
Aspects of Reproductive Biology of the European Hake (Merluccius merluccius) in the Northern and Central Adriatic Sea (GSA 17-Central Mediterranean Sea)


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Reproductive Seasonality and Fish Condition Indices
2.3. Histological Analysis
2.4. Size at First Maturity (L50) and Fecundity
2.5. Statistical Analysis
3. Results
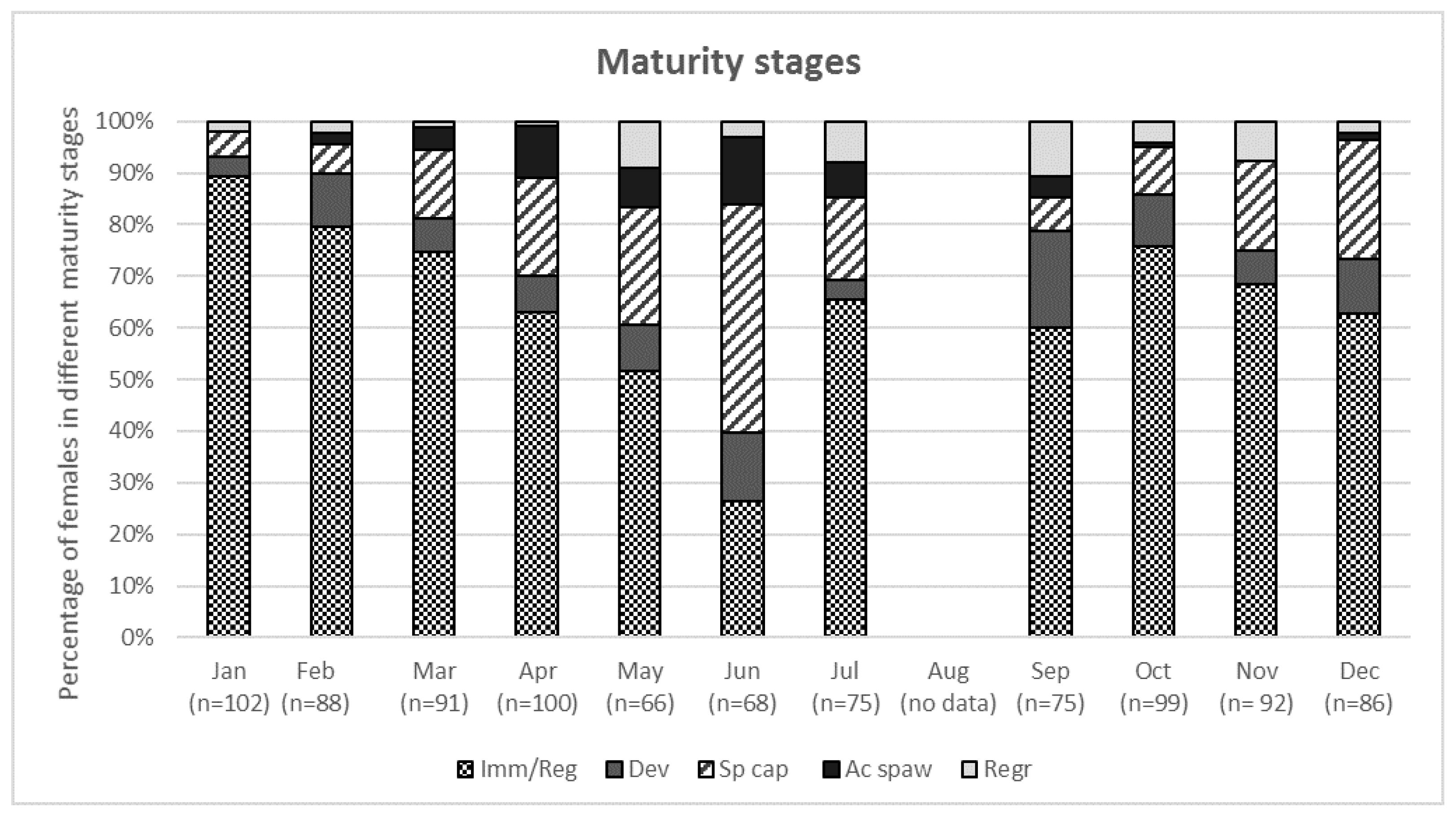
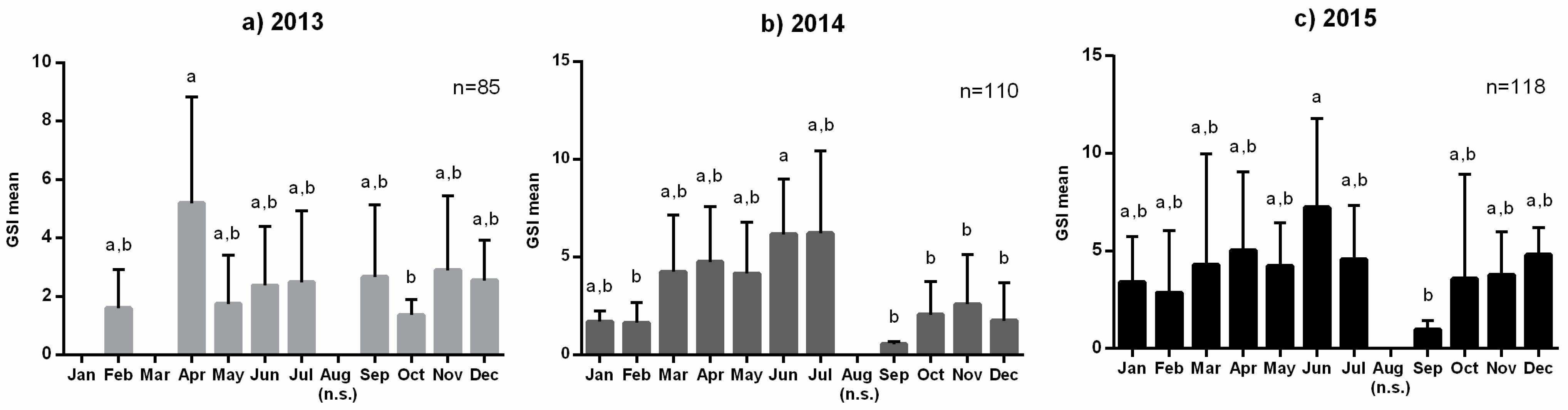
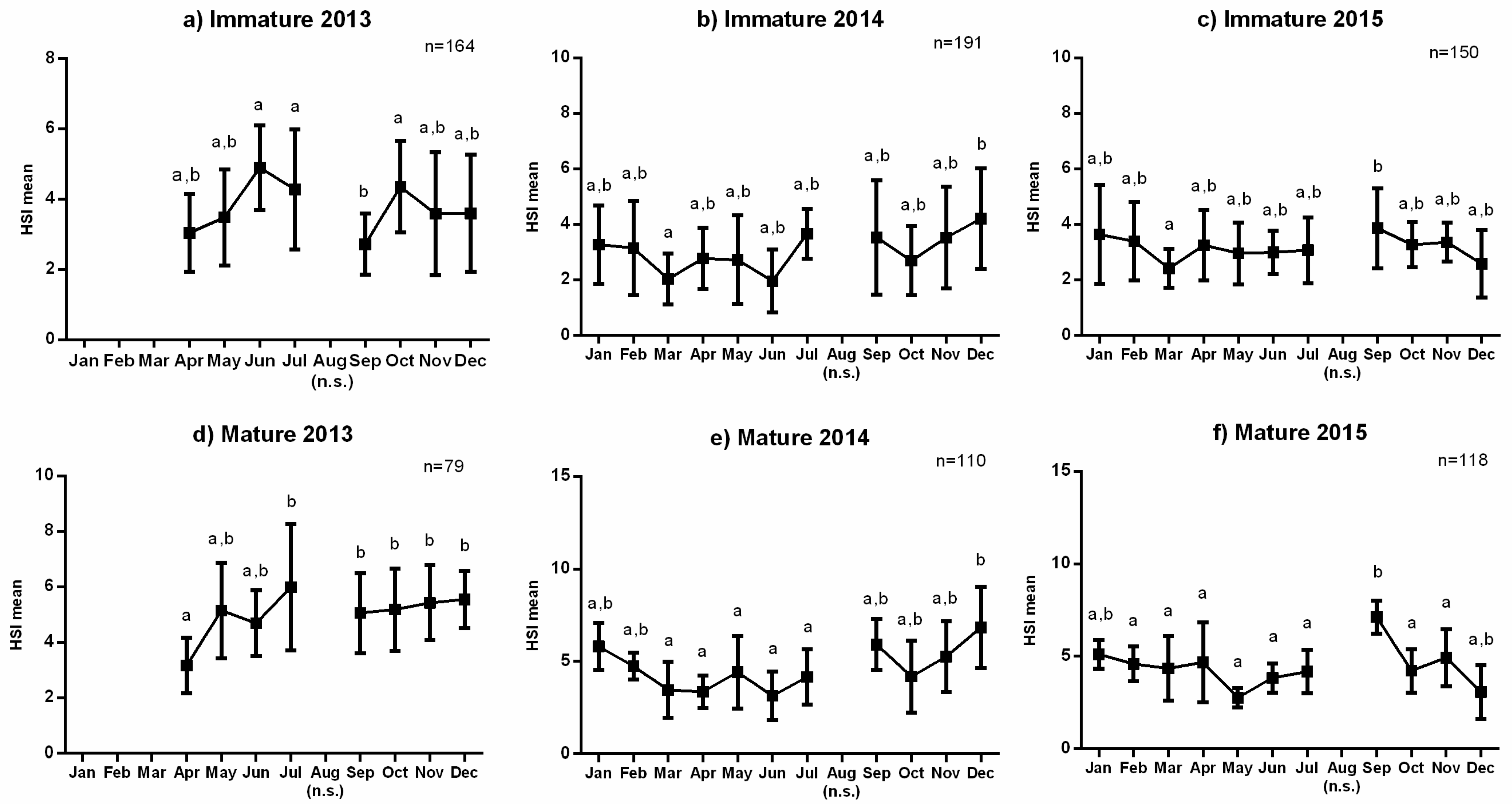
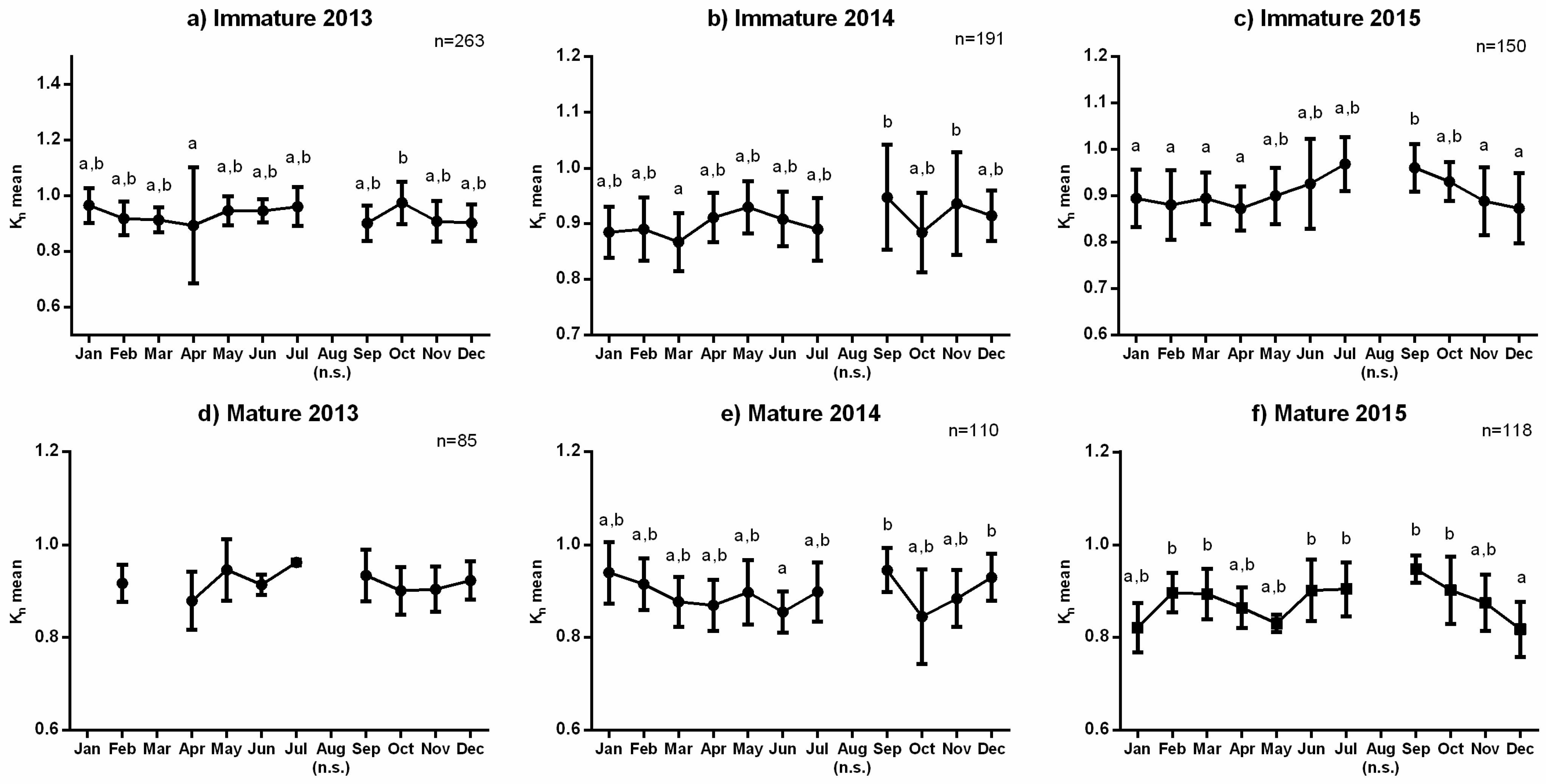
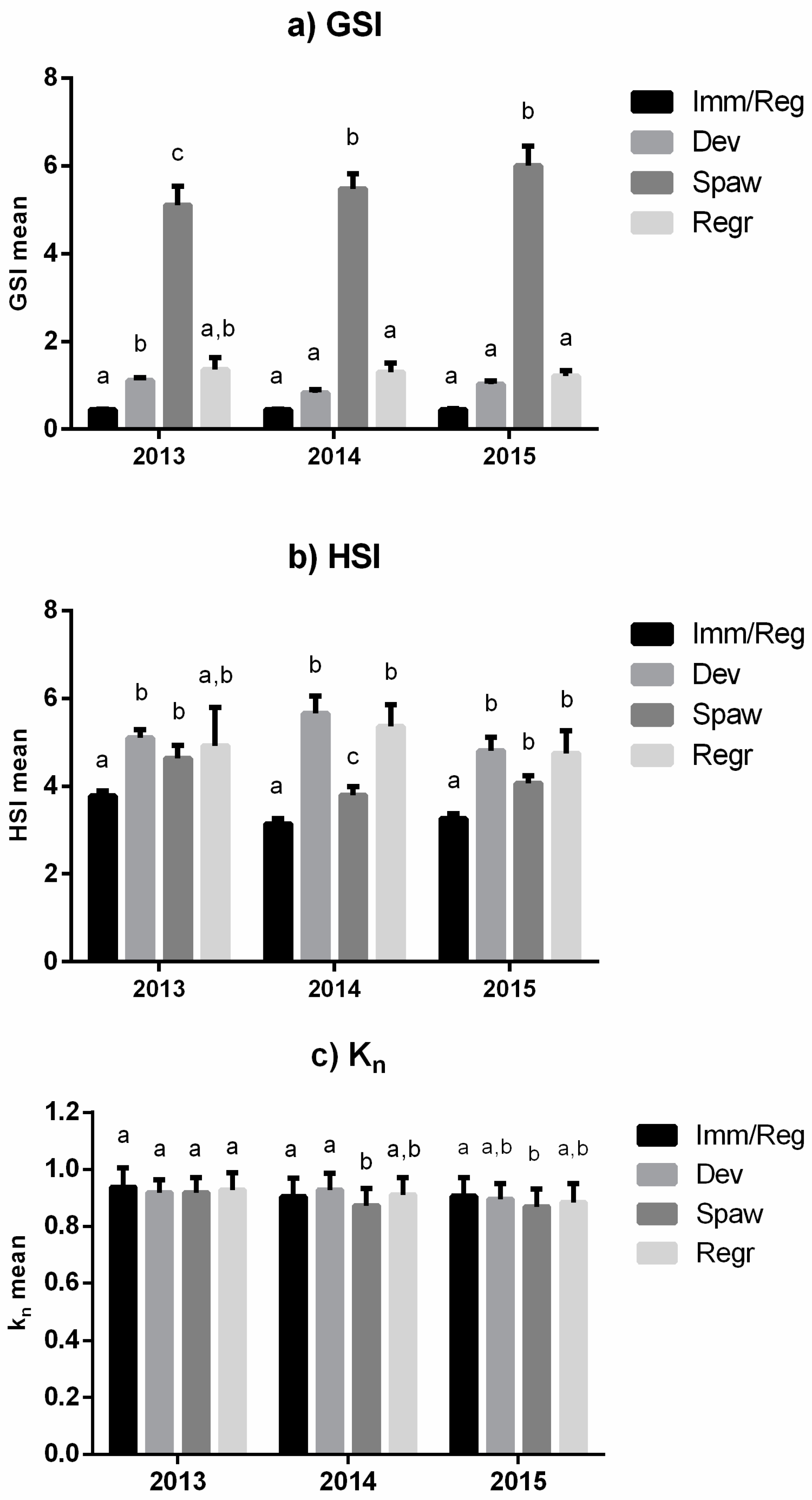
3.1. Reproductive Seasonality and Fish Condition
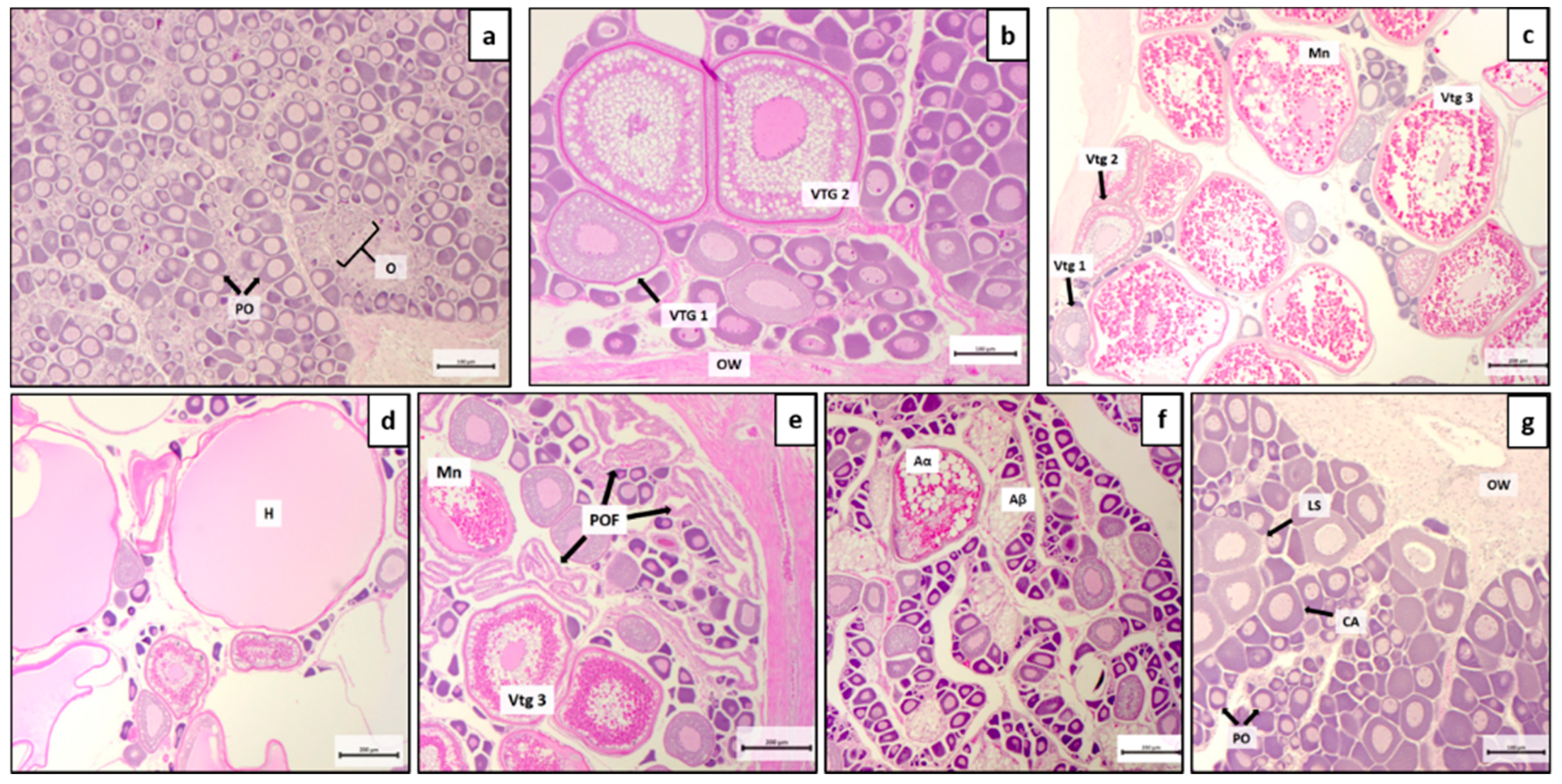
3.2. Histological Analysis: Ovarian Classification and Patterns
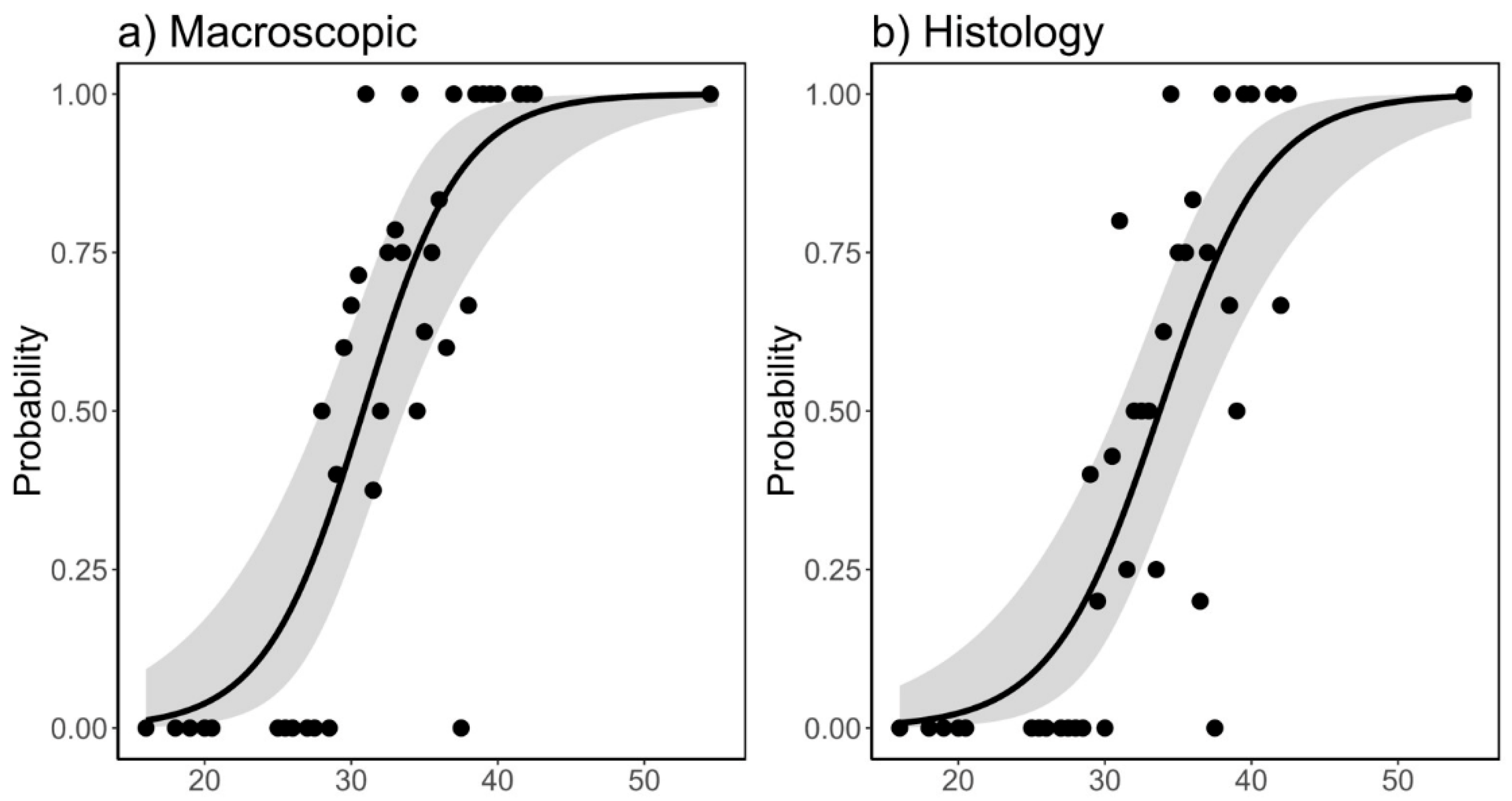
3.3. Size at First Maturity (L50) and Fecundity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Review of the state of world marine fishery resources. FAO Fish. Aquac. Tech. Pap. 2011, 569, 334. [Google Scholar]
- Colloca, F.; Cardinale, M.; Maynou, F.; Giannoulaki, M.; Scarcella, G.; Jenko, K.; Bellido, J.M.; Fiorentino, F. Rebuilding Mediterranean fisheries: A new paradigm for ecological sustainability. Fish Fish. 2013, 14, 89–109. [Google Scholar] [CrossRef]
- Tsikliras, A.C.; Dinouli, A.; Tsiros, V.-Z.; Tsalkou, E. The Mediterranean and Black Sea Fisheries at Risk from Overexploitation. PLoS ONE 2015, 10, e0121188. [Google Scholar] [CrossRef] [Green Version]
- Mullon, C.; Freon, P.; Cury, P. The dynamics of collapse in world fisheries. Fish Fish. 2005, 6, 111–120. [Google Scholar] [CrossRef]
- Colloca, F.; Scarcella, G.; Libralato, S. Recent trends and impacts of fisheries exploitation on Mediterranean stocks and ecosystems. Front. Mar. Sci. 2017, 4, 244. [Google Scholar] [CrossRef] [Green Version]
- Druon, J.N.; Fiorentino, F.; Murenu, M.; Knittweis, L.; Colloca, F.; Osio, C.; Mérigot, B.H.; Garofalo, G.; Mannini, A.; Jadaud, A.H.; et al. Modelling of European hake nurseries in the Mediterranean Sea: An ecological niche approach. Prog. Oceanogr. 2015, 130, 188–204. [Google Scholar] [CrossRef]
- Cerviño, S.; Domínguez-Petit, R.; Jardim, E.; Mehault, S.; Piñeiro, C.; Saborido-Rey, F. Impact of egg production and stock structure on MSY reference points and its management implications for southern hake (Merluccius merluccius). Fish. Res. 2013, 138, 168–178. [Google Scholar] [CrossRef] [Green Version]
- Sabatella, E.C.; Colloca, F.; Coppola, G.; Fiorentino, F.; Gambino, M.; Malvarosa, L.; Sabatella, R. Key Economic Characteristics of Italian Trawl Fisheries and Management Challenges. Front. Mar. Sci. 2017, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Grati, F.; Aladžuz, A.; Azzurro, E.; Bolognini, L.; Car, P.; Spedicato, M.T.; Stagličić, N.; Vrgoć, N.; Zerem, N.; Arneri, E.; et al. Seasonal dynamics of small-scale fisheries in the Adriatic Sea. Mediterr. Mar. Sci. 2018, 19, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Murua, H.; Saborido-Rey, F. Female reproductive strategies of marine fish species of the North Atlantic. J. Northwest Atl. Fish. Sci. 2003, 33, 23–31. [Google Scholar] [CrossRef]
- Recasens, L.; Chiericoni, V.; Belcari, P. Spawning pattern and batch fecundity of the European hake (Merluccius merluccius (Linnaeus, 1758)) in the western Mediterranean. Sci. Mar. 2008, 72, 721–732. [Google Scholar] [CrossRef] [Green Version]
- El Habouz, H.; Recasens, L.; Kifani, S.; Moukrim, A.; Bouhaimi, A.; El Ayoubi, S. Maturity and batch fecundity of the European hake (Merluccius merluccius, Linnaeus, 1758) in the eastern central Atlantic. Sci. Mar. 2011, 75, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Bouaziz, A.; Bennoui, A.; Djabali, F.; Maurin, C. Reproduction du merlu Merluccius merluccius (Linnaeus, 1758) dans la région de Bou-Ismail. CIHEAM Cah. Opt. Méditerr. 1998, 35, 109–117. [Google Scholar]
- Al-Absawy, M.A. The reproductive biology and the histological and ultrastructural characteristics in ovaries of the female gadidae fish Merluccius merluccius from the Egyptian Mediterranean water. Afr. J. Biotechnol. 2010, 9, 2544–2559. [Google Scholar]
- Nannini, N.; Pinna, D.; Chiericoni, V.; Biagi, F.; Belcari, P. Ciclo ovarico di Merluccius merluccius (Linnaeus, 1758) nel Mar Tirreno settentrionale. Biol. Mar. Mediterr. 2001, 8, 745–748. [Google Scholar]
- Carbonara, P.; Porcu, C.; Donnaloia, M.; Pesci, P.; Sion, L.; Spedicato, M.T.; Zupa, W.; Vitale, F.; Follesa, M.C. The spawning strategy of European hake (Merluccius merluccius, L. 1758) across the Western and Central Mediterranean Sea. Fish. Res. 2019, 219, 105333. [Google Scholar] [CrossRef]
- Carpentieri, P.; Colloca, F.; Cardinale, M.; Belluscio, A.; Ardizzone, G.D. Feeding habits of European hake (Merluccius merluccius) in the central Mediterranean Sea. Fish. Bull. 2005, 103, 411–416. [Google Scholar]
- Carrozzi, V.; Di Lorenzo, M.; Massi, D.; Titone, A.; Ardizzone, G.; Colloca, F. Prey preferences and ontogenetic diet shift of European hake Merluccius merluccius (Linnaeus, 1758) in the central Mediterranean Sea. Reg. Stud. Mar. Sci. 2019, 25, 100440. [Google Scholar] [CrossRef]
- Lloret, J.; Demestre, M.; Sanchez-Pardo, J. Lipid (energy) reserves of European hake (Merluccius merluccius) in the north-western Mediterranean. Vie Milieu-Life Environ. 2008, 58, 75–85. [Google Scholar]
- Cantafaro, A.; Ardizzone, G.; Enea, M.; Ligas, A.; Colloca, F. Assessing the importance of nursery areas of European hake (Merluccius merluccius) using a body condition index. Ecol. Indic. 2017, 81, 383–389. [Google Scholar] [CrossRef]
- Ligas, A.; Colloca, F.; Lundy, M.G.; Mannini, A.; Sartor, P.; Sbrana, M.; Voliani, A.; Belcari, P. Modeling the growth of recruits of European hake (Merluccius merluccius) in the northwestern Mediterranean Sea with generalized additive models. Fish. Bull. 2015, 113, 69–81. [Google Scholar] [CrossRef]
- Deniz, T.; Göktürk, D.; Ateş, C. Selectivity parameters of European hake gillnets for target and by-catch species with a perspective on small-scale fisheries management in the Sea of Marmara, Turkey. Reg. Stud. Mar. Sci. 2020, 33, 100934. [Google Scholar] [CrossRef]
- Angelini, S.; Hillary, R.; Morello, E.B.; Plagányi, É.E.; Martinelli, M.; Manfredi, C.; Isajlović, I.; Santojanni, A. An Ecosystem Model of Intermediate Complexity to test management options for fisheries: A case study. Ecol. Model. 2016, 319, 218–232. [Google Scholar] [CrossRef]
- Tsagarakis, K.; Palialexis, A.; Vassilopoulou, V. Mediterranean fishery discards: Review of the existing knowledge. ICES J. Mar. Sci. 2014, 71, 1219–1234. [Google Scholar] [CrossRef] [Green Version]
- Candelma, M.; Fontaine, R.; Colella, S.; Santojanni, A.; Weltzien, F.-A.; Carnevali, O. Gonadotropin characterization, localization and expression in the European hake (Merluccius merluccius). Reproduction 2017, 153, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Candelma, M.; Valle, L.D.; Colella, S.; Santojanni, A.; Carnevali, O. Cloning, characterization, and molecular expression of gonadotropin receptors in European hake (Merluccius merluccius), a multiple-spawning species. Fish Physiol. Biochem. 2018, 44, 895–910. [Google Scholar] [CrossRef] [PubMed]
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; Macewicz, B.J.; Lowerre-Barbieri, S.K. A standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Murua, H.; Motos, L.; Lucio, P. Reproductive modality and batch fecundity of the European hake (Merluccius merluccius L.) in the Bay of Biscay. Calif. Coop. Ocean. Fish. Investig. Rep. 1998, 39, 196–203. [Google Scholar]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Murua, H.; Kraus, G.; Saborido-Rey, F.; Witthames, P.R.; Thorsen, A.; Junquera, S. Procedures to estimate fecundity of marine fish species in relation to their reproductive strategy. J. Northwest Atl. Fish. Sci. 2003, 33, 33–54. [Google Scholar] [CrossRef]
- Murua, H.; Lucio, P.; Santurtún, M.; Motos, L. Seasonal variation in egg production and batch fecundity of European hake Merluccius merluccius (L.) in the Bay of Biscay. J. Fish Biol. 2006, 69, 1304–1316. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 April 2021).
- Ogle, D.; Wheeler, P.; Dinno, A. FSA: Fisheries stock analysis. In R Package Version 0.8.25; Available online: https://github.com/droglenc/FSA (accessed on 1 April 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Lo Martire, R. R package version 1.3.1. Reliab. Coeff. 2017. Available online: https://CRAN.R-project.org/package=rel (accessed on 1 April 2021).
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Petit, R.; Korta, M.; Saborido-Rey, F.; Murua, H.; Sainza, M.; Piñeiro, C. Changes in size at maturity of European hake Atlantic populations in relation with stock structure and environmental regimes. J. Mar. Syst. 2008, 71, 260–278. [Google Scholar] [CrossRef] [Green Version]
- Vitale, F.; Svedäng, H.; Cardinale, M. Histological analysis invalidates macroscopically determined maturity ogives of the Kattegat cod (Gadus morhua) and suggests new proxies for estimating maturity status of individual fish. ICES J. Mar. Sci. 2006, 63, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Marisaldi, L.; Basili, D.; Candelma, M.; Sesani, V.; Pignalosa, P.; Gioacchini, G.; Carnevali, O. Maturity assignment based on histology-validated macroscopic criteria: Tackling the stock decline of the Mediterranean swordfish (Xiphias gladius). Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1–12. [Google Scholar] [CrossRef]
- Murua, H.; Motos, L. Reproductive strategy and spawning activity of the European hake Merluccius merluccius (L.) in the Bay of Biscay. J. Fish Biol. 2006, 69, 1288–1303. [Google Scholar] [CrossRef]
- Recasens, L.; Lombarte, A.; Morales-Nin, B.; Tores, G.J. Spatiotemporal variation in the population structure of the European hake in the NW Mediterranean. J. Fish Biol. 1998, 53, 387–401. [Google Scholar] [CrossRef]
- Mihanović, H.; Vilibić, I.; Carniel, S.; Tudor, M.; Russo, A.; Bergamasco, A.; Bubić, N.; Ljubešić, Z.; Viličić, D.; Boldrin, A.; et al. Exceptional dense water formation on the Adriatic shelf in the winter of 2012. Ocean Sci. 2013, 9, 561–572. [Google Scholar] [CrossRef] [Green Version]
- Janekovic, I.; Mihanovic, H.; Vilibic, I.; Tudor, M. Extreme cooling and dense water formation estimates in open and coastal regions of the Adriatic Sea during the winter of 2012. J. Geophys. Res. Ocean. 2014, 119, 3200–3218. [Google Scholar] [CrossRef]
- Benetazzo, A.; Bergamasco, A.; Bonaldo, D.; Falcieri, F.M.; Sclavo, M.; Langone, L.; Carniel, S. Response of the Adriatic Sea to an intense cold air outbreak: Dense water dynamics and wave-induced transport. Prog. Oceanogr. 2014, 128, 115–138. [Google Scholar] [CrossRef]
- Dominguez-Petit, R.; Saborido-Rey, F.; Medina, I. Changes of proximate composition, energy storage and condition of European hake (Merluccius merluccius, L. 1758) through the spawning season. Fish. Res. 2010, 104, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Lloret, J.; Shulman, G.; Love, M.R. Condition and Health Indicators of Exploited Marine Fishes; Wiley Blackwell: Hoboken, NJ, USA, 2014; ISBN 978-0-470-67024-8. [Google Scholar]
- Jones, R.E.; Petrell, R.J.; Pauly, D. Using modified length-weight relationships to assess the condition of fish. Aquac. Eng. 1999, 20, 261–276. [Google Scholar] [CrossRef]
- Flores, A.; Wiff, R.; Ganias, K.; Marshall, C.T. Accuracy of gonadosomatic index in maturity classification and estimation of maturity ogive. Fish. Res. 2019, 210, 50–62. [Google Scholar] [CrossRef]
- Lucio, P.; Murua, H.; Santurtun, M. Growth and reproduction of hake (Merluccius merluccius) in the Bay of Biscay during the period 1996–1997. Ozeanografika 2000, 3, 325–354. [Google Scholar]
- Zupanovic, S.; Jardas, I. A contribution to the study of biology and population dynamics of the Adriatic hake, Merluccius merluccius (L.). Acta Adriat. 1986, 27, 97–146. [Google Scholar]
- Alegria Hernandez, V.; Jukic, S. Abundance dynamics of the hake (Merluccius merluccius L.) from the middle Adriatic sea. Bull. l’Inst. Océanogr. 1992, 11, 151–161. [Google Scholar]
- Anderson, C.N.K.; Hsieh, C.; Sandin, S.A.; Hewitt, R.; Hollowed, A.; Beddington, J.; May, R.M.; Sugihara, G. Why fishing magnifies fluctuations in fish abundance. Nature 2008, 452, 835–839. [Google Scholar] [CrossRef]
- Korta, M.; Dominguez-Petit, R.; Murua, H.; Saborido-Rey, F. Regional variability in reproductive traits of European hake Merluccius merluccius L. populations. Fish. Res. 2010, 104, 64–72. [Google Scholar] [CrossRef]
- FAO. Working Group on Stock Assessment of Demersal Species (WGSAD). Gen. Fish. Comm. Mediterr. 2017. Available online: http://www.fao.org/gfcm/reports/technical-meetings/detail/en/c/1105308/ (accessed on 1 April 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ovarian Phases | Macroscopic Morphology | Histological Aspect | Follicle Diameter (µm) |
|---|---|---|---|
| Immature | Orange, semi-transparent | Presence of oogonia (O), primary oocytes (PO). Scarce connective tissue and well compact ovigerous lamellae. | <250 |
| Developing | Small pink but some oocytes visible | Presence of early (vtg1) and middle (vtg2) vitellogenic oocytes. Yolk vesicles form in the ooplasm. Lipid granules occupy a larger cytoplasmic area than yolk droplets. The zona radiata thickens. | 250–550 |
| Spawning capable and actively spawning subphase | Large ovaries, oocytes visible macroscopically. An abundance of hydrated oocytes in actively spawning subphase. | Presence of late vitellogenic oocytes (vtg3). Lipid globules occupy a cytoplasmic area like that occupied by yolk granules. Actively spawning subphase: lipid globules fuse into a single larger oil droplet, yolk droplets start to coalescence, the nucleus starts to migrate peripherally to the animal pole (Mn) and POFs. Follicles grow due to water uptake and become transparent (H). Zona radiata is thinner than in previous stages | 550–1150 |
| Regressing (postspawning) | Flaccid and small ovaries, blood vessels prominent. | Atresia (any stage) and POFs present. Some CA and/or vtg1, vtg2 oocytes present. | <250 |
| Regenerating | Pinkish and small ovaries that occupy 1/3 of the body cavity, blood vessels reduced but present. | Presence of oogonia (O), primary oocytes (PO), cortical alveoli (CA), and lipid stage oocytes (LS). The ovarian wall is thickened. | <250 |
| Model Estimates HISTOLOGY | ||||
| Estimate | Std. Error | z value | p-value | |
| Intercept | −9.17104 | 2.00365 | −4.577 | p < 0.001 |
| Total_length_cm | 0.27183 | 0.05925 | 4.588 | p < 0.0001 |
| Model Estimates MACROSCOPIC | ||||
| Estimate | Std. Error | z value | p-value | |
| Intercept | −9.15307 | 2.13655 | −4.284 | p < 0.0001 |
| Total_length_cm | 0.29703 | 0.06505 | 4.566 | p < 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Candelma, M.; Marisaldi, L.; Bertotto, D.; Radaelli, G.; Gioacchini, G.; Santojanni, A.; Colella, S.; Carnevali, O. Aspects of Reproductive Biology of the European Hake (Merluccius merluccius) in the Northern and Central Adriatic Sea (GSA 17-Central Mediterranean Sea). J. Mar. Sci. Eng. 2021, 9, 389. https://doi.org/10.3390/jmse9040389
Candelma M, Marisaldi L, Bertotto D, Radaelli G, Gioacchini G, Santojanni A, Colella S, Carnevali O. Aspects of Reproductive Biology of the European Hake (Merluccius merluccius) in the Northern and Central Adriatic Sea (GSA 17-Central Mediterranean Sea). Journal of Marine Science and Engineering. 2021; 9(4):389. https://doi.org/10.3390/jmse9040389
Chicago/Turabian StyleCandelma, Michela, Luca Marisaldi, Daniela Bertotto, Giuseppe Radaelli, Giorgia Gioacchini, Alberto Santojanni, Sabrina Colella, and Oliana Carnevali. 2021. "Aspects of Reproductive Biology of the European Hake (Merluccius merluccius) in the Northern and Central Adriatic Sea (GSA 17-Central Mediterranean Sea)" Journal of Marine Science and Engineering 9, no. 4: 389. https://doi.org/10.3390/jmse9040389