
The Identification of a Target Gene of the Transcription Factor KojR and Elucidation of Its Role in Carbon Metabolism for Kojic Acid Biosynthesis in Aspergillus oryzae

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. A. oryzae Strains, Growth Conditions, and Transformation
2.2. The Construction of a KojR FL/BD Expression Vector
2.3. The Expression of MBP-Fused KojR Using a Cell-Free Protein Synthesis System
2.4. The In Vitro Selection of KojR-Bound DNA Fragments from an A. oryzae Genomic Library Using gSELEX-Seq
2.5. DNA Sequencing and Data Analysis in gSELEX-Seq
2.6. The Construction of an A. oryzae ΔkojR Strain Using CRISPR/Cas9
2.7. The Identifications of DEGs Associated with KojR Expression Using RNA-Seq
2.8. The KEGG Pathway Analysis of DEGs
3. Results
3.1. The Preparation of MBP-Fused KojR for Use in gSELEX
3.2. The Genome-Wide Identification of KojR Binding Sites Using gSELEX-Seq
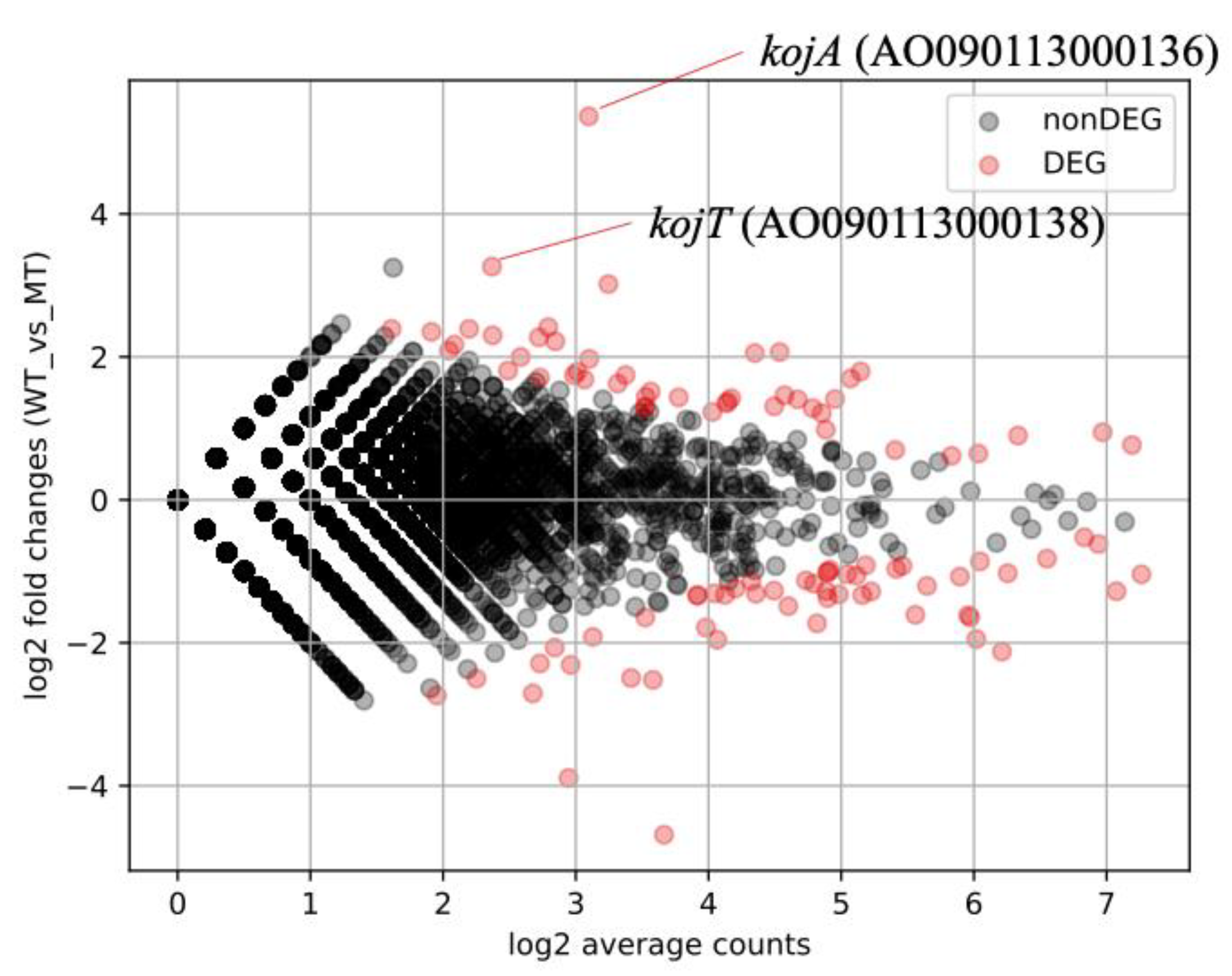
3.3. The Identification of KojR Expression-Dependent DEGs Using RNA-Seq
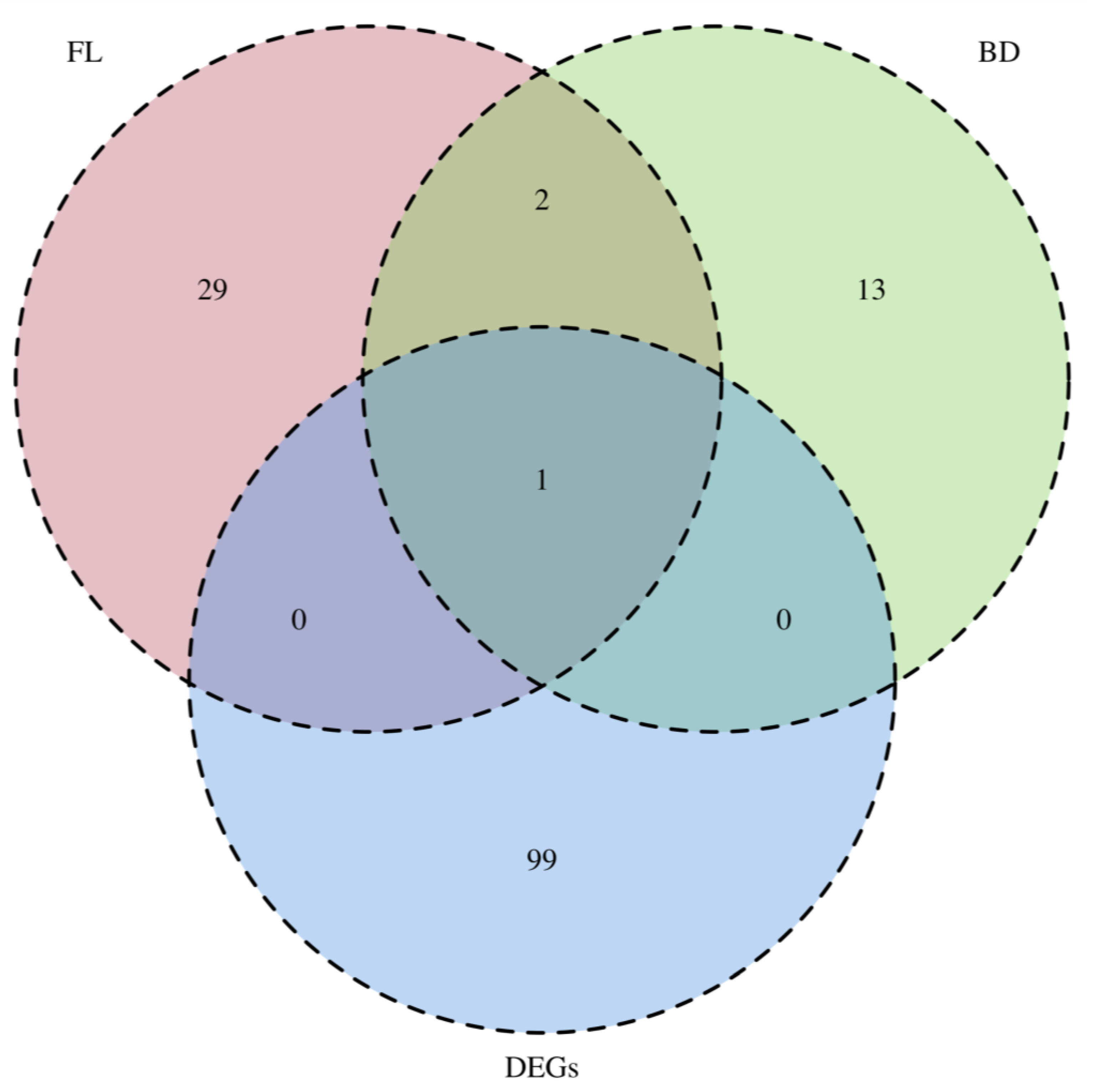
3.4. The Comprehensive Identification of the Target Genes of KojR Using Integrated Analysis-Based Binding Sites on A. oryzae Genome and DEGs Information
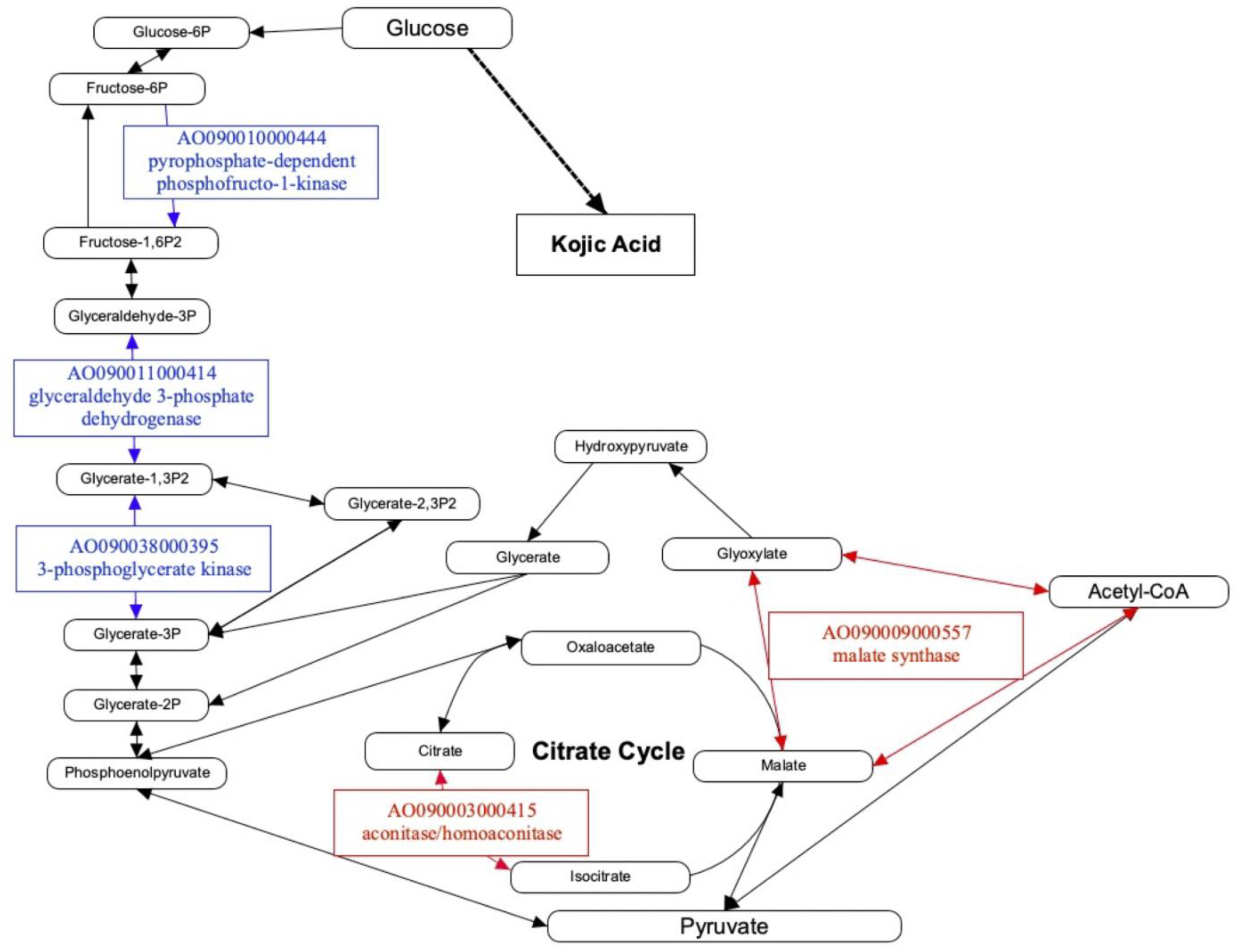
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saeedi, M.; Eslamifar, M.; Khezri, K. Kojic Acid Applications in Cosmetic and Pharmaceutical Preparations. Biomed. Pharmacother. 2019, 110, 582–593. [Google Scholar] [CrossRef]
- Brtko, J. Biological Functions of Kojic Acid and Its Derivatives in Medicine, Cosmetics, and Food Industry: Insights into Health Aspects. Arch. Pharm. 2022, 355, 2200215. [Google Scholar] [CrossRef]
- Bentley, R. From Miso, Sake and Shoyu to Cosmetics: A Century of Science for Kojic Acid. ChemInform 2007, 38, 18. [Google Scholar] [CrossRef]
- Chib, S.; Jamwal, V.L.; Kumar, V.; Gandhi, S.G.; Saran, S. Fungal Production of Kojic Acid and Its Industrial Applications. Appl. Microbiol. Biotechnol. 2023, 107, 2111–2130. [Google Scholar] [CrossRef]
- Terabayashi, Y.; Sano, M.; Yamane, N.; Marui, J.; Tamano, K.; Sagara, J.; Dohmoto, M.; Oda, K.; Ohshima, E.; Tachibana, K.; et al. Identification and Characterization of Genes Responsible for Biosynthesis of Kojic Acid, an Industrially Important Compound from Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 953–961. [Google Scholar] [CrossRef]
- Marui, J.; Yamane, N.; Ohashi-Kunihiro, S.; Ando, T.; Terabayashi, Y.; Sano, M.; Ohashi, S.; Ohshima, E.; Tachibana, K.; Higa, Y.; et al. Kojic Acid Biosynthesis in Aspergillus oryzae Is Regulated by a Zn(II)2Cys6 Transcriptional Activator and Induced by Kojic Acid at the Transcriptional Level. J. Biosci. Bioeng. 2011, 112, 40–43. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Zeng, B.; Zhang, Z. Construction of Single, Double, or Triple Mutants within Kojic Acid Synthesis Genes kojA, kojR, and kojT by the CRISPR/Cas9 Tool in Aspergillus oryzae. Folia Microbiol. 2022, 67, 459–468. [Google Scholar] [CrossRef]
- Chang, P.-K.; Scharfenstein, L.L.; Mahoney, N.; Kong, Q. Kojic Acid Gene Clusters and the Transcriptional Activation Mechanism of Aspergillus flavus KojR on Expression of Clustered Genes. J. Fungi 2023, 9, 259. [Google Scholar] [CrossRef]
- De Castro, P.A.; Colabardini, A.C.; Moraes, M.; Horta, M.A.C.; Knowles, S.L.; Raja, H.A.; Oberlies, N.H.; Koyama, Y.; Ogawa, M.; Gomi, K.; et al. Regulation of Gliotoxin Biosynthesis and Protection in Aspergillus Species. PLoS Genet. 2022, 18, e1009965. [Google Scholar] [CrossRef]
- Kojima, T.; Kunitake, E.; Ihara, K.; Kobayashi, T.; Nakano, H. A Robust Analytical Pipeline for Genome-Wide Identification of the Genes Regulated by a Transcription Factor: Combinatorial Analysis Performed Using gSELEX-Seq and RNA-Seq. PLoS ONE 2016, 11, e0159011. [Google Scholar] [CrossRef]
- Oka, H.; Kojima, T.; Ihara, K.; Kobayashi, T.; Nakano, H. Comprehensive Investigation of the Gene Expression System Regulated by an Aspergillus oryzae Transcription Factor XlnR Using Integrated Mining of gSELEX-Seq and Microarray Data. BMC Genom. 2019, 20, 16. [Google Scholar] [CrossRef]
- Oka, H.; Kojima, T.; Kato, R.; Ihara, K.; Nakano, H. Construction of Transcript Regulation Mechanism Prediction Models Based on Binding Motif Environment of Transcription Factor AoXlnR in Aspergillus oryzae. bioRxiv 2021. [Google Scholar] [CrossRef]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.-I.; Arima, T.; Akita, O.; Kashiwagi, Y.; et al. Genome Sequencing and Analysis of Aspergillus oryzae. Nature 2005, 438, 1157–1161. [Google Scholar] [CrossRef]
- Okegawa, Y.; Motohashi, K. A Simple and Ultra-Low Cost Homemade Seamless Ligation Cloning Extract (SLiCE) as an Alternative to a Commercially Available Seamless DNA Cloning Kit. Biochem. Biophys. Rep. 2015, 4, 148–151. [Google Scholar] [CrossRef]
- Tani, S.; Itoh, T.; Kato, M.; Kobayashi, T.; Tsukagoshi, N. In vivo and in vitro analyses of the AmyR binding site of the Aspergillus nidulans agdA promoter; requirement of the CGG direct repeat for induction and high affinity binding of AmyR. Biosci. Biotechnol. Biochem. 2001, 65, 1568–1574. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Katayama, T.; Nakamura, H.; Zhang, Y.; Pascal, A.; Fujii, W.; Maruyama, J. Forced Recycling of an AMA1-Based Genome-Editing Plasmid Allows for Efficient Multiple Gene Deletion/Integration in the Industrial Filamentous Fungus Aspergillus oryzae. Appl. Environ. Microbiol. 2019, 85, e01896-18. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for Inferring Cellular Functions from Protein Sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef]
- Kutmon, M.; Van Iersel, M.P.; Bohler, A.; Kelder, T.; Nunes, N.; Pico, A.R.; Evelo, C.T. PathVisio 3: An Extendable Pathway Analysis Toolbox. PLoS Comput. Biol. 2015, 11, e1004085. [Google Scholar] [CrossRef]
- Tamano, K.; Kuninaga, M.; Kojima, N.; Umemura, M.; Machida, M.; Koike, H. Use of the kojA Promoter, Involved in Kojic Acid Biosynthesis, for Polyketide Production in Aspergillus oryzae: Implications for Long-Term Production. BMC Biotechnol. 2019, 19, 70. [Google Scholar] [CrossRef]
- Arakawa, G.; Kudo, H.; Yanase, A.; Eguchi, Y.; Kodama, H.; Ogawa, M.; Koyama, Y.; Shindo, H.; Hosaka, M.; Tokuoka, M. A Unique Zn(II)2-Cys6-Type Protein, KpeA, Is Involved in Secondary Metabolism and Conidiation in Aspergillus oryzae. Fungal Genet. Biol. 2019, 127, 35–44. [Google Scholar] [CrossRef]
- Sano, M. Aspergillus oryzae nrtA Affects Kojic Acid Production. Biosci. Biotechnol. Biochem. 2016, 80, 1776–1780. [Google Scholar] [CrossRef]
- Oda, K.; Kobayashi, A.; Ohashi, S.; Sano, M. Aspergillus oryzae laeA Regulates Kojic Acid Synthesis Genes. Biosci. Biotechnol. Biochem. 2011, 75, 1832–1834. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Zeng, B.; Zhang, Z. Identification and Characterization of a Novel Gene Aokap1 Involved in Growth and Kojic Acid Synthesis in Aspergillus oryzae. Arch. Microbiol. 2022, 204, 67. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Xiao, Y.; Jie, J.; Zeng, B.; Zhang, Z. Overexpression of a Novel Gene Aokap2 Affects the Growth and Kojic Acid Production in Aspergillus oryzae. Mol. Biol. Rep. 2022, 49, 2745–2754. [Google Scholar] [CrossRef]
- Chen, T.; Chen, Z.; Li, Y.; Zeng, B.; Zhang, Z. A Novel Major Facilitator Superfamily Transporter Gene Aokap4 near the Kojic Acid Gene Cluster Is Involved in Growth and Kojic Acid Production in Aspergillus oryzae. J. Fungi 2022, 8, 885. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; Zhang, F.; Chen, T.; Fan, J.; Deng, X.; Lei, X.; Zeng, B.; Zhang, Z. The C2H2-Type Zinc-Finger Regulator AoKap5 Is Required for the Growth and Kojic Acid Synthesis in Aspergillus oryzae. Fungal Genet. Biol. 2023, 167, 103813. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, T.; Wang, H.; Jiang, C.; Liu, Y.; Wu, X.; Li, Y.; Zeng, B.; Zhang, Z. Disruption of Aokap6 near the Kojic Acid Gene Cluster Affects the Growth and Kojic Acid Production in Aspergillus oryzae. World J. Microbiol. Biotechnol. 2022, 38, 175. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, J.; Long, C.; He, B.; Hu, Z.; Jiang, C.; Li, Y.; Ma, L.; Wen, J.; Zou, X.; et al. Identification and Characterization of the ZRT, IRT-like Protein (ZIP) Family Genes Reveal Their Involvement in Growth and Kojic Acid Production in Aspergillus oryzae. J. Ind. Microbiol. Biotechnol. 2019, 46, 1769–1780. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, T.; Zhang, H.; Li, Y.; Fan, J.; Yao, L.; Zeng, B.; Zhang, Z. Functional Role of a Novel Zinc Finger Protein, AoZFA, in Growth and Kojic Acid Synthesis in Aspergillus oryzae. Appl. Environ. Microbiol. 2023, 89, e00909-23. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Region | Gene ID | Original Description |
|---|---|---|
| Chr5_A_oryzae_RIB40:4372480-4372692 | AO090113000138 | Putative transporter; present in the kojic acid biosynthetic gene cluster |
| Chr6_A_oryzae_RIB40:3584694-3584906 | AO090038000029 | Has domain(s) with predicted peroxiredoxin activity and role in oxidation-reduction process |
| Chr5_A_oryzae_RIB40:1165262-1165474 | AO090701000448 | Has domain(s) with predicted heme binding activity |
| Chr5_A_oryzae_RIB40:643370-643582 | AO090701000654 | Protein of unknown function |
| Chr5_A_oryzae_RIB40:1158260-1158472 | AO090701000450 | Has domain(s) with predicted catalytic activity, hydrolase activity, and role in metabolic process |
| Chr1_A_oryzae_RIB40:5569621-5569833 | AO090005000336 | Has domain(s) with predicted sequence-specific DNA binding RNA polymerase II transcription factor activity, zinc ion binding activity, and role in regulation of transcription, DNA-templated and nucleus localization |
| Chr7_A_oryzae_RIB40:229700-229912 | AO090011000078 | Ortholog of A. niger CBS 513.88: An01g06430, A. versicolor: Aspve1_0082366, A. niger ATCC 1015: 36036-mRNA, and A. zonatus: Aspzo1_0026572 |
| Chr1_A_oryzae_RIB40:4941497-4941709 | AO090005000571 | Has domain(s) with predicted NAD binding, oxidoreductase activity, activity on the CH-OH group of donors, NAD or NADP as acceptor activity, and role in oxidation-reduction process |
| Chr2_A_oryzae_RIB40:4505321-4505533 | AO090003000890 | Protein of unknown function |
| Chr4_A_oryzae_RIB40:4149411-4149623 | AO090102000151 | Ortholog(s) have ubiquitin–protein transferase activity, have a role in protein import into peroxisome matrix and are an integral component of peroxisomal membrane localization |
| Chr1_A_oryzae_RIB40:6476509-6476721 | AO090308000007 | Protein of unknown function |
| Chr3_A_oryzae_RIB40:391752-391964 | AO090023000155 | Protein of unknown function |
| Chr6_A_oryzae_RIB40:3081726-3081938 | AO090038000221 | Protein of unknown function |
| Chr7_A_oryzae_RIB40:1923126-1923338 | AO090011000751 | Protein of unknown function |
| Chr2_A_oryzae_RIB40:5473553-5473765 | AO090003001242 | Has domain(s) with predicted ATP binding and inositol pentakisphosphate 2-kinase activity |
| Chr6_A_oryzae_RIB40:1409858-1410070 | AO090020000161 | Ortholog of A. nidulans FGSC A4: AN6458, A. fumigatus Af293: Afu3g07420, A. niger CBS 513.88: An02g10960, An01g01420, An12g05390, and A. oryzae RIB40: AO090005000921, AO090003001427 |
| Chr6_A_oryzae_RIB40:1409858-1410070 | AO090020000162 | Has domain(s) with predicted UDP-N-acetylmuramate dehydrogenase activity, flavin adenine dinucleotide binding, oxidoreductase activity, and activity on the CH-OH group of donors |
| Chr6_A_oryzae_RIB40:4172129-4172341 | AO090138000010 | Has domain(s) with predicted 2-dehydropantoate 2-reductase activity, NADP binding, coenzyme binding, oxidoreductase activity, activity on the CH-OH group of donors, and NAD or NADP as acceptor activity |
| Chr5_A_oryzae_RIB40:1114034-1114246 | AO090701000470 | Ortholog of A. fumigatus Af293: Afu2g16985, A. wentii: Aspwe1_0171101, A. clavatus NRRL 1: ACLA_075940, and A. zonatus: Aspzo1_0015705 |
| Chr3_A_oryzae_RIB40:1794701-1794913 | AO090023000683 | Protein of unknown function |
| Chr6_A_oryzae_RIB40:3558859-3559071 | AO090038000040 | Has domain(s) with predicted catalytic activity, catechol 1,2-dioxygenase activity, ferric iron binding, iron ion binding and oxidoreductase activity, in addition to other activities |
| Chr1_A_oryzae_RIB40:3825460-3825672 | AO090005000971 | Has domain(s) with a predicted role in the biosynthetic process |
| Chr5_A_oryzae_RIB40:4076380-4076592 | AO090113000012 | Ortholog of A. nidulans FGSC A4: AN6909/BEST2, A. niger CBS 513.88: An14g05100, A. wentii: Aspwe1_0153378, A. sydowii: Aspsy1_0054771, and A. terreus NIH2624: ATET_06151 |
| Chr7_A_oryzae_RIB40:2355701-2355913 | AO090011000905 | Ortholog(s) have sequence-specific DNA binding transcription factor activity |
| Chr7_A_oryzae_RIB40:2498610-2498822 | AO090011000954 | Ortholog(s) have cytosol localization |
| Chr8_A_oryzae_RIB40:1654282-1654494 | AO090010000667 | Has domain(s) with predicted iron ion binding and oxidoreductase activity; role in fatty acid biosynthetic process and oxidation-reduction process |
| Chr3_A_oryzae_RIB40:244139-244351 | AO090023000096 | Has domain(s) with predicted catalytic activity, coenzyme binding activity, and role in cellular metabolic process |
| Chr8_A_oryzae_RIB40:1372821-1373033 | AO090010000775 | Ortholog(s) have UDP-N-acetylglucosamine transmembrane transporter activity and role in UDP-N-acetylglucosamine transport, UDP-glucose transport, fungal-type cell wall chitin biosynthetic process, and transmembrane transport |
| Chr1_A_oryzae_RIB40:3919266-3919478 | AO090005000937 | Ortholog of A. nidulans FGSC A4: AN1323 and A. flavus NRRL 3357: AFL2T_00913 |
| Chr6_A_oryzae_RIB40:442735-442947 | AO090020000539 | Ortholog(s) have Golgi apparatus, endoplasmic reticulum localization |
| Chr8_A_oryzae_RIB40:2804269-2804481 | AO090010000224 | Ortholog(s) have role in ethanol metabolic process and mitochondrial inner membrane localization |
| Chr8_A_oryzae_RIB40:2804269-2804481 | AO090010000223 | 40S ribosomal protein S2-like protein; predominantly expressed in the hyphal tip region |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizutani, T.; Oka, H.; Goto, R.; Tsurigami, R.; Maruyama, J.-i.; Shimizu, M.; Kato, M.; Nakano, H.; Kojima, T. The Identification of a Target Gene of the Transcription Factor KojR and Elucidation of Its Role in Carbon Metabolism for Kojic Acid Biosynthesis in Aspergillus oryzae. J. Fungi 2024, 10, 113. https://doi.org/10.3390/jof10020113
Mizutani T, Oka H, Goto R, Tsurigami R, Maruyama J-i, Shimizu M, Kato M, Nakano H, Kojima T. The Identification of a Target Gene of the Transcription Factor KojR and Elucidation of Its Role in Carbon Metabolism for Kojic Acid Biosynthesis in Aspergillus oryzae. Journal of Fungi. 2024; 10(2):113. https://doi.org/10.3390/jof10020113
Chicago/Turabian StyleMizutani, Tomoka, Hiroya Oka, Riko Goto, Ryoga Tsurigami, Jun-ichi Maruyama, Motoyuki Shimizu, Masashi Kato, Hideo Nakano, and Takaaki Kojima. 2024. "The Identification of a Target Gene of the Transcription Factor KojR and Elucidation of Its Role in Carbon Metabolism for Kojic Acid Biosynthesis in Aspergillus oryzae" Journal of Fungi 10, no. 2: 113. https://doi.org/10.3390/jof10020113