

Environmental Assessment with Cage Exposure in the Neva Estuary, Baltic Sea: Metal Bioaccumulation and Physiologic Activity of Bivalve Molluscs

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
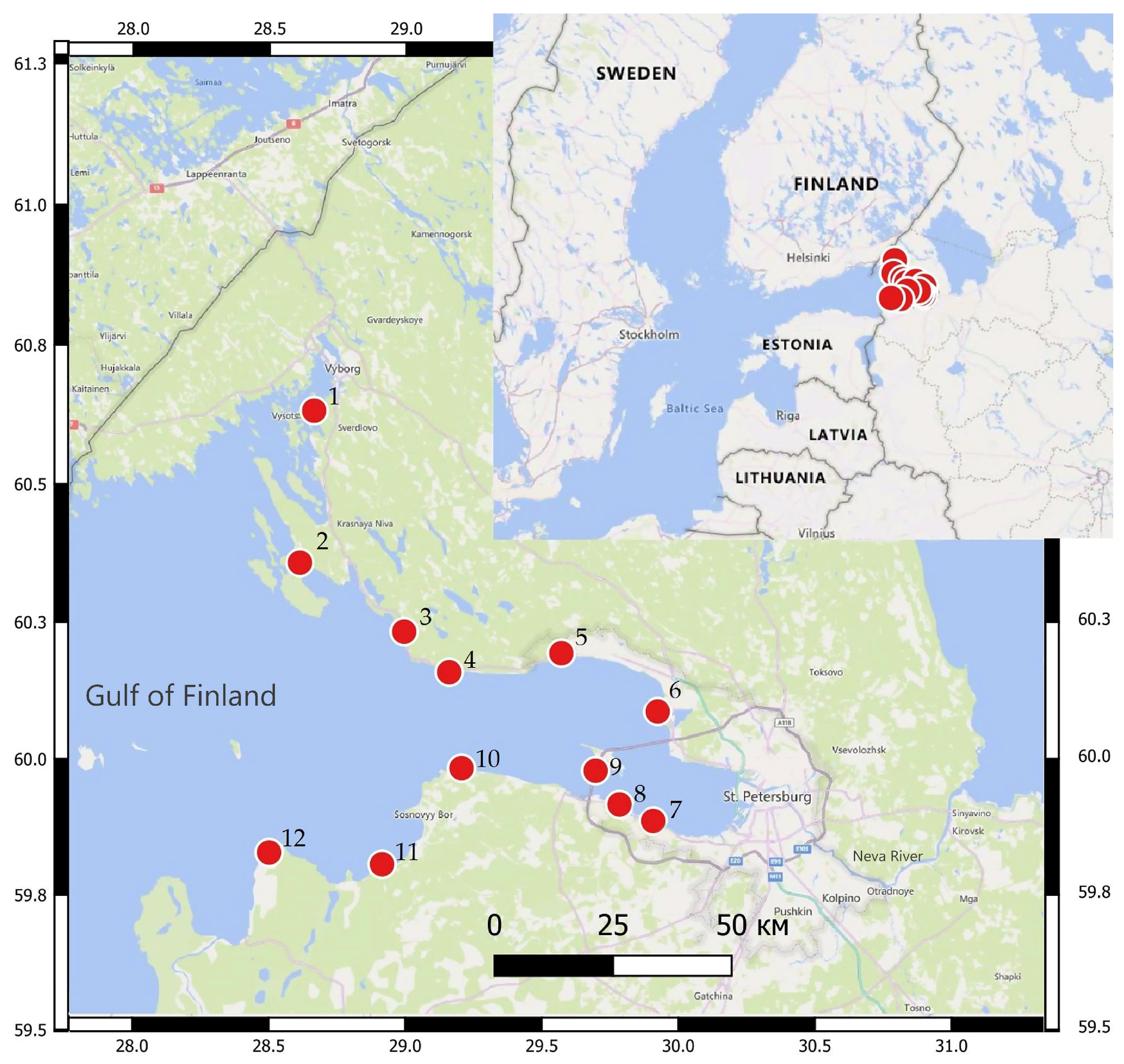
2.1. Study Area
2.2. Sampling Design
2.3. Biotesting
2.4. Chemical Analyses
2.5. Bioaccumulation Factor
2.6. Mollusc Physiological State
2.7. Statistics
3. Results
3.1. Physical and Chemical Parameters of Sediment Statistics
3.2. Metal and PAHs Concentrations in Biota and Sediments
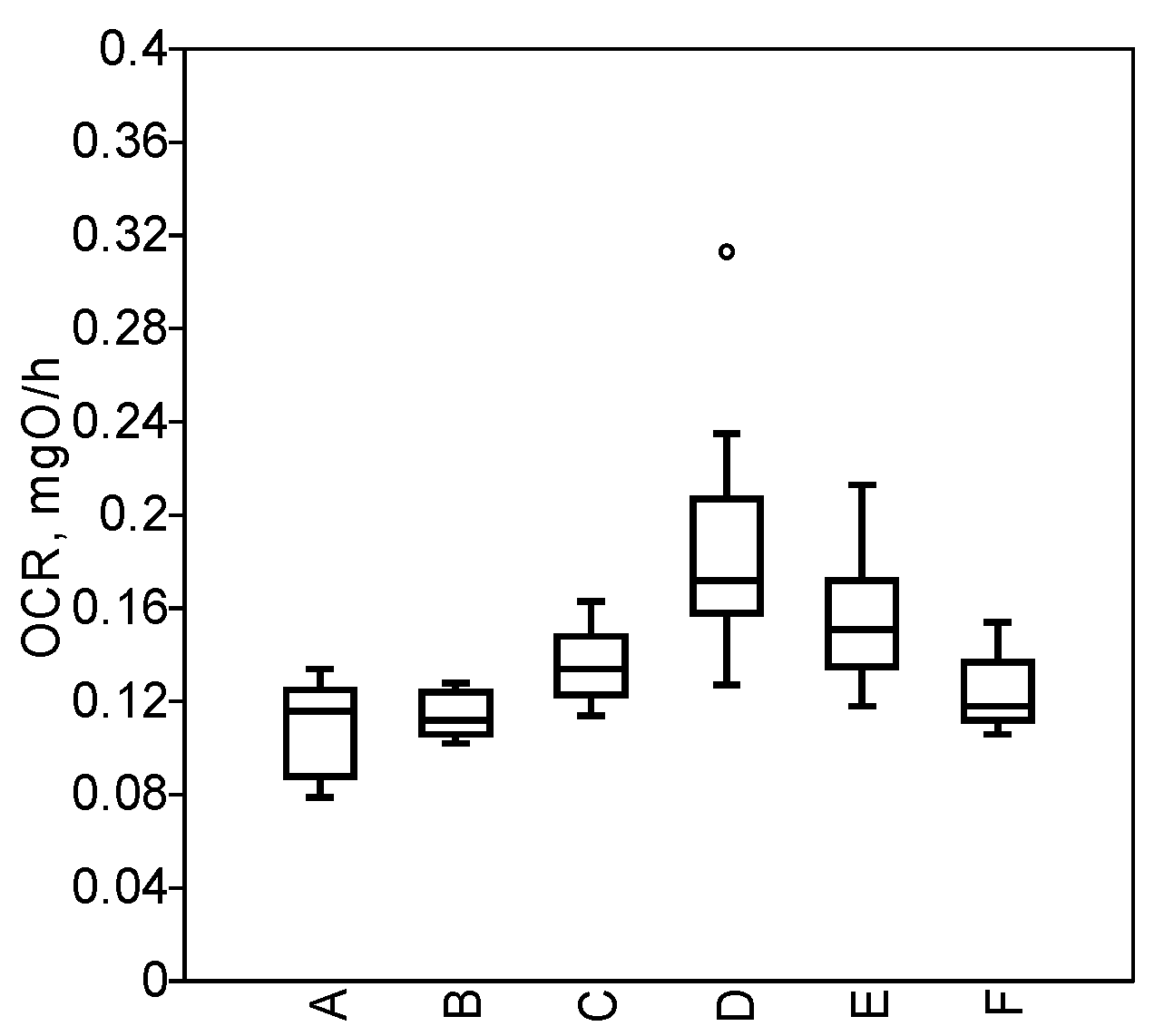
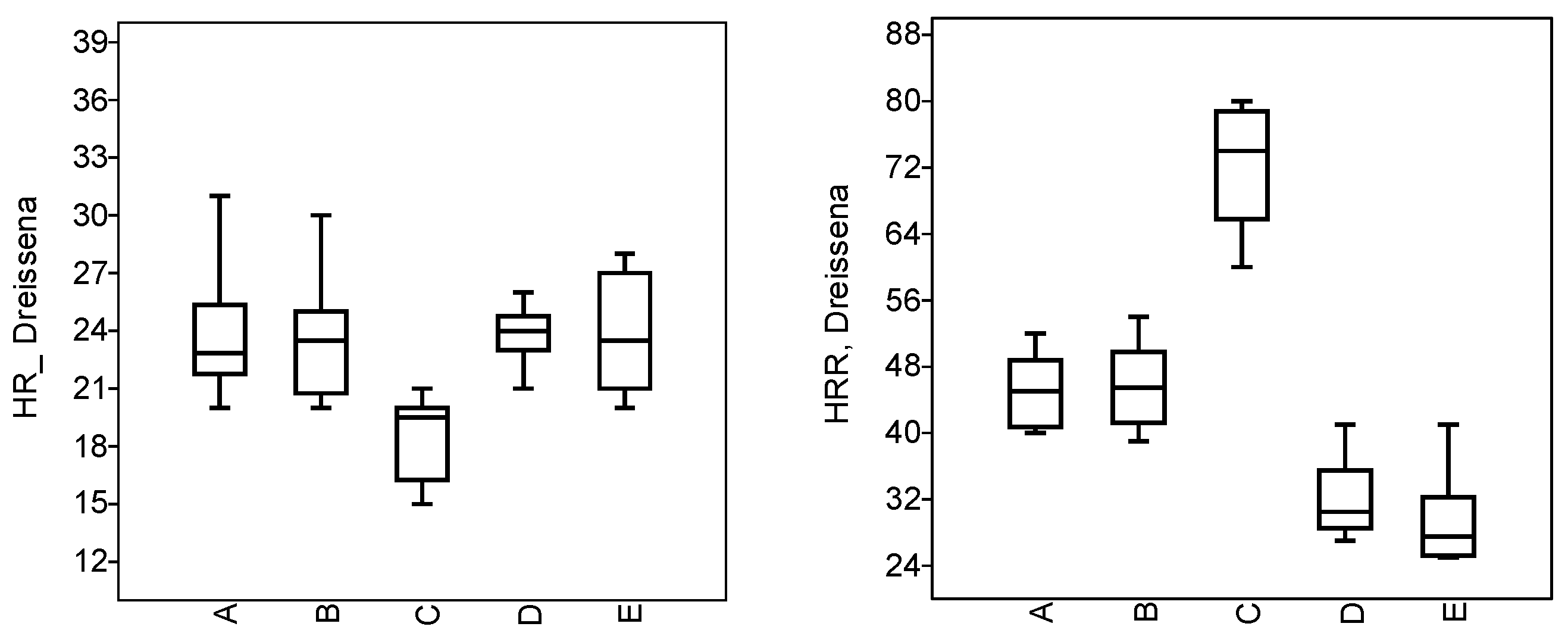
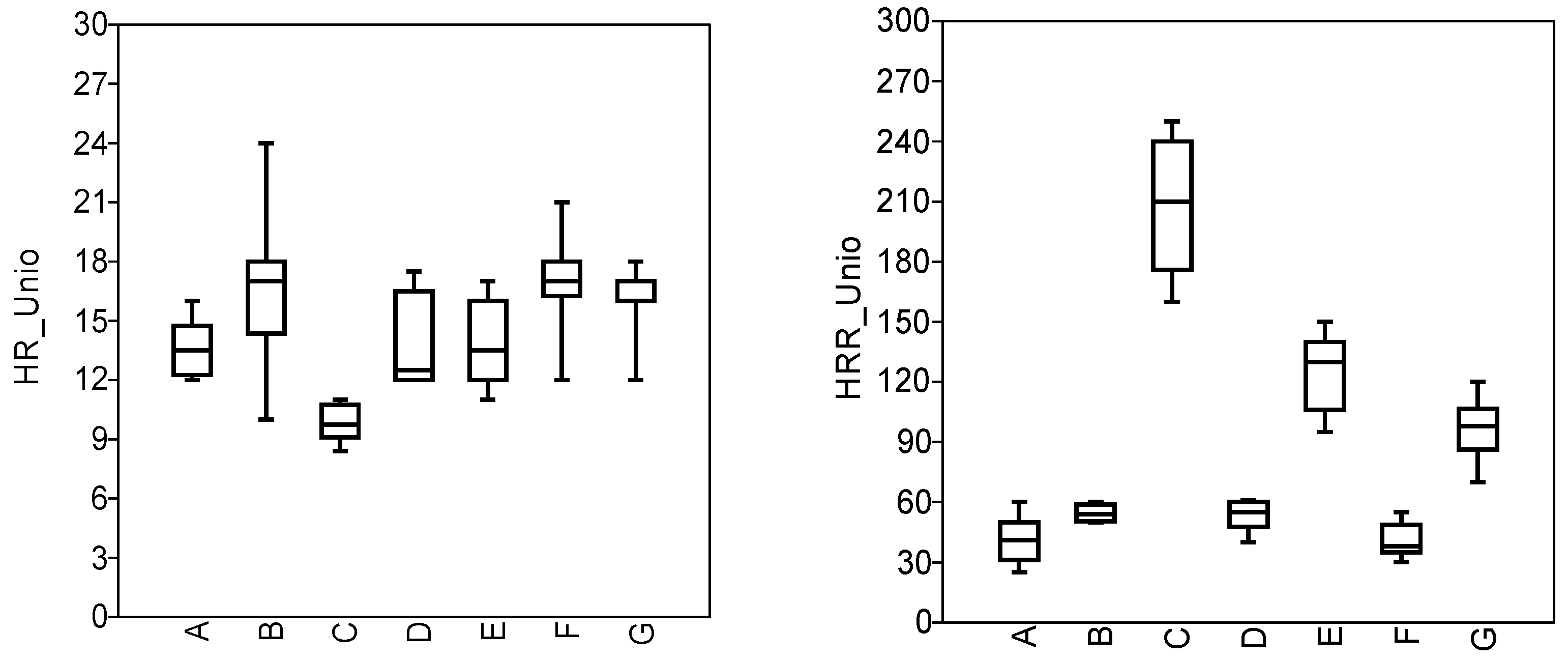
3.3. Physiological State of Molluscs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McLusky, D.S.; Elliott, M. The Estuarine Ecosystem: Ecology, Threats and Management; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Halpern, B.S.; Ebert, C.M.; Kappel, C.V.; Madin, E.M.P.; Micheli, F.; Perry, M.; Selkoe, K.A.; Walbridge, S. Global priority areas for incorporating land–sea connections in marine conservation. Conserv. Let. 2009, 2, 189–196. [Google Scholar] [CrossRef]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef] [PubMed]
- Maximov, A.A.; Berezina, N.A. Benthic Opportunistic Polychaete/Amphipod Ratio: An indicator of pollution or modification of the environment by macroinvertebrates? J. Mar. Sci. Eng. 2023, 11, 190. [Google Scholar] [CrossRef]
- Kuprijanov, I.; Väli, G.; Sharov, A.; Berezina, N.; Liblik, T.; Lips, U.; Kolesova, N.; Maanio, J.; Junttila, V.; Lips, I. Hazardous substances in the sediments and their pathways from potential sources in the eastern Gulf of Finland. Mar. Pollut. Bull. 2021, 170, 112642. [Google Scholar] [CrossRef] [PubMed]
- Saleh, H.N.; Panahande, M.; Yousefi, M.; Asghari, F.B.; Oliveri Conti, G.; Talaee, E.; Mohammadi, A.A. Carcinogenic and non-carcinogenic risk assessment of heavy metals in groundwater wells in Neyshabur Plain, Iran. Biol. Trace Elem. Res. 2019, 190, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Liu, Y.; Feng, X.; Mao, P.; Sun, A.; Wang, M.; Wang, M. Pollution effect assessment of industrial activities on potentially toxic metal distribution in windowsill dust and surface soil in central China. Sci. Total Environ. 2021, 759, 144023. [Google Scholar] [CrossRef]
- Bosch, A.C.; O’Neill, B.; Sigge, G.O.; Kerwath, S.E.; Hoffman, L.C. Heavy metals in marine fish meat and consumer health: A review. J. Sci. Food Agric. 2016, 96, 32–48. [Google Scholar] [CrossRef]
- Ateş, A.; Türkmen, M.; Tepe, Y. Assessment of heavy metals in fourteen marine fish species of four Turkish seas. Indian J. Geo-Mar. Sci. 2015, 44, 49–55. [Google Scholar]
- Senoro, D.B.; De Jesus, K.L.M.; Monjardin, C.E.F. Pollution and Risk Evaluation of Toxic Metals and Metalloid in Water Resources of San Jose, Occidental Mindoro, Philippines. Sustainability 2023, 15, 3667. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic mechanisms of five heavy metals: Mercury, lead, chromium, cadmium, and arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, X.; Zhang, Y. Heavy Metals and Their Ecological Risk Assessment in Surface Sediments of the Changjiang River Estuary and Contiguous East China Sea. Sustainability 2023, 15, 4323. [Google Scholar] [CrossRef]
- Ogunola, O.S. Physiological, immunological, genotoxic and histopathological biomarker responses of molluscs to heavy metal and water quality parameter exposures: A Critical Review. J. Oceanogr. Mar. Sci. 2017, 5, 1. [Google Scholar] [CrossRef]
- DeForest, D.K.; Brix, K.V.; Adams, W.J. Assessing metal bioaccumulation in aquatic environments: The inverse relationship between bioaccumulation factors, trophic transfer factors and exposure concentration. Aquat. Toxicol. 2007, 84, 236. [Google Scholar] [CrossRef] [PubMed]
- Singovszka, E.; Balintova, M.; Demcak, S.; Pavlikova, P. Metal Pollution Indices of Bottom Sediment and Surface Water Affected by Acid Mine Drainage. Metals 2017, 7, 284. [Google Scholar] [CrossRef]
- Gubelit, Y.I.; Shigaeva, T.D.; Kudryavtseva, V.A.; Berezina, N.A. Heavy metal content in macroalgae as a tool for environmental quality assessment: The eastern Gulf of Finland case study. J. Mar. Sci. Eng. 2023, 11, 1640. [Google Scholar] [CrossRef]
- Rajfur, M. Algae—Heavy metal biosorbernt. Ecol. Chem. Eng. 2013, 20, 23–40. [Google Scholar]
- Chakraborty, S.; Bhattacharya, T.; Singh, G.; Maity, J.P. Benthic macroalgae as biological indicators of heavy metal pollution in the marine environments: A biomonitoring approach for pollution assessment. Ecotoxicol. Environ. Saf. 2014, 100, 61–68. [Google Scholar] [CrossRef]
- Da Ros, L.; Nasci, C.; Marigomez, I.; Soto, M. Biomarkers and trace metals in the digestive gland of indigenous and transplanted mussels, Mytilus galloprovincialis, in Venice Lagoon, Italy. Mar. Environ. Res. 2000, 50, 417–423. [Google Scholar] [CrossRef]
- Lehtonen, K.K.; d’Errico, G.; Korpinen, S.; Regoli, F.; Ahkola, H.; Kinnunen, T.; Lastumäki, A. Mussel caging and the weight of evidence approach in the assessment of chemical contamination in coastal waters of Finland (Baltic Sea). Front. Mar. Sci. 2019, 6, 688. [Google Scholar] [CrossRef]
- Schøyen, M.; Allan, I.J.; Ruus, A.; Håvardstun, J.; Hjermann, D.Ø.; Beyer, J. Comparison of caged and native blue mussels (Mytilus edulis spp.) for environmental monitoring of PAH, PCB and trace metals. Mar. Environ. Res. 2017, 130, 221–232. [Google Scholar] [CrossRef]
- Faria, M.; Ochoa, V.; Blazquez, M.; Juan, M.F.S.; Lazzara, R.; Lacorte, S.; Soares, A.M.V.M.; Barata, C. Separating natural from anthropogenic causes of impairment in Zebra mussel (Dreissena polymorpha) populations living across a pollution gradient. Aquat. Toxicol. 2014, 152, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Evariste, L.; David, E.; Cloutier, P.-L.; Brousseau, P.; Auffret, M.; Desrosiers, M.; Groleau, P.E.; Fournier, M.; Betoulle, S. Field biomonitoring using the zebra mussel Dreissena polymorpha and the quagga mussel Dreissena bugensis following immunotoxic reponses. Is there a need to separate the two species? Environ. Pollut. 2018, 238, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, E.; Antal, L.; Stoyanova, S.; Arnaudova, D.; Velcheva, I.; Iliev, I.; Vasileva, T.; Bivolarski, V.; Mitkovska, V.; Chassovnikarova, T.; et al. Biomarkers for pollution in caged mussels from three reservoirs in Bulgaria: A pilot study. Heliyon 2022, 8, e09069. [Google Scholar] [CrossRef]
- Moiseenko, T.I. Bioavailability and ecotoxicity of metals in aquatic systems: Critical levels of pollution. Geochem. Int. 2019, 64, 675–688. [Google Scholar] [CrossRef]
- Kholodkevich, S.V.; Kuznetsova, T.V.; Sharov, A.N.; Kurakin, A.S.; Lips, U.; Kolesova, N.; Lehtonen, K.K. Applicability of a bioelectronic cardiac monitoring system for the detection of biological effects of pollution in bioindicator species in the Gulf of Finland. J. Mar. Syst. 2017, 171, 151–158. [Google Scholar] [CrossRef]
- Kholodkevich, S.; Sharov, A.; Chuiko, G.; Kuznetsova, T.; Gapeeva, M.; Lozhkina, R. Quality assessment of freshwater ecosystems by the functional state of bivalved mollusks. Water Resour. 2019, 46, 249–257. [Google Scholar] [CrossRef]
- Berezina, N.A.; Strode, E.; Lehtonen, K.K.; Balode, M.; Golubkov, S.M. Sediment quality assessment using Gmelinoides fasciatus and Monoporeia affinis (Amphipoda, Gammaridea) in the northeastern Baltic Sea. Crustaceana 2013, 86, 780–801. [Google Scholar] [CrossRef]
- Depledge, M.H.; Lundebye, A.K.; Curtis, T.; Aagaard, A.; Andersen, B.B. Automated interpulse-durationassessment (AIDA): A new technique for detecting disturbances in cardiac activity in selected invertebrates. Mar. Biol. 1996, 126, 313–319. [Google Scholar] [CrossRef]
- Golubkov, M.; Golubkov, S. Eutrophication in the Neva Estuary (Baltic Sea): Response to temperature and precipitation patterns. Mar. Freshw. Res. 2020, 71, 583–595. [Google Scholar] [CrossRef]
- Polyak, Y.M.; Berezina, N.A.; Polev, D.E.; Sharov, A.N. The state of the intestinal bacterial community in mollusks for assessing habitat pollution in the Gulf of Finland (Baltic Sea). Estuar. Coast. Shelf Sci. 2022, 278, 108095. [Google Scholar] [CrossRef]
- Kostamo, K.; Blomster, J.; Korpelainen, H.; Kelly, J.; Maggs, C.A.; Mineur, F. New microsatellite markers for Ulva intestinalis (Chlorophyta) and the transferability of markers across species of Ulvaceae. Phycologia 2008, 47, 580–587. [Google Scholar] [CrossRef]
- Green-Gavrielidis, L.A.; MacKechnie, F.; Thornber, C.S.; Gomez-Chiarri, M. Bloom-forming macroalgae (Ulva spp.) inhibit the growth of co-occurring macroalgae and decrease eastern oyster larval survival. Mar. Ecol. Prog. Ser. 2018, 595, 27–37. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, Z.; Wu, Y.; Deng, Y.; Chen, Q.; Zhao, C.; Cui, L.; Huang, X. Macroalgae bloom decay decreases the sediment organic carbon sequestration potential in tropical seagrass meadows of the South China Sea. Mar. Pollut. Bull. 2019, 138, 598–603. [Google Scholar] [CrossRef]
- Lanari, M.; Copertino, M. Drift macroalgae in the Patos Lagoon Estuary (southern Brazil): Effects of climate, hydrology and wind action on the onset and magnitude of blooms. Mar. Biol. Res. 2017, 13, 36–47. [Google Scholar] [CrossRef]
- Jorhem, L.; Engman, J.; Sundström, B.; Thim, A.M. Trace elements in crayfish: Regional differences and changes induced by cooking. Arch. Environ. Contam. Toxicol. 1994, 26, 137–142. [Google Scholar] [CrossRef]
- Kholodkevich, S.V.; Ivanov, A.V.; Kurakin, A.S.; Kornienko, E.L.; Fedotov, V.P. Real time biomonitoring of surface water toxicity level at water supply stations. J. Environ. Bioindic. 2008, 3, 23–34. [Google Scholar] [CrossRef]
- Gubelit, Y.; Polyak, Y.; Dembska, G.; Pazikowska-Sapota, G.; Zegarowski, L.; Kochura, D.; Krivorotov, D.; Podgornaya, E.; Burova, O.; Maazouzi, C. Nutrient and metal pollution of the eastern Gulf of Finland coastline: Sediments, macroalgae, microbiota. Sci. Total Environ. 2016, 550, 806–819. [Google Scholar] [CrossRef] [PubMed]
- Zalewska, T.; Danowska, B. Marine environment status assessment based on macrophytobenthic plants as bio-indicators of heavy metals pollution. Mar. Pollut. Bull. 2017, 118, 281–288. [Google Scholar] [CrossRef]
- Polyak, Y.; Gubelit, Y.; Bakina, L.; Shigaeva, T.; Kudryavtseva, V. Impact of macroalgal blooms on biogeochemical processes in estuarine systems: A case study in the eastern Gulf of Finland, Baltic Sea. J. Soils Sediments 2023, 23. [Google Scholar] [CrossRef]
- Vallius, H.; Leivuori, M. Classification of Heavy metal contaminated sediments of the Gulf of Finland. Baltica 2003, 16, 3–12. [Google Scholar]
- Marine Habitat Committee. Report of the Working Group on Marine Sediments in Relation to Pollution; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2003; Available online: https://www.ices.dk/sites/pub/CM%20Doccuments/2003/E/E0403.PDF (accessed on 23 August 2023).
- Oros, A.; Gomoiu, M.-T. Comparative data on the accumulation of five heavy metals (cadmium, chromium, copper, nickel, lead) in some marine species (molluscs, fish) from the Romanian sector of the Black Sea. Cercet. Mar. 2010, 39, 89–108. [Google Scholar]
- Damir, N.-A.; Danilov, D.A.; Oros, A.; Lazăr, L.; Coatu, V. Chemical status evaluation of the Romanian Black Sea marine environment based on benthic organisms’ contamination. Cercet. Mar. 2022, 52, 52–77. [Google Scholar] [CrossRef]
- Besada, V.; Fumega, J.; Vaamonde, A. Temporal trends of Cd, Cu, Hg, Pb and Zn in mussel (Mytilus galloprovincialis) from the Spanish North-Atlantic coast 1991–1999. Sci. Total Environ. 2002, 288, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Bryan, G.W. Concentrations of zinc and copper in the tissues of decapod crustaceans. J. Mar. Biol. Assoc. UK 1968, 48, 303–321. [Google Scholar] [CrossRef]
- Ferns, P.N. Birds of the Bristol Channel and Severn Estuary. Mar. Pollut. Bull. 1984, 15, 76–81. [Google Scholar] [CrossRef]
- Rzymski, P.; Niedzielski, P.; Klimaszyk, P.; Poniedziałek, B. Bioaccumulation of selected metals in bivalves (Unionidae) and Phragmites australis inhabiting a municipal water reservoir. Environ. Monit. Assess. 2014, 186, 3199–3212. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, F.E.A.; Minervino, A.H.H.; Miranda, M.; Herrero-Latorre, C.; Barrêto Júnior, R.A.; Oliveira, F.L.C.; Sucupira, M.C.A.; Ortolani, E.L.; López-Alonso, M. Toxic and essential trace element concentrations in fish species in the Lower Amazon, Brazil. Sci. Total Environ. 2020, 732, 138983. [Google Scholar] [CrossRef]
- Depledge, M.H.; Rainbow, P.S. Models of regulation and accumulation of trace metals in marine invertebrates. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 1990, 97, 1–7. [Google Scholar] [CrossRef]
- Amiard, J.C.; Triquet, C.; Berthet, B.; Metayer, C. Comparative study of the patterns of bioaccumulation of essential (Cu, Zn) and non-essential (Cd, Pb) trace metals in various estuarine and coastal organisms. J. Exp. Mar. Biol. Ecol. 1987, 106, 73–89. [Google Scholar] [CrossRef]
- Ansari, T.M.; Marr, I.L.; Tariq, N. Heavy metals in marine pollution perspective—A mini review. J. Appl. Sci. 2004, 4, 1–20. [Google Scholar] [CrossRef]
- Cordeli, A.N.; Oprea, L.; Crețu, M.; Dediu, L.; Coadă, M.T.; Mînzală, D.-N. Bioaccumulation of Metals in Some Fish Species from the Romanian Danube River: A Review. Fishes 2023, 8, 387. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, F.; Yan, X. Eutrophication likely prompts metal bioaccumulation in edible clams. Ecotoxicol. Environ. Saf. 2021, 224, 112671. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, P.S.; Poirier, L.; Smith, B.D.; Brix, K.V.; Luoma, S.N. Trophic transfer of trace metals from the polychaete worm Nereis diversicolor to the polychaete N. virens and the decapod crustacean Palaemonetes varians. Mar. Ecol. Prog. Ser. 2006, 321, 167–181. [Google Scholar] [CrossRef]
- Voigt, H.-R. Heavy metal concentrations in four-horn sculpin Triglopsis quadricornis (L.) (Pisces), its main food organism Saduria entomon L. (Crustacea), and in bottom sediments in the Archipelago Sea and the Gulf of Finland (Baltic Sea). Proc. Est. Acad. Sci. Biol. Ecol. 2007, 56, 224–238. [Google Scholar] [CrossRef]
- Curtis, T.M.; Williamson, R.; Depledge, M.H. Simultaneous, long-term monitoring of valve and cardiac activity in the blue mussel Mytilus edulis exposed to copper. Mar. Biol. 2000, 136, 837–846. [Google Scholar] [CrossRef]
- Xing, Q.; Zhang, L.; Li, Y.; Zhu, X.; Li, Y.; Guo, H.; Bao, Z.; Wang, S. Development of novel cardiac indices and assessment of factors affecting cardiac activity in a bivalve mollusc Chlamys farreri. Front. Physiol. 2019, 10, 293. [Google Scholar] [CrossRef]
- Bakhmet, I.N. Cardiac activity and oxygen consumption of blue mussels (Mytilus edulis) from the White Sea in relation to body mass, ambient temperature and food availability. Polar Biol. 2017, 40, 1959–1964. [Google Scholar] [CrossRef]
- Burnett, N.P.; Seabra, R.; De Pirro, M.; Wethey, D.S.; Woodin, S.A.; Helmuth, B.; Zippay, M.L.; Sara, G.; Monaco, C.; Lima, F.P. An improved non-invasive method for measuring heartbeat of intertidal animals. Limnol. Oceanogr.-Meth. 2013, 11, 91–100. [Google Scholar] [CrossRef]
- Depledge, M.H. Disruption of circulatory and respiratory activity in shore crabs (Carcinus maenas) exposed to heavy metal pollution. Comp. Biochem. Physiol. C 1984, 78, 445–459. [Google Scholar] [CrossRef]
- Cheung, S.G.; Cheung, R.Y.H. Effects of heavy metals on oxygen consumption and ammonia excretion in green-lipped mussels (Perna viridis). Mar. Pollut. Bull. 1995, 31, 381–386. [Google Scholar] [CrossRef]
- Sobrino-Figueroa, A.S.; Cáceres-Martinez, C. Evaluation of the effects of the metals Cd, Cr, Pb and their mixture on the filtration and oxygen consumption rates in catarina scallop, Argopecten ventricosus juveniles. J. Environ. Biol. 2014, 35, 1–8. [Google Scholar] [PubMed]
- Capparelli, M.V.; Abessa, D.M.; McNamara, J.C. Effects of metal contamination in situ on osmoregulation and oxygen consumption in the mudflat fiddler crab Uca rapax (Ocypodidae, Brachyura). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 185–186, 102–111. [Google Scholar] [CrossRef]
- Bakhmet, I.; Fokina, N.; Ruokolainen, T. Changes of heart rate and lipid composition in Mytilus edulis and Modiolus modiolus caused by crude oil pollution and low salinity effects. J. Xenobiot. 2021, 11, 46–60. [Google Scholar] [CrossRef]
- Bakhmet, I.N.; Kantserova, N.P.; Lysenko, L.A.; Nemova, N.N. Effect of copper and cadmium ions on heart function and calpain activity in blue mussel Mytilus edulis. J. Environ. Sci. Health A 2012, 47, 1528–1535. [Google Scholar] [CrossRef]
- Azizi, G.; Akodad, M.; Baghour, M.; Mostafa, L.; Moumen, A. The use of Mytilus spp. mussels as bioindicators of heavy metal pollution in the coastal environment. A review. J. Mater. Environ. Sci. 2018, 9, 1170–1181. [Google Scholar]
- Beggel, S.; Hinzmann, M.; Machado, J.; Geist, J. Combined impact of acute exposure to ammonia and temperature stress on the freshwater mussel Unio pictorum. Water 2017, 9, 455. [Google Scholar] [CrossRef]
- Vereycken, J.E.; Aldridge, D.C. Bivalve molluscs as biosensors of water quality: State of the art and future directions. Hydrobiologia 2023, 850, 231–256. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name | Method | Latitude Longitude | H | Sal | TP | THC | Corg | 16PAHs | Ma | Surv | State |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Zimino | Caged molluscs Sediment, Fish | 60.6348 28.6620 | 4 | 1.7 | 25 | 0.016 | 0.3 | 64 | <0.1 | 90 | GES |
| 2 | Primorsk | Caged molluscs Sediment, Macroalgae | 60.3605 28.6097 | 6 | 2.7 | 34 | 0.02 | 0.5 | 595 | 87 (10) | 60 | Sub-GES |
| 3 | Okunevaya | Sediment, Fish | 60.2352 28.9913 | 4 | 2.5 | 25 | 0.008 | 0.45 | 49 | <0.1 | 90 | GES |
| 4 | Flotsky | Sediment, Fish, Macroalgae | 60.1613 29.1563 | 1 | 1.2 | 25 | 0.012 | 0.5 | 91 | 97 (31) | 90 | GES |
| 5 | Serovo | Sediment, Field molluscs, Macroalgae | 60.196 29.567 | 1 | 0.8 | 27 | 0.01 | 0.8 | 57 | 35 (52) | 80 | GES |
| 6 | Dubki | Caged molluscs, Sediment, Fish | 60.0896 29.9195 | 2 | 0.3 | 50 | 0.011 | 0.8 | 49 | 47 (33) | 95 | GES |
| 7 | Petergof WPT | Sediment, Field molluscs, Macroalgae | 59.8888 29.9033 | 1 | 0.2 | 43 | 0.009 | 0.9 | 250 | 19 (55) | 65 | Sub-GES |
| 8 | Lomonosov beach | Sediment, Field molluscs, Macroalgae | 59.9193 29.7796 | 1 | 0.4 | 47 | 0.01 | 0.8 | 65 | 15 (40) | 70 | Sub-GES |
| 9 | Dam, Port | Caged molluscs, Sediment | 59.9812 29.6922 | 5.5 | 0.67 | 25 | 0.024 | 0.96 | 1033 | 10 (30) | 60 | Sub-GES |
| 10 | Grafskaya | Caged molluscs, Sediment, Macroalgae | 59.9858 29.2014 | 3 | 2.6 | 48 | 0.023 | 0.45 | 725 | 80 (17) | 20 | BES |
| 11 | River Sista mouth | Caged molluscs, Sediment, Macroalgae | 59.8092 28.9104 | 2 | 3.2 | 30 | 0.032 | 0.59 | 314 | 17 (30) | 80 | GES |
| 12 | Luga Bay | Sediment, Macroalgae | 59.8309 28.4969 | 1 | 3.8 | 25 | 0.008 | 0.29 | <16 | 67 (32) | 90 | GES |
| Metal Metalloid | St 1 | St 2 | St 3 | St 4 | St 5 | St 6 | St 7 | St 8 | St 9 | St 10 | St 11 | St 12 | MAC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sediment | |||||||||||||
| Cd | 1.6 | 0.38 | 0.12 | 0.18 | 0.2 | 0.2 | 0.39 | 0.42 | 0.39 | 0.16 | <0.05 | 0.24 | 1 |
| Pb | 42 | 13 | 2.9 | 7.9 | 11 | 5 | 2.2 | 7.8 | 8.7 | 4.2 | 1.5 | 1.7 | 10 |
| Zn | 250 | 33 | 16 | 24 | 26 | 26 | 20 | 33 | 34 | 10 | 7.7 | 4.2 | 199 |
| Cu | 46 | 7.1 | 1.6 | 4.4 | 6.7 | 5.1 | 20 | 12 | 10 | 1.3 | 1.4 | 0.84 | 50 |
| Hg | 0.005 | 0.032 | 0.17 | 0.022 | 0.005 | 0.005 | 0.09 | 0.08 | 0.032 | 0.005 | 0.005 | 0.005 | 0.05 |
| As | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.20 | 0.05 | 0.05 | 0.05 | 0.38 | 0.5 |
| Molluscs | |||||||||||||
| Cd | 4.7 | 2.7 | - | 0.4 | 2.6 | 2.2 | 2.1 | 2 | 0.8 | 1.8 | 1.9 | 2.1 | 5.00 |
| Pb | 0.92 | 0.68 | - | 1.2 | 6.9 | 0.68 | 2.5 | 1.7 | 1.1 | 0.1 | 3.1 | 1.7 | 7.50 |
| Zn | 150 | 108 | - | 14.8 | 47 | 116 | 200 | 170 | 25 | 5 | 23 | 21 | 170 |
| Cu | 5.3 | 5.2 | - | 3 | 10.2 | 3.4 | 20 | 6.9 | 1.6 | 0.56 | 8.8 | 2.8 | 50 |
| Hg | 0.001 | 0.001 | - | 0.05 | 0.1 | 0.14 | 0.06 | 0.04 | 0.05 | 0.04 | 0.06 | 0.001 | 0.1 |
| Macroalgae | |||||||||||||
| Cd | - | 3.6 | - | 0.4 | 2.6 | 4.9 | 0.18 | 5.6 | 3.5 | 1.3 | 1.4 | 1.6 | 5.00 |
| Pb | - | 10 | - | 1.02 | 6.9 | 3.2 | <0.1 | 1.0 | 22 | 5.9 | 2.1 | 1.1 | 7.50 |
| Zn | - | 45 | - | 12.8 | 47 | 27 | 7.9 | 16 | 40 | 28 | 23 | 11.1 | 170 |
| Cu | - | 11 | - | 3 | 10.2 | 5.6 | 0.37 | 3.9 | 8.7 | 7.9 | 5.8 | 3.8 | 50 |
| Fish | |||||||||||||
| Cd | 10 | 11 | 21 | 4 | 5.2 | - | 0.11 | 0.18 | - | - | - | - | 5.00 |
| Pb | 0.17 | 0.1 | 0.1 | 0.8 | 0.95 | - | 0.1 | 0.1 | - | - | - | - | 7.50 |
| Zn | 73 | 41 | 44 | 108 | 112 | - | 5 | 7.9 | - | - | - | - | 170 |
| Cu | 0.8 | 0.6 | 1 | 5 | 4.4 | - | 0.37 | 0.37 | - | - | - | - | 50 |
| Hg | 0.14 | 0.25 | 0.21 | 0.22 | 0.24 | - | 0.21 | 0.05 | - | - | - | - | 0.1 |
| Metals | Cd | Pb | Zn | Cu | Hg |
|---|---|---|---|---|---|
| Macroalgae | |||||
| Mean | 25.5 | 24.2 | 3.1 | 3.8 | - |
| Standard Error | 7.6 | 8.5 | 0.8 | 1.6 | - |
| Median | 18.5 | 14.0 | 1.8 | 1.4 | - |
| Standard Deviation | 26.4 | 29.4 | 2.9 | 5.4 | - |
| Minimum | 0.5 | 0.2 | 0.4 | 0.0 | - |
| Maximum | 100.0 | 100.0 | 9.1 | 15.6 | - |
| 95% Confidence Level | 16.8 | 18.7 | 1.8 | 3.4 | - |
| Unionid molluscs | |||||
| Mean | 24.2 | 0.9 | 5.5 | 2.7 | 9.2 |
| Standard Error | 8.5 | 0.4 | 1.4 | 1.1 | 3.5 |
| Median | 14.0 | 0.3 | 5.3 | 1.2 | 2.2 |
| Standard Deviation | 29.4 | 1.3 | 4.8 | 3.9 | 12.1 |
| Minimum | 0.2 | 0.1 | 0.8 | 0.2 | 0.1 |
| Maximum | 100.0 | 3.5 | 17.2 | 11.5 | 35.0 |
| 95% Confidence Level | 18.7 | 0.8 | 3.1 | 2.5 | 7.7 |
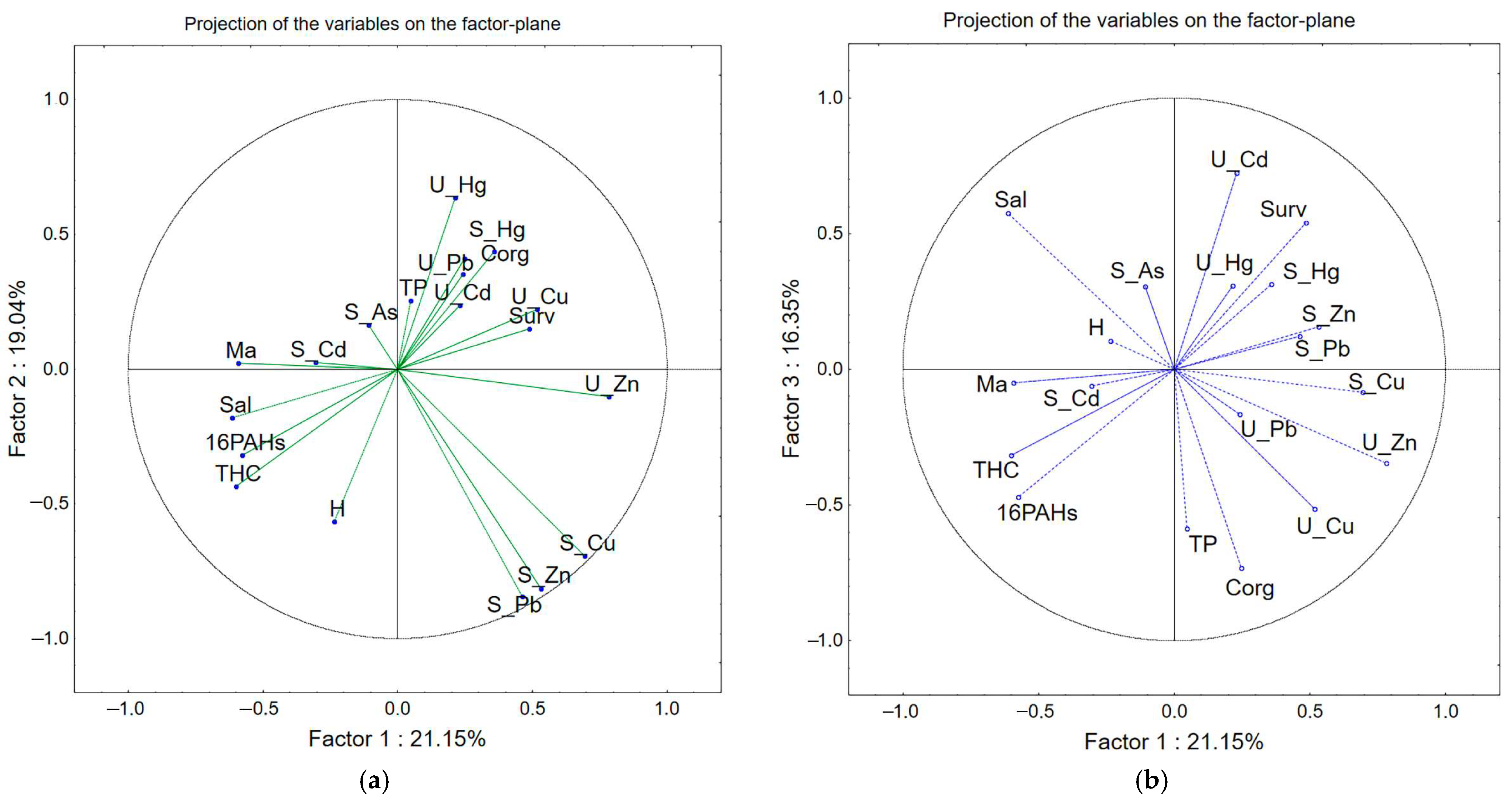
| Variable | Factor 1 | Factor 2 | Factor 3 |
|---|---|---|---|
| H | −0.24 | −0.57 | 0.1 |
| Sal | −0.61 | −0.18 | 0.58 |
| TP | 0.05 | 0.25 | −0.59 |
| THC | −0.6 | −0.44 | −0.32 |
| Corg | 0.25 | 0.41 | −0.73 |
| 16PAHs | −0.58 | −0.32 | −0.47 |
| Ma | −0.59 | 0.02 | −0.05 |
| Surv | 0.49 | 0.15 | 0.54 |
| S_Cd | −0.3 | 0.02 | −0.06 |
| S_Pb | 0.46 | −0.84 | 0.12 |
| S_Zn | 0.53 | −0.81 | 0.16 |
| S_Cu | 0.69 | −0.69 | −0.09 |
| S_Hg | 0.36 | 0.44 | 0.31 |
| S_As | −0.11 | 0.16 | 0.31 |
| U_Cd | 0.23 | 0.24 | 0.72 |
| U_Pb | 0.24 | 0.35 | −0.17 |
| U_Zn | 0.78 | −0.1 | −0.35 |
| U_Cu | 0.52 | 0.22 | −0.52 |
| U_Hg | 0.21 | 0.64 | 0.31 |
| Caged Unionid Molluscs | |||||||
| PAH | St 1 | St 2 | St 6 | St 9 | St 10 | St 11 | Ref. |
| Benzo-a-pyrene | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 |
| Benzo-a-anthracene | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| Benz-β-fluorantene | <2 | <2 | <2 | 36 | 5.8 | 39 | <2 |
| Benz-k-fluorantene | <0.5 | 1 | 1.2 | 5.7 | 10 | 1.4 | <0.5 |
| Benz(-g, h, i-)perylene | <0.5 | <0.5 | <0.5 | 6.2 | <0.5 | 2.8 | <0.5 |
| Dibenz[a,h]anthracene | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| Indeno (1,2,3-cd) pyrene | <2 | <2 | <2 | <2 | <2 | <2 | <2 |
| Pyrene | <2 | <2 | <2 | <2 | <2 | <2 | <2 |
| Chrysene | <2 | <2 | <2 | <2 | <2 | <2 | <2 |
| Field Fish and Molluscs | |||||||
| Site | St 3 | St 4 | St 5 | St 6 | St 7 | St 8 | Ref. |
| Benzo-a-pyrene | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | 0.62 | <0.5 |
| Benzo-a-anthracene | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| Benz-β-fluorantene | <2 | 1.5 | <2 | <2 | <2 | 38 | <2 |
| Benz-k-fluorantene | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | 10 | <0.5 |
| Benz(-g, h, i-)perylene | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 | <0.5 |
| Dibenz[a,h]anthracene | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| Indeno (1,2,3-cd) pyrene | <2 | <2 | <2 | <2 | <2 | <2 | <2 |
| Pyrene | <2 | <2 | <2 | <2 | <2 | <2 | <2 |
| Chrysene | <2 | <2 | <2 | <2 | <2 | <2 | <2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berezina, N.; Maximov, A.; Sharov, A.; Gubelit, Y.; Kholodkevich, S. Environmental Assessment with Cage Exposure in the Neva Estuary, Baltic Sea: Metal Bioaccumulation and Physiologic Activity of Bivalve Molluscs. J. Mar. Sci. Eng. 2023, 11, 1756. https://doi.org/10.3390/jmse11091756
Berezina N, Maximov A, Sharov A, Gubelit Y, Kholodkevich S. Environmental Assessment with Cage Exposure in the Neva Estuary, Baltic Sea: Metal Bioaccumulation and Physiologic Activity of Bivalve Molluscs. Journal of Marine Science and Engineering. 2023; 11(9):1756. https://doi.org/10.3390/jmse11091756
Chicago/Turabian StyleBerezina, Nadezhda, Alexey Maximov, Andrey Sharov, Yulia Gubelit, and Sergei Kholodkevich. 2023. "Environmental Assessment with Cage Exposure in the Neva Estuary, Baltic Sea: Metal Bioaccumulation and Physiologic Activity of Bivalve Molluscs" Journal of Marine Science and Engineering 11, no. 9: 1756. https://doi.org/10.3390/jmse11091756