“Firefly” of the Submarine Mountains: New Data on Physiculus cynodon (Moridae, Teleostei) from Emperor Seamounts and Northwestern Hawaiian Ridge

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
Analysis of Biological Data
3. Results
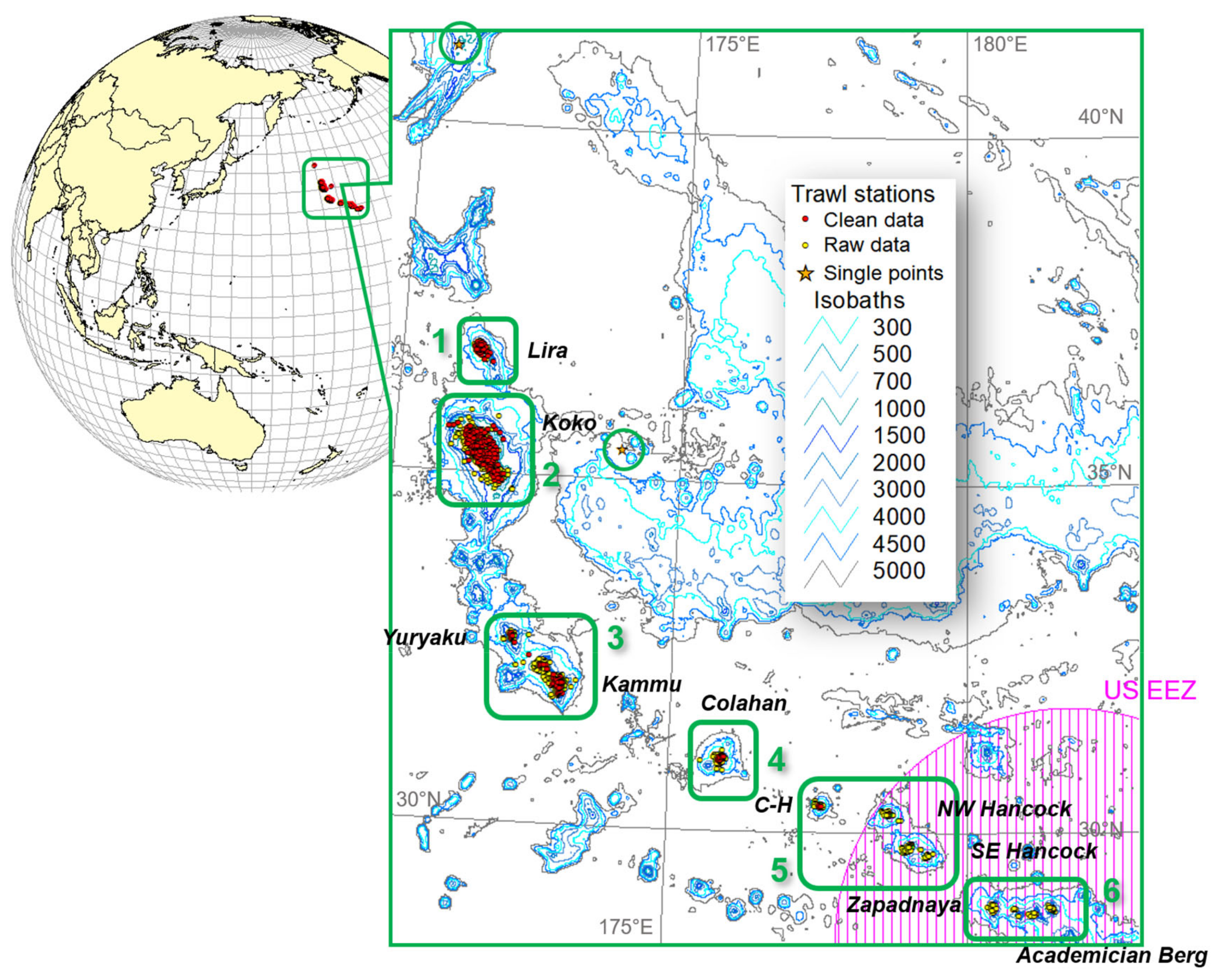
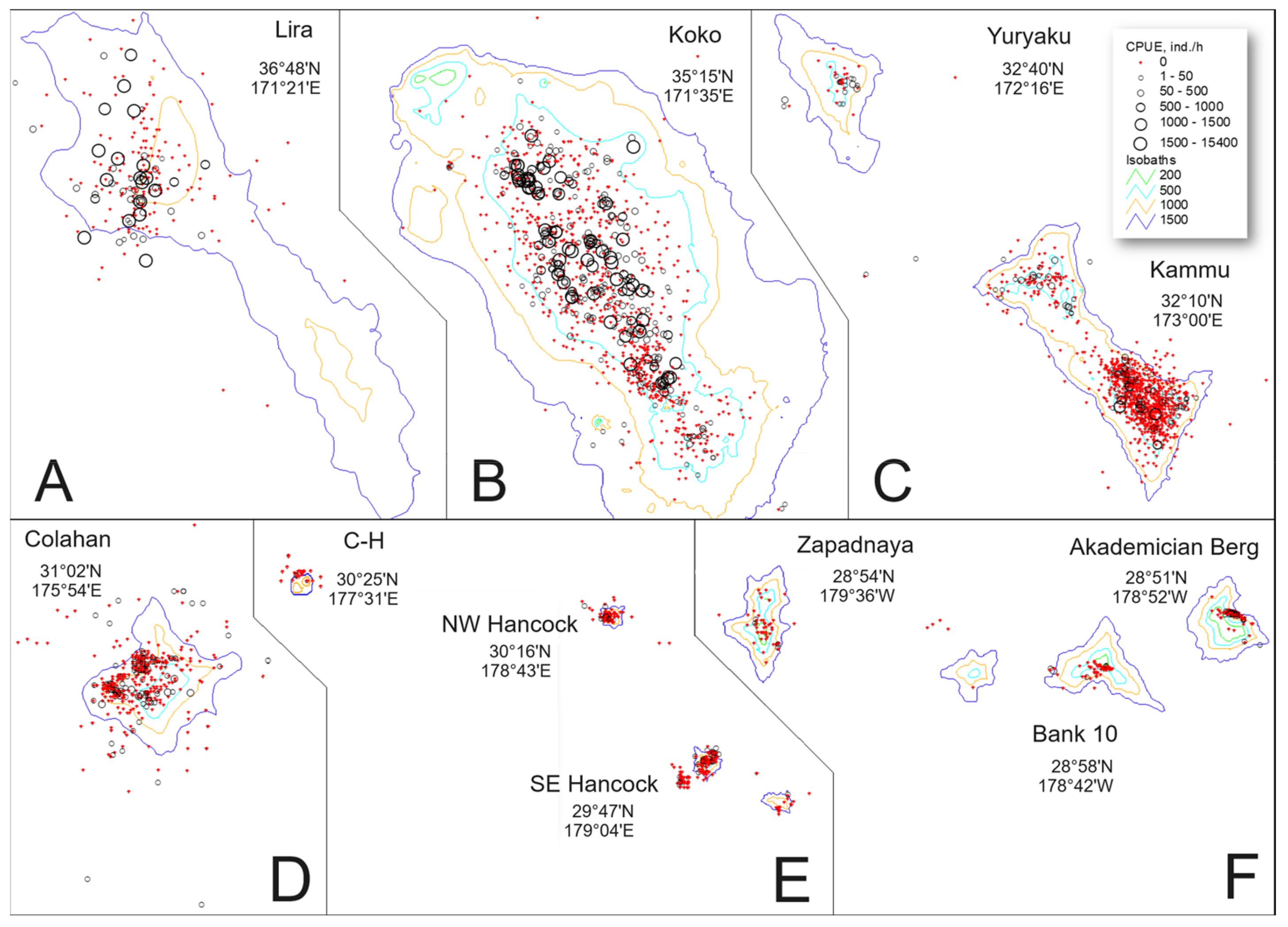
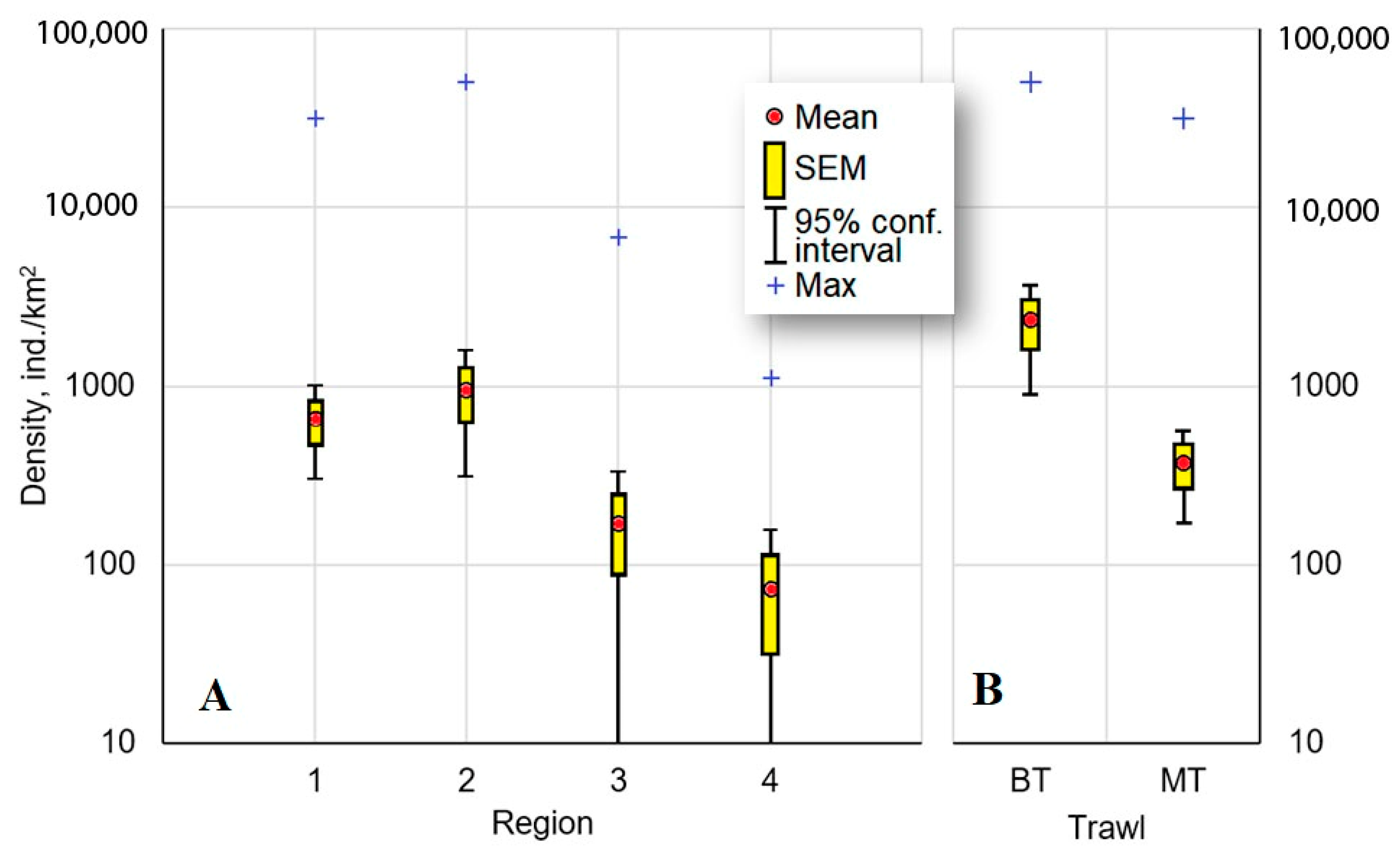
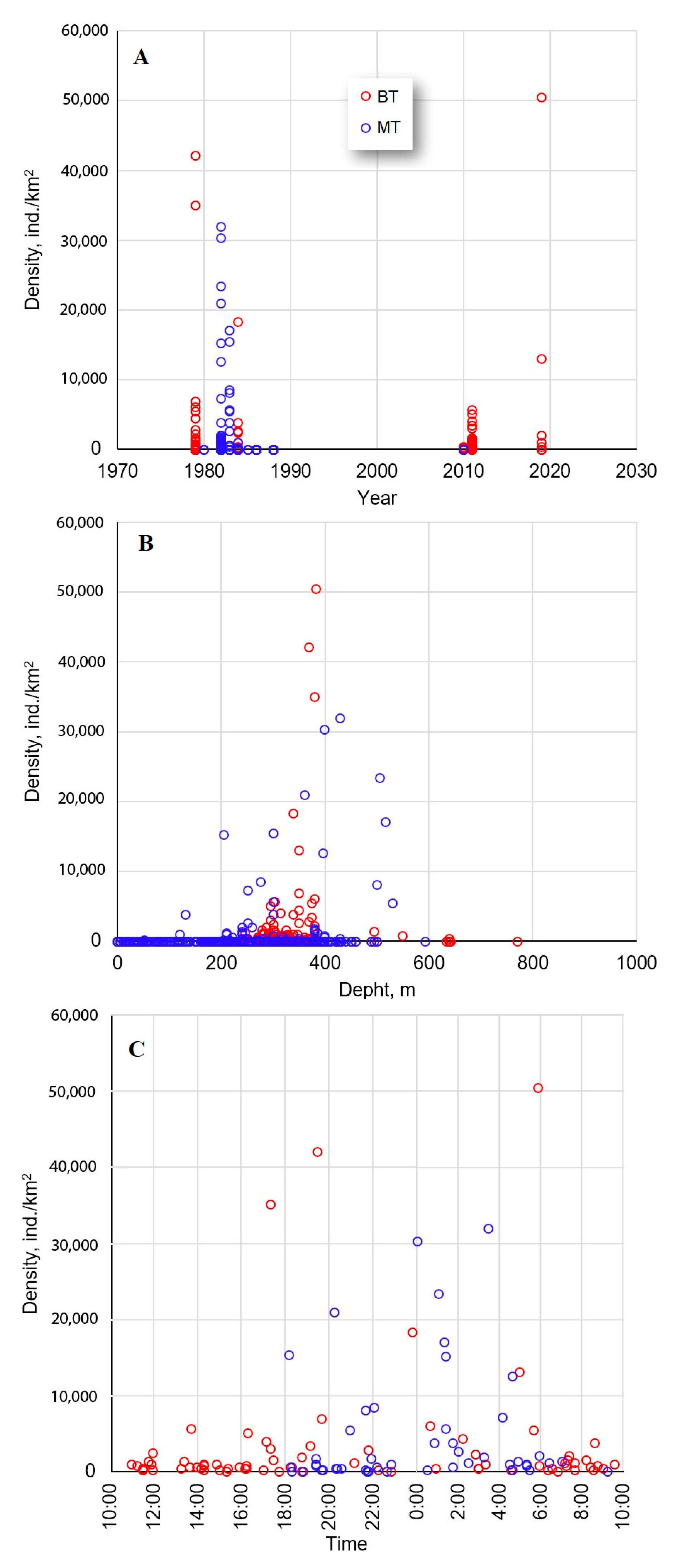
3.1. Catch Rate and Distribution
3.2. Biological Features
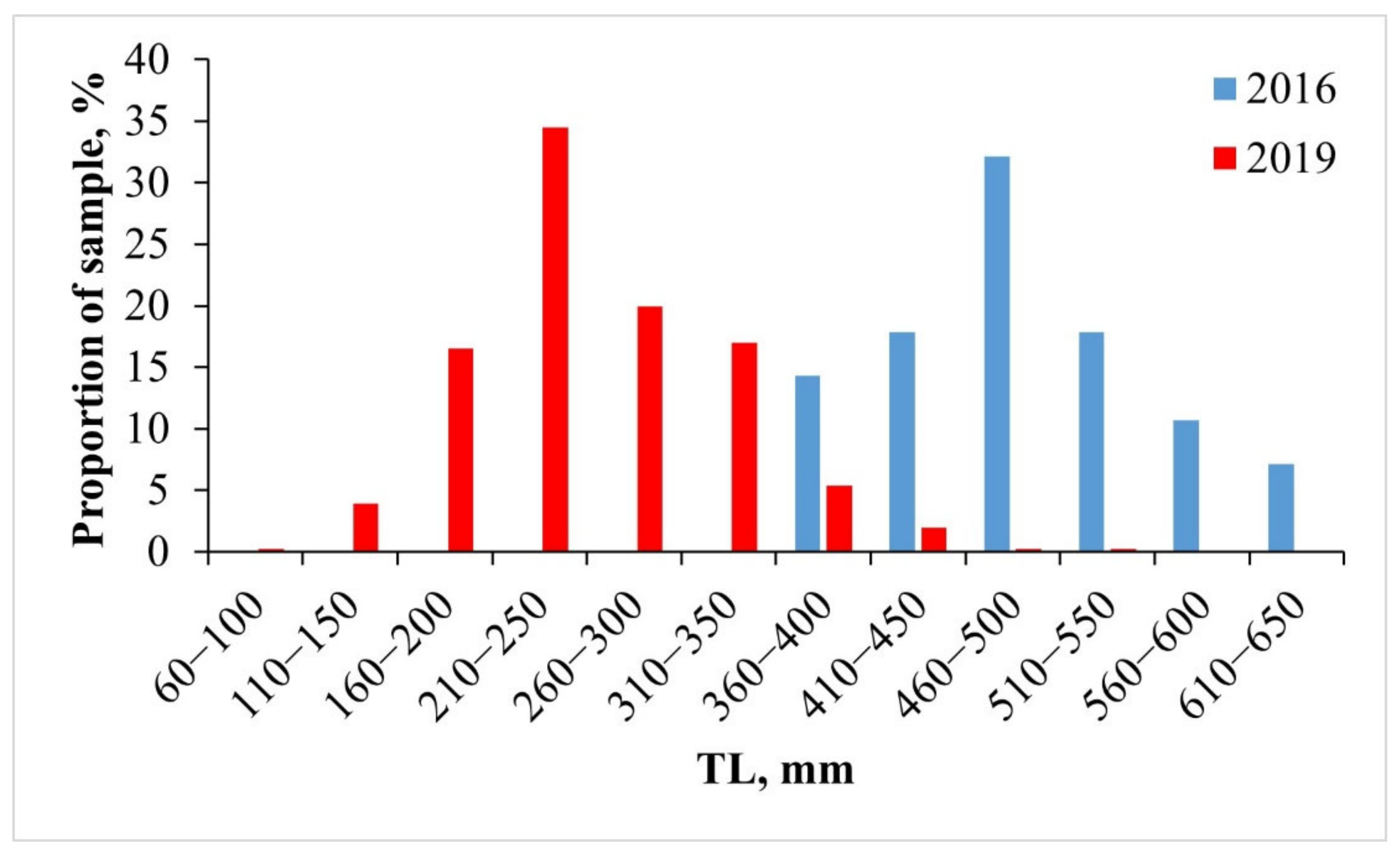
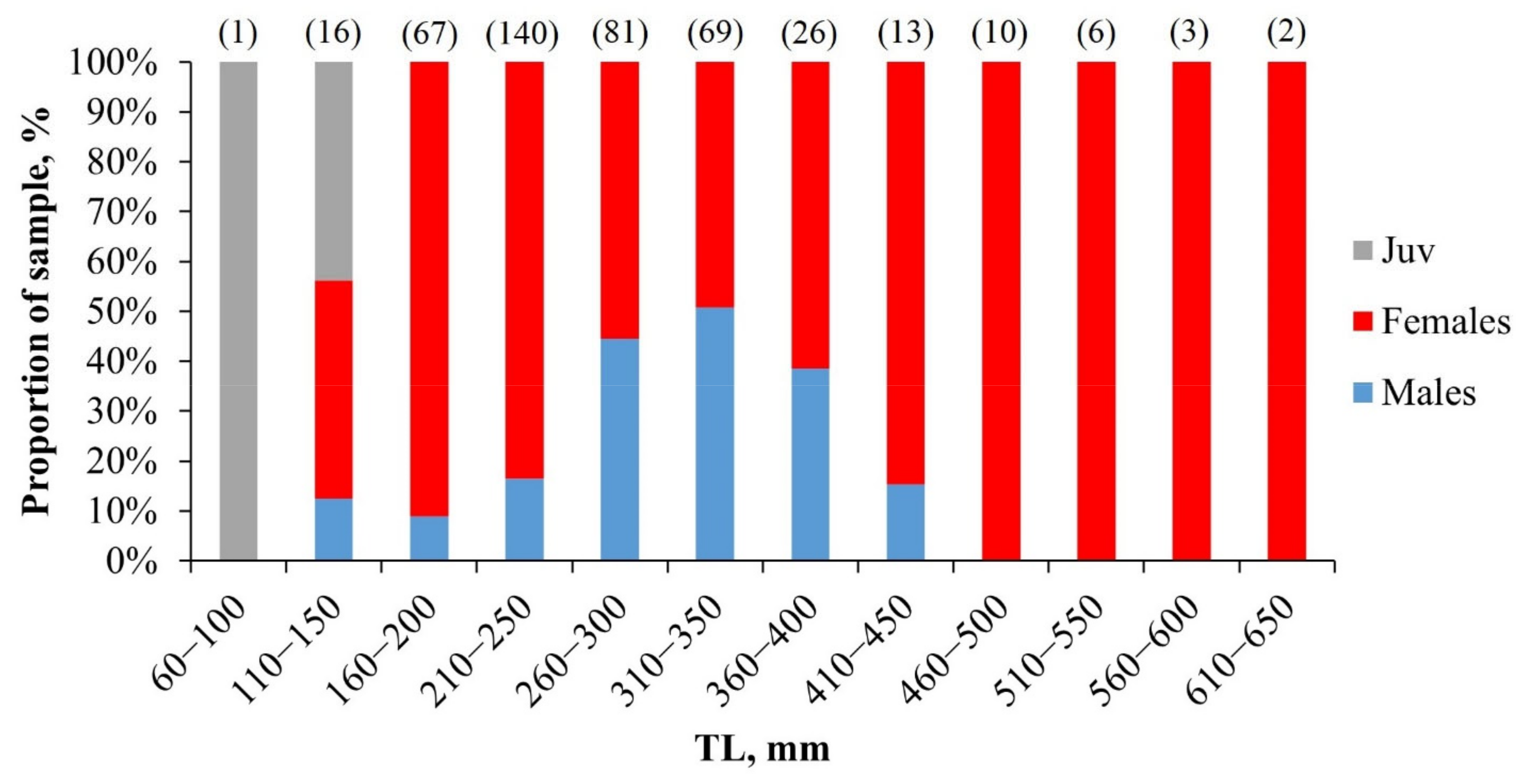
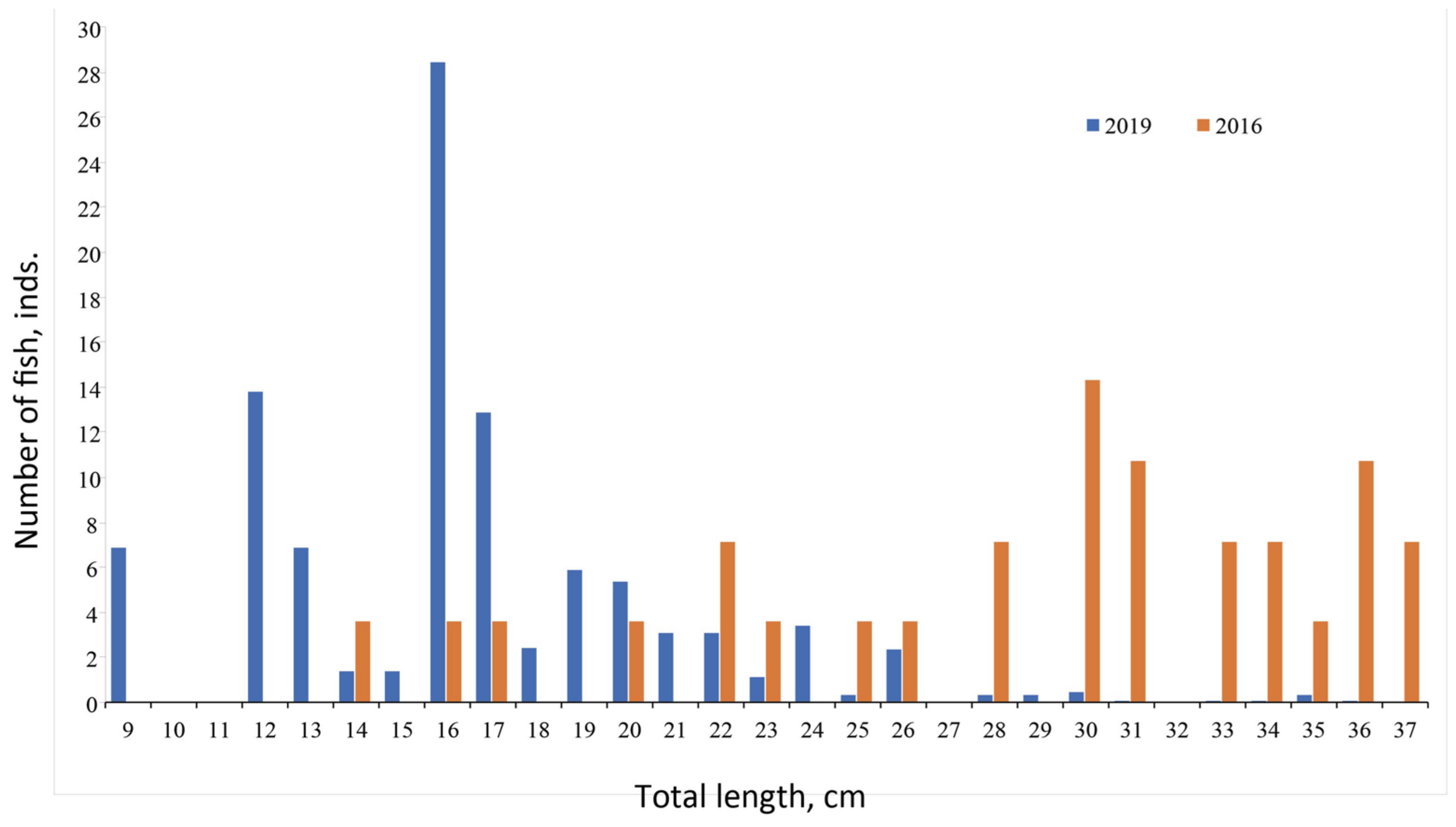
3.2.1. Size Composition of Catches
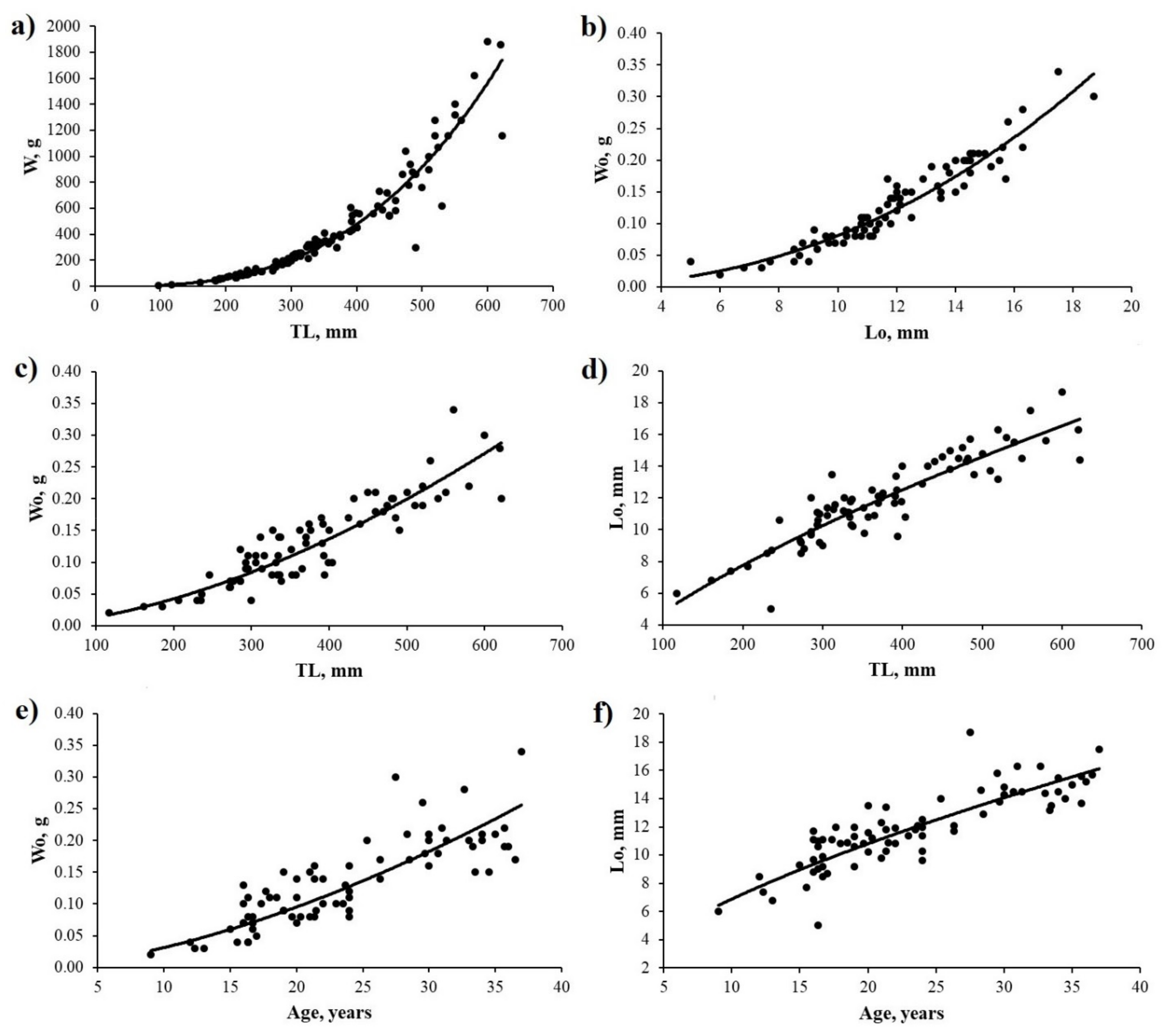
3.2.2. Relationships between Body and Otolith Sizes
4. Discussion
4.1. Catch Rate and Distribution
4.2. Biological Features
4.2.1. Size Composition
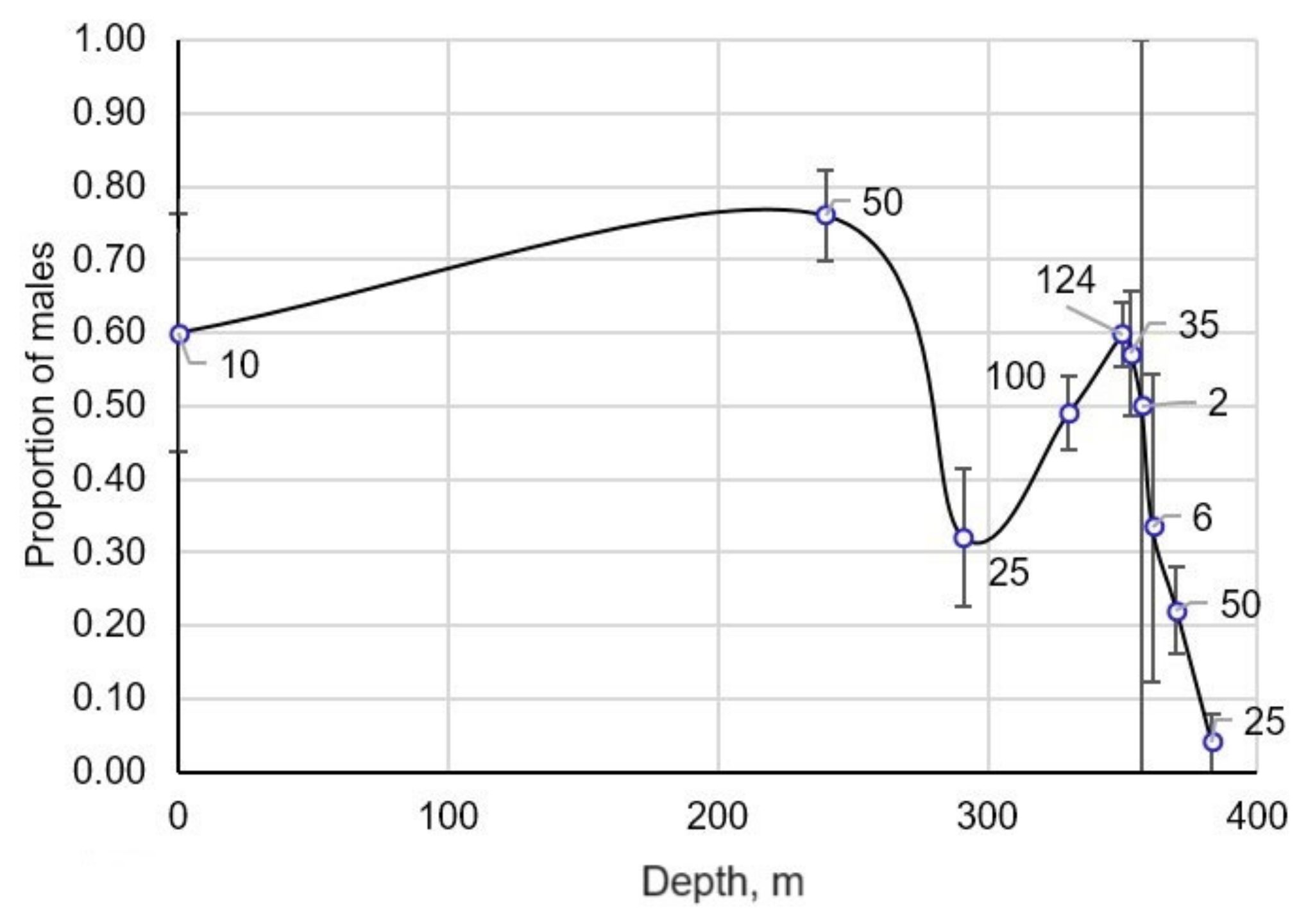
4.2.2. Sex Ratio
4.2.3. Size-Related Relationships
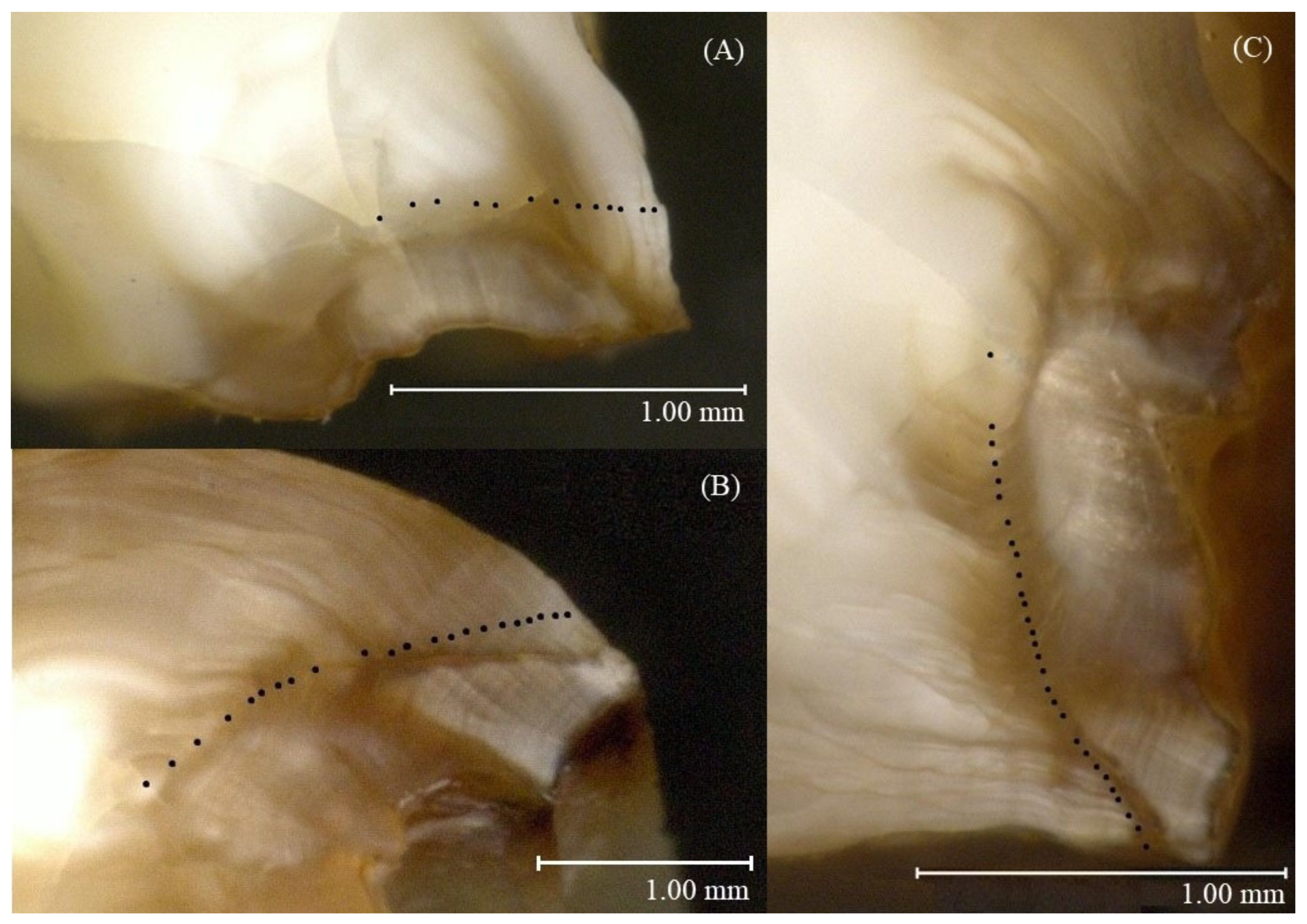
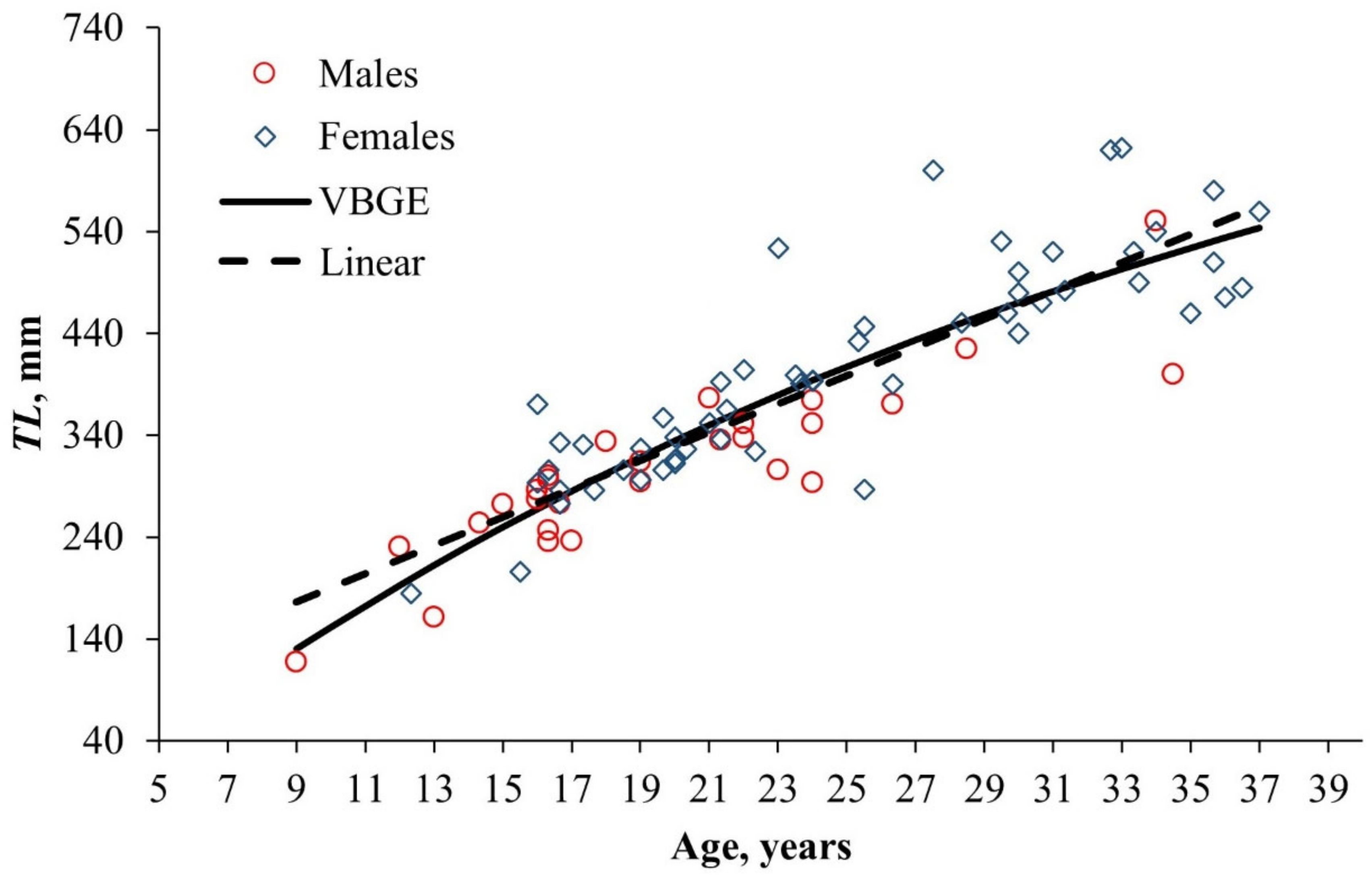
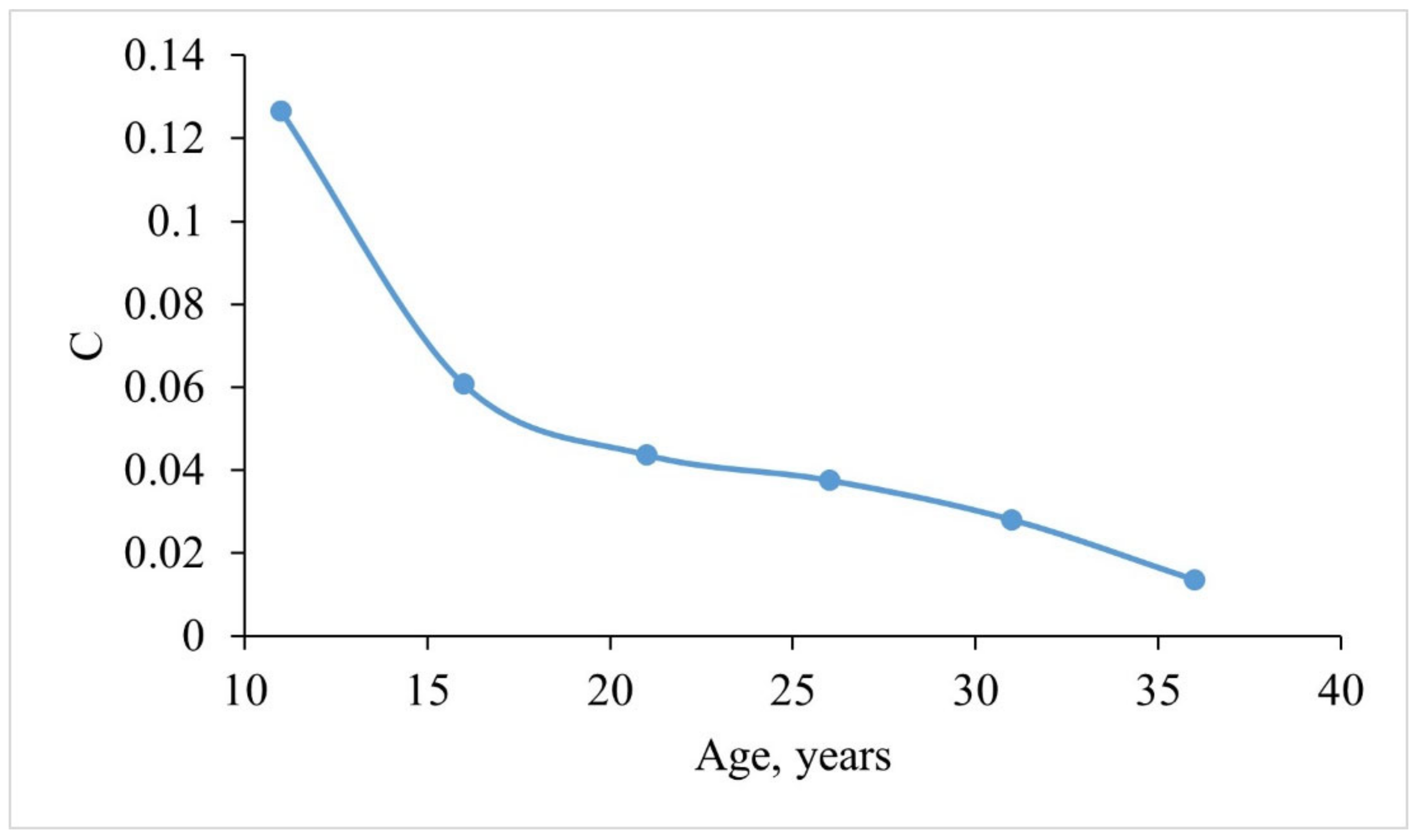
4.2.4. Age and Growth
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Genera/Species by Family/Subfamily. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 26 September 2023).
- Paulin, C.D. Review of the Morid Genera Gadella, Physiculus, and Salilota (Teleostei: Gadiformes) with Descriptions of Seven New Species. N. Z. J. Zool. 1989, 16, 93–133. [Google Scholar] [CrossRef]
- Paulin, C.D.; Roberts, C.D. Review of the Morid Cods (Teleostei, Paracanthopterygii, Moridae) of New Caledonia, with Description of a New Species of Gadella. Résult. Camp. Musorstom 1997, 17, 17–41. [Google Scholar]
- Tang, C.N. Description of a New Codling Species of Physiculus from Taiwan (Gadiformes: Moridae). Zootaxa 2021, 5052, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Idrees Babu, K.K.; Ho, H.C.; Mariyambi, P.C.; Sureshkumar, S. Two New Species of the Codling Fish Genus Physiculus from Lakshadweep, India (Gadiformes: Moridae). Zootaxa 2022, 5104, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Froese, R.; Pauly, D. Fishbase. World Wide Web Electronic Publication. 2023. Available online: https://www.fishbase.org (accessed on 23 August 2023).
- Okamoto, M.; Sato, N.; Watanabe, Y. Pelagic Juveniles of Two Morids (Teleostei: Gadiformes: Moridae), Antimora microlepis and Physiculus japonicus, from the Western North Pacific. Species Divers. 2007, 12, 17–27. [Google Scholar] [CrossRef]
- Kitagawa, D.; Nagahora, S. Estimation of the Spawning Season of the Morid Fish Physiculus maximowiczi Collected from the Coastal Waters of Iwate Prefecture, Japan. Bull. Jpn. Soc. Sci. Fish. 1983, 49, 1649–1654. [Google Scholar] [CrossRef]
- Borets, L.A. Ichthyofauna of Northwestern and Hawaiian Underwater Ridges. Vopr. Ikhtiologii 1986, 26, 208–220. [Google Scholar]
- Watabe, S.; Ikeda, D.; Mashiro, T.; Kagetakubo, Y.; Takahashi, Y.; Uemura, M.; Mizusawa, N.; Koyama, H.; Yasumoto, K.; Jimbo, M.; et al. Suitability of Japanese Codling as a Raw Material for Surimi-Based Products Revealed by Primary Sequence Analysis of Myosin Heavy Chain and Thermal Gel Properties. Fish. Sci. 2020, 86, 711–719. [Google Scholar] [CrossRef]
- Cohen, D.M.; Inada, T.; Iwamoto, T.; Scialabba, N. FAO Species Catalogue; FAO: Rome, Italy, 1990; Volume 1, ISBN 9251028907. [Google Scholar]
- Koh, J.-R.; Moon, D.-Y. First Record of Japanese Codling, Physiculus japonica Hilgendorf (Moridae, Gadiformes) from Korea. Fish. Aquat. Sci. 2003, 6, 97–100. [Google Scholar] [CrossRef]
- Yu, Y.; Ho, H.-C. Review of Codfish Family Moridae (Teleostei: Gadiformes) from Taiwan. Platax 2012, 9, 33–59. [Google Scholar] [CrossRef]
- Jang, S.H.; Kim, J.-K.; Park, J.-H.; Song, Y.S. New Record of Gadella jordani and Redescription of Physiculus japonicus (Pisces: Moridae) in Korea. Anim. Syst. Evol. Divers. 2016, 32, 28–37. [Google Scholar] [CrossRef]
- Koeda, K.; Takashima, S.; Yamakita, T.; Tsuchida, S.; Fujiwara, Y. Deep-Sea Fish Fauna on the Seamounts of Southern Japan with Taxonomic Notes on the Observed Species. J. Mar. Sci. Eng. 2021, 9, 1294. [Google Scholar] [CrossRef]
- Gooding, R.M. Exploratory Fishing on the Hancock Seamounts by the Townsend Cromwell, 1976–1979. In Proceedings of the Symposium on Status of Resource Investigations in the Northwestern Hawaiian Islands, Honolulu, HI, USA, 25–27 May 1980; pp. 241–250. [Google Scholar]
- Uchida, R.; Tagami, D. Groundfish Fisheries and Research in the Vicinity of Seamounts in the North Pacific Ocean. Mar. Fish. Rev. 1984, 46, 1–17. [Google Scholar]
- Mejía-Mercado, B.E.; Mundy, B.; Baco, A.R. Variation in the Structure of the Deep-Sea Fish Assemblages on Necker Island, Northwestern Hawaiian Islands. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2019, 152, 103086. [Google Scholar] [CrossRef]
- Baytalyuk, A.A.; Karyakin, K.A.; Orlov, A.M. Thalassobathyal Resources of the Emperor: Fishery, Stock Status and Possibility for Distant Fishery. Probl. Fish. 2010, 11, 801–816. [Google Scholar]
- Somov, A.A.; Kanzeparova, A.N.; Vazhova, A.S.; Khleborodov, A.S.; Zuev, M.A.; Slabinsky, A.M.; Orlova, S.Y.; Kurnosov, D.S.; Balyaev, V.A.; Orlov, A.M. Some Preliminary Results of Research on Emperor Seamounts in April 2019. Tr. VNIRO 2019, 175, 208–219. [Google Scholar] [CrossRef]
- Sazonov, Y.I. Physiculus cynodon sp. n.(Gadiformes, Moridae) from Submarine Mountains in the Northern Part of Pacific Ocean. Zool. Zhurnal 1986, 65, 305–308. [Google Scholar]
- Prokofiev, A.M. New Findings of Rare Fish Species in Indian and Pacific Oceans with the Description of Two New Species from the Families Gobiidae and Platycephalidae. J. Ichthyol. 2017, 57, 803–820. [Google Scholar] [CrossRef]
- Volvenko, I.V.; Orlov, A.M.; Gebruk, A.V.; Katugin, O.N.; Vinogradov, G.M.; Maznikova, O.A. Species Richness and Taxonomic Composition of Trawl Macrofauna of the North Pacific and Its Adjacent Seas. Sci. Rep. 2018, 8, 16604. [Google Scholar] [CrossRef]
- Volvenko, I.V.; Gebruk, A.V.; Katugin, O.N.; Ogorodnikova, A.A.; Vinogradov, G.M.; Maznikova, O.A.; Orlov, A.M. Commercial Value of Trawl Macrofauna of the North Pacific and Adjacent Seas. Environ. Rev. 2020, 28, 269–283. [Google Scholar] [CrossRef]
- Kitagawa, D.; Kuroda, K.; Tsuruta, Y. Description and Distribution of Eggs and Larvae of the Brown Hakeling Physiculus maximowiczi in Japanese Waters. Bull. Jpn. Soc. Sci. Fish. 1985, 51, 1627–1630. [Google Scholar] [CrossRef]
- Fujita, T.; Kitagawa, D.; Okuyama, Y.; Ishito, Y.; Inada, T.; Jin, Y. Diets of the Demersal Fishes on the Shelf off Iwate, Northern Japan. Mar. Biol. 1995, 123, 219–233. [Google Scholar] [CrossRef]
- Aguzzi, J.; Fanelli, E.; Ciuffardi, T.; Schirone, A.; De Leo, F.C.; Doya, C.; Kawato, M.; Miyazaki, M.; Furushima, Y.; Costa, C.; et al. Faunal Activity Rhythms Influencing Early Community Succession of an Implanted Whale Carcass Offshore Sagami Bay, Japan. Sci. Rep. 2018, 8, 11163. [Google Scholar] [CrossRef]
- Haneda, Y. The Luminescence of Some Deep-Sea Fishes of the Families Gadidae and Macrouridae. Pac. Sci. 1951, 5, 372–378. [Google Scholar]
- Makiguchi, N.; Arita, M.; Asai, Y. Optimum Cultural Conditions for Strong Light Production by Photobacterium phosphoreum. J. General. Appl. Microbiol. 1980, 26, 75–84. [Google Scholar] [CrossRef]
- Kuwae, T.; Andoh, M.; Fukasawa, S.; Kurata, M. Relationship between Luminous Fish and Symbiosis: I. Comparative Studies of Lipopolysaccharides Isolated from Symbiotic Luminous Bacteria of the Luminous Marine Fish, Physiculus japonicus. Microbiol. Immunol. 1983, 27, 847–859. [Google Scholar] [CrossRef]
- Kuwae, T.; Andoh, M.; Kurata, M. Relationship between Luminous Fish and Symbiosis II. Chemical Composition of Lipopolysaccharides Isolated from Symbiotic Luminous Bacteria in the Luminous Marine Fish, Physiculus japonicus. Microbiol. Immunol. 1986, 30, 75–80. [Google Scholar] [CrossRef]
- Pires, A.; Carvalho-Filho, A.; Ferreira, R.; Viana, D.; Nunes, D.; Hazin, F. Review of the Brazilian species of Physiculus (Gadiformes: Moridae), with description of a new species from Saint Peter and Saint Paul Archipelago, equatorial Atlantic. Zootaxa 2019, 4671, 67–80. [Google Scholar] [CrossRef]
- Bensch, A.; Gianni, M.; Sanders, J.; Hjort, A.; Bensch, A.; Gréboval, D. Worldwide Review of Bottom Fisheries in the High Seas: South West Atlantic Ocean; FAO Fisheries and Aquaculture Technical Paper; FAO: Rome, Italy, 2009; Volume 522, 145p. [Google Scholar]
- Volvenko, I.V. The Role of the Regional Data Center (RDC) of the Pacific Research Fisheries Center (TINRO-Center) in North Pacific Ecosystem and Fisheries Research. Int. J. Eng. Res. Sci. (IJOER) 2015, 1, 47–54. [Google Scholar]
- Volvenko, I.V. The Concept of Information Support for Bioresource and Ecosystem Research in the North-West Pacific: Theory and Practical Implementation. Nat. Resour. 2016, 7, 40–50. [Google Scholar] [CrossRef]
- Volvenko, I.V.; Kulik, V.V. Updated and Extended Database of the Pelagic Trawl Surveys in the Far Eastern Seas and North Pacific Ocean in 1979–2009. Russ. J. Mar. Biol. 2011, 37, 513–532. [Google Scholar] [CrossRef]
- Volvenko, I.V. The New Large Database of the Russian Bottom Trawl Surveys in the Far Eastern Seas and the North Pacific Ocean in 1977–2010. Int. J. Environ. Monit. Anal. 2014, 2, 302–312. [Google Scholar] [CrossRef]
- Laevastu, T. Manual of Methods in Fisheries Biology; FAO: Rome, Italy, 1965. [Google Scholar]
- Pravdin, I.F. Guidelines to Studies of Fishes; Pishchevaya Promyshlennost: Moscow, USSR, 1966. [Google Scholar]
- Beamish, R.J.; McFarlane, G.A. Current Trends in Age Determination Methodology. In Age and Growth of Fish; Summerfelt, R.C., Hall, G.E., Eds.; Iowa State University Press: Ames, IA, USA, 1987; pp. 15–42. [Google Scholar]
- Horn, P.L.; Sutton, C.P. An Assessment of Age and Growth of Violet Cod (Antimora rostrata) in the Ross Sea, Antarctica. Polar Biol. 2015, 38, 1553–1558. [Google Scholar] [CrossRef]
- Frey, P.H.; Keller, A.A.; Simon, V. Dynamic Population Trends Observed in the Deep-Living Pacific Flatnose, Antimora microlepis, on the U.S. West Coast. Deep. Sea Res. Part. I Oceanogr. Res. Pap. 2017, 122, 105–112. [Google Scholar] [CrossRef]
- Orlov, A.M.; Vedishcheva, E.V.; Trofimova, A.O.; Orlova, S.Y. Age and Growth of Blue Antimora Antimora rostrata (Moridae) in Southwestern Greenland Waters. J. Ichthyol. 2018, 58, 217–225. [Google Scholar] [CrossRef]
- Vedishcheva, E.V.; Korostelev, N.B.; Gordeev, I.I.; Orlov, A.M. A First Attempt to Evaluate the Age and Growth of Blue Hake Antimora rostrata (Moridae, Gadiformes, Teleostei) from the Lazarev and Weddell Seas (Antarctic). Polar Rec. 2019, 55, 25–31. [Google Scholar] [CrossRef]
- Korostelev, N.B.; Vedishcheva, E.V.; Orlov, A.M. Age and Growth of Antimora rostrata (Moridae, Gadiformes, Teleostei) from the Kerguelen and Crozet Islands in the Southern Indian Ocean. Polar Rec. 2019, 55, 452–459. [Google Scholar] [CrossRef]
- Korostelev, N.B.; Frey, P.H.; Orlov, A.M. Using Different Hard Structures to Estimate the Age of Deep-Sea Fishes: A Case Study of the Pacific Flatnose, Antimora microlepis (Moridae, Gadiformes, Teleostei). Fish. Res. 2020, 232, 105731. [Google Scholar] [CrossRef]
- Korostelev, N.B.; Baytalyuk, A.A.; Maltsev, I.V.; Orlov, A.M. First Data on The Age and Growth in Pacific Flatnose Antimora microlepis (Moridae) from the Waters of the Underwater Emperor Mountain Range (Northwestern Pacific). J. Ichthyol. 2020, 60, 891–899. [Google Scholar] [CrossRef]
- Korostelev, N.B.; Welsford, D.C.; Belyakov, V.V.; Bush, A.G.; Orlov, A.M. So Close Yet So Far: Age and Growth of Blue Antimora Antimora rostrata (Moridae, Gadiformes, Teleostei) off New Zealand and Macquarie Island (Southwestern Pacific Ocean). J. Mar. Sci. Eng. 2022, 10, 956. [Google Scholar] [CrossRef]
- Korostelev, N.B.; Maltsev, I.V.; Orlov, A.M. First Data on the Age and Growth of Schmidt’s Cod Lepidion Schmidti (Moridae) from Waters of the Emperor Seamounts (Northwestern Pacific). J. Mar. Sci. Eng. 2023, 11, 1212. [Google Scholar] [CrossRef]
- Beamish, R.J.; Fournier, D.A. A Method for Comparing the Precision of a Set of Age Determinations. Can. J. Fish. Aquat. Sci. 1981, 38, 982–983. [Google Scholar] [CrossRef]
- Arkhipkin, A.I.; Baumgartner, N.; Brickle, P.; Laptikhovsky, V.V.; Pompert, J.H.W.; Shcherbich, Z.N. Biology of the Skates Bathyraja brachyurops and B. griseocauda in Waters around the Falkland Islands, Southwest Atlantic. ICES J. Mar. Sci. 2008, 65, 560–570. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Maravelias, C.D. Modelling Fish Growth: Multi-Model Inference as a Better Alternative to a Priori Using von Bertalanffy Equation. Fish Fish. 2008, 9, 178–187. [Google Scholar] [CrossRef]
- Mina, M.V.; Klevezal, G.A. Growth of the Animals; Nauka: Moscow, USSR, 1976. [Google Scholar]
- Alimov, A.F. Introduction into Production Hydrobiology; Gidrometeoizdat: Leningrad, USSR, 1989. [Google Scholar]
- Hammer, Ø.; Harper, D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 178. [Google Scholar]
- Morozov, A.V. Methodology for Collecting and Processing Ichthyological Materials. Rybn. Khozyaistvo Turkm./Tr. NIIRH 1929, 5, 172–189. [Google Scholar]
- Bystrov, Y.N.; Mikhailovsky, A.P. (Eds.) Description of Seamounts and Underwater Elevations of the World Oceans Fishing Grounds (High Seas); Ministry of Defense of the USSR, Ministry of Fisheries of the USSR: Moscow, USSR, 1989; Volume 2.
- Novikov, N.P.; Kodolov, L.S.; Gavrilov, G.M. Preliminary List of Fishes of the Emperor Underwater Ridge. Fishes Open Ocean. 1981, 32–35. [Google Scholar]
- Miyamoto, M.; Kiyota, M.; Hayashibara, T.; Nonaka, M.; Imahara, Y.; Tachikawa, H. Megafaunal Composition of Cold-Water Corals and Other Deep-Sea Benthos in the Southern Emperor Seamounts Area, North Pacific Ocean. Galaxea J. Coral Reef. Stud. 2017, 19, 19–30. [Google Scholar] [CrossRef]
- Baco, A.R.; Morgan, N.B.; Roark, E.B. Observations of Vulnerable Marine Ecosystems and Significant Adverse Impacts on High Seas Seamounts of the Northwestern Hawaiian Ridge and Emperor Seamount Chain. Mar. Policy 2020, 115, 103834. [Google Scholar] [CrossRef]
- Morgan, N.B.; Baco, A.R. Recent Fishing Footprint of the High-Seas Bottom Trawl Fisheries on the Northwestern Hawaiian Ridge and Emperor Seamount Chain: A Finer-Scale Approach to a Large-Scale Issue. Ecol. Indic. 2021, 121, 107051. [Google Scholar] [CrossRef]
- Baco, A.R.; Morgan, N.B.; Brendan Roark, E.; Biede, V. Bottom-Contact Fisheries Disturbance and Signs of Recovery of Precious Corals in the Northwestern Hawaiian Islands and Emperor Seamount Chain. Ecol. Indic. 2023, 148, 110010. [Google Scholar] [CrossRef]
- Hoff, G.R.; Stevenson, D.E.; Orr, J.W. Guide to the Gadiform Fishes of the Eastern North Pacific; NOAA Technical Memorandum NMFS-AFSC; National Oceanic and Atmospheric Administration, U.S. Department of Commerce: Washington, DC, USA, 2015; Volume 309, 68p. [CrossRef]
- Borets, L.A.; Kulikov, M.Y. Thalassobathyal. In Biological Resources of the Pacific Ocean; Nauka: Moscow, USSR, 1986; pp. 505–520. [Google Scholar]
- Orlov, A.M. Trophic Relationships of Commercial Fishes in the Pacific Waters off Southeastern Kamchatka and the Northern Kuril Islands. In Proceedings of the Ecosystem Approaches for Fishery Management, Anchorage, AK, USA, 30 October–3 November 1998; University of Alaska: Fairbanks, AK, USA, 1999; pp. 231–263. [Google Scholar]
- Orlov, A.M. Trophic Interrelations in Predatory Fishes of Pacific Waters Circumambient the Northern Kuril Islands and Southeastern Kamchatka. Hydrobiol. J. 2004, 40, 106–124. [Google Scholar] [CrossRef]
- Savinykh, V.F.; Baytalyuk, A.A. New Data on Biology of the Pearlfish Maurolicus imperatorius (Sternopthychidae) from the Emperor Seamount Chain. J. Ichthyol. 2010, 50, 148–158. [Google Scholar] [CrossRef]
- Orlov, A.M. Mesopelagic Fishes as Prey of Atka Mackerel (Pleurogrammus monopterygius, Hexagrammidae, Scorpaeniformes) off the Northern Kuril Islands. In Proceedings of the Forage Fishes in Marine Ecosystems, Anchorage, AK, USA, 13–16 November 1997; University of Alaska: Fairbanks, AK, USA, 1997; pp. 323–335. [Google Scholar]
- Orlov, A.M. On the Feeding of Atka Mackerel Pleurogrammus monopterygius in the Pacific Waters of the Northern Kuril Islands. J. Ichthyol. 1997, 37, 226–231. [Google Scholar]
- Wenner, C.A.; Musick, J.A. Biology of the Morid Fish Antimora rostrata in the Western North Atlantic. J. Fish. Res. Board. Can. 1977, 34, 2362–2368. [Google Scholar] [CrossRef]
- González, J.A.; Triay-Portella, R.; Correia, S.; Martins, A.; González-Lorenzo, G.; Lorenzo, J.M.; Pajuelo, J.G. Length–Weight Relationships of Five Selected Demersal Fishes from the Cabo Verde Islands (Eastern-Central Atlantic). J. Appl. Ichthyol. 2021, 37, 350–353. [Google Scholar] [CrossRef]
- Froese, R.; Thorson, J.T.; Reyes, R.B. A Bayesian Approach for Estimating Length-Weight Relationships in Fishes. J. Appl. Ichthyol. 2014, 30, 78–85. [Google Scholar] [CrossRef]
- Triay-Portella, R.; González, J.A.; Pajuelo, J.G. Caudal Region Regeneration in a Natural Population of the Morid Fish Physiculus cyanostrophus in the Tropical Eastern Atlantic Ocean. Deep. Sea Res. Part. I Oceanogr. Res. Pap. 2019, 150, 103062. [Google Scholar] [CrossRef]
- Parker, R.R.; Larkin, P.A. A Concept of Growth in Fishes. J. Fish. Res. Board. Can. 1959, 16, 721–745. [Google Scholar] [CrossRef]
- Paloheimo, J.E.; Dickie, L.M. Food and Growth of Fishes.: I. A Growth Curve Derived from Experimental Data. J. Fish. Res. Board. Can. 1965, 22, 521–542. [Google Scholar] [CrossRef]
- Ursin, E. A Mathematical Model of Some Aspects of Fish Growth, Respiration, and Mortality. J. Fish. Res. Board. Can. 1967, 24, 2355–2453. [Google Scholar] [CrossRef]
- Roff, D.A. A Motion for the Retirement of the Von Bertalanffy Function. Can. J. Fish. Aquat. Sci. 1980, 37, 127–129. [Google Scholar] [CrossRef]
- Schnute, J. A Versatile Growth Model with Statistically Stable Parameters. Can. J. Fish. Aquat. Sci. 1981, 38, 1128–1140. [Google Scholar] [CrossRef]
- Froese, R. Cube Law, Condition Factor and Weight-Length Relationships: History, Meta-Analysis and Recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Von Bertalanffy, L. A Quantitative Theory of Organic Growth (Inquiries on Growth Laws. II). Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Cailliet, G.M.; Andrews, A.H.; Burton, E.J.; Watters, D.L.; Kline, D.E.; Ferry-Graham, L.A. Age Determination and Validation Studies of Marine Fishes: Do Deep-Dwellers Live Longer? Exp. Gerontol. 2001, 36, 739–764. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Raw Data, 58 Cruises | Clean Data, 19 Cruises | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Years | Depth Range, m | BT | MT | Total | Years | Depth Range, m | BT | MT | Total | |
| 1 | 1971–2019 | 0–770 | 21 | 188 | 209 | 1979–2019 | 0–770 | 6 | 349 | 355 |
| 2 | 1969–2019 | 0–1500 | 659 | 457 | 1116 | 1979–2019 | 0–550 | 69 | 172 | 241 |
| 3 | 1969–2013 | 0–800 | 324 | 788 | 1112 | 1979–2011 | 5–459 | 13 | 92 | 105 |
| 4 | 1969–2013 | 0–2000 | 655 | 45 | 700 | 1979–1982 | 40–500 | 19 | 11 | 30 |
| 5 | 1969–1976 | 0–999 | 498 | 76 | 574 | 1982–1982 | 120–260 | 0 | 5 | 5 |
| 6 | 1968–1976 | 0–700 | 79 | 158 | 237 | - | - | 0 | 0 | 0 |
| Total | 1968–2019 | 0–2000 | 2236 | 1712 | 3948 | 1979–2019 | 0–770 | 107 | 629 | 736 |
| Area | Capture Depths, m | Highest CPUEs | Highest Mean CPUEs | ||||
|---|---|---|---|---|---|---|---|
| Min. | Max. | Mean | Depth Range, m | Inds. Caught per h | Depth Range, m | Inds. Caught per h | |
| 1 | 120 | 700 | 373.9 | 400–450 | 7013.3–7627.8 | 275–430 | 3320.0–4380.0 |
| 2 | 53 | 550 | 303.8 | 290–383 | 3258.0–4140.0 | 278–383 | 657.1–3528.0 |
| 3 | 285 | 500 | 373.6 | 330–385 | 384.0–432.0 | 354–392 | 192.0–232.0 |
| 4 | 250 | 900 | 345.4 | 275–280 | 248.6–466.7 | 224–350 | 20.3–28.8 |
| 5 | 260 | 400 | 296.8 | 270–300 | 72.0–96.0 | 270–295 | 2.5–8.0 |
| 6 | 190 | 280 | 240.0 | 195–220 | 34.3–109.1 | 195–220 | 15.6–34.3 |
| Function | AIC |
|---|---|
| Linear: TL = at + b | 2.0542 |
| Power: TL = atb + c | 2.0213 |
| VBGE: | 1.9664 |
| 1.9735 | |
| 1.9694 |
| Species | Habitat Depth, m | Age Max, Years | L∞ | k | t0 | R2 | Reference |
|---|---|---|---|---|---|---|---|
| A. microlepis | 175–3048 | 46 | 1311.0 | 0.016 | −0.90 | 0.7 | [47] |
| L. schmidti | 375–2404 | 49 | 2225.3 | 0.0007 | −6.2 | 0.7 | [49] |
| P. cynodon | 139–1007 | 37 | 858.6 | 0.030 | 3.5 | 0.8 | Present study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korostelev, N.B.; Volvenko, I.V.; Belyakov, V.V.; Baytaliuk, A.A.; Bush, A.G.; Kanzeparova, A.N.; Orlov, A.M. “Firefly” of the Submarine Mountains: New Data on Physiculus cynodon (Moridae, Teleostei) from Emperor Seamounts and Northwestern Hawaiian Ridge. J. Mar. Sci. Eng. 2023, 11, 2355. https://doi.org/10.3390/jmse11122355
Korostelev NB, Volvenko IV, Belyakov VV, Baytaliuk AA, Bush AG, Kanzeparova AN, Orlov AM. “Firefly” of the Submarine Mountains: New Data on Physiculus cynodon (Moridae, Teleostei) from Emperor Seamounts and Northwestern Hawaiian Ridge. Journal of Marine Science and Engineering. 2023; 11(12):2355. https://doi.org/10.3390/jmse11122355
Chicago/Turabian StyleKorostelev, Nikolai B., Igor V. Volvenko, Vladimir V. Belyakov, Aleksey A. Baytaliuk, Andrey G. Bush, Albina N. Kanzeparova, and Alexei M. Orlov. 2023. "“Firefly” of the Submarine Mountains: New Data on Physiculus cynodon (Moridae, Teleostei) from Emperor Seamounts and Northwestern Hawaiian Ridge" Journal of Marine Science and Engineering 11, no. 12: 2355. https://doi.org/10.3390/jmse11122355