
Towards an Integrated and Accurate Planktonic-Foraminiferal-Deduced Bio-Chrono-Stratigraphic Framework of Late Quaternary Mediterranean Marine Cores
 , , and
, , and 
Abstract
:1. Introduction
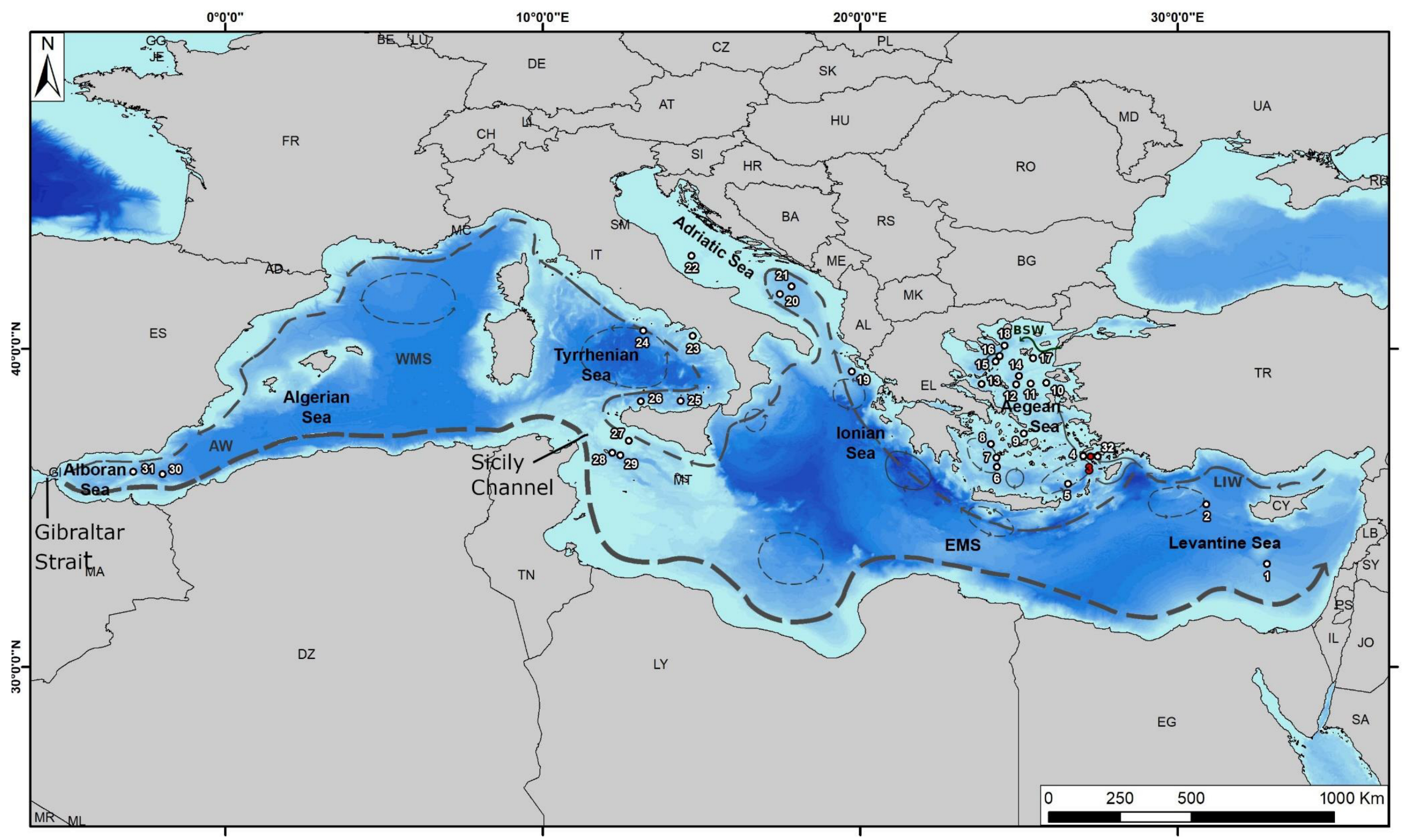
2. Regional Setting
3. Material and Methods
3.1. Study Core
3.2. Radiocarbon Analyses
3.3. Micropaleontological Analyses
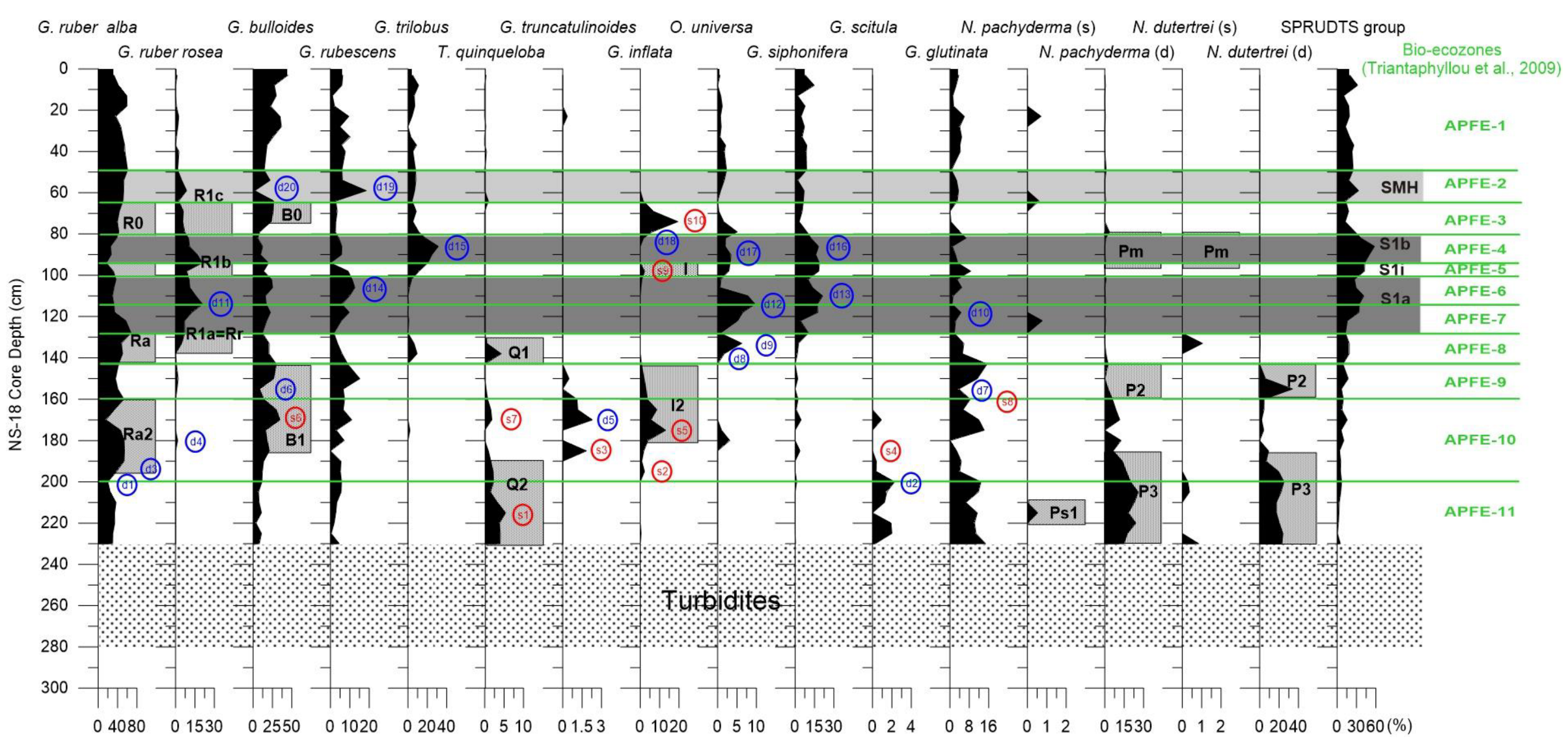
4. Results
4.1. Planktonic Foraminiferal Assemblages
4.2. Bioevent Stratigraphy

5. Discussion
5.1. High-Resolution Mediterranean Bio-Ecochronological Events Deduced from Planktonic Foraminiferal Assemblages
5.2. Planktonic Foraminifera Fluctuations along the Mediterranean Basin: Calibrating the Synchronous Bioevents
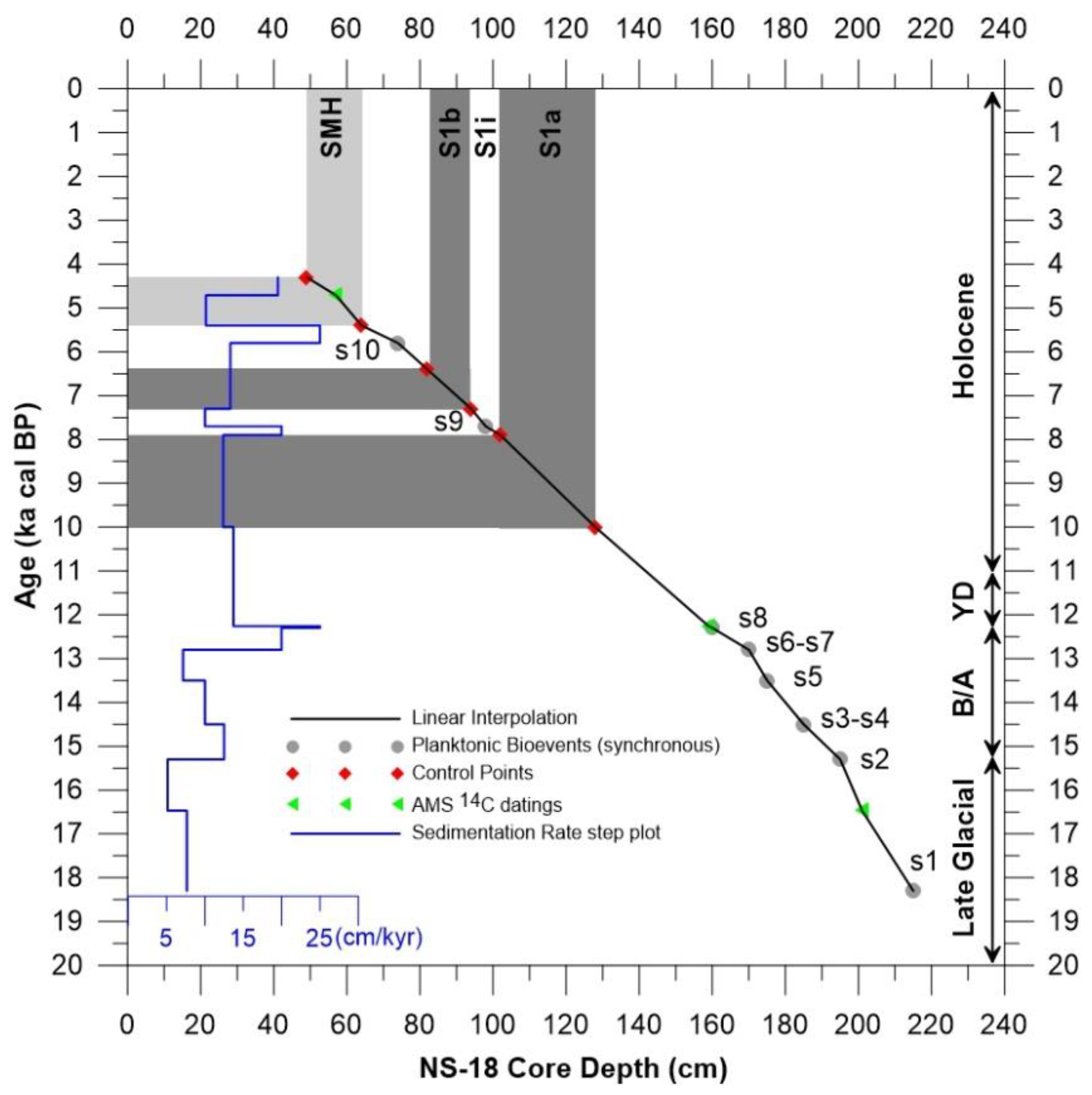
5.3. Integrated Age Model
5.4. Diachronous Bioevents for Improving Local Aegean Biostratigraphic Schemes and Their Paleoceanographic Implications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Label in Figure 1 | Core Name | Reference |
| 1 | MD84-641 | [46] |
| 2 | LC-31 | [44] |
| 3 | NS-18 | This study |
| 4 | NS-14 | [3,90] |
| 5 | LC-21 | [44] |
| 6 | KIM-2A | [5] |
| 7 | C69 | [6] |
| 8 | C40 | [7,44] |
| 9 | SL-9 | [44] |
| 10 | SL-11 | [44] |
| 11 | SL-21 | [44] |
| 12 | SL-31 | [44] |
| 13 | SK-1 | [9,44] |
| 14 | MNB-3 | [111] |
| 15 | AEX-15 | [4,19] |
| 16 | AEX-23 | [4] |
| 17 | M4-G | [118] |
| 18 | SL-152 | [119,120] |
| 19 | Z1 | [63] |
| 20 | MD90-917 | [61] |
| 21 | IN68-9 | [44,57] |
| 22 | CM92-43 | [59,60] |
| 23 | GNS84-C106 | [54] |
| 24 | KET80-19 | [46] |
| 25 | BS79-22 | [52] |
| 26 | BS79-38 | [53] |
| 27 | ODP SITE-963 | [51] |
| 28 | REC13-53 | [46] |
| 29 | Hole 963 D | [51] |
| 30 | MD95-2043 | [48] |
| 31 | ODP977 | [48] |
| 32 | ST5 | [88] |
References
- Marino, G.; Rohling, E.J.; Sangiorgi, F.; Hayes, A.; Casford, J.L.; Lotter, A.F.; Kucera, M.; Brinkhuis, H. Early and middle Holocene in the Aegean Sea: Interplay between high and low latitude climate variability. Quat. Sci. Rev. 2009, 28, 3246–3262. [Google Scholar] [CrossRef]
- Rohling, E.J.; Marino, G.; Grant, K.M. Mediterranean climate and oceanography, and the periodic development of anoxic events (sapropels). Earth-Sci. Rev. 2015, 143, 62–97. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Antonarakou, A.; Kouli, K.; Dimiza, M.; Kontakiotis, G.; Papanikolaou, M.D.; Ziveri, P.; Mortyn, P.G.; Lianou, V.; Lykousis, V.; et al. Late Glacial–Holocene ecostratigraphy of the south-eastern Aegean Sea, based on plankton and pollen assemblages. Geo-Mar. Lett. 2009, 29, 249–267. [Google Scholar] [CrossRef]
- Giamali, C.; Koskeridou, E.; Antonarakou, A.; Ioakim, C.; Kontakiotis, G.; Karageorgis, A.P.; Roussakis, G.; Karakitsios, V. Multiproxy ecosystem response of abrupt Holocene climatic changes in the northeastern Mediterranean sedimentary archive and hydrologic regime. Quat. Res. 2019, 92, 665–685. [Google Scholar] [CrossRef]
- Giamali, C.; Kontakiotis, G.; Koskeridou, E.; Ioakim, C.; Antonarakou, A. Key Environmental Factors Controlling Planktonic Foraminiferal and Pteropod Community’s Response to Late Quaternary Hydroclimate Changes in the South Aegean Sea (Eastern Mediterranean). J. Mar. Sci. Eng. 2020, 8, 709. [Google Scholar] [CrossRef]
- Geraga, M.; Tsaila-Monopolis, S.; Ioakim, C.; Papatheodorou, G.; Ferentinos, G. Short-term climate changes in the southern Aegean Sea over the last 48,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 220, 311–332. [Google Scholar] [CrossRef]
- Geraga, M.; Tsaila-Monopolis, S.; Ioakim, C.; Papatheodorou, G.; Ferentinos, G. Evaluation of palaeoenvironmental changes during the last 18,000 years in the Myrtoon basin, SW Aegean Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 156, 1–17. [Google Scholar] [CrossRef]
- Geraga, M.; Ioakim, C.; Lykousis, V.; Tsaila-Monopolis, S.; Mylona, G. The high-resolution palaeoclimatic and palaeoceanographic history of the last 24,000 years in the central Aegean Sea, Greece. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 287, 101–115. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Mortyn, G.P.; Antonarakou, A.; Drinia, H. Assessing the reliability of foraminiferal Mg/Ca thermometry by comparing field-samples and culture experiments: A review. Geol. Q. 2016, 60, 547–560. [Google Scholar] [CrossRef]
- Dämmer, L.K.; de Nooijer, L.; van Sebille, E.; Haak, J.G.; Reichart, G.J. Evaluation of oxygen isotopes and trace elements in planktonic foraminifera from the Mediterranean Sea as recorders of seawater oxygen isotopes and salinity. Clim. Past 2020, 16, 2401–2414. [Google Scholar] [CrossRef]
- Wit, J.C.; Reichart, G.J.; Jung, S.J.A.; Kroon, D. Approaches to unravel seasonality in sea surface temperatures using paired single-specimen foraminiferal δ18O and Mg/Ca analyses. Paleoceanography 2010, 25, 58. [Google Scholar] [CrossRef]
- Dissard, D.; Reichart, G.J.; Menkes, C.; Mangeas, M.; Frickenhaus, S.; Bijma, J. Mg/Ca, Sr/Ca and stable isotopes from the planktonic foraminifera T. sacculifer: Testing a multi-proxy approach for inferring paleotemperature and paleosalinity. Biogeosciences 2021, 18, 423–439. [Google Scholar] [CrossRef]
- Le Houedec, S.; Erez, J.; Rosenthal, Y. Testing the Influence of Changing Seawater Ca Concentration on Elements/Ca Ratios in Planktic Foraminifera: A Culture Experiment. Geochem. Geophys. Geosystems 2021, 22, e2020GC009496. [Google Scholar] [CrossRef]
- Hoogakker, B.A.A.; Klinkhammer, G.P.; Elderfield, H.; Rohling, E.J.; Hayward, C. Mg/Ca paleothermometry in high salinity environments. Earth Planet. Sci. Lett. 2009, 284, 583–589. [Google Scholar] [CrossRef]
- Kısakürek, B.; Eisenhauer, A.; Böhm, F.; Garbe-Schönberg, D.; Erez, J. Controls on shell Mg/Ca and Sr/Ca in cultured planktonic foraminiferan, Globigerinoides ruber (white). Earth Planet. Sci. Lett. 2008, 273, 260–269. [Google Scholar] [CrossRef]
- Zarkogiannis, S.; Kontakiotis, G.; Antonarakou, A. Recent planktonic foraminifera population and size response to Eastern Mediterranean hydrography. Rev. De Micropaléontologie 2020, 69, 100450. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Antonarakou, A.; Mortyn, P.G.; Drinia, H.; Anastasakis, G.; Zarkogiannis, S.; Möbius, J. Morphological recognition of Globigerinoides ruber morphotypes and their susceptibility to diagenetic alteration in the eastern Mediterranean Sea. J. Mar. Syst. 2017, 174, 12–24. [Google Scholar] [CrossRef]
- Thunell, R.C. Distribution of recent planktonic foraminifera in surface sediments of the Mediterranean Sea. Mar. Micropaleontol. 1978, 3, 147–173. [Google Scholar] [CrossRef]
- Giamali, C.; Kontakiotis, G.; Antonarakou, A.; Koskeridou, E. Ecological Constraints of Plankton Bio-Indicators for Water Column Stratification and Productivity: A Case Study of the Holocene North Aegean Sedimentary Record. J. Mar. Sci. Eng. 2021, 9, 1249. [Google Scholar] [CrossRef]
- Rebotim, A.; Voelker, A.H.L.; Jonkers, L.; Waniek, J.J.; Meggers, H.; Schiebel, R.; Fraile, I.; Schulz, M.; Kucera, M. Factors controlling the depth habitat of planktonic foraminifera in the subtropical eastern North Atlantic. Biogeosciences 2017, 14, 827–859. [Google Scholar] [CrossRef]
- Wilke, I.; Meggers, H.; Bickert, T. Depth habitats and seasonal distributions of recent planktic foraminifers in the Canary Islands region (29°N) based on oxygen isotopes. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 89–106. [Google Scholar] [CrossRef]
- Takagi, H.; Kimoto, K.; Fujiki, T.; Saito, H.; Schmidt, C.; Kucera, M.; Moriya, K. Characterizing photosymbiosis in modern planktonic foraminifera. Biogeosciences 2019, 16, 3377–3396. [Google Scholar] [CrossRef]
- Zarkogiannis, S.D.; Antonarakou, A.; Tripati, A.; Kontakiotis, G.; Mortyn, P.G.; Drinia, H.; Greaves, M. Influence of surface ocean density on planktonic foraminifera calcification. Sci. Rep. 2019, 9, 533. [Google Scholar] [CrossRef] [PubMed]
- Weinkauf, M.F.G.; Kunze, J.G.; Waniek, J.J.; Kučera, M. Seasonal Variation in Shell Calcification of Planktonic Foraminifera in the NE Atlantic Reveals Species-Specific Response to Temperature, Productivity, and Optimum Growth Conditions. PLoS ONE 2016, 11, e0148363. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Li, T.; Liu, J.; Sun, H.; Chang, F. Spatial distribution and controlling factors of planktonic foraminifera in the modern western Pacific. Quat. Int. 2018, 468, 14–23. [Google Scholar] [CrossRef]
- Mallo, M.; Ziveri, P.; Mortyn, P.G.; Schiebel, R.; Grelaud, M. Low planktic foraminiferal diversity and abundance observed in a spring 2013 west–east Mediterranean Sea plankton tow transect. Biogeosciences 2017, 14, 2245–2266. [Google Scholar] [CrossRef]
- Pujol, C.; Grazzini, C.V. Distribution patterns of live planktic foraminifers as related to regional hydrography and productive systems of the Mediterranean Sea. Mar. Micropaleontol. 1995, 25, 187–217. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Efstathiou, E.; Zarkogiannis, S.D.; Besiou, E.; Antonarakou, A. Latitudinal Differentiation among Modern Planktonic Foraminiferal Populations of Central Mediterranean: Species–Specific Distribution Patterns and Size Variability. J. Mar. Sci. Eng. 2021, 9, 551. [Google Scholar] [CrossRef]
- Avnaim-Katav, S.; Herut, B.; Rahav, E.; Katz, T.; Weinstein, Y.; Alkalay, R.; Berman-Frank, I.; Zlatkin, O.; Almogi-Labin, A. Sediment trap and deep sea coretop sediments as tracers of recent changes in planktonic foraminifera assemblages in the southeastern ultra-oligotrophic Levantine Basin. Deep Sea Res. Part II Top. Stud. Oceanogr. 2020, 171, 104669. [Google Scholar] [CrossRef]
- Schmidt, D.N.; Renaud, S.; Bollmann, J.; Schiebel, R.; Thierstein, H.R. Size distribution of Holocene planktic foraminifer assemblages: Biogeography, ecology and adaptation. Mar. Micropaleontol. 2004, 50, 319–338. [Google Scholar] [CrossRef]
- Schmidt, D.N.; Thierstein, H.R.; Bollmann, J. The evolutionary history of size variation of planktic foraminiferal assemblages in the Cenozoic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 212, 159–180. [Google Scholar] [CrossRef]
- Schmidt, D.N.; Lazarus, D.; Young, J.R.; Kucera, M. Biogeography and evolution of body size in marine plankton. Earth-Sci. Rev. 2006, 78, 239–266. [Google Scholar] [CrossRef]
- Roussakis, G.; Karageorgis, A.P.; Conispoliatis, N.; Lykousis, V. Last glacial–Holocene sediment sequences in N. Aegean basins: Structure, accumulation rates and clay mineral distribution. Geo-Mar. Lett. 2004, 24, 97–111. [Google Scholar] [CrossRef]
- Giresse, P.; Buscail, R.; Charrière, B. Late Holocene multisource material input into the Aegean Sea: Depositional and post-depositional processes. Oceanol. Acta 2003, 26, 657–672. [Google Scholar] [CrossRef]
- Kontakiotis, G. Late Quaternary paleoenvironmental reconstruction and paleoclimatic implications of the Aegean Sea (eastern Mediterranean) based on paleoceanographic indexes and stable isotopes. Quat. Int. 2016, 401, 28–42. [Google Scholar] [CrossRef]
- Emeis, K.-C.; Struck, U.; Schulz, H.-M.; Rosenberg, R.; Bernasconi, S.; Erlenkeuser, H.; Sakamoto, T.; Martinez-Ruiz, F. Temperature and salinity variations of Mediterranean Sea surface waters over the last 16,000 years from records of planktonic stable oxygen isotopes and alkenone unsaturation ratios. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 158, 259–280. [Google Scholar] [CrossRef]
- Budillon, F.; Lirer, F.; Iorio, M.; Macrì, P.; Sagnotti, L.; Vallefuoco, M.; Ferraro, L.; Garziglia, S.; Innangi, S.; Sahabi, M.; et al. Integrated stratigraphic reconstruction for the last 80kyr in a deep sector of the Sardinia Channel (Western Mediterranean). Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 725–737. [Google Scholar] [CrossRef]
- Drinia, H.; Antonarakou, A.; Tsourou, T.; Kontakiotis, G.; Psychogiou, M.; Anastasakis, G. Foraminifera eco-biostratigraphy of the southern Evoikos outer shelf, central Aegean Sea, during MIS 5 to present. Cont. Shelf Res. 2016, 126, 36–49. [Google Scholar] [CrossRef]
- Aksu, A.E.; Jenner, G.; Hiscott, R.N.; İşler, E.B. Occurrence, stratigraphy and geochemistry of Late Quaternary tephra layers in the Aegean Sea and the Marmara Sea. Mar. Geol. 2008, 252, 174–192. [Google Scholar] [CrossRef]
- Satow, C.; Tomlinson, E.L.; Grant, K.M.; Albert, P.G.; Smith, V.C.; Manning, C.J.; Ottolini, L.; Wulf, S.; Rohling, E.J.; Lowe, J.J.; et al. A new contribution to the Late Quaternary tephrostratigraphy of the Mediterranean: Aegean Sea core LC21. Quat. Sci. Rev. 2015, 117, 96–112. [Google Scholar] [CrossRef]
- Petrosino, P.; Morabito, S.; Jicha, B.R.; Milia, A.; Sprovieri, M.; Tamburrino, S. Multidisciplinary tephrochronological correlation of marker events in the eastern Tyrrhenian Sea between 48 and 105 ka. J. Volcanol. Geotherm. Res. 2016, 315, 79–99. [Google Scholar] [CrossRef]
- Wulf, S.; Keller, J.; Satow, C.; Gertisser, R.; Kraml, M.; Grant, K.M.; Appelt, O.; Vakhrameeva, P.; Koutsodendris, A.; Hardiman, M.; et al. Advancing Santorini’s tephrostratigraphy: New glass geochemical data and improved marine-terrestrial tephra correlations for the past ~360 kyrs. Earth-Sci. Rev. 2020, 200, 102964. [Google Scholar] [CrossRef]
- Morabito, S.; Petrosino, P.; Milia, A.; Sprovieri, M.; Tamburrino, S. A multidisciplinary approach for reconstructing the stratigraphic framework of the last 40ka in a bathyal area of the eastern Tyrrhenian Sea. Glob. Planet. Chang. 2014, 123, 121–138. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Abu-Zied, R.; Rohling, E.J.; Cooke, S.; Fontanier, C.; Leng, M.; Millard, A.; Thomson, J. A stratigraphically controlled multiproxy chronostratigraphy for the eastern Mediterranean. Paleoceanography 2007, 22, 102964. [Google Scholar] [CrossRef]
- Antonarakou, A.; Kontakiotis, G.; Karageorgis, A.P.; Besiou, E.; Zarkogiannis, S.; Drinia, H.; Mortyn, G.P.; Tripsanas, E. Eco-biostratigraphic advances on late Quaternary geochronology and palaeoclimate: The marginal Gulf of Mexico analogue. Geol. Q. 2019, 63, 178–191. [Google Scholar] [CrossRef]
- Zouari, S.; Boussetta, S.; Siani, G.; Tisnerat-Laborde, N.; Thil, F.; Kallel, A.; Michel, E.; Kallel, N. Mediterranean faunal evolution and biochronological events during the last 24 kyr. Mar. Micropaleontol. 2021, 165, 101997. [Google Scholar] [CrossRef]
- Lirer, F.; Foresi, L.M.; Iaccarino, S.M.; Salvatorini, G.; Turco, E.; Cosentino, C.; Sierro, F.J.; Caruso, A. Mediterranean Neogene planktonic foraminifer biozonation and biochronology. Earth-Sci. Rev. 2019, 196, 102869. [Google Scholar] [CrossRef]
- Pérez-Folgado, M.; Sierro, F.J.; Flores, J.-A.; Cacho, I.; Grimalt, J.O.; Zahn, R.; Shackleton, N.J. Planktonic foraminiferal distribution during the last 70 kyr in the western Mediterranean Sea. Mar. Micropaleontol. 2003, 48, 49–70. [Google Scholar] [CrossRef]
- Pérez-Folgado, M.; Sierro, F.J.; Flores, J.A.; Grimalt, J.O.; Zahn, R. Paleoclimatic variations in foraminifer assemblages from the Alboran Sea (Western Mediterranean) during the last 150 ka in ODP Site 977. Mar. Geol. 2004, 212, 113–131. [Google Scholar] [CrossRef]
- Suárez-Ibarra, J.Y.; Petró, S.M.; Frozza, C.F.; Freire, T.M.; Portilho-Ramos, R.D.C.; Pivel, M.A.G. Time-spatial boundaries of bioecozonations (planktonic foraminifera) in the latest Quaternary: A case study from the western South Atlantic. Rev. Micropaléontol. 2021, 73, 100554. [Google Scholar] [CrossRef]
- Sprovieri, R.; Stefano, E.; Incarbona, A.; Gargano, M. A high-resolution record of the last deglaciation in the Sicily Channel based on foraminifera and calcareous nannofossil quantitative distribution. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 202, 119–142. [Google Scholar] [CrossRef]
- Sbaffi, L.; Wezel, F.; Kallel, N.; Paterne, M.; Cacho, I.; Ziveri, P.; Shackleton, N. Response of the pelagic environment to palaeoclimatic changes in the central Mediterranean Sea during the Late Quaternary. Mar. Geol. 2001, 178, 39–62. [Google Scholar] [CrossRef]
- Sbaffi, L.; Wezel, F.C.; Curzi, G.; Zoppi, U. Millennial- to centennial-scale palaeoclimatic variations during Termination I and the Holocene in the central Mediterranean Sea. Glob. Planet. Chang. 2004, 40, 201–217. [Google Scholar] [CrossRef]
- Di Donato, V.; Esposito, P.; Russo-Ermolli, E.; Scarano, A.; Cheddadi, R. Coupled atmospheric and marine palaeoclimatic reconstruction for the last 35 ka in the Sele Plain–Gulf of Salerno area (southern Italy). Quat. Int. 2008, 190, 146–157. [Google Scholar] [CrossRef]
- Lirer, F.; Sprovieri, M.; Ferraro, L.; Vallefuoco, M.; Capotondi, L.; Cascella, A.; Petrosino, P.; Insinga, D.D.; Pelosi, N.; Tamburrino, S.; et al. Integrated stratigraphy for the Late Quaternary in the eastern Tyrrhenian Sea. Quat. Int. 2013, 292, 71–85. [Google Scholar] [CrossRef]
- Amore, F.O.; Caffau, M.; Massa, B.; Morabito, S. Late Pleistocene–Holocene paleoclimate and related paleoenvironmental changes as recorded by calcareous nannofossils and planktonic foraminifera assemblages in the southern Tyrrhenian Sea (Cape Palinuro, Italy). Mar. Micropaleontol. 2004, 52, 255–276. [Google Scholar] [CrossRef]
- Jorissen, F.J.; Asioli, A.; Borsetti, A.M.; Capotondi, L.; de Visser, J.P.; Hilgen, F.J.; Rohling, E.J.; van der Borg, K.; Vergnaud Grazzini, C.; Zachariasse, W.J. Late Quaternary central Mediterranean biochronology. Mar. Micropaleontol. 1993, 21, 169–189. [Google Scholar] [CrossRef]
- Capotondi, L.; Maria Borsetti, A.; Morigi, C. Foraminiferal ecozones, a high resolution proxy for the late Quaternary biochronology in the central Mediterranean Sea. Mar. Geol. 1999, 153, 253–274. [Google Scholar] [CrossRef]
- Asioli, A.; Trincardi, F.; Lowe, J.J.; Ariztegui, D.; Langone, L.; Oldfield, F. Sub-millennial scale climatic oscillations in the central Adriatic during the Lateglacial: Palaeoceanographic implications. Quat. Sci. Rev. 2001, 20, 1201–1221. [Google Scholar] [CrossRef]
- Asioli, A.; Trincardi, F.; Lowe, J.J.; Oldfield, F. Short-term climate changes during the Last Glacial–Holocene transition: Comparison between Mediterranean records and the GRIP event stratigraphy. J. Quat. Sci. 1999, 14, 373–381. [Google Scholar] [CrossRef]
- Siani, G.; Paterne, M.; Colin, C. Late glacial to Holocene planktic foraminifera bioevents and climatic record in the South Adriatic Sea. J. Quat. Sci. 2010, 25, 808–821. [Google Scholar] [CrossRef]
- Principato, M.S.; Giunta, S.; Corselli, C.; Negri, A. Late Pleistocene–Holocene planktonic assemblages in three box-cores from the Mediterranean Ridge area (west–southwest of Crete): Palaeoecological and palaeoceanographic reconstruction of sapropel S1 interval. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 190, 61–77. [Google Scholar] [CrossRef]
- Geraga, M.; Mylona, G.; Tsaila-Monopoli, S.; Papatheodorou, G.; Ferentinos, G. Northeastern Ionian Sea: Palaeoceanographic variability over the last 22 ka. J. Mar. Syst. 2008, 74, 623–638. [Google Scholar] [CrossRef]
- Louvari, M.A.; Drinia, H.; Kontakiotis, G.; Di Bella, L.; Antonarakou, A.; Anastasakis, G. Impact of latest-glacial to Holocene sea-level oscillations on central Aegean shelf ecosystems: A benthic foraminiferal palaeoenvironmental assessment of South Evoikos Gulf, Greece. J. Mar. Syst. 2019, 199, 103181. [Google Scholar] [CrossRef]
- Lykousis, V.; Chronis, G.; Tselepides, A.; Price, N.B.; Theocharis, A.; Siokou-Frangou, I.; Van Wambeke, F.; Danovaro, R.; Stavrakakis, S.; Duineveld, G.; et al. Major outputs of the recent multidisciplinary biogeochemical researches undertaken in the Aegean Sea. J. Mar. Syst. 2002, 33–34, 313–334. [Google Scholar] [CrossRef]
- Ignatiades, L.; Psarra, S.; Zervakis, V.; Pagou, K.; Souvermezoglou, E.; Assimakopoulou, G.; Gotsis-Skretas, O. Phytoplankton size-based dynamics in the Aegean Sea (Eastern Mediterranean). J. Mar. Syst. 2002, 36, 11–28. [Google Scholar] [CrossRef]
- Price, N.B.; Lindsay, F.S.; Pates, J.M. The biogeochemistry of major elements of the suspended particulate matter of the Cretan Sea. Prog. Oceanogr. 1999, 44, 677–699. [Google Scholar] [CrossRef]
- Psarra, S.; Tselepides, A.; Ignatiades, L. Primary productivity in the oligotrophic Cretan Sea (NE Mediterranean): Seasonal and interannual variability. Prog. Oceanogr. 2000, 46, 187–204. [Google Scholar] [CrossRef]
- Tselepides, A.; Zervakis, V.; Polychronaki, T.; Danovaro, R.; Chronis, G. Distribution of nutrients and particulate organic matter in relation to the prevailing hydrographic features of the Cretan Sea (NE Mediterranean). Prog. Oceanogr. 2000, 46, 113–142. [Google Scholar] [CrossRef]
- Karageorgis, A.P.; Gardner, W.D.; Georgopoulos, D.; Mishonov, A.V.; Krasakopoulou, E.; Anagnostou, C. Particle dynamics in the Eastern Mediterranean Sea: A synthesis based on light transmission, PMC, and POC archives (1991–2001). Deep Sea Res. Part I Oceanogr. Res. Pap. 2008, 55, 177–202. [Google Scholar] [CrossRef]
- Theocharis, A.; Georgopoulos, D.; Lascaratos, A.; Nittis, K. Water masses and circulation in the central region of the Eastern Mediterranean: Eastern Ionian, South Aegean and Northwest Levantine, 1986–1987. Deep Sea Res. Part II Top. Stud. Oceanogr. 1993, 40, 1121–1142. [Google Scholar] [CrossRef]
- Papanikolaou, D.; Alexandri, M.; Nomikou, P.; Ballas, D. Morphotectonic structure of the western part of the North Aegean Basin based on swath bathymetry. Mar. Geol. 2002, 190, 465–492. [Google Scholar] [CrossRef]
- Poulos, S.E. Origin and distribution of the terrigenous component of the unconsolidated surface sediment of the Aegean floor: A synthesis. Cont. Shelf Res. 2009, 29, 2045–2060. [Google Scholar] [CrossRef]
- Roether, W.; Klein, B.; Manca, B.B.; Theocharis, A.; Kioroglou, S. Transient Eastern Mediterranean deep waters in response to the massive dense-water output of the Aegean Sea in the 1990s. Prog. Oceanogr. 2007, 74, 540–571. [Google Scholar] [CrossRef]
- Theocharis, A.; Balopoulos, E.; Kioroglou, S.; Kontoyiannis, H.; Iona, A. A synthesis of the circulation and hydrography of the South Aegean Sea and the Straits of the Cretan Arc (March 1994–January 1995). Prog. Oceanogr. 1999, 44, 469–509. [Google Scholar] [CrossRef]
- Pinardi, N.; Zavatarelli, M.; Adani, M.; Coppini, G.; Fratianni, C.; Oddo, P.; Simoncelli, S.; Tonani, M.; Lyubartsev, V.; Dobricic, S.; et al. Mediterranean Sea large-scale low-frequency ocean variability and water mass formation rates from 1987 to 2007: A retrospective analysis. Prog. Oceanogr. 2015, 132, 318–332. [Google Scholar] [CrossRef]
- Zervakis, V.; Georgopoulos, D.; Drakopoulos, P.G. The role of the North Aegean in triggering the recent Eastern Mediterranean climatic changes. J. Geophys. Res. Ocean. 2000, 105, 26103–26116. [Google Scholar] [CrossRef]
- Lykousis, V. Subaqueous bedforms on the Cyclades Plateau (NE Mediterranean)—Evidence of Cretan Deep Water Formation? Cont. Shelf Res. 2001, 21, 495–507. [Google Scholar] [CrossRef]
- Mercone, D.; Thomson, J.; Croudace, I.W.; Siani, G.; Paterne, M.; Troelstra, S. Duration of S1, the most recent sapropel in the eastern Mediterranean Sea, as indicated by accelerator mass spectrometry radiocarbon and geochemical evidence. Paleoceanography 2000, 15, 336–347. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V. Coccolithophore assemblages during the Holocene Climatic Optimum in the NE Mediterranean (Aegean and northern Levantine Seas, Greece): Paleoceanographic and paleoclimatic implications. Quat. Int. 2014, 345, 56–67. [Google Scholar] [CrossRef]
- Reimer, P.J.; McCormac, F.G. Marine Radiocarbon Reservoir Corrections for the Mediterranean and Aegean Seas. Radiocarbon 2016, 44, 159–166. [Google Scholar] [CrossRef]
- Stuiver, M.; Reimer, P.J.; Reimer, R.W. CALIB 8.2 [WWW Program]. 2021, Volume 13. Available online: http://calib.org (accessed on 1 January 2023).
- Zarkogiannis, S.D.; Kontakiotis, G.; Gkaniatsa, G.; Kuppili, V.S.C.; Marathe, S.; Wanelik, K.; Lianou, V.; Besiou, E.; Makri, P.; Antonarakou, A. An Improved Cleaning Protocol for Foraminiferal Calcite from Unconsolidated Core Sediments: HyPerCal—A New Practice for Micropaleontological and Paleoclimatic Proxies. J. Mar. Sci. Eng. 2020, 8, 998. [Google Scholar] [CrossRef]
- Capotondi, L.; Soroldoni, E.; Principato, M.S.; Corselli, C. Late Quaternary planktonic foraminiferal distributions: Problems related to size fraction. In Proceedings of the First Italian Meeting on EnvironmentalMicropaleontology, Urbino, Italy, 4–6 June 2002; Coccioni, R.G.S., Lirer, F., Eds.; Grzybowski Foundation, Special Publication: Krakow, Poland, 2004; Volume 9, pp. 1–6. [Google Scholar]
- Fatela, F.; Taborda, R. Confidence limits of species proportions in microfossil assemblages. Mar. Micropaleontol. 2002, 45, 169–174. [Google Scholar] [CrossRef]
- Hemleben, C.; Anderson, O.R.; Spindler, M. Modern Planktonic Foraminifera; Springer-Verlag: New York, NY, USA, 1989. [Google Scholar]
- Brummer, G.J.A.; Kučera, M. Taxonomic review of living planktonic foraminifera. J. Micropalaeontol. 2022, 41, 29–74. [Google Scholar] [CrossRef]
- Kozanoglou, C.; Triantaphyllou, M.V.; Geraga, M.; Rousakis, G.; Papatheodorou, G.; Arabas, A.; Dimiza, M.D.; Gogou, A. A high-resolution study of planktonic foraminifera during the Holocene at the Tilos-Symi sea basin in the SE Aegean Sea. Holocene 2023, 33, 1317–1332. [Google Scholar] [CrossRef]
- Rohling, E.J.; Jorissen, F.J.; Grazzini, C.V.; Zachariasse, W.J. Northern Levantine and Adriatic Quaternary planktic foraminifera; Reconstruction of paleoenvironmental gradients. Mar. Micropaleontol. 1993, 21, 191–218. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Ziveri, P.; Gogou, A.; Marino, G.; Lykousis, V.; Bouloubassi, I.; Emeis, K.C.; Kouli, K.; Dimiza, M.; Rosell-Melé, A.; et al. Late Glacial–Holocene climate variability at the south-eastern margin of the Aegean Sea. Mar. Geol. 2009, 266, 182–197. [Google Scholar] [CrossRef]
- Essallami, L.; Sicre, M.A.; Kallel, N.; Labeyrie, L.; Siani, G. Hydrological changes in the Mediterranean Sea over the last 30,000 years. Geochem. Geophys. Geosystems 2007, 8, 59. [Google Scholar] [CrossRef]
- Rouis-Zargouni, I.; Turon, J.-L.; Londeix, L.; Essallami, L.; Kallel, N.; Sicre, M.-A. Environmental and climatic changes in the central Mediterranean Sea (Siculo–Tunisian Strait) during the last 30ka based on dinoflagellate cyst and planktonic foraminifera assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 285, 17–29. [Google Scholar] [CrossRef]
- Zachariasse, W.; Jorissen, F.; Perissoratis, C.; Rohling, E.; Tsapralis, V. Late Quaternary foraminiferal changes and the nature of sapropel S1 in Skopelos Basin. In Proceedings of the 5th Hellenic Symposium of Oceanography and Fisheries, Kavala, Greece, 27 July 1997; pp. 391–394. [Google Scholar]
- Hayes, A.; Rohling, E.J.; De Rijk, S.; Kroon, D.; Zachariasse, W.J. Mediterranean planktonic foraminiferal faunas during the last glacial cycle. Mar. Geol. 1999, 153, 239–252. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Rohling, E.J.; Abu-Zied, R.; Cooke, S.; Fontanier, C.; Leng, M.; Lykousis, V. Circulation changes and nutrient concentrations in the late Quaternary Aegean Sea: A nonsteady state concept for sapropel formation. Paleoceanography 2002, 17, 14-1–14-11. [Google Scholar] [CrossRef]
- Rohling, E.J.; Jorissen, F.J.; De Stigter, H.C. 200 Year interruption of Holocene sapropel formation in the Adriatic Sea. J. Micropalaeontol. 1997, 16, 97–108. [Google Scholar] [CrossRef]
- De Rijk, S.; Hayes, A.; Rohling, E.J. Eastern Mediterranean sapropel S1 interruption: An expression of the onset of climatic deterioration around 7 ka BP. Mar. Geol. 1999, 153, 337–343. [Google Scholar] [CrossRef]
- Papanikolaou, D.; Nomikou, P. Tectonic structure and volcanic centers at the eastern edge of the aegean volcanic arc around Nisyros island. Bull. Geol. Soc. Greece 2001, 34, 289–296. [Google Scholar] [CrossRef]
- Pujol, C.; Vergnaud Grazzini, C. Palaeoceanography of the last deglaciation in the Alboran Sea (western Mediterranean). Stable isotopes and planktonic foraminiferal records. Mar. Micropaleontol. 1989, 15, 153–179. [Google Scholar] [CrossRef]
- Rohling, E.J.; Gieskes, W.W.C. Late Quaternary changes in Mediterranean intermediate water density and formation rate. Paleoceanography 1989, 4, 531–545. [Google Scholar] [CrossRef]
- Cacho, I.; Grimalt, J.O.; Canals, M.; Sbaffi, L.; Shackleton, N.J.; Schönfeld, J.; Zahn, R. Variability of the western Mediterranean Sea surface temperature during the last 25,000 years and its connection with the Northern Hemisphere climatic changes. Paleoceanography 2001, 16, 40–52. [Google Scholar] [CrossRef]
- Buccheri, G.; Capretto, G.; Di Donato, V.; Esposito, P.; Ferruzza, G.; Pescatore, T.; Russo Ermolli, E.; Senatore, M.R.; Sprovieri, M.; Bertoldo, M.; et al. A high resolution record of the last deglaciation in the southern Tyrrhenian Sea: Environmental and climatic evolution. Mar. Geol. 2002, 186, 447–470. [Google Scholar] [CrossRef]
- Carboni, M.G.; Bergamin, L.; Di Bella, L.; Landini, B.; Manfra, L.; Vesica, P. Late Quaternary paleoclimatic and paleoenviromental changes in the Tyrrhenian Sea. Quat. Sci. Rev. 2005, 24, 2069–2082. [Google Scholar] [CrossRef]
- Paterne, M.; Kallel, N.; Labeyrie, L.; Vautravers, M.; Duplessy, J.-C.; Rossignol-Strick, M.; Cortijo, E.; Arnold, M.; Fontugne, M. Hydrological relationship between the North Atlantic Ocean and the Mediterranean Sea during the past 15–75 kyr. Paleoceanography 1999, 14, 626–638. [Google Scholar] [CrossRef]
- Bordon, A.; Peyron, O.; Lézine, A.-M.; Brewer, S.; Fouache, E. Pollen-inferred Late-Glacial and Holocene climate in southern Balkans (Lake Maliq). Quat. Int. 2009, 200, 19–30. [Google Scholar] [CrossRef]
- Larocque, I.; Finsinger, W. Late-glacial chironomid-based temperature reconstructions for Lago Piccolo di Avigliana in the southwestern Alps (Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 257, 207–223. [Google Scholar] [CrossRef]
- Dormoy, I.; Peyron, O.; Combourieu Nebout, N.; Goring, S.; Kotthoff, U.; Magny, M.; Pross, J. Terrestrial climate variability and seasonality changes in the Mediterranean region between 15 000 and 4000 years BP deduced from marine pollen records. Clim. Past 2009, 5, 615–632. [Google Scholar] [CrossRef]
- Björck, S.; Walker, M.J.C.; Cwynar, L.C.; Johnsen, S.; Knudsen, K.-L.; Lowe, J.J.; Wohlfarth, B. An event stratigraphy for the Last Termination in the North Atlantic region based on the Greenland ice-core record: A proposal by the INTIMATE group. J. Quat. Sci. 1998, 13, 283–292. [Google Scholar] [CrossRef]
- Peyron, O.; Bégeot, C.; Brewer, S.; Heiri, O.; Magny, M.; Millet, L.; Ruffaldi, P.; Van Campo, E.; Yu, G. Lateglacial climate in the Jura mountains (France) based on different quantitative reconstruction approaches from pollen, lake-levels, and chironomids. Quat. Res. 2005, 64, 197–211. [Google Scholar] [CrossRef]
- Rasmussen, S.O.; Andersen, K.K.; Svensson, A.M.; Steffensen, J.P.; Vinther, B.M.; Clausen, H.B.; Siggaard-Andersen, M.L.; Johnsen, S.J.; Larsen, L.B.; Dahl-Jensen, D.; et al. A new Greenland ice core chronology for the last glacial termination. J. Geophys. Res. Atmos. 2006, 111, 1348. [Google Scholar] [CrossRef]
- Gogou, A.; Bouloubassi, I.; Lykousis, V.; Arnaboldi, M.; Gaitani, P.; Meyers, P.A. Organic geochemical evidence of Late Glacial–Holocene climate instability in the North Aegean Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 256, 1–20. [Google Scholar] [CrossRef]
- De Lange, G.J.; Thomson, J.; Reitz, A.; Slomp, C.P.; Speranza Principato, M.; Erba, E.; Corselli, C. Synchronous basin-wide formation and redox-controlled preservation of a Mediterranean sapropel. Nat. Geosci. 2008, 1, 606–610. [Google Scholar] [CrossRef]
- Alley, R.B.; Mayewski, P.A.; Sowers, T.; Stuiver, M.; Taylor, K.C.; Clark, P.U. Holocene climatic instability: A prominent, widespread event 8200 yr ago. Geology 1997, 25, 483–486. [Google Scholar] [CrossRef]
- Ariztegui, D.; Asioli, A.; Lowe, J.J.; Trincardi, F.; Vigliotti, L.; Tamburini, F.; Chondrogianni, C.; Accorsi, C.A.; Bandini Mazzanti, M.; Mercuri, A.M.; et al. Palaeoclimate and the formation of sapropel S1: Inferences from Late Quaternary lacustrine and marine sequences in the central Mediterranean region. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 158, 215–240. [Google Scholar] [CrossRef]
- Rohling, E.; Mayewski, P.; Abu-Zied, R.; Casford, J.; Hayes, A. Holocene atmosphere-ocean interactions: Records from Greenland and the Aegean Sea. Clim. Dyn. 2002, 18, 587–593. [Google Scholar] [CrossRef]
- Bazzicalupo, P.; Maiorano, P.; Girone, A.; Marino, M.; Combourieu-Nebout, N.; Pelosi, N.; Salgueiro, E.; Incarbona, A. Holocene climate variability of the Western Mediterranean: Surface water dynamics inferred from calcareous plankton assemblages. Holocene 2020, 30, 691–708. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Rohling, E.J.; Abu-Zied, R.H.; Fontanier, C.; Jorissen, F.J.; Leng, M.J.; Schmiedl, G.; Thomson, J. A dynamic concept for eastern Mediterranean circulation and oxygenation during sapropel formation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 190, 103–119. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Gogou, A.; Dimiza, M.D.; Kostopoulou, S.; Parinos, C.; Roussakis, G.; Geraga, M.; Bouloubassi, I.; Fleitmann, D.; Zervakis, V.; et al. Holocene Climatic Optimum centennial-scale paleoceanography in the NE Aegean (Mediterranean Sea). Geo-Mar. Lett. 2016, 36, 51–66. [Google Scholar] [CrossRef]
- Kotthoff, U.; Müller, U.C.; Pross, J.; Schmiedl, G.; Lawson, I.T.; van de Schootbrugge, B.; Schulz, H. Lateglacial and Holocene vegetation dynamics in the Aegean region: An integrated view based on pollen data from marine and terrestrial archives. Holocene 2008, 18, 1019–1032. [Google Scholar] [CrossRef]
- Kotthoff, U.; Pross, J.; Müller, U.C.; Peyron, O.; Schmiedl, G.; Schulz, H.; Bordon, A. Climate dynamics in the borderlands of the Aegean Sea during formation of sapropel S1 deduced from a marine pollen record. Quat. Sci. Rev. 2008, 27, 832–845. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Core Depth (cm) | AMS Lab Code | Conventional 14C Age (yr) | Calibrated Age (yr cal BP) | 2σ Age Range |
|---|---|---|---|---|
| 56–58 | Beta-253709 | 4690 ± 40 | 4710 | 4570–4850 |
| 158–160 | UCIAMS-256864 | 11,020 ± 25 | 11,020 | 11,956–12,572 |
| 200–202 | UCIAMS-256865 | 14,350 ± 35 | 16,472 | 16,144–16,800 |
| 254–256 | Beta-303651 | 18,300 ± 80 | 21,260 | 21,110–21,410 |
| Synchronous Planktonic Foraminiferal Bioevents within the Mediterranean Sea | Control Points | AMS 14C | Depth (cm) | Age (ka cal BP) |
|---|---|---|---|---|
| (s1) Last glacial peak of T. quinqueloba | 215 | 18.3 | ||
| AMS | 201 | 16.47 | ||
| (s2) First post-glacial occurrence of G. inflata | 195 | 15.3 | ||
| (s3-s4) Last post-glacial occurrence of G. truncatulinoides and disappearance of G. scitula | 185 | 14.5 | ||
| (s5) G. inflata maximum abundance during B/A | 175 | 13.5 | ||
| (s6-s7) T. quinqueloba and G. bulloides peaks during YD | 170 | 12.8 | ||
| (s8) G. glutinata peak at the B/A-YD boundary | 160 | 12.3 | ||
| AMS | 159 | 12.26 | ||
| Base S1a | 128 | 10 | ||
| Top S1a | 102 | 7.9 | ||
| (s9) Frequency increase of G. inflata during the sapropel interruption S1i | 98 | 7.7 | ||
| Base S1b | 94 | 7.3 | ||
| Top S1b | 82 | 6.4 | ||
| (s10) G. inflata peak after the end of sapropel S1 | 74 | 5.8 | ||
| Base SMH | 64 | 5.4 | ||
| AMS | 57 | 4.71 | ||
| Top SMH | 49 | 4.3 |
| Newly Defined Diachronous Planktonic Bioevents in Aegean Sea | Depth (cm) | Age (ka cal BP) |
|---|---|---|
| (d1) Decrease of G. ruber (w) at the end of the Latest Glacial | 200 | 16.2 |
| (d2) Increase of G. scitula during the Oldest Dryas | 200 | 16.2 |
| (d3) Increase of G. ruber (w) at the start of Bølling–Allerød (base of bioevent Ra2) | 190 | 14.9 |
| (d4) First occurrence of G. ruber rosea during the deglaciation | 180 | 14.0 |
| (d5) G. truncatulinoides maximum during the Bølling–Allerød | 170 | 12.8 |
| (d6) Minimum abundance of G. bulloides during the Younger Dryas | 155 | 11.97 |
| (d7) Minimum abundance of G. glutinata during the Younger Dryas | 155 | 11.97 |
| (d8) Re-occurrence of O. universa with the start of Holocene | 138 | 10.73 |
| (d9) Maximum abundance of O. universa below the sapropel S1 | 133 | 10.36 |
| (d10) Peak in abundance of G. glutinata within S1a | 118 | 9.2 |
| (d11) Peak in abundance of G. ruber rosea within S1a | 114 | 8.87 |
| (d12) Peak in abundance of O. universa within S1a | 114 | 8.87 |
| (d13) Maximum abundance of G. siphonifera within S1a | 110 | 8.55 |
| (d14) G. rubescens peak within S1a | 106 | 8.23 |
| (d15) G. trilobus peak within S1b | 86 | 6.7 |
| (d16) G. siphonifera peak within s1b | 86 | 6.7 |
| (d17) O. universa peak within s1b | 90 | 7.0 |
| (d18) Near absence of G. inflata at the top of S1b | 82 | 6.40 |
| (d19) G. rubescens peak within SMH | 59 | 4.91 |
| (d20) G. bulloides decline during SMH (top of bioevent B0) | 59 | 4.91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kontakiotis, G.; Antonarakou, A.; Besiou, E.; Skampa, E.; Triantaphyllou, M.V. Towards an Integrated and Accurate Planktonic-Foraminiferal-Deduced Bio-Chrono-Stratigraphic Framework of Late Quaternary Mediterranean Marine Cores. J. Mar. Sci. Eng. 2023, 11, 2345. https://doi.org/10.3390/jmse11122345
Kontakiotis G, Antonarakou A, Besiou E, Skampa E, Triantaphyllou MV. Towards an Integrated and Accurate Planktonic-Foraminiferal-Deduced Bio-Chrono-Stratigraphic Framework of Late Quaternary Mediterranean Marine Cores. Journal of Marine Science and Engineering. 2023; 11(12):2345. https://doi.org/10.3390/jmse11122345
Chicago/Turabian StyleKontakiotis, George, Assimina Antonarakou, Evangelia Besiou, Elisavet Skampa, and Maria V. Triantaphyllou. 2023. "Towards an Integrated and Accurate Planktonic-Foraminiferal-Deduced Bio-Chrono-Stratigraphic Framework of Late Quaternary Mediterranean Marine Cores" Journal of Marine Science and Engineering 11, no. 12: 2345. https://doi.org/10.3390/jmse11122345