
Past Analogues of Deoxygenation Events in the Mediterranean Sea: A Tool to Constrain Future Impacts
 , , , and
, , , and 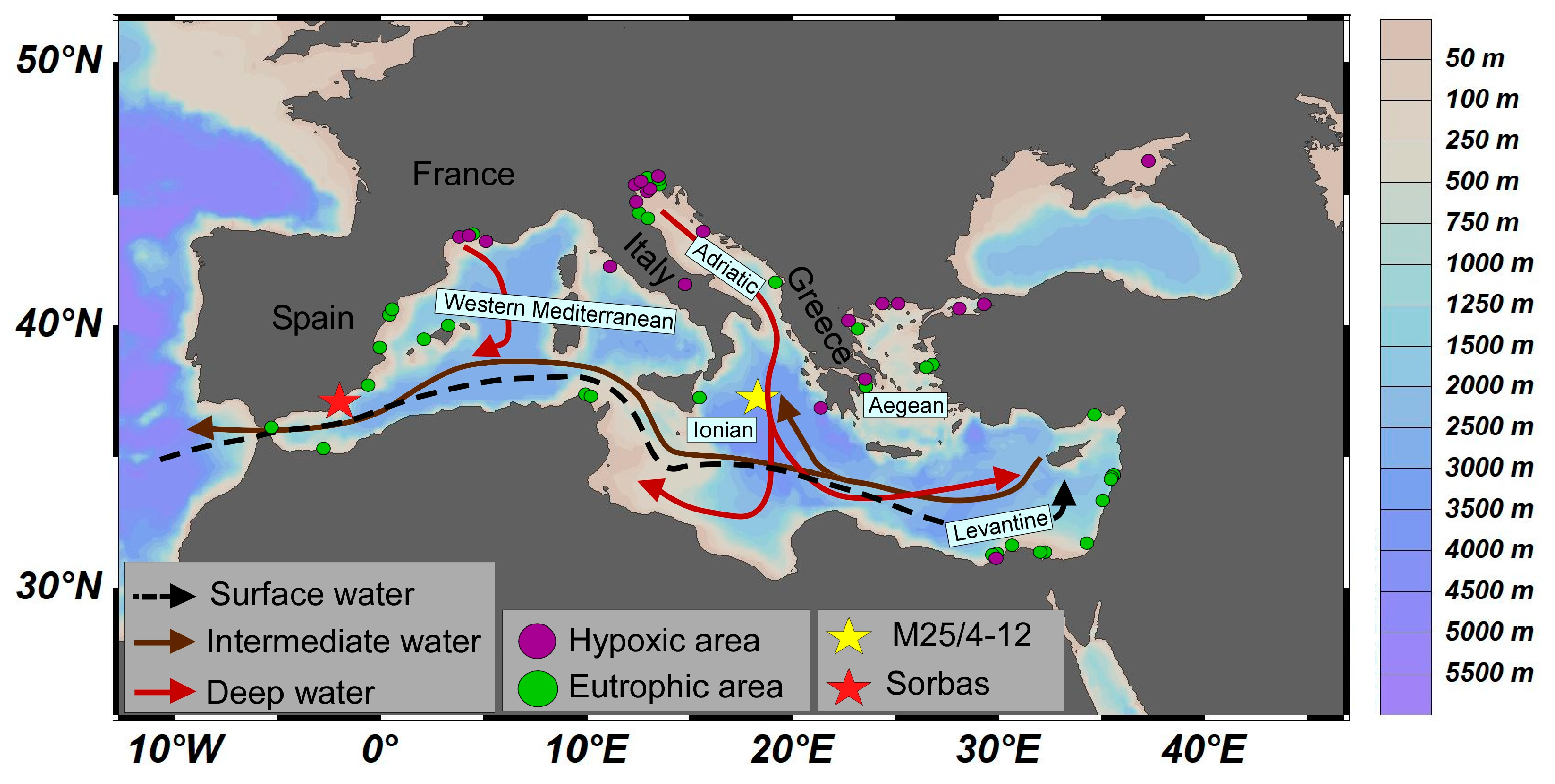{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Oceanographic and Climatic Framework
Sapropel and Deep-Sea Anoxia
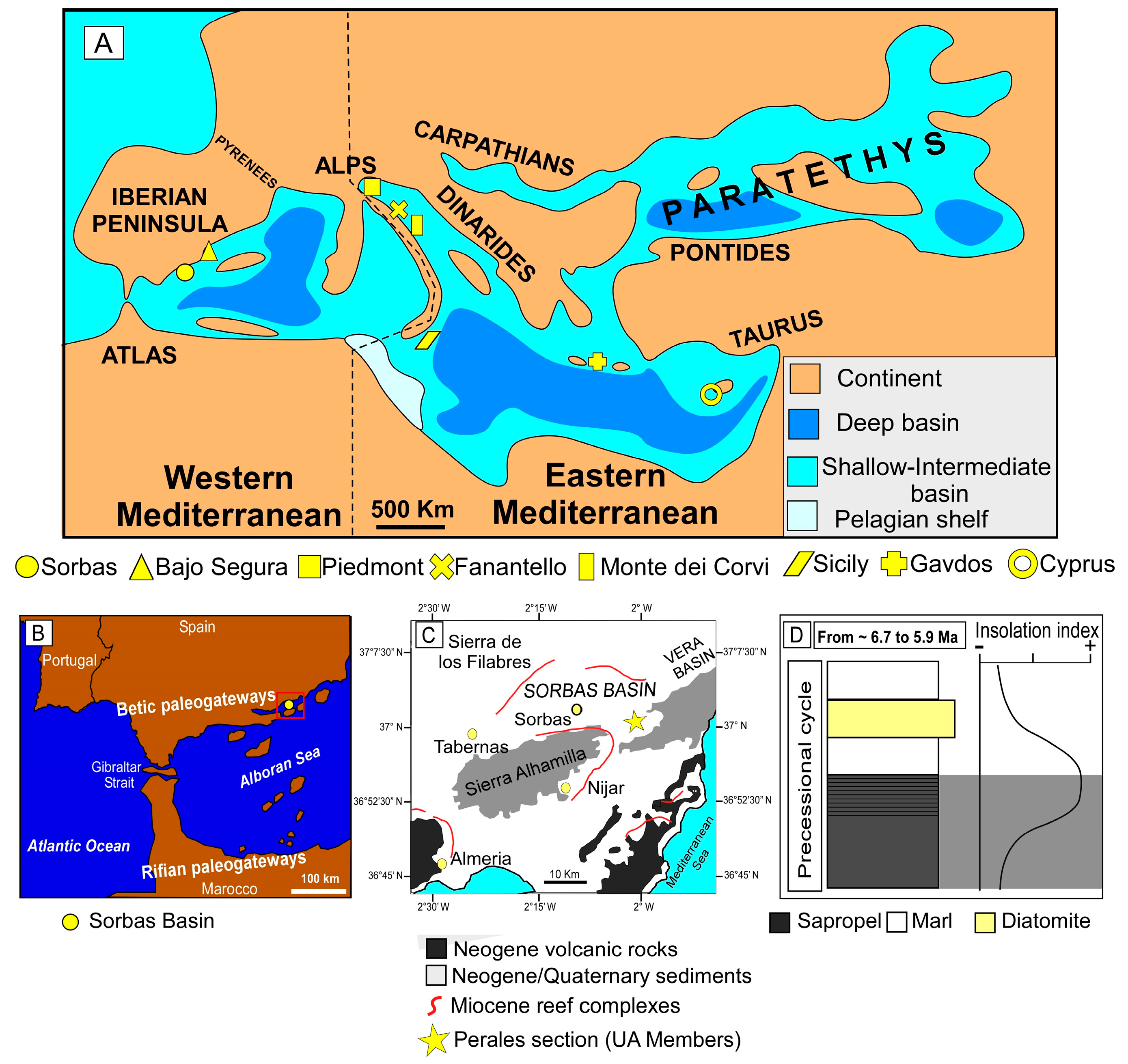
3. Material and Methods
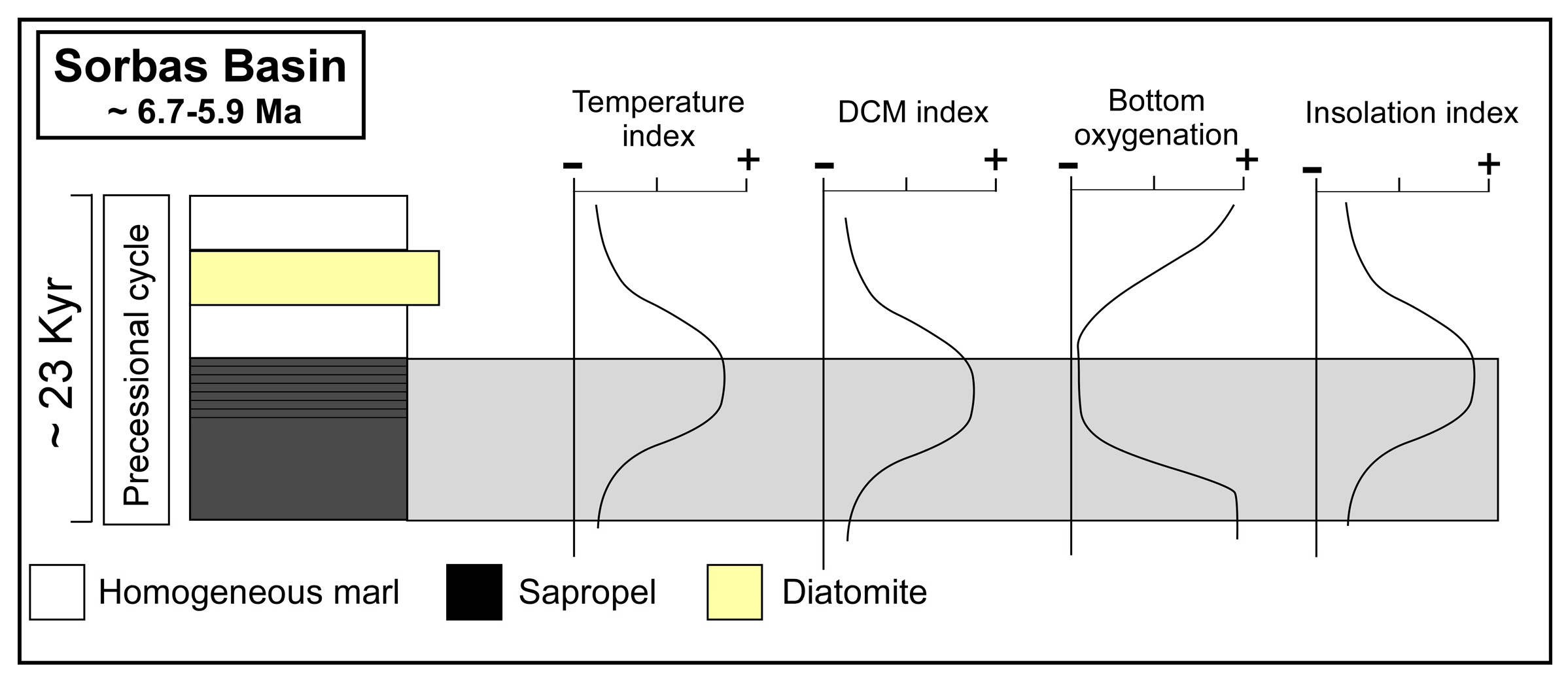
3.1. The Sapropels of the Sorbas Basin
3.2. The Sapropel S1 in the M25/4-12 Core
3.3. Micropaleontological-Based Indexes as a Proxy of Deoxygenation Event
4. Results
4.1. Micropaleontological Signature of the Sorbas Basin Sapropel
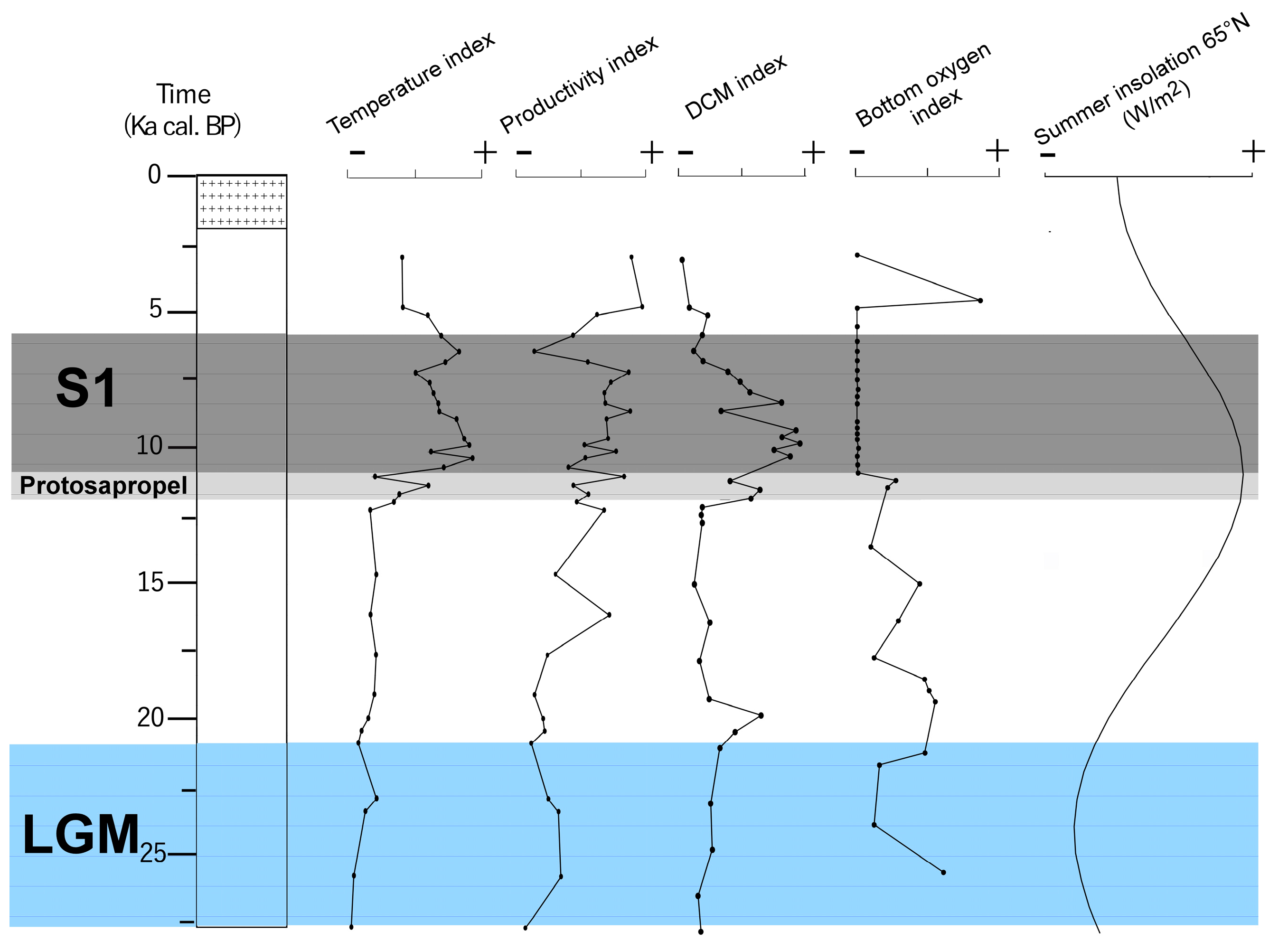
4.2. Micropaleontological Signature of Sapropel S1
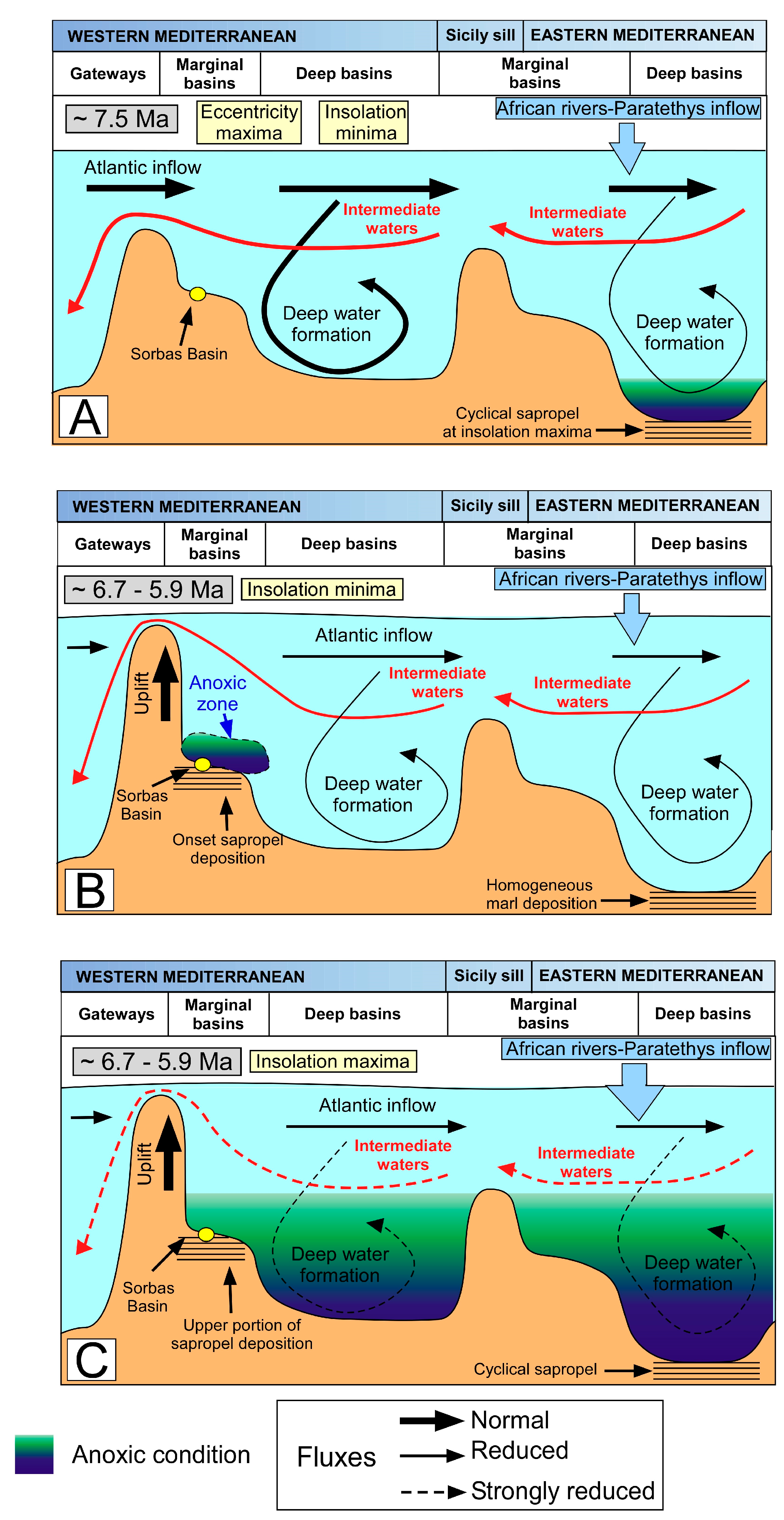
5. Discussion
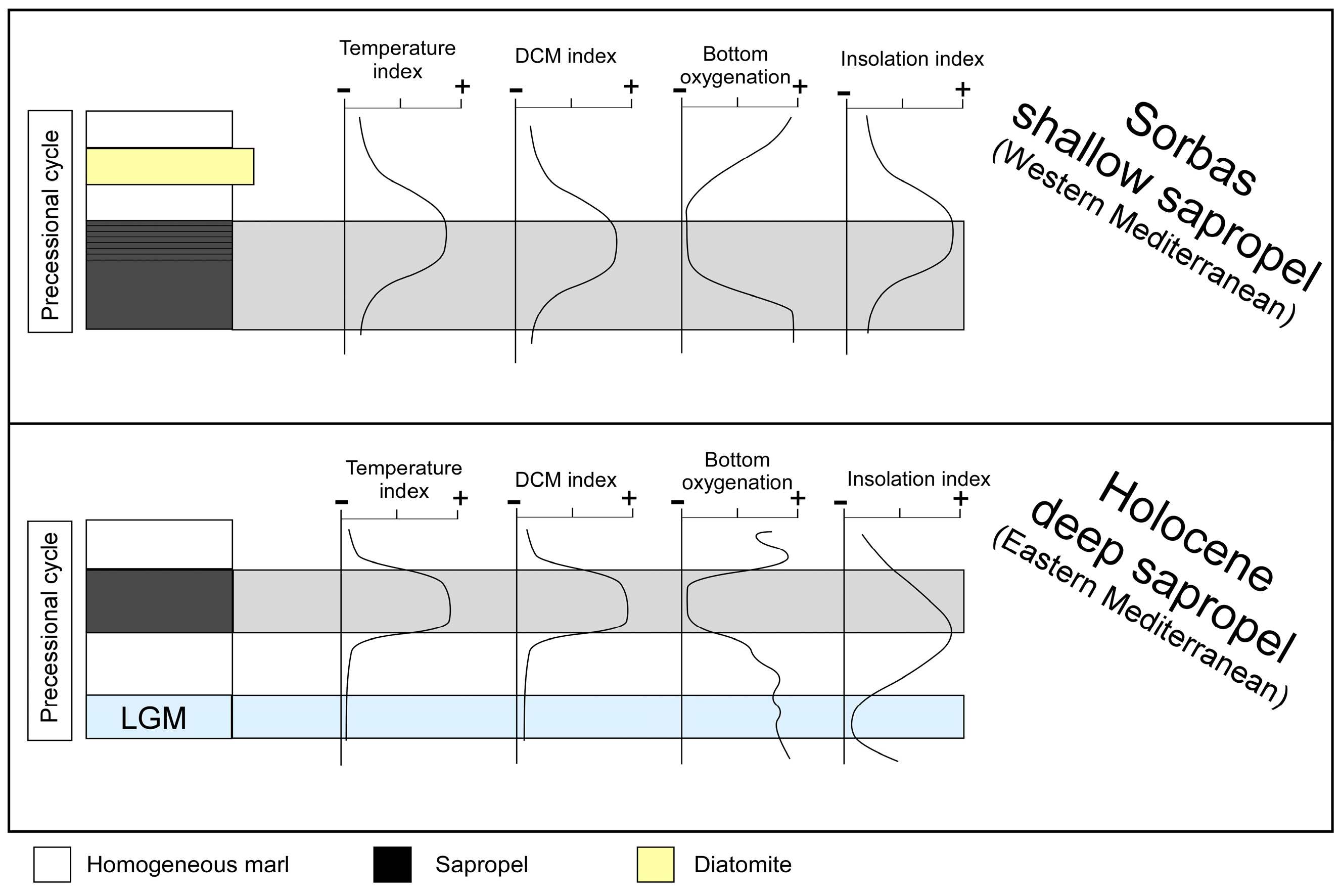
5.1. Paleoceanographic Changes of Messinian and Holocene Sapropels
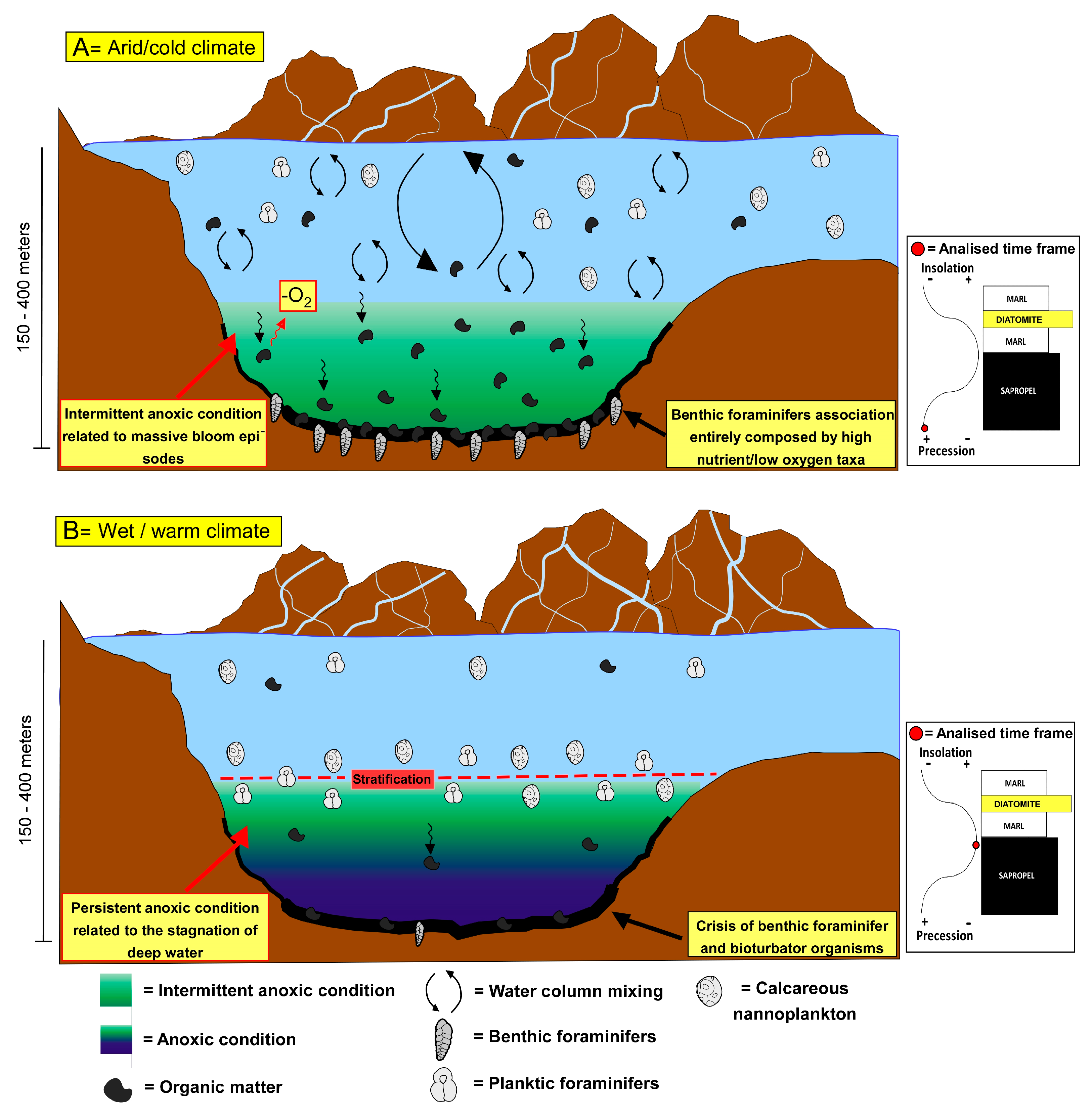
5.2. Sorbas Shallow-Sea vs. Holocene Deep-Sea Sapropels: What Are the Differences?
5.3. A Clue on Future Deoxygenation: Are Sapropels a Good Analogue?
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar] [CrossRef]
- Limburg, K.E.; Breitburg, D.; Swaney, D.P.; Jacinto, G. Ocean deoxygenation: A primer. One Earth 2020, 2, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Mavropoulou, A.-M.; Vervatis, V.; Sofianos, S. Dissolved oxygen variability in the Mediterranean Sea. J. Mar. Syst. 2020, 208, 103348. [Google Scholar] [CrossRef]
- Danovaro, R. Climate change impacts on the biota and on vulnerable habitats of the deep Mediterranean Sea. Rendiconti Lince-Sci. Fis. Nat. 2018, 29, 525–541. [Google Scholar] [CrossRef]
- Robinson, C. Microbial respiration, the engine of ocean deoxygenation. Front. Mar. Sci. 2019, 5, 533. [Google Scholar] [CrossRef] [Green Version]
- Schmidtko, S.; Stramma, L.; Visbeck, M. Decline in global oceanic oxygen content during the past five decades. Nature 2017, 542, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef]
- Stramma, L.; Schmidtko, S.; Levin, L.A.; Johnson, G.C. Ocean oxygen minima expansions and their biological impacts. Deep Sea Res. Part I Oceanogr. Res. Pap. 2010, 57, 587–595. [Google Scholar] [CrossRef]
- Zhou, X.; Thomas, E.; Rickaby, R.E.M.; Winguth, A.M.E.; Lu, Z. I/Ca evidence for upper ocean deoxygenation during the PETM. Paleoceanography 2014, 29, 964–975. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.; Paytan, A.; Wortmann, U.G. Large-scale ocean deoxygenation during the Paleocene-Eocene Thermal Maximum. Science 2019, 361, 804–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, R.J.; Rosenberg, R. Marine benthic hypoxia: A review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 03. [Google Scholar]
- Roman, M.R.; Brandt, S.B.; Houde, E.D.; Pierson, J.J. Interactive effects of hypoxia and temperature on coastal pelagic zooplankton and fish. Front. Mar. Sci. 2019, 6, 139. [Google Scholar] [CrossRef] [Green Version]
- Pauly, D.; Dimarchopoulou, D. Introduction: Fishes in a warming and deoxygenating world. Environ. Biol. Fishes 2022, 105, 1261–1267. [Google Scholar] [CrossRef]
- Doney, S.C.; Ruckelshaus, M.; Duffy, J.E.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate change impacts on marine ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidd, R.B.; Cita, M.B.; Ryan, W.B.F. Stratigraphy of eastern Mediterranean sapropel sequences recovered during DSDP Leg 42A and their paleoenvironmental significance. Affil. Lamont-Doherty Geol. Obs. 1978, 42, 421. [Google Scholar]
- Emeis, K.C.; Party, S.S. Paleoceanography and sapropel introduction. Proc. Ocean. Drill. Program Initial. Rep. 1996, 160, 21–28. [Google Scholar]
- Rohling, E.; Marino, G.; Grant, K. Mediterranean climate and oceanography, and the periodic development of anoxic events (sapropels). Earth-Sci. Rev. 2015, 143, 62–97. [Google Scholar] [CrossRef]
- Rohling, E.J.; Hilgen, F.J. The eastern Mediterranean climate at times of sapropel formation: A review. Neth. J. Geosci./Geol. En Mijnb. 2007, 70, 253–264. [Google Scholar]
- Schenau, S.J.; Antonarakou, A.; Hilgen, F.J.; Lourens, L.J.; Nijenhuis, I.A.; van der Weijden, C.H.; Zachariasse, W.J. Organic-rich layers in the Metochia section (Gavdos, Greece): Evidence for a single mechanism of sapropel formation during the past 10 My. Mar. Geol. 1999, 153, 117–135. [Google Scholar] [CrossRef]
- Mancini, A.M.; Gennari, R.; Ziveri, P.; Mortyn, P.G.; Stolwijk, D.J.; Lozar, F. Calcareous nannofossil and foraminiferal trace element records in the Sorbas Basin: A new piece of the Messinian Salinity Crisis onset puzzle. Palaeogeogr. Palaeoclim. Palaeoecol. 2020, 554, 109796. [Google Scholar] [CrossRef]
- Justić, D.; Legović, T.; Rottini-Sandrini, L. Trends in oxygen content 1911–1984 and occurrence of benthic mortality in the northern Adriatic Sea. Estuar. Coast. Shelf Sci. 1987, 25, 435–445. [Google Scholar] [CrossRef]
- UNEP-MAP. State of the Mediterranean Marine and Coastal Environment. Highlights for Policy Makers. 2012. Available online: www.unepmap.org/index.php (accessed on 2 December 2022).
- Turley, C.M. The changing Mediterranean Sea—A sensitive ecosystem? Prog. Oceanogr. 1999, 44, 387–400. [Google Scholar] [CrossRef]
- Viaroli, P.; Nizzoli, D.; Pinardi, M.; Soana, E.; Bartoli, M. Eutrophication of the Mediterranean Sea: A watershed—Cascading aquatic filter approach. Rend. Lince Sci. Fis. Nat. 2014, 26, 13–23. [Google Scholar] [CrossRef]
- MedECC. Climate and Environmental Change in the Mediterranean Basin–Current Situation and Risks for the Future; First Mediter-ranean Assessment Report; Cramer, W., Guiot, J., Marini, K., Eds.; Union for the Mediterranean, Plan Bleu, UNEP/MAP: Marseille, France, 2020; 632p. [Google Scholar] [CrossRef]
- Schlitzer, R. Ocean Data View, Version 4.4. 2. 2014-03-31. 2011. Available online: https://odv.awi.de (accessed on 5 October 2021).
- Somot, S.; Sevault, F.; Déqué, M. Transient climate change scenario simulation of the Mediterranean Sea for the twenty-first century using a high-resolution ocean circulation model. Clim. Dyn. 2006, 27, 851–879. [Google Scholar] [CrossRef] [Green Version]
- Powley, H.R.; Krom, M.D.; Van Cappellen, P. Circulation and oxygen cycling in the Mediterranean Sea: Sensitivity to future climate change. J. Geophys. Res. Ocean. 2016, 121, 8230–8247. [Google Scholar] [CrossRef] [Green Version]
- Reale, M.; Cossarini, G.; Lazzari, P.; Lovato, T.; Bolzon, G.; Masina, S.; Solidoro, C.; Salon, S. Acidification, deoxygenation, and nutrient and biomass declines in a warming Mediterranean Sea. Biogeosciences 2022, 19, 4035–4065. [Google Scholar] [CrossRef]
- Berger, A.L. Long-Term variations of caloric insolation resulting from the Earth’s orbital elements. Quat. Res. 1978, 9, 139–167. [Google Scholar] [CrossRef]
- Taylforth, J.E.; McCay, G.A.; Ellam, R.; Raffi, I.; Kroon, D.; Robertson, A.H. Middle Miocene (Langhian) sapropel formation in the easternmost Mediterranean deep-water basin: Evidence from northern Cyprus. Mar. Pet. Geol. 2014, 57, 521–536. [Google Scholar] [CrossRef]
- Olausson, E. Studies of deep-sea cores. Rep. Swed. Deep Sea Exped. 1961, 1947–1948, 353–391. [Google Scholar]
- Tesi, T.; Asioli, A.; Minisini, D.; Maselli, V.; Valle, G.D.; Gamberi, F.; Langone, L.; Cattaneo, A.; Montagna, P.; Trincardi, F. Large-scale response of the Eastern Mediterranean thermohaline circulation to African monsoon intensification during sapropel S1 formation. Quat. Sci. Rev. 2017, 159, 139–154. [Google Scholar] [CrossRef] [Green Version]
- Pinardi, N.; Masetti, E. Variability of the large scale general circulation of the Mediterranean Sea from observations and modelling: A review. Palaeogeogr. Palaeoclim. Palaeoecol. 2000, 158, 153–173. [Google Scholar] [CrossRef]
- Krijgsman, W.; Hilgen, F.J.; Raffi, I.; Sierro, F.J.; Wilson, D.S. Chronology, causes and progression of the Messinian salinity crisis. Nature 1999, 400, 652–655. [Google Scholar] [CrossRef] [Green Version]
- Hilgen, F.J.; Krijgsman, W.; Langereis, C.G.; Lourens, L.J.; Santarelli, A.; Zachariasse, W.J. Extending the astronomical (polarity) time scale into the Miocene. Earth Planet. Sci. Lett. 1995, 136, 495–510. [Google Scholar] [CrossRef] [Green Version]
- Hilgen, F.J.; Krijgsman, W. Cyclostratigraphy and astrochronology of the Tripolo diatomite formation pre-evaporite Messinian, Sicily, Italy. Terra Nova 1999, 11, 16–22. [Google Scholar] [CrossRef]
- Flecker, R.; Krijgsman, W.; Capella, W.; Martíns, C.D.C.; Dmitrieva, E.; Mayser, J.P.; Marzocchi, A.; Modestou, S.; Ochoa, D.; Simon, D.; et al. Evolution of the Late Miocene Mediterranean–Atlantic gateways and their impact on regional and global environmental change. Earth Sci. Rev. 2015, 150, 365–392. [Google Scholar] [CrossRef] [Green Version]
- Kouwenhoven, T.; Morigi, C.; Negri, A.; Giunta, S.; Krijgsman, W.; Rouchy, J.-M. Paleoenvironmental evolution of the eastern Mediterranean during the Messinian: Constraints from integrated microfossil data of the Pissouri Basin (Cyprus). Mar. Micropaleontol. 2006, 60, 17–44. [Google Scholar] [CrossRef]
- Corbí, H.; Soria, J.M.; Giannetti, A.; Yébenes, A. The step-by-step restriction of the Mediterranean (start, amplification, and consolidation phases) preceding the Messinian Salinity Crisis (climax phase) in the Bajo Segura basin. Geo-Mar. Lett. 2020, 40, 341–361. [Google Scholar] [CrossRef]
- Bulian, F.; Kouwenhoven, T.J.; Jiménez-Espejo, F.J.; Krijgsman, W.; Andersen, N.; Sierro, F.J. Impact of the Mediterranean-Atlantic connectivity and the late Miocene carbon shift on deep-sea communities in the Western Alboran Basin. Palaeogeogr. Palaeoclim. Palaeoecol. 2022, 589, 110841. [Google Scholar] [CrossRef]
- Hilgen, F.J.; Aziz, H.A.; Krijgsman, W.; Raffi, I.; Turco, E. Integrated stratigraphy and astronomical tuning of the Serravallian and lower Tortonian at Monte dei Corvi (Middle–Upper Miocene, northern Italy). Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 199, 229–264. [Google Scholar] [CrossRef]
- Kouwenhoven, T.J.; Seidenkrantz, M.-S.; van der Zwaan, G.J. Deep-water changes: The near-synchronous disappearance of a group of benthic foraminifera from the Late Miocene Mediterranean. Palaeogeogr. Palaeoclim. Palaeoecol. 1999, 152, 259–281. [Google Scholar] [CrossRef]
- Gladstone, R.; Flecker, R.; Valdes, P.; Lunt, D.; Markwick, P. The Mediterranean hydrologic budget from a Late Miocene global climate simulation. Palaeogeogr. Palaeoclim. Palaeoecol. 2007, 251, 254–267. [Google Scholar] [CrossRef]
- Karakitsios, V.; Roveri, M.; Lugli, S.; Manzi, V.; Gennari, R.; Antonarakou, A.; Triantaphyllou, M.; Agiadi, K.; Kontakiotis, G.; Kafousia, N.; et al. A record of the Messinian salinity crisis in the eastern Ionian tectonically active domain (Greece, eastern Mediterranean). Basin Res. 2017, 29, 203–233. [Google Scholar] [CrossRef]
- Sierro, F.J.; Flores, J.A.; Zamarreño, I.; Vázquez, A.; Utrilla, R.; Francés, G.; Hilgen, F.J.; Krijgsman, W. Messinian pre-evaporite sapropels and precession-induced oscillations in western Mediterranean climate. Mar. Geol. 1999, 153, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Iaccarino, S.M.; Bertini, A.; Di Stefano, A.; Ferraro, L.; Gennari, R.; Grossi, F.; Lirer, F.; Manzi, V.; Menichetti, E.; Lucchi, M.R.; et al. The Trave section (Monte dei Corvi, Ancona, Central Italy): An integrated paleontological study of the Messinian deposits. Stratigraphy 2008, 5, 281–306. [Google Scholar]
- Di Stefano, A.; Verducci, M.; Lirer, F.; Ferraro, L.; Iaccarino, S.M.; Hüsing, S.K.; Hilgen, F.J. Paleoenvironmental conditions preceding the Messinian Salinity Crisis in the Central Mediterranean: Integrated data from the Upper Miocene Trave section (Italy). Palaeogeogr. Palaeoclim. Palaeoecol. 2010, 297, 37–53. [Google Scholar] [CrossRef]
- Sierro, F.J.; Flores, J.A.; Frances, G.; Vazquez, A.; Utrilla, R.; Zamarreño, I.; Erlenkeuser, H.; Barcena, M.A. Orbitally-controlled oscillations in planktic communities and cyclic changes in western Mediterranean hydrography during the Messinian. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 190, 289–316. [Google Scholar] [CrossRef]
- AlHammoud, B.; Meijer, P.T.; Dijkstra, H.A. Sensitivity of Mediterranean thermohaline circulation to gateway depth: A model investigation. Paleoceanography 2010, 25, PA222. [Google Scholar] [CrossRef]
- Gennari, R.; Lozar, F.; Turco, E.; Pierre, F.D.; Lugli, S.; Manzi, V.; Natalicchio, M.; Roveri, M.; Schreiber, B.C.; Taviani, M. Integrated stratigraphy and paleoceanographic evolution of the pre-evaporitic phase of the Messinian salinity crisis in the Eastern Mediterranean as recorded in the Tokhni section (Cyprus island). Newsl. Strat. 2018, 51, 33–55. [Google Scholar] [CrossRef] [Green Version]
- Popov, S.V.; Rogl, F.; Rozanov, A.Y.; Steininger, F.F.; Shcherba, I.G.; Kovac, M. Lithological-paleogeographic maps of paratethys. CFS Cour. Forsch. Senckenberg 2004, 254, 1–46. [Google Scholar]
- Manzi, V.; Roveri, M.; Gennari, R.; Bertini, A.; Biffi, U.; Giunta, S.; Iaccarino, S.M.; Lanci, L.; Lugli, S.; Negri, A.; et al. The deep-water counterpart of the Messinian lower Evaporites in the Apennine foredeep: The Fanantello section (Northern Apennines, Italy). Palaeogeogr. Palaeoclim. Palaeoecol. 2007, 251, 470–499. [Google Scholar] [CrossRef]
- Lozar, F.; Violanti, D.; Pierre, F.D.; Bernardi, E.; Cavagna, S.; Clari, P.; Irace, A.; Martinetto, E.; Trenkwalder, S. Calcareous nannofossils and foraminifers herald the Messinian Salinity Crisis: The Pollenzo section (Alba, Cuneo; NW Italy). Geobios 2010, 43, 21–32. [Google Scholar] [CrossRef]
- Corbí, H.; Soria, J.M.; Lancis, C.; Giannetti, A.; Tent-Manclús, J.E.; Dinarès-Turell, J. Sedimentological and paleoenvironmental scenario before, during, and after the Messinian Salinity Crisis: The San Miguel de Salinas composite section (western Mediterranean). Mar. Geol. 2016, 379, 246–266. [Google Scholar] [CrossRef] [Green Version]
- Krijgsman, W.; Capella, W.; Simon, D.; Hilgen, F.J.; Kouwenhoven, T.J.; Meijer, P.T.; Sierro, F.J.; Tulbure, M.A.; van den Berg, B.C.; van der Schee, M.; et al. The Gibraltar Corridor: Watergate of the Messinian Salinity Crisis. Mar. Geol. 2018, 403, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Capella, W.; Flecker, R.; Hernández-Molina, F.J.; Simon, D.; Meijer, P.T.; Rogerson, M.; Sierro, F.J.; Krijgsman, W. Mediterranean isolation preconditioning the Earth System for late Miocene climate cooling. Sci. Rep. 2019, 9, 3795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskar, J.; Robutel, P.; Joutel, F.; Gastineau, M.; Correia, A.C.M.; Levrard, B. A long-term numerical solution for the insolation quantities of the Earth. Astron. Astrophys. 2004, 428, 261–285. [Google Scholar] [CrossRef] [Green Version]
- Sierro, F.J.; Hilgen, F.J.; Krijgsman, W.; Flores, J.A. The Abad composite (SE Spain): A Messinian reference section for the Mediterranean and the APTS. Palaeogeogr. Palaeoclim. Palaeoecol. 2001, 168, 141–169. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Folgado, M.; Sierro, F.J.; Bárcena, M.A.; Flores, J.; Vázquez, A.; Utrilla, R.; Hilgen, F.; Krijgsman, W.; Filippelli, G. Western versus eastern Mediterranean paleoceanographic response to astronomical forcing: A high-resolution microplankton study of precession-controlled sedimentary cycles during the Messinian. Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 190, 317–334. [Google Scholar] [CrossRef]
- Flores, J.-A.; Sierro, F.J.; Filippelli, G.M.; Bárcena, M.; Pérez-Folgado, M.; Vázquez, A.; Utrilla, R. Surface water dynamics and phytoplankton communities during deposition of cyclic late Messinian sapropel sequences in the western Mediterranean. Mar. Micropaleontol. 2005, 56, 50–79. [Google Scholar] [CrossRef]
- Reghizzi, M.; Gennari, R.; Douville, E.; Lugli, S.; Manzi, V.; Montagna, P.; Roveri, M.; Sierro, F.J.; Taviani, M. Isotope stratigraphy (87Sr/86Sr, δ18O, δ13C) of the Sorbas basin (Betic Cordillera, Spain): Paleoceanographic evolution across the onset of the Messinian salinity crisis. Palaeogeogr. Palaeoclim. Palaeoecol. 2017, 469, 60–73. [Google Scholar] [CrossRef]
- Hemleben, C.; Spindler, M.; Anderson, O.R. Modern Planktonic Foraminifera; Springer: New York, NY, USA, 1989; pp. 1–363. [Google Scholar] [CrossRef]
- Pujol, C.; Grazzini, C.V. Distribution patterns of live planktic foraminifers as related to regional hydrography and productive systems of the Mediterranean Sea. Mar. Micropaleontol. 1995, 25, 187–217. [Google Scholar] [CrossRef]
- Jannink, N.T.; Zachariasse, W.J.; Van Der Zwaan, G.J. Living (Rose Bengal stained) benthic foraminifera from the Pakistan continental margin (northern Arabian Sea). Deep Sea Res. Part I Oceanogr. Res. Pap. 1998, 45, 1483–1513. [Google Scholar] [CrossRef]
- Jorissen, F.J. Benthic foraminiferal microhabitats below the sediment-water interface. In Modern Foraminifera; Springer: Dordrecht, The Netherlands, 1999; pp. 161–179. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006; pp. 1–426. [Google Scholar]
- Morigi, C.; Jorissen, F.J.; Gervais, A.; Guichard, S.; Borsetti, A.M. Benthic foraminiferal faunas in surface sediments off NW Africa: Relationship with organic flux to the ocean floor. J. Foraminifer. Res. 2001, 31, 350–368. [Google Scholar] [CrossRef]
- Morigi, C. Benthic environmental changes in the Eastern Mediterranean Sea during sapropel S5 deposition. Palaeogeogr. Palaeoclim. Palaeoecol. 2009, 273, 258–271. [Google Scholar] [CrossRef]
- Gibbs, S.; Shackleton, N.; Young, J. Orbitally forced climate signals in mid-Pliocene nannofossil assemblages. Mar. Micropaleontol. 2004, 51, 39–56. [Google Scholar] [CrossRef]
- Negri, A.; Capotondi, L.; Keller, J. Calcareous nannofossils, planktonic foraminifera and oxygen isotopes in the late Quaternary sapropels of the Ionian Sea. Mar. Geol. 1999, 157, 89–103. [Google Scholar] [CrossRef]
- Fallon, M.F.; Papadopoulos, G.; Leonard, J.J.; Patrikalakis, N.M. Cooperative AUV navigation using a single maneuvering surface craft. Int. J. Robot. Res. 2010, 29, 1461–1474. [Google Scholar] [CrossRef] [Green Version]
- De Lange, G.J.; Thomson, J.; Reitz, A.; Slomp, C.P.; Principato, M.S.; Erba, E.; Corselli, C. Synchronous basin-wide formation and redox-controlled preservation of a Mediterranean sapropel. Nat. Geosci. 2008, 1, 606–610. [Google Scholar] [CrossRef]
- Keller, J.; Ryan, W.B.F.; Ninkovich, D.; Altherr, R. Explosive volcanic activity in the Mediterranean over the past 200,000 yr as recorded in deep-sea sediments. GSA Bull. 1978, 89, 591–604. [Google Scholar] [CrossRef]
- Rohling, E.J.; Sprovieri, M.; Cane, T.R.; Casford, J.S.L.; Cooke, S.; Bouloubassi, I.; Emeis, K.C.; Schiebel, R.; Rogerson, M.; Hayes, A.; et al. Reconstructing past planktic foraminiferal habitats using stable isotope data: A case history for Mediterranean sapropel S5. Mar. Micropaleontol. 2004, 50, 89–123. [Google Scholar] [CrossRef]
- Castradori, D. Calcareous nannofossils and the origin of eastern Mediterranean sapropels. Paleoceanography 1993, 8, 459–471. [Google Scholar] [CrossRef]
- Triantaphyllou, M.; Ziveri, P.; Gogou, A.; Marino, G.; Lykousis, V.; Bouloubassi, I.; Emeis, K.-C.; Kouli, K.; Dimiza, M.; Rosell-Melé, A.; et al. Late Glacial–Holocene climate variability at the south-eastern margin of the Aegean Sea. Mar. Geol. 2009, 266, 182–197. [Google Scholar] [CrossRef]
- Kemp, A.E.; Pike, J.; Pearce, R.B.; Lange, C.B. The “Fall dump”—A new perspective on the role of a “shade flora” in the annual cycle of diatom production and export flux. Deep Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 2129–2154. [Google Scholar] [CrossRef]
- Brodie, I.; Kemp, A.E.S. Variation in biogenic and detrital fluxes and formation of laminae in late Quaternary sediments from the Peruvian coastal upwelling zone. Mar. Geol. 1994, 116, 385–398. [Google Scholar] [CrossRef]
- Gooday, A.J. Benthic foraminifera (Protista) as tools in deep-water palaeoceanography: Environmental influences on faunal characteristics. Adv. Mar. Biol. 2003, 46, 1–90. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Sen Gupta, B.K. Foraminifera of oxygen-depleted environments. In Modern Foraminifera; Springer: Dordrecht, The Netherlands, 2003; pp. 201–216. [Google Scholar]
- Glock, N.; Roy, A.-S.; Romero, D.; Wein, T.; Weissenbach, J.; Revsbech, N.P.; Høgslund, S.; Clemens, D.; Sommer, S.; Dagan, T. Metabolic preference of nitrate over oxygen as an electron acceptor in foraminifera from the Peruvian oxygen minimum zone. Proc. Natl. Acad. Sci. USA 2019, 116, 2860–2865. [Google Scholar] [CrossRef] [Green Version]
- Cornuault, M.; Tachikawa, K.; Vidal, L.; Guihou, A.; Siani, G.; Deschamps, P.; Bassinot, F.; Revel, M. Circulation Changes in the Eastern Mediterranean Sea Over the Past 23,000 Years Inferred From Authigenic Nd Isotopic Ratios. Paleoceanography 2018, 33, 264–280. [Google Scholar] [CrossRef]
- Schmiedl, G.; Kuhnt, T.; Ehrmann, W.; Emeis, K.-C.; Hamann, Y.; Kotthoff, U.; Dulski, P.; Pross, J. Climatic forcing of eastern Mediterranean deep-water formation and benthic ecosystems during the past 22 000 years. Quat. Sci. Rev. 2010, 29, 3006–3020. [Google Scholar] [CrossRef]
- Rossignol-Strick, M. African monsoons, an immediate climate response to orbital insolation. Nature 1983, 304, 46–49. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Rohling, E.J.; Abu-Zied, R.; Cooke, S.; Fontanier, C.; Leng, M.; Lykousis, V. Circulation changes and nutrient concentrations in the late Quaternary Aegean Sea: A nonsteady state concept for sapropel formation. Paleoceanography 2002, 17, 14-1–14-11. [Google Scholar] [CrossRef] [Green Version]
- Abu-Zied, R.H.; Rohling, E.J.; Jorissen, F.J.; Fontanier, C.; Casford, J.S.L.; Cooke, S. Benthic foraminiferal response to changes in bottom-water oxygenation and organic carbon flux in the eastern Mediterranean during LGM to Recent times. Mar. Micropaleontol. 2008, 67, 46–68. [Google Scholar] [CrossRef]
- Marino, G.; Rohling, E.J.; Sangiorgi, F.; Hayes, A.; Casford, J.S.L.; Lotter, A.F.; Kucera, M.; Brinkhuis, H. Early and middle Holocene in the Aegean Sea: Interplay between high and low latitude climate variability. Quat. Sci. Rev. 2009, 28, 3246–3262. [Google Scholar] [CrossRef]
- Hennekam, R.; Jilbert, T.; Schnetger, B.; de Lange, G.J. Solar forcing of Nile discharge and sapropel S1 formation in the early to middle Holocene eastern Mediterranean. Paleoceanography 2014, 29, 343–356. [Google Scholar] [CrossRef]
- Blanchet, C.L.; Tjallingii, R.; Schleicher, A.M.; Schouten, S.; Frank, M.; Brauer, A. Deoxygenation dynamics on the western Nile deep-sea fan during sapropel S1 from seasonal to millennial timescales. Clim. Past 2021, 17, 1025–1050. [Google Scholar] [CrossRef]
- Castañeda, I.S.; Schouten, S.; Pätzold, J.; Lucassen, F.; Kasemann, S.; Kuhlmann, H.; Schefuß, E. Hydroclimate variability in the Nile River Basin during the past 28,000 years. Earth Planet. Sci. Lett. 2016, 438, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Weldeab, S.; Menke, V.; Schmiedl, G. The pace of East African monsoon evolution during the Holocene. Geophys. Res. Lett. 2014, 41, 1724–1732. [Google Scholar] [CrossRef]
- Mojtahid, M.; Manceau, R.; Schiebel, R.; Hennekam, R.; de Lange, G.J. Thirteen thousand years of southeastern Mediterranean climate variability inferred from an integrative planktic foraminiferal-based approach. Paleoceanography 2015, 30, 402–422. [Google Scholar] [CrossRef]
- Mercone, D.; Thomson, J.; Croudace, I.W.; Siani, G.; Paterne, M.; Troelstra, S. Duration of S1, the most recent sapropel in the eastern Mediterranean Sea, as indicated by accelerator mass spectrometry radiocarbon and geochemical evidence. Paleoceanography 2000, 15, 336–347. [Google Scholar] [CrossRef]
- Rohling, E.J.; Cane, T.R.; Cooke, S.; Sprovieri, M.; Bouloubassi, I.; Emeis, K.C.; Schiebel, R.; Kroon, D.; Jorissen, F.J.; Lorre, A.; et al. African monsoon variability during the previous interglacial maximum. Earth Planet. Sci. Lett. 2002, 202, 61–75. [Google Scholar] [CrossRef]
- Giunta, S.; Negri, A.; Morigi, C.; Capotondi, L.; Combourieu-Nebout, N.; Emeis, K.C.; Sangiorgi, F.; Vigliotti, L. Coccolithophorid ecostratigraphy and multi-proxy paleoceanographic reconstruction in the Southern Adriatic Sea during the last deglacial time (Core AD91-17). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 190, 39–59. [Google Scholar] [CrossRef]
- Casford, J.S.L.; Abu-Zied, R.; Rohling, E.J.; Cooke, S.; Fontanier, C.; Leng, M.; Millard, A.; Thomson, J. A strat-igraphically controlled multi-proxy chronostratigraphy for the eastern Mediterranean. Paleoceanography 2007, 22, PA4215. [Google Scholar] [CrossRef]
- Mojtahid, M.; Hennekam, R.; De Nooijer, L.; Reichart, G.-J.; Jorissen, F.; Boer, W.; Le Houedec, S.; De Lange, G. Evaluation and application of foraminiferal element/calcium ratios: Assessing riverine fluxes and environmental conditions during sapropel S1 in the southeastern Mediterranean. Mar. Micropaleontol. 2019, 153, 101783. [Google Scholar] [CrossRef]
- Cascella, A.; Bonomo, S.; Lirer, F.; Margaritelli, G.; Checa, H.; Cacho, I.; Pena, L.D.; Frigola, J. The response of calcareous plankton to the Sapropel S1 interval in North Ionian Sea. Glob. Planet. Chang. 2021, 205, 103599. [Google Scholar] [CrossRef]
- Schimmelmann, A.; Lange, C.B. Tales of 1001 varves: A review of Santa Barbara Basin sediment studies. In Palaeoclimatology and Palaeoceanography from Laminated Sediments; Kemp, A.E.S., Ed.; Geological Society of London Special Publication: London, UK, 1996; Volume 116, pp. 121–141. [Google Scholar] [CrossRef]
- Lückge, A.; Horsfield, B.; Littke, R.; Scheeder, G. Organic matter preservation and sulfur uptake in sediments from the continental margin off Pakistan. Org. Geochem. 2002, 33, 477–488. [Google Scholar] [CrossRef]
- Dronkert, H. Late Miocene evaporites in the Sorbas Basin and adjoining areas. Mem. Soc. Geol. Ital. 1976, 16, 341–361. [Google Scholar]
- Troelstra, S.R.; Van Der Poel, H.M.; Huisman, C.H.A.; Geerlings, L.P.; Dronkert, H. Paleoecological changes in the latest Miocene of Sorbas basin, S.E. Spain. Géologie Méditerranéenne 1980, 7, 115–125. [Google Scholar] [CrossRef]
- van de Poel, H.M. Foraminiferal biostratigraphy and palaeoenvironments of the Miocene-Pliocene Carboneras-Nijar basin (SE Spain). Scr. Geol. 1992, 102, 1–32. [Google Scholar]
- Baggley, K.A. The late tortonian-early messinian foraminiferal record of the Abad Member (Turre formation), Sorbas Basin, Almeria, south-east Spain. Palaeontology 2000, 43, 1069–1112. [Google Scholar] [CrossRef]
- Clauzon, G.; Suc, J.-P.; Couto, D.D.; Jouannic, G.; Melinte-Dobrinescu, M.C.; Jolivet, L.; Quillévéré, F.; Lebret, N.; Mocochain, L.; Popescu, S.-M.; et al. New insights on the Sorbas Basin (SE Spain): The onshore reference of the Messinian salinity crisis. Mar. Pet. Geol. 2015, 66, 71–100. [Google Scholar] [CrossRef] [Green Version]
- Modestou, S.; Simon, D.; Gutjahr, M.; Marzocchi, A.; Kouwenhoven, T.J.; Ellam, R.M.; Flecker, R. Precessional variability of87Sr/86Sr in the late Miocene Sorbas Basin: An interdisciplinary study of drivers of interbasin exchange. Paleoceanography 2017, 32, 531–552. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, L08707. [Google Scholar] [CrossRef]
- Bethoux, J.P.; Gentili, B.; Raunet, J.; Tailliez, D. Warming trend in the western Mediterranean deep water. Nature 1990, 347, 660–662. [Google Scholar] [CrossRef]
- Nykjaer, L. Mediterranean Sea surface warming 1985–2006. Clim. Res. 2009, 39, 11–17. [Google Scholar] [CrossRef]
- Sakalli, A. Sea surface temperature change in the Mediterranean Sea under climate change: A linear model for simulation of the sea surface temperature up to 2100. Appl. Ecol. Environ. Res. 2017, 15, 707–716. [Google Scholar] [CrossRef]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Skliris, N. Past, Present and future patterns of the thermohaline circulation and characteristic water masses of the Mediterranean Sea. In The Mediterranean Sea; Springer: Dordrecht, The Netherlands, 2014; pp. 29–48. [Google Scholar] [CrossRef]
- Coppola, L.; Legendre, L.; Lefevre, D.; Prieur, L.; Taillandier, V.; Riquier, E.D. Seasonal and inter-annual variations of dissolved oxygen in the northwestern Mediterranean Sea (DYFAMED site). Prog. Oceanogr. 2018, 162, 187–201. [Google Scholar] [CrossRef]
- Keeling, R.F.; Körtzinger, A.; Gruber, N. Ocean deoxygenation in a warming world. Annu. Rev. Mar. Sci. 2010, 2, 199–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzanova, A.; Herbert, T.D.; Peterson, L. Cooling Mediterranean Sea surface temperatures during the Late Miocene provide a climate context for evolutionary transitions in Africa and Eurasia. Earth Planet. Sci. Lett. 2015, 419, 71–80. [Google Scholar] [CrossRef]
- Vasiliev, I.; Karakitsios, V.; Bouloubassi, I.; Agiadi, K.; Kontakiotis, G.; Antonarakou, A.; Triantaphyllou, M.; Gogou, A.; Kafousia, N.; De Rafélis, M.; et al. Large Sea Surface Temperature, Salinity, and Productivity-Preservation Changes Preceding the Onset of the Messinian Salinity Crisis in the Eastern Mediterranean Sea. Paleoceanogr. Paleoclimatol. 2019, 34, 182–202. [Google Scholar] [CrossRef]
- Kontakiotis, G.; Butiseacă, G.A.; Antonarakou, A.; Agiadi, K.; Zarkogiannis, S.D.; Krsnik, E.; Besiou, E.; Zachariasse, W.J.; Lourens, L.; Thivaiou, D.; et al. Hypersalinity accompanies tectonic restriction in the eastern Mediterranean prior to the Messinian Salinity Crisis. Palaeogeogr. Palaeoclim. Palaeoecol. 2022, 592, 110903. [Google Scholar] [CrossRef]
- Rohling, E.J.; Bryden, H.L. Man-induced salinity and temperature increases in Western Mediterranean Deep Water. J. Geophys. Res. Atmos. 1992, 97, 11191–11198. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Gomez, E.; Somot, S.; Mariotti, A. Future changes in the Mediterranean water budget projected by an ensemble of regional climate models. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef]
- Wu, C.-H.; Lee, S.-Y.; Tsai, P.-C. Role of eccentricity in early Holocene African and Asian summer monsoons. Sci. Rep. 2021, 11, 24089. [Google Scholar] [CrossRef] [PubMed]
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, eaam7240. [Google Scholar] [CrossRef] [Green Version]
- Mancini, L.; Milandri, A.; Nizzoli, C.; Pirini, M.; Poletti, R.; Pompei, M.; Viviani, R. L’eutrofizzazione in rapporto ad alcuni aspetti igienico-sanitari delle acque costiere e dei prodotti della pesca. Nova Thalass. 1986, 8, 325–330. [Google Scholar]
- Artegiani, A.; Solazzi, A.; Tolomio, C.; Marzocchi, M.; Favero, P.; Cavolo, F. Parametri fisico-chimici e fitoplancton in una stazione a Sud del Conero (Zona di ripopolamento). Convegno Sci. Naz. PF Oceanogr. CNR 1979, 5, 1979. [Google Scholar]
- Colleoni, F.; Masina, S.; Negri, A.; Marzocchi, A. Plio–Pleistocene high–low latitude climate interplay: A Mediterranean point of view. Earth Planet. Sci. Lett. 2012, 319–320, 35–44. [Google Scholar] [CrossRef]
- Batziakas, S.; Frangoulis, C.; Tsiola, A.; Nikolioudakis, N.; Tsagaraki, T.M.; Somarakis, S. Hypoxia changes the shape of the biomass size spectrum of planktonic communities: A case study in the eastern Mediterranean (Elefsina Bay). J. Plankton Res. 2020, 42, 752–766. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancini, A.M.; Bocci, G.; Morigi, C.; Gennari, R.; Lozar, F.; Negri, A. Past Analogues of Deoxygenation Events in the Mediterranean Sea: A Tool to Constrain Future Impacts. J. Mar. Sci. Eng. 2023, 11, 562. https://doi.org/10.3390/jmse11030562
Mancini AM, Bocci G, Morigi C, Gennari R, Lozar F, Negri A. Past Analogues of Deoxygenation Events in the Mediterranean Sea: A Tool to Constrain Future Impacts. Journal of Marine Science and Engineering. 2023; 11(3):562. https://doi.org/10.3390/jmse11030562
Chicago/Turabian StyleMancini, Alan Maria, Giacomo Bocci, Caterina Morigi, Rocco Gennari, Francesca Lozar, and Alessandra Negri. 2023. "Past Analogues of Deoxygenation Events in the Mediterranean Sea: A Tool to Constrain Future Impacts" Journal of Marine Science and Engineering 11, no. 3: 562. https://doi.org/10.3390/jmse11030562