
Effects of Ocean Acidification and Summer Thermal Stress on the Physiology and Growth of the Atlantic Surfclam (Spisula solidissima)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Husbandry
2.2. Experimental Design
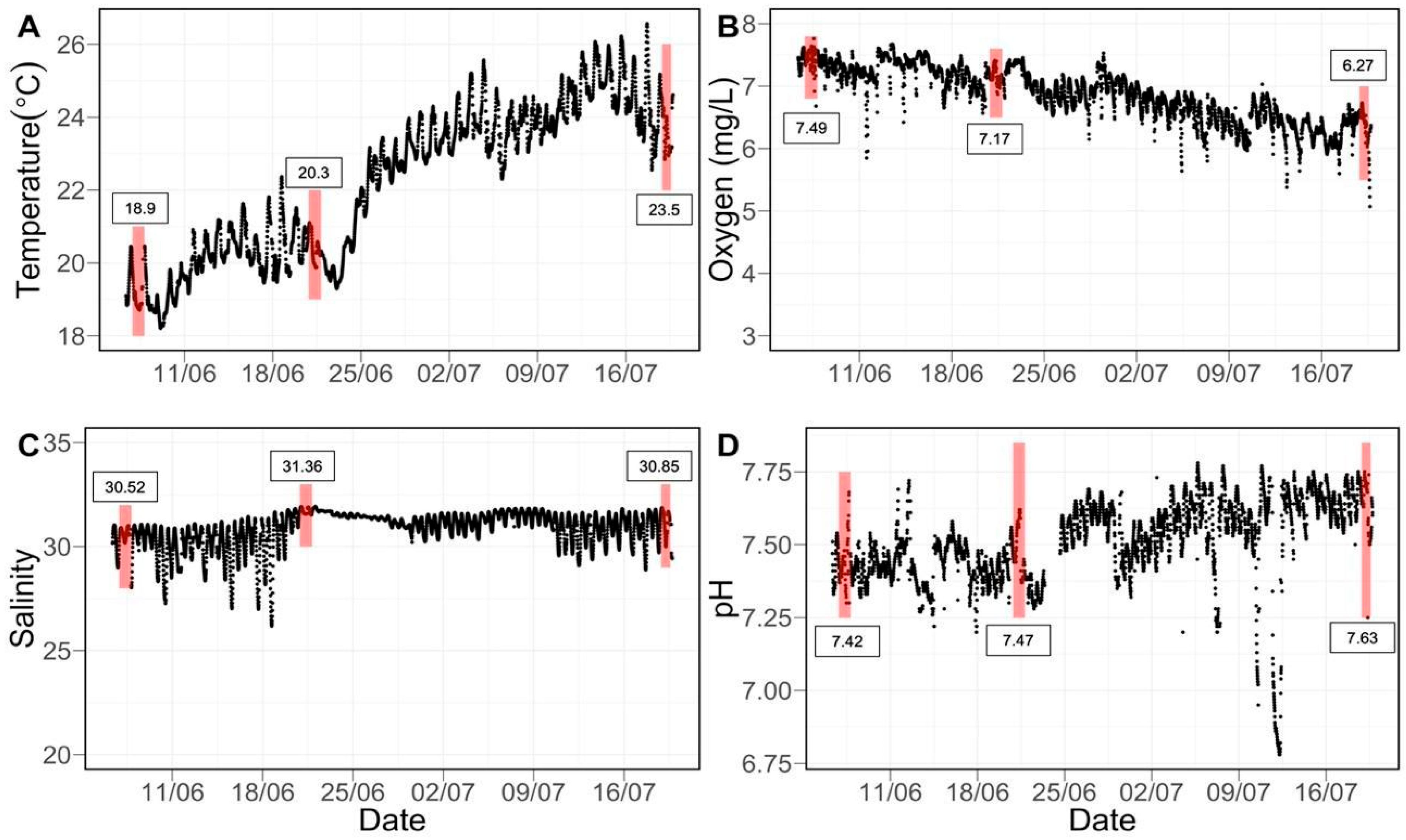
2.3. Environmental Monitoring
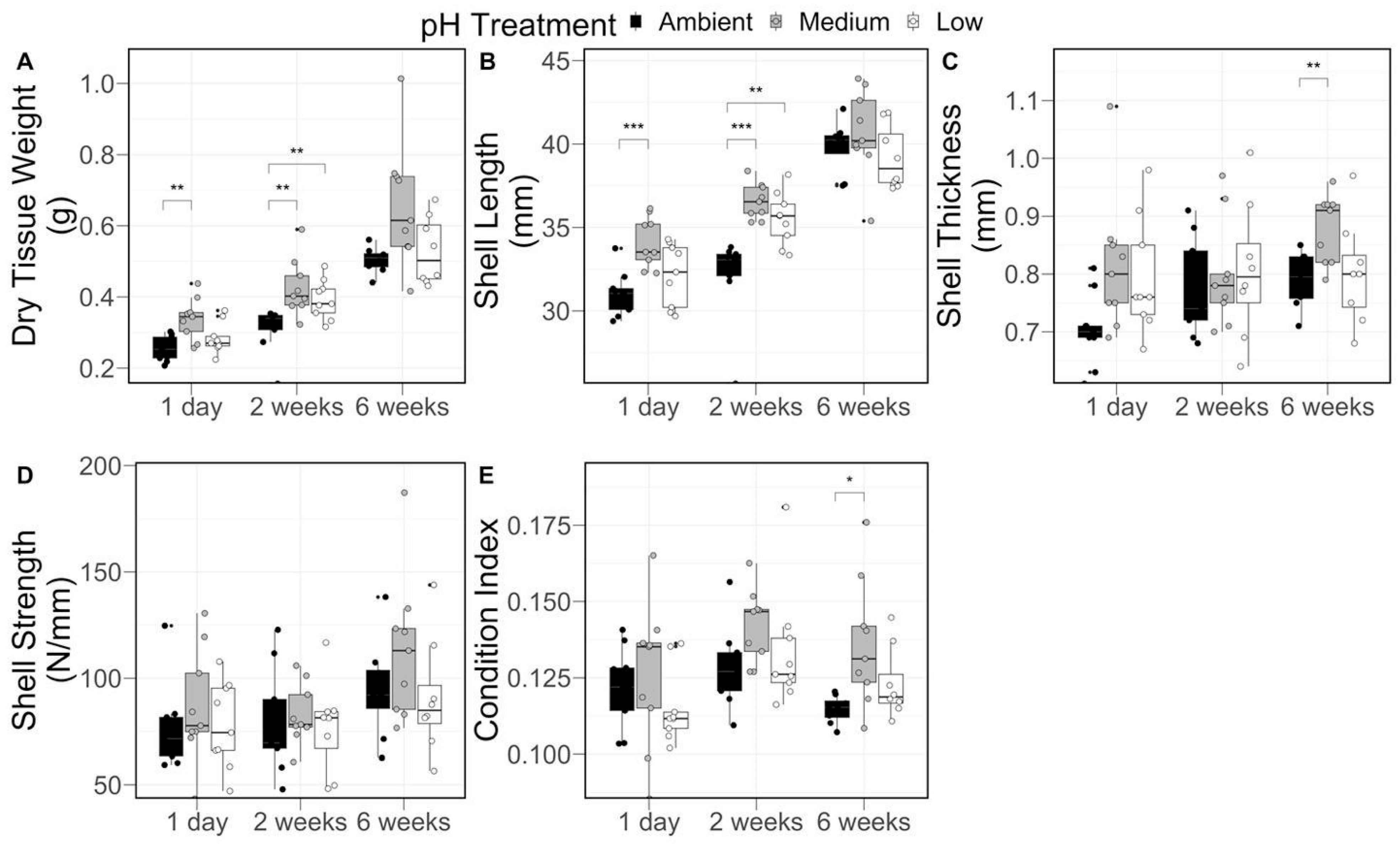
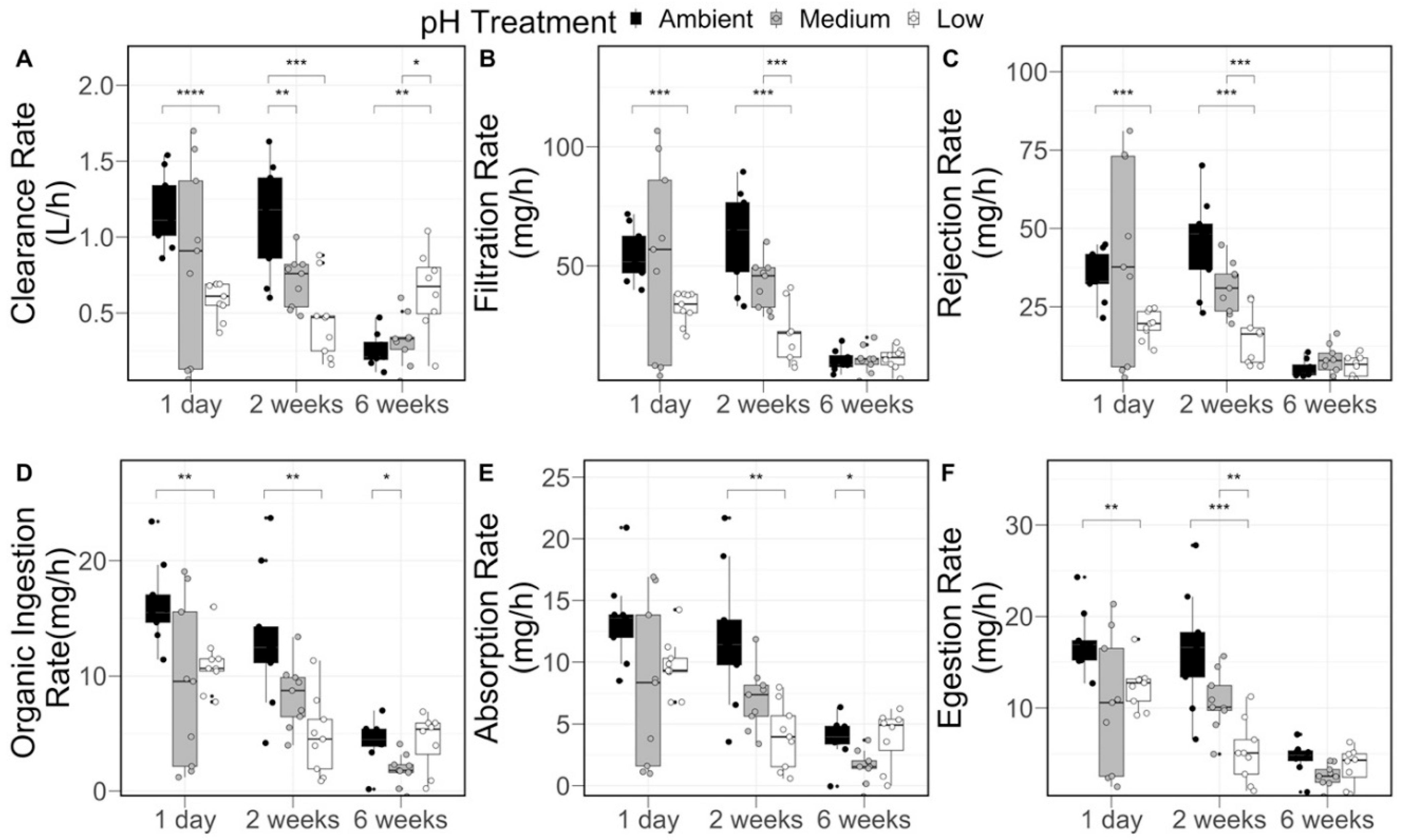
2.4. Surfclam Physiology and Growth Measurements
2.5. Statistical Analyses
3. Results
3.1. Water Quality and Experimental Conditions
3.2. Clam Growth and Physiology
4. Discussion
4.1. Effects of Ocean Acidification and Warming on Bivalve Physiology
4.2. Design of Laboratory Experiments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robert, G. Dynamics of an Exploited Population of Bar Clam, Spisula solidissima. In Canadian Manuscript Report of Fisheries and Aquatic Sciences; Department of Fisheries and Oceans Canada: Ottawa, ON, Canada, 1981; Volume 1670, iv + 12 p. [Google Scholar]
- Marzec, R.J.; Kim, Y.; Powell, E.N. Geographical Trends in Weight and Condition Index of Surfclams (Spisula solidissima) in the Mid-Atlantic Bight. J. Shellfish Res. 2010, 29, 117–128. [Google Scholar] [CrossRef]
- NOAA Fisheries Landings. 2023. Available online: https://www.fisheries.noaa.gov/foss/f?p=215:200 (accessed on 1 March 2023).
- Weinberg, J.R. Bathymetric Shift in the Distribution of Atlantic Surfclams: Response to Warmer Ocean Temperature. ICES J. Mar. Sci. 2005, 62, 1444–1453. [Google Scholar] [CrossRef]
- Hofmann, E.E.; Powell, E.N.; Klinck, J.M.; Munroe, D.M.; Mann, R.; Haidvogel, D.B.; NarvÁEz, D.A.; Zhang, X.; Kuykendall, K.M. An Overview of Factors Affecting Distribution of the Atlantic Surfclam (Spisula solidissima), a Continental Shelf Biomass Dominant, During a Period of Climate Change. J. Shellfish Res. 2018, 37, 821–831. [Google Scholar] [CrossRef]
- Feely, R.; Doney, S.; Cooley, S. Ocean Acidification: Present Conditions and Future Changes in a High-CO2 World. Oceanography 2009, 22, 36–47. [Google Scholar] [CrossRef]
- Ma, D.; Gregor, L.; Gruber, N. Four Decades of Trends and Drivers of Global Surface Ocean Acidification. Glob. Biogeochem. Cycles 2023, 37, e2023GB007765. [Google Scholar] [CrossRef]
- Wanninkhof, R.; Barbero, L.; Byrne, R.; Cai, W.-J.; Huang, W.-J.; Zhang, J.-Z.; Baringer, M.; Langdon, C. Ocean Acidification along the Gulf Coast and East Coast of the USA. Cont. Shelf Res. 2015, 98, 54–71. [Google Scholar] [CrossRef]
- Wright-Fairbanks, E.K.; Miles, T.N.; Cai, W.; Chen, B.; Saba, G.K. Autonomous Observation of Seasonal Carbonate Chemistry Dynamics in the Mid-Atlantic Bight. JGR Ocean. 2020, 125, e2020JC016505. [Google Scholar] [CrossRef]
- Green, M.A.; Waldbusser, G.G.; Reilly, S.L.; Emerson, K.; O’Donnell, S. Death by Dissolution: Sediment Saturation State as a Mortality Factor for Juvenile Bivalves. Limnol Ocean. 2009, 54, 1037–1047. [Google Scholar] [CrossRef]
- Ventura, A.; Schulz, S.; Dupont, S. Maintained Larval Growth in Mussel Larvae Exposed to Acidified Under-Saturated Seawater. Sci. Rep. 2016, 6, 23728. [Google Scholar] [CrossRef] [PubMed]
- Ekstrom, J.A.; Suatoni, L.; Cooley, S.R.; Pendleton, L.H.; Waldbusser, G.G.; Cinner, J.E.; Ritter, J.; Langdon, C.; Van Hooidonk, R.; Gledhill, D.; et al. Vulnerability and Adaptation of US Shellfisheries to Ocean Acidification. Nat. Clim. Chang. 2015, 5, 207–214. [Google Scholar] [CrossRef]
- Xu, X.; Yang, F.; Zhao, L.; Yan, X. Seawater Acidification Affects the Physiological Energetics and Spawning Capacity of the Manila Clam Ruditapes philippinarum during Gonadal Maturation. Comp. Biochem. Phys. A 2016, 196, 20–29. [Google Scholar] [CrossRef]
- Navarro, J.M.; Villanueva, P.; Rocha, N.; Torres, R.; Chaparro, O.R.; Benítez, S.; Andrade-Villagrán, P.V.; Alarcón, E. Plastic Response of the Oyster Ostrea chilensis to Temperature and pCO2 within the Present Natural Range of Variability. PLoS ONE 2020, 15, e0234994. [Google Scholar] [CrossRef]
- Pousse, E.; Poach, M.E.; Redman, D.H.; Sennefelder, G.; White, L.E.; Lindsay, J.M.; Munroe, D.; Hart, D.; Hennen, D.; Dixon, M.S.; et al. Energetic Response of Atlantic Surfclam Spisula solidissima to Ocean Acidification. Mar. Pollut. Bull. 2020, 161, 111740. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, X.; Rastrick, S.P.S.; Wang, J.; Zhang, Y.; Strand, Ø.; Fang, J.; Jiang, Z. Effects of Elevated p CO2 on the Physiological Energetics of Pacific Oyster, Crassostrea gigas. ICES J. Mar. Sci. 2021, fsab139. [Google Scholar] [CrossRef]
- Czaja, R.; Holmberg, R.; Pales Espinosa, E.; Hennen, D.; Cerrato, R.; Lwiza, K.; O’Dwyer, J.; Beal, B.; Root, K.; Zuklie, H.; et al. Behavioral and Physiological Effects of Ocean Acidification and Warming on Larvae of a Continental Shelf Bivalve. Mar. Pollut. Bull. 2023, 192, 115048. [Google Scholar] [CrossRef] [PubMed]
- Range, P.; Chícharo, M.A.; Ben-Hamadou, R.; Piló, D.; Matias, D.; Joaquim, S.; Oliveira, A.P.; Chícharo, L. Calcification, Growth and Mortality of Juvenile Clams Ruditapes decussatus under Increased pCO2 and Reduced pH: Variable Responses to Ocean Acidification at Local Scales? J. Exp. Mar. Biol. Ecol. 2011, 396, 177–184. [Google Scholar] [CrossRef]
- Acquafredda, M.P.; Ximing, G.; Munroe, D.M. Transcriptomic Response of the Atlantic Surfclam (Spisula solidissima) to Acute Heat Stress. Mar. Biotechnol. 2024, 26, 149–168. [Google Scholar] [CrossRef]
- Pousse, É.; Munroe, D.; Hart, D.; Hennen, D.; Cameron, L.P.; Rheuban, J.E.; Wang, Z.A.; Wikfors, G.H.; Meseck, S.L. Dynamic Energy Budget Modeling of Atlantic Surfclam, Spisula solidissima, under Future Ocean Acidification and Warming. Mar. Environ. Res. 2022, 177, 105602. [Google Scholar] [CrossRef]
- Bockmon, E.E.; Dickson, A.G. An Inter-Laboratory Comparison Assessing the Quality of Seawater Carbon Dioxide Measurements. Mar. Chem. 2015, 171, 36–43. [Google Scholar] [CrossRef]
- Dickson, A.G.; Sabine, C.L.; Christian, J.R. Guide to Best Practices for Ocean CO2 Measurements. North Pacific Marine Science Organization: Sidney, BC, Canada, 2007; ISBN 1-897176-07-4. [Google Scholar]
- Pierrot, D.; Lewis, E.; Wallace, D. MS Excel Program Developed for CO2 System Calculations; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy: Oak Ridge, TN, USA, 2006; ORNL/CDIAC-105a.
- Lueker, T.J.; Dickson, A.G.; Keeling, C.D. Ocean pCO2 Calculated from Dissolved Inorganic Carbon, Alkalinity, and Equations for K1 and K2: Validation Based on Laboratory Measurements of CO2 in Gas and Seawater at Equilibrium. Mar. Chem. 2000, 70, 105–119. [Google Scholar] [CrossRef]
- Dickson, A.G.; Wesolowski, D.J.; Palmer, D.A.; Mesmer, R.E. Dissociation Constant of Bisulfate Ion in Aqueous Sodium Chloride Solutions to 250.Degree.C. J. Phys. Chem. 1990, 94, 7978–7985. [Google Scholar] [CrossRef]
- Lee, K.; Kim, T.-W.; Byrne, R.H.; Millero, F.J.; Feely, R.A.; Liu, Y.-M. The Universal Ratio of Boron to Chlorinity for the North Pacific and North Atlantic Oceans. Geochim. Cosmochim. Acta 2010, 74, 1801–1811. [Google Scholar] [CrossRef]
- Iglesias, J.I.P.; Urrutia, M.B.; Navarro, E.; Ibarrola, I. Measuring Feeding and Absorption in Suspension-Feeding Bivalves: An Appraisal of the Biodeposition Method. J. Exp. Mar. Biol. Ecol. 1998, 219, 71–86. [Google Scholar] [CrossRef]
- Galimany, E.; Rose, J.M.; Dixon, M.S.; Wikfors, G.H. Transplant Experiment to Evaluate the Feeding Behaviour of the Atlantic Ribbed Mussel, Geukensia demissa, Moved to a High Inorganic Seston Area. Mar. Freshw. Res. 2015, 66, 220. [Google Scholar] [CrossRef]
- Galimany, E.; Lunt, J.; Domingos, A.; Paul, V.J. Feeding Behavior of the Native Mussel Ischadium recurvum and the Invasive Mussels Mytella charruana and Perna viridis in FL, USA, Across a Salinity Gradient. Estuar. Coasts 2018, 41, 2378–2388. [Google Scholar] [CrossRef]
- Galimany, E.; Rose, J.M.; Dixon, M.S.; Wikfors, G.H. Quantifying Feeding Behavior of Ribbed Mussels (Geukensia demissa) in Two Urban Sites (Long Island Sound, USA) with Different Seston Characteristics. Estuar. Coasts 2013, 36, 1265–1273. [Google Scholar] [CrossRef]
- Galimany, E.; Lunt, J.; Freeman, C.; Reed, S.; Segura-García, I.; Paul, V. Feeding Behavior of Eastern Oysters Crassostrea virginica and Hard Clams Mercenaria mercenaria in Shallow Estuaries. Mar. Ecol. Prog. Ser. 2017, 567, 125–137. [Google Scholar] [CrossRef]
- Coughlan, J. The Estimation of Filtering Rate from the Clearance of Suspensions. Mar. Biol. 1969, 2, 356–358. [Google Scholar] [CrossRef]
- Winter, J.E. The Filtration Rate of Mytilus Edulis and Its Dependence on Algal Concentration, Measured by a Continuous Automatic Recording Apparatus. Mar. Biol. 1973, 22, 317–328. [Google Scholar] [CrossRef]
- Urrutia, M.; Iglesias, J.; Navarro, E.; Prou, J. Feeding and Absorption in Cerastoderma Edule under Environmental Conditions in the Bay of Marennesoleron (Western France). J. Mar. Biol. Assoc. UK 1996, 76, 431–450. [Google Scholar] [CrossRef]
- Roy, K.; Miller, D.J.; Labarbera, M. Taphonomic Bias in Analyses of Drilling Predation: Effects of Gastropod Drill Holes on Bivalve Shell Strength. Palaios 1994, 9, 413–421. [Google Scholar] [CrossRef]
- DiRenzi, B.; Munroe, D.; Saba, G.; López, A. Biological Response of Spisula solidissima (Atlantic Surfclams) to Varying Carbonate Chemistry in the Mid-Atlantic Bight. In Proceedings of the OCEANS 2023–MTS/IEEE U.S. Gulf Coast, Biloxi, MS, USA, 25–28 September 2023; pp. 1–7. [Google Scholar]
- Navarro, J.M.; Duarte, C.; Manríquez, P.H.; Lardies, M.A.; Torres, R.; Acuña, K.; Vargas, C.A.; Lagos, N.A. Ocean Warming and Elevated Carbon Dioxide: Multiple Stressor Impacts on Juvenile Mussels from Southern Chile. ICES J. Mar. Sci. 2016, 73, 764–771. [Google Scholar] [CrossRef]
- Rastrick, S.P.S.; Collier, V.; Graham, H.; Strohmeier, T.; Whiteley, N.M.; Strand, Ø. Feeding Plasticity More than Metabolic Rate Drives the Productivity of Economically Important Filter Feeders in Response to Elevated CO2 and Reduced Salinity. ICES J. Mar. Sci. 2018, 75, 2117–2128. [Google Scholar] [CrossRef]
- Meseck, S.L.; Sennefelder, G.; Krisak, M.; Wikfors, G.H. Physiological Feeding Rates and Cilia Suppression in Blue Mussels (Mytilus edulis) with Increased Levels of Dissolved Carbon Dioxide. Ecol. Indic. 2020, 117, 106675. [Google Scholar] [CrossRef]
- Liang, J.; Liu, Y.; Zhu, F.; Li, Y.; Liang, S.; Guo, Y. Impact of Ocean Acidification on the Physiology of Digestive Gland of Razor Clams Sinonovacula constricta. Front. Mar. Sci. 2022, 9, 1010350. [Google Scholar] [CrossRef]
- Hiebenthal, C.; Philipp, E.E.R.; Eisenhauer, A.; Wahl, M. Effects of Seawater pCO2 and Temperature on Shell Growth, Shell Stability, Condition and Cellular Stress of Western Baltic Sea Mytilus edulis (L.) and Arctica islandica (L.). Mar. Biol. 2013, 160, 2073–2087. [Google Scholar] [CrossRef]
- Poach, M.; Munroe, D.; Vasslides, J.; Abrahamsen, I.; Coffey, N. Monitoring Coastal Acidification along the U.S. East Coast: Conerns for Shellfish Production. In Proceedings of the 45th Scientific Symposium of UJNR Aquaculture Panel, Hiroshima, Japan, 16–17 October 2017; Volume 49, pp. 53–64. [Google Scholar]
- Hennen, D.R.; Mann, R.; Munroe, D.M.; Powell, E.N. Biological Reference Points for Atlantic Surfclam (Spisula solidissima) in Warming Seas. Fish. Res. 2018, 207, 126–139. [Google Scholar] [CrossRef]
- Hornstein, J.; Pales Espinosa, E.; Cerrato, R.M.; Lwiza, K.M.M.; Allam, B. The Influence of Temperature Stress on the Physiology of the Atlantic Surfclam, Sp. Solidissima. Comp. Biochem. Phys. A 2018, 222, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Acquafredda, M.P.; Munroe, D.M.; Ragone Calvo, L.M.; De Luca, M. The Effect of Rearing Temperature on the Survival and Growth of Early Juvenile Atlantic Surfclams (Spisula solidissima). Aquac. Rep. 2019, 13, 100176. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Farrell, A.P. Physiology and Climate Change. Science 2008, 322, 690–692. [Google Scholar] [CrossRef]
- Parker, L.M.; Scanes, E.; O’Connor, W.A.; Coleman, R.A.; Byrne, M.; Pörtner, H.-O.; Ross, P.M. Ocean Acidification Narrows the Acute Thermal and Salinity Tolerance of the Sydney Rock Oyster Saccostrea glomerata. Mar. Pollut. Bull. 2017, 122, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.C. Elevated Temperature Has Adverse Effects on GABA-Mediated Avoidance Behaviour to Sediment Acidification in a Wide-Ranging Marine Bivalve. Mar. Biol. 2017, 11, 56. [Google Scholar] [CrossRef]
- Velasco, L.; Navarro, J. Feeding Physiology of Two Bivalves under Laboratory and Field Conditions in Response to Variable Food Concentrations. Mar. Ecol. Prog. Ser. 2005, 291, 115–124. [Google Scholar] [CrossRef]
- Bayne, B.L.; Iglesias, J.I.P.; Hawkins, A.J.S.; Navarro, E.; Heral, M.; Deslous-Paoli, J.M. Feeding Behaviour of the Mussel, Mytilus edulis: Responses to Variations in Quantity and Organic Content of the Seston. J. Mar. Biol. Ass. 1993, 73, 813–829. [Google Scholar] [CrossRef]
- Marsden, I.D. Respiration and Feeding of the Surf Clam Paphies Donacina from New Zealand. Hydrobologia 1999, 405, 179–188. [Google Scholar] [CrossRef]
- Iglesias, J.I.P.; Navarro, E.; Alvarez Jorna, P.; Armentina, I. Feeding, Particle Selection and Absorption in Cockles Cerastoderma edule (L.) Exposed to Variable Conditions of Food Concentration and Quality. J. Exp. Mar. Biol. Ecol. 1992, 162, 177–198. [Google Scholar] [CrossRef]
- Widdows, J. Physiological Adaptation of Mytilus edulis to Cyclic Temperatures. J. Comp. Physiol. B 1976, 105, 115–128. [Google Scholar] [CrossRef]
- Small, D.P.; Milazzo, M.; Bertolini, C.; Graham, H.; Hauton, C.; Hall-Spencer, J.M.; Rastrick, S.P.S. Temporal Fluctuations in Seawater pCO2 May Be as Important as Mean Differences When Determining Physiological Sensitivity in Natural Systems. ICES J. Mar. Sci. 2016, 73, 604–612. [Google Scholar] [CrossRef]
- Connor, K.; Gracey, A.Y. Cycles of Heat and Aerial-Exposure Induce Changes in the Transcriptome Related to Cell Regulation and Metabolism in Mytilus californianus. Mar. Biol. 2020, 167, 132. [Google Scholar] [CrossRef]
- Bitter, M.C.; Kapsenberg, L.; Silliman, K.; Gattuso, J.-P.; Pfister, C.A. Magnitude and Predictability of pH Fluctuations Shape Plastic Responses to Ocean Acidification. Am. Nat. 2021, 197, 486–501. [Google Scholar] [CrossRef]
- Zhang, H.; Shin, P.K.S.; Cheung, S.G. Physiological Responses and Scope for Growth upon Medium-Term Exposure to the Combined Effects of Ocean Acidification and Temperature in a Subtidal Scavenger Nassarius conoidalis. Mar. Environ. Res. 2015, 106, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, J.; Casties, I.; Pansch, C.; Körtzinger, A.; Melzner, F. Food Availability Outweighs Ocean Acidification Effects in Juvenile Mytilus edulis: Laboratory and Field Experiments. Glob. Chang. Biol. 2013, 19, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Swanson, L.; Sindermann, C.J. Oxygen Depletion and Associated Benthic Mortalities in New York Bight, 1976. NOAA Prof. Pap. 1979, 11, 345. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Term | Definition | Units | References |
|---|---|---|---|
| Clearance Rate | Volume of water cleared of particles, by the bivalve, per unit time | L h−1 | Coughlan 1969 ref. [32] |
| Filtration Rate | Volume of water cleared of particles, by the bivalve, per unit time | mg h−1 | Winter 1973 ref. [33] |
| Rejection Rate | Mass of pseudofeces production (rejected material, captured but not ingested) per unit time | mg h−1 | Iglesias et al., 1998 ref. [27] |
| Organic Ingestion Rate | Mass of organic material ingested per unit time | mg h−1 | Iglesias et al., 1998 ref. [27] |
| Absorption Rate | Uptake of nutrients across gut surface per time, often measured as organic ingestion rate minus organic egestion rate | mg h−1 | (Urrutia et al., 1996 ref. [34] |
| Egestion Rate | Mass of feces production per unit time | mg h−1 | Iglesias et al., 1998 ref. [27] |
| Diet | 1 Day | 2 Weeks | 6 Weeks |
|---|---|---|---|
| TPM (mg L−1) | 55.43 ± 22.31 a | 50.06 ± 15.20 a | 29.23 ± 14.34 b |
| POM (mg L−1) | 7.81 ± 3.00 a | 6.10 ± 1.97 b | 4.67 ± 1.22 c |
| %Organic | 14.39 ± 21.3 a | 12.46 ± 3.12 a | 17.50 ± 4.50 b |
| pH Treatment | |||
|---|---|---|---|
| Ambient | Medium | Low | |
| Temperature (°C) | 22.13 ± 2.06 | 22.07 ± 2.09 | 21.99 ± 2.10 |
| Oxygen (mgL−1) | 6.92 ± 0.43 | 6.92 ± 0.45 | 6.99 ± 0.43 |
| Salinity | 30.66 ± 1.29 | 30.64 ± 1.28 | 30.63 ± 1.27 |
| pH in situ (seawater scale) | 7.81 ± 0.07 a | 7.51 ± 0.11 b | 7.20 ± 0.10 c |
| DIC (μmol kg−1) | 1992.68 ± 70.31 a | 2097.79 ± 66.61 a,b | 2178.28 ± 98.04 b |
| Total alkalinity (μmol/kg) | 2080.79 ± 58.15 | 2084.18 ± 54.96 | 2085.90 ± 53.64 |
| ρCO2 * (μatm) | 1002.10 ± 288.35 a | 2274.74 ± 369.15 b | 4293.57 ± 1289.67 c |
| Ω * Aragonite | 1.30 ± 0.27 a | 0.62 ± 0.09 b | 0.37 ± 0.11 c |
| Growth Measurement | pH Treatment | Sampling Time (Temperature °C) | Interaction p-Value | ||
|---|---|---|---|---|---|
| 1 Day (18.9 °C) | 2 Weeks (20.3 °C) | 6 Weeks (23.5 °C) | |||
| Dry Tissue Weight (g) | Ambient | 0.25 ± 0.04 a | 0.31 ± 0.06 a | 0.50 ± 0.04 b | 0.389 |
| Medium | 0.34 ± 0.06 a | 0.43 ± 0.08 a | 0.66 ± 0.17 b | ||
| Low | 0.28 ± 0.28 a | 0.39 ± 0.06 a | 0.53 ± 0.09 b | ||
| Shell Length (mm) | Ambient | 31.03 ± 1.33 a | 32.17 ± 2.54 a | 39.86 ± 1.56 b | 0.019 |
| Medium | 34.13 ± 1.51 a | 36.56 ± 1.07 a | 40.69 ± 2.61 b | ||
| Low | 32.16 ± 1.87 a | 35.53 ± 1.58 b | 39.19 ± 1.89 c | ||
| Shell Thickness (mm) | Ambient | 0.70 ± 0.06 | 0.78 ± 0.09 | 0.79 ± 0.05 | 0.376 |
| Medium | 0.81 ± 0.12 | 0.80 ± 0.09 | 0.88 ± 0.06 | ||
| Low | 0.79 ± 0.10 | 0.79 ± 0.12 | 0.80 ± 0.09 | ||
| Shell Strength (N/mm) | Ambient | 76.70 ± 19.96 | 80.05 ± 24.67 | 94.61 ± 23.03 | 0.828 |
| Medium | 86.61 ± 26.69 | 83.02 ± 14.24 | 113.37 ± 34.14 | ||
| Low | 77.84 ± 20.21 | 61.16 ± 19.23 | 90.96 ± 27.21 | ||
| Condition Index | Ambient | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.11 ± 0.00 | 0.480 |
| Medium | 0.13 ± 0.02 | 0.14 ± 0.01 | 0.14 ± 0.02 | ||
| Low | 0.12 ± 0.01 | 0.13 ± 0.02 | 0.12 ± 0.01 | ||
| Physiological Rate | pH Treatment | Sampling Time | Interaction p-Value | ||
|---|---|---|---|---|---|
| 1 Day (18.9 °C) | 2 Weeks (20.3 °C) | 6 Weeks (23.5 °C) | |||
| Clearance Rate (L h−1) | Ambient | 1.18 ± 0.24 a | 1.13 ± 0.36 a | 0.25 ± 0.12 b | p < 0.001 |
| Medium | 0.85 ± 0.64 a | 0.71 ± 0.17 a,b | 0.32 ± 0.17 b | ||
| Low | 0.59 ± 0.12 | 0.45 ± 0.26 | 0.64 ± 0.28 | ||
| Filtration Rate (mg h−1) | Ambient | 54.80 ± 11.14 a | 62.25 ± 19.75 a | 10.05 ± 4.55 b | p < 0.001 |
| Medium | 53.08 ± 39.89 a | 42.59 ± 10.40 a | 10.59 ± 5.54 b | ||
| Low | 32.42 ± 6.65 | 21.14 ± 11.96 | 11.00 ± 4.73 | ||
| Rejection Rate (mg h−1) | Ambient | 34.94 ± 7.94 a | 44.97± 14.77 a | 5.20 ± 2.86 b | p < 0.001 |
| Medium | 40.07 ± 31.26 a | 30.97 ± 8.32 a | 7.90 ± 4.86 b | ||
| Low | 19.22 ± 4.63 | 14.99 ± 8.56 | 6.07 ± 3.45 | ||
| Organic Ingestion Rate (mg h−1) | Ambient | 16.24 ± 3.52 a | 13.24 ± 5.87 a | 4.29 ± 2.01 b | p < 0.05 |
| Medium | 9.13 ± 7.19 a | 8.28 ± 2.84 a | 1.85 ± 1.38 b | ||
| Low | 10.99 ± 2.40 a | 4.76 ± 3.39 b | 4.32 ± 2.47 b | ||
| Absorption Rate (mg h−1) | Ambient | 13.42 ± 3.53 a | 11.93 ± 5.56 a | 3.80 ± 1.88 b | p < 0.05 |
| Medium | 8.00 ± 6.56 a | 7.04 ± 2.53 a | 1.58 ± 1.35 b | ||
| Low | 9.67 ± 2.27 a | 4.02 ± 2.65 b | 3.91 ± 2.31 b | ||
| Egestion Rate (mg h−1) | Ambient | 17.22 ± 3.35 a | 16.30 ± 6.33 a | 4.54 ± 1.83 b | p < 0.01 |
| Medium | 10.34 ± 7.45 a | 10.72 ± 3.22 a | 2.61 ± 1.23 b | ||
| Low | 12.40 ± 2.49 a | 5.18 ± 3.39 a,b | 3.65 ± 2.05 b | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steeves, L.; Honecker, M.; Meseck, S.L.; Munroe, D. Effects of Ocean Acidification and Summer Thermal Stress on the Physiology and Growth of the Atlantic Surfclam (Spisula solidissima). J. Mar. Sci. Eng. 2024, 12, 673. https://doi.org/10.3390/jmse12040673
Steeves L, Honecker M, Meseck SL, Munroe D. Effects of Ocean Acidification and Summer Thermal Stress on the Physiology and Growth of the Atlantic Surfclam (Spisula solidissima). Journal of Marine Science and Engineering. 2024; 12(4):673. https://doi.org/10.3390/jmse12040673
Chicago/Turabian StyleSteeves, Laura, Molly Honecker, Shannon L. Meseck, and Daphne Munroe. 2024. "Effects of Ocean Acidification and Summer Thermal Stress on the Physiology and Growth of the Atlantic Surfclam (Spisula solidissima)" Journal of Marine Science and Engineering 12, no. 4: 673. https://doi.org/10.3390/jmse12040673