysis of the Floral Fragrance Compounds of Panax notoginseng Flowers under the Panax notoginseng-pinus Agroforestry System Using SPME-GC-MS. Molecules 2022, 27, 3565. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.J.; Wang, S.; He, X.H.; Zhao, X.Y. Different factors drive the assembly of pine and Panax notoginseng-associated microbiomes in Panax notoginseng-pine agroforestry systems. Front. Microbiol. 2022, 13, 1018989. [Google Scholar] [CrossRef]
- Hu, A.; Nie, Y.; Yu, G.; Han, C.; He, J.; He, N.; Liu, S.; Deng, J.; Shen, W.; Zhang, G. Diurnal Temperature Variation and Plants Drive Latitudinal Patterns in Seasonal Dynamics of Soil Microbial Community. Front. Microbiol. 2019, 10, 674. [Google Scholar] [CrossRef]
- Wu, L.S. Diversity and Secondary Metabolites of Endophytic fungi from Taxus Chinensis var. mairei and Their Correlations with the Host. Ph.D. Thesis, Second Military Medical University, Shanghai, China, 2012. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CDFD1214&filename=1012404569.n (accessed on 29 November 2022).
- Zhang, P.L.; Wang, G.K.; Yu, Y.; Liu, J.S.; Wang, J.T.; Xu, F.Q.; Ma, Z.H.; Zhang, N.; Wang, G. Isolation and Identification of Endophytic Fungi from Paeonia ostii and Their Diversity in Different Growth Periods. J. Anhui Univ. Chin. Med. 2016, 35, 78–82. [Google Scholar]
- Ercole, E.; Adamo, M.; Rodda, M.; Gebauer, G.; Girlanda, M.; Perotto, S. Temporal variation in mycorrhizal diversity and carbon and nitrogen stable isotope abundance in the wintergreen meadow orchid Anacamptis morio. New Phytol. 2015, 205, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Chen, X.; Kennedy, D.W.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying community assembly processes and identifying features that impose them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef]
- Donna, J.L.; Juliette, T.J.; Colleen, J.D. Only a matter of time: The impact of daily and seasonal rhythms on phytochemicals. Phytochem. Rev. Fundam. Perspect. Nat. Prod. Res. 2019, 18, 1409–1433. [Google Scholar]
- Dudareva, N.; Gershenzon, P.J. Biochemistry of Plant Volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Liu, Z.P.; Sun, L.Y.; Zhang, K.X.; Xu, J.; Wen, D.; Zhao, R.; Liu, X.B. Endophytic Fungi Diversity of Ginseng Seeds by Using 454 GS FLX. Inf. Tradit. Chin. Med. 2017, 34, 28–32. [Google Scholar]
- Yang, L.C.; Wu, Q.S.; Gao, Y.; Chen, Q.Q.; Wang, Y.Q.; Niu, X.J.; Weng, Q.B. Analysis of Composition of Endophytic Community of Dendrobium nobile Lindl. Seeds Based on High-throughput Sequencing. Seed 2020, 39, 5. [Google Scholar]
- Yang, J.; Dong, C.B.; Chen, W.H.; Liang, J.D.; Han, Y.F.; Liang, Z.Q. Community composition and ecological functional structural analysis of endophytic fungi in bark of Eucommia ulmoides in different areas. China J. Chin. Mater. Medica 2019, 6, 1126–1134. [Google Scholar]
- Zhang, S.P.; Xu, T.; Yang, L.Q.; Song, Y.; Li, X.Y.; Zhang, H.W. Endophytic fungal communities of Stipa sp. roots in different types of steppes in northern China. Chin. J. Appl. Ecol. 2014, 12, 3475–3482. [Google Scholar]
- Yang, Q.; Zhou, K.; Chen, A.J.; Guo, X.; Liu, X.R.; Liang, Y.L. Endophytic fungal communities structure and variation in Ginkgo biloba leaf tissues with different ages. J. Hunan Agric. Univ. (Nat. Sci.) 2019, 45, 42–49. [Google Scholar]
- Serra, S.; De Simeis, D. Fungi-mediated biotransformation of the isomeric forms of the apocarotenoidsionone, damascone and theaspirane. Molecules 2019, 24, 19. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.H.; Yuan, M.Q.; Ao, X.; Ren, A.Y.; Zhang, H.B.; Yang, M.Z. Endophytic fungi specifically introduce novel metabolites into grape flesh cells in vitro. PLoS ONE 2018, 13, e0196996. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.F. Diversity of Endophytic Fung in Five Flower Plants in Yunnan and Their Effects on Plant Growth. Master’s Thesis, Kunming University of Science and Technology, Kunming, China, 2013. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1016037043.nh&DbName=CMFD2016 (accessed on 24 July 2022).
- Huang, Y. The Study of the Didymellaceae and Plectosphaerella in Endophytic fungi of Aquatic Plants. Master’s Thesis, Yunnan University, Kunming, China, 2018. [Google Scholar] [CrossRef]
- Xu, Z.L.; Yan, D.J.; Tan, X.M.; Niu, S.B.; Yu, M.; Sun, B.D.; Ding, C.F.; Zhang, Y.G.; Ding, G. Phaeosphspirone (1/1′), a pair of unique polyketide enantiomers with an unusual 6/5/5/6 tetracyclic ring from the desert plant endophytic fungus Phaeosphaeriaceae sp. Phytochemistry 2022, 194, 112969. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.Q.; Chen, W.F.; Fan, Y.K.; Zhu, Z.D.; Ma, G.X.; Chen, D.L.; Tian, R.R. Research progress on secondary metabolites and activities of endophytic fungi of genuses Aspergillus and Trichoderma from mangroves. J. Trop. Oceanogr. 2023, 42, 12–24. Available online: http://kns.cnki.net/kcms/detail/44.1500.P.20221208.1416.002.html (accessed on 13 June 2023).
- Zhang, J. Research on Diversity of Endosymbiotic Fungi in Roots of Cymbidium. Master’s Thesis, Chinese Academy of Forestry, Beijing, China, 2012. Available online: https://kns.cnki.net/kcms/detail/detail.aspx?FileName=1012423572.nh&DbName=CMFD2012 (accessed on 29 June 2022).
- Rai, M.; Agarkar, G. Plant-fungal interactions: What triggers the fungi to switch among lifestyles. Crit. Rev. Microbiol. 2016, 42, 428–438. [Google Scholar] [CrossRef]
- Huang, W.Y.; Cai, Y.Z.; Hyde, K.D.; Corke, H.; Sun, M. Endophytic fungi from Nerium oleander L (Apocynaceae): Main constituents and antioxidant activity. World J. Microbiol. Biotechnol. 2007, 23, 1253–1263. [Google Scholar] [CrossRef]
- Cai, J.; Zhou, W.Y.; Xu, G.M.; Zhou, X.J. Research progress on natural products of fungi of Diaporthe sp. China J. Chin. Mater. Medica 2021, 46, 1717–1726. [Google Scholar] [CrossRef]
- Cui, Y.F.; Huang, Y.; Jiang, L.H. Research Progress on Gibberella Species of Agricultural Importance. Chin. Agric. Sci. Bull. 2007, 7, 441–446. [Google Scholar]
- Li, H.K. Correlation between the Diversity of Endophytic Fungi and Taxoids Accumulation in Taxus cuspidata and the Application of the High-Yield Strain. Master’s Thesis, Northeast Forestry University, Heilongjiang, China, 2021. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxonomy Levels | Top Five | GUA | GUB | GUC | GOA | GOB | GOC |
|---|---|---|---|---|---|---|---|
| Phylum | Ascomycota | 96.57% | 90.84% | 87.70% | 98.40% | 85.16% | 82.32% |
| Basidiomycota | 2.88% | 7.03% | 8.85% | 1.55% | 13.60% | 13.60% | |
| unclassified_k__Fungi | 0.29% | 1.95% | 3.07% | 0.04% | 0.86% | 3.31% | |
| Class | Dothideomycetes | 46.20% | 37.16% | 44.57% | 44.03% | 36.64% | 19.90% |
| unclassified_p__Ascomycota | 35.23% | 20.61% | 7.43% | 50.52% | 26.31% | 37.55% | |
| Eurotiomycetes | 8.30% | 7.34% | 8.39% | 2.41% | 8.81% | 4.32% | |
| Sordariomycetes | 5.73% | 16.97% | 4.06% | 0.89% | 1.30% | 14.92% | |
| Tremellomycetes | 1.94% | 5.66% | 6.93% | 1.00% | 10.83% | 11.45% | |
| Order | Pleosporales | 33.37% | 31.01% | 18.37% | 40.96% | 50.53% | 30.30% |
| unclassified_p__Ascomycota | 46.20% | 20.61% | 41.66% | 7.43% | 41.98% | 26.13% | |
| Eurotiales | 7.81% | 4.45% | 16.93% | 5.94% | 1.30% | 7.42% | |
| Hypocreales | 4.51% | 16.82% | 1.57% | 1.99% | 0.49% | 0.29% | |
| Trichosporonales | 1.73% | 5.49% | 1.63% | 5.06% | 0.73% | 9.67% | |
| Family | unclassified_p__Ascomycota | 46.20% | 20.61% | 7.43% | 50.52% | 26.13% | 37.55% |
| Didymellaceae | 18.86% | 23.55% | 25.44% | 28.41% | 19.57% | 9.13% | |
| Phaeosphaeriaceae | 10.68% | 6.00% | 14.21% | 4.20% | 7.45% | 4.29% | |
| Aspergillaceae | 7.80% | 4.45% | 5.94% | 1.29% | 7.20% | 2.45% | |
| Nectriaceae | 4.49% | 16.46% | 1.62% | 0.47% | 1.63% | 13.67% | |
| Genus | unclassified_p__Ascomycota | 46.20% | 20.61% | 24.58% | 50.52% | 26.13% | 37.55% |
| Didymella | 17.39% | 19.57% | 7.43% | 26.91% | 14.77% | 8.70% | |
| unclassified_f__Phaeosphaeriaceae | 9.88% | 4.61% | 12.15% | 2.84% | 4.05% | 3.57% | |
| Aspergillus | 7.29% | 5.49% | 5.06% | 1.19% | 6.32% | 1.63% | |
| Gibberalla | 3.86% | 13.87% | 1.28% | 0.73% | 1.63% | 13.28% | |
| Species | unclassified_p__Ascomycota | 46.20% | 20.61% | 24.58% | 50.52% | 26.13% | 37.56% |
| unclassified_g__Didymella | 17.39% | 19.56% | 7.43% | 26.91% | 14.77% | 8.70% | |
| unclassified_f__Phaeosphaeriaceae | 9.88% | 4.61% | 12.15% | 2.84% | 4.05% | 3.57% | |
| Aspergillus amstelodami | 6.49% | 2.08% | 2.12% | 0.84% | 1.41% | 10.71% | |
| unclassified_g__Gibberella | 3.61% | 13.13% | 1.05% | 8.99% | 9.67% | 13.28% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Hei, J.; He, X.; Rui, R.; Wang, S. The Relationship between Endophytic Fungi of Chimonanthus praecox and Volatile Metabolites under Different Circadian Rhythms and Blooming Stages. J. Fungi 2024, 10, 145. https://doi.org/10.3390/jof10020145
Li Y, Hei J, He X, Rui R, Wang S. The Relationship between Endophytic Fungi of Chimonanthus praecox and Volatile Metabolites under Different Circadian Rhythms and Blooming Stages. Journal of Fungi. 2024; 10(2):145. https://doi.org/10.3390/jof10020145
Chicago/Turabian StyleLi, Yue, Jingying Hei, Xiahong He, Rui Rui, and Shu Wang. 2024. "The Relationship between Endophytic Fungi of Chimonanthus praecox and Volatile Metabolites under Different Circadian Rhythms and Blooming Stages" Journal of Fungi 10, no. 2: 145. https://doi.org/10.3390/jof10020145