
First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours

Abstract
:1. Introduction
2. Materials and Methods
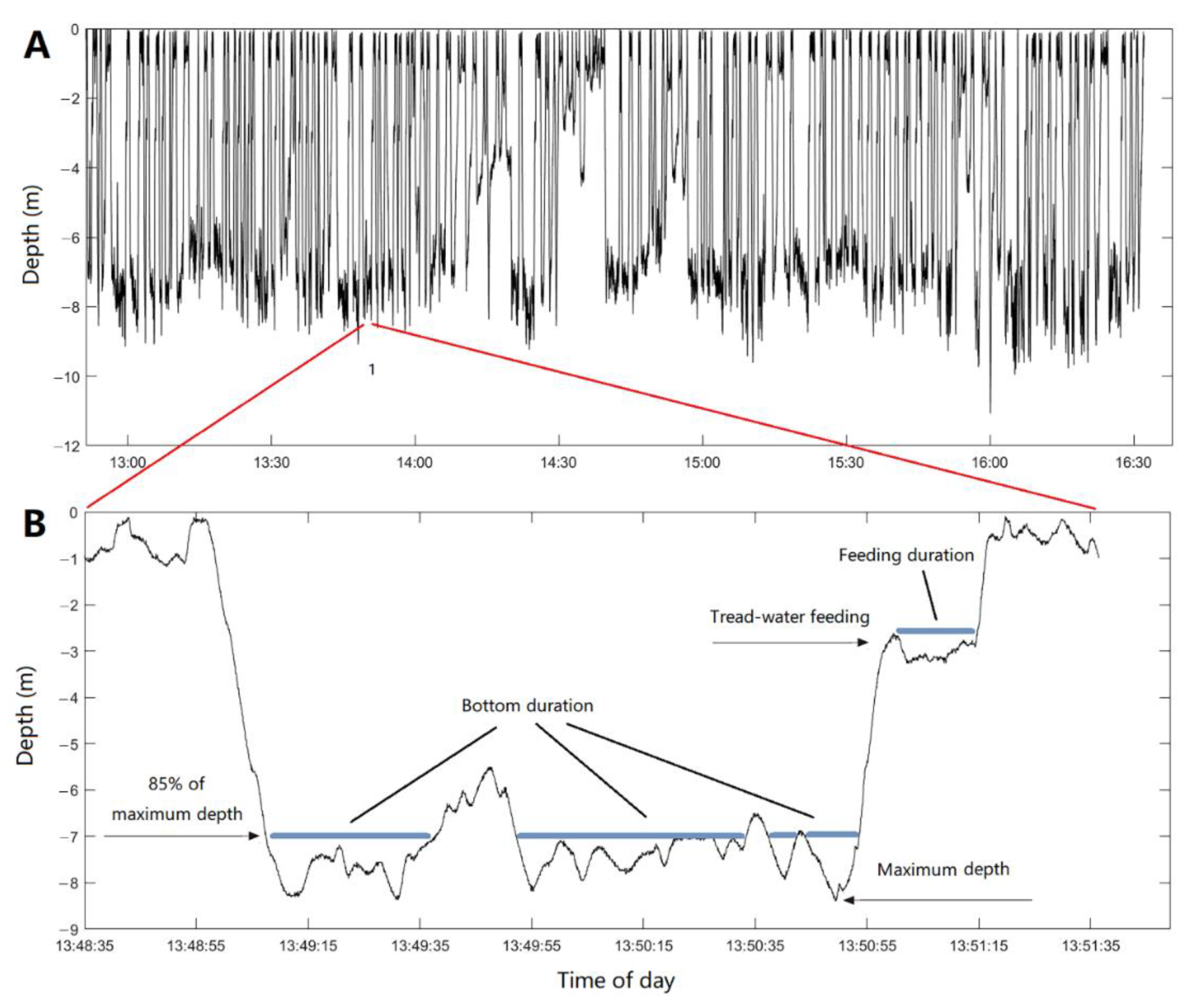
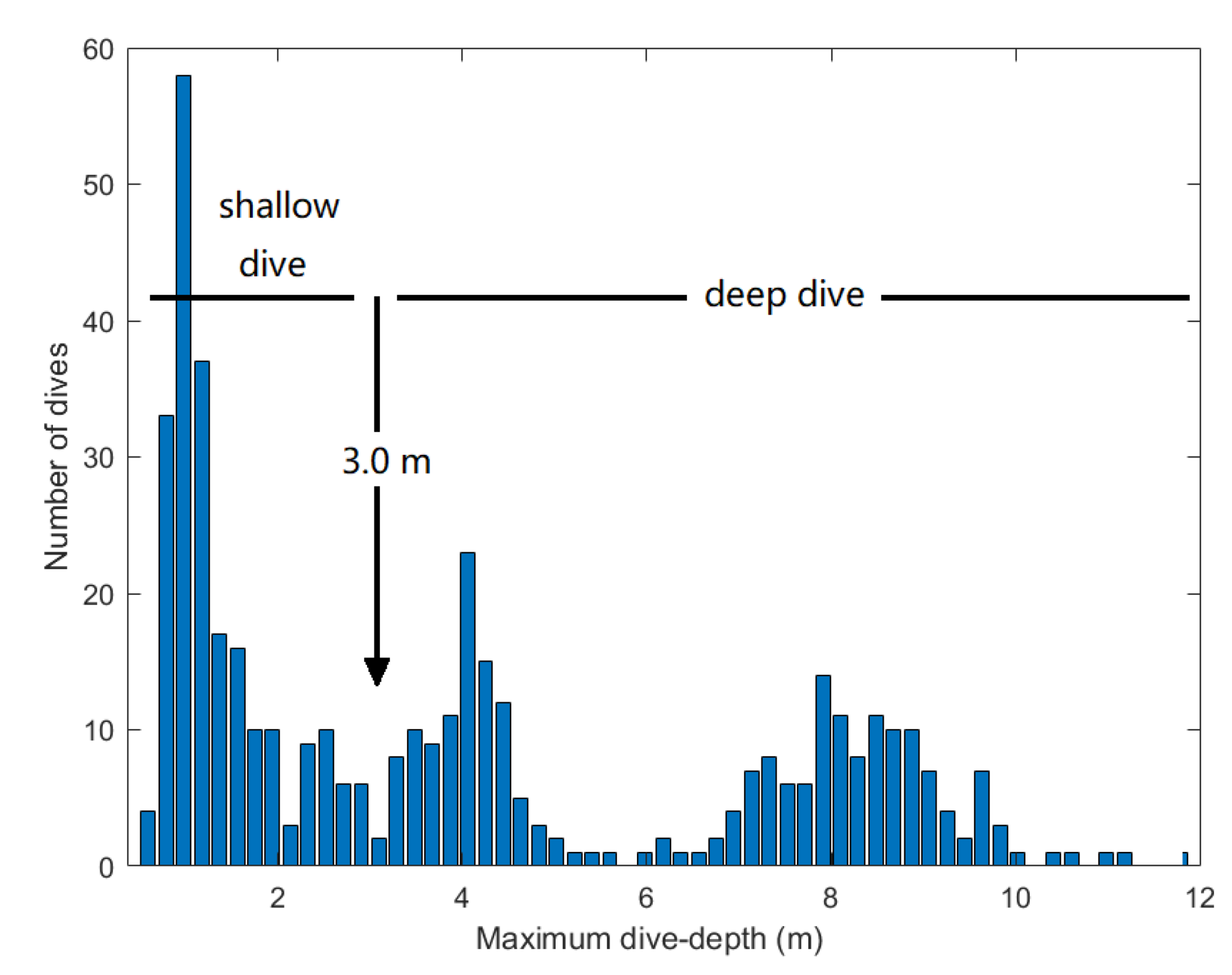
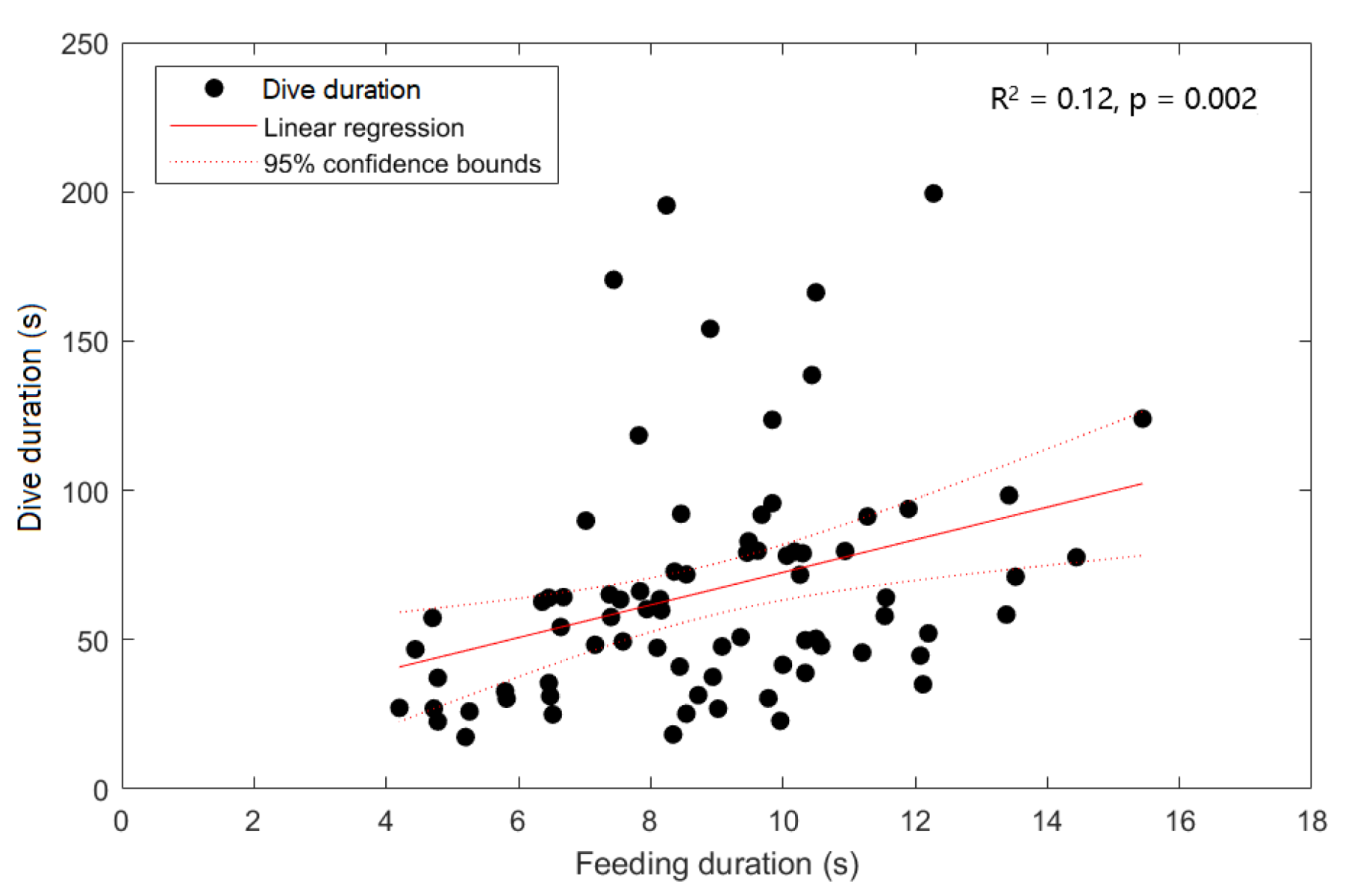
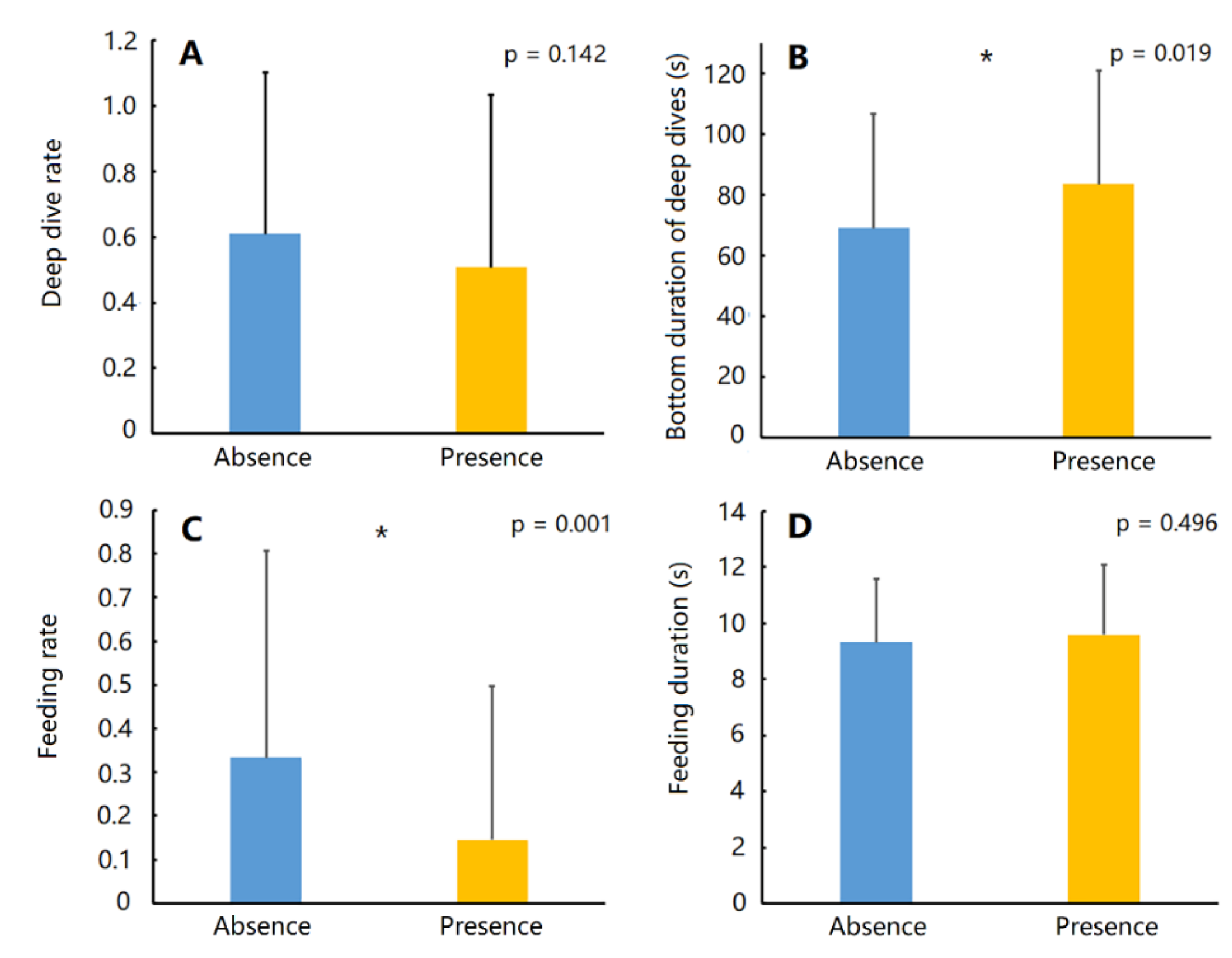
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boyd, I.L.; Kato, A.; Ropert-Coudert, Y. Bio-logging science: Sensing beyond the boundaries. Mem. Natl. Inst. Polar Res. 2004, 58, 1–14. [Google Scholar]
- Grothues, T.M. A review of acoustic telemetry technology and a perspective on its diversification relative to coastal tracking arrays. In Tagging and Tracking of Marine Animals with Electronic Devices; Nielsen, J.L., Ed.; Springer Science & Business Media: Heidelberg, Germany, 2009; pp. 77–90. [Google Scholar]
- Rutz, C.; Hays, G.C. New frontiers in biologging science. Biol. Lett. 2009, 5, 289–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerschlag, N.; Gallagher, A.; Lazarre, D. A review of shark satellite tagging studies. J. Exp. Mar. Biol. Ecol. 2011, 398, 1–8. [Google Scholar] [CrossRef]
- Stone, G.; Schubel, J.; Tausig, H. Electronic marine animal tagging: New frontier in ocean science. Oceanography 1999, 12, 24–27. [Google Scholar] [CrossRef] [Green Version]
- Bograd, S.J.; Block, B.A.; Costa, D.P.; Godley, B.J. Biologging technologies: New tools for conservation. Introduction. Endang. Species Res. 2010, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Evans, K.; Lea, M.A.; Patterson, T.A. Recent advances in bio-logging science: Technologies and methods for understanding animal behaviour and physiology and their environments. Deep Sea Res. II Top. Stud. Oceanogr. 2013, 88–89, 1–6. [Google Scholar] [CrossRef]
- Baumgartner, M.F.; Hammar, T.; Robbins, J. Development and assessment of a new dermal attachment for short-term tagging studies of baleen whales. Methods Ecol. Evol. 2015, 6, 289–297. [Google Scholar] [CrossRef]
- Szesciorka, A.R.; Calambokidis, J.; Harvey, J.T. Testing tag attachments to increase the attachment duration of archival tags on baleen whales. Anim. Biotelemet. 2016, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Guzman, H.M.; Capella, J.J. Short-term recovery of humpback whales after percutaneous satellite tagging. J. Wildl. Manag. 2017, 81, 728–733. [Google Scholar] [CrossRef]
- Liu, M.; Lin, W.; Lin, M.; Liu, B.; Dong, L.; Zhang, P.; Yang, Z.; Wang, K.; Dai, L.; Li, S. The First Attempt of Satellite Tracking on Occurrence and Migration of Bryde’s Whale (Balaenoptera edeni) in the Beibu Gulf. J. Mar. Sci. Eng. 2021, 9, 796. [Google Scholar] [CrossRef]
- Tyson, R.B.; Friedlaender, A.S.; Ware, C.; Stimpert, A.K.; Nowacek, D.P. Synchronous mother and calf foraging behaviour in humpback whales Megaptera novaeangliae: Insights from multi-sensor suction cup tags. Mar. Ecol. Prog. Ser. 2012, 457, 209–220. [Google Scholar] [CrossRef]
- Goldbogen, J.A.; Calambokidis, J.; Friedlaender, A.S.; Francis, J.; DeRuiter, S.L.; Stimpert, A.K.; Falcone, E.; Southall, B.L. Underwater acrobatics by the world’s largest predator: 360 rolling manoeuvres by lunge-feeding blue whales. Biol. Lett. 2013, 9, 20120986. [Google Scholar] [CrossRef] [PubMed]
- Tackaberry, J.E.; Cade, D.E.; Goldbogen, J.A.; Wiley, D.N.; Friedlaender, A.S.; Stimpert, A.K. From a calf’s perspective: Humpback whale nursing behavior on two US feeding grounds. PeerJ 2020, 8, e8538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldbogen, J.; Calambokidis, J.; Shadwick, R.; Oleson, E.; McDonald, M.; Hildebrand, J. Kinematics of diving and lunge-feeding in fin whales. J. Exp. Biol. 2006, 209, 1231–1244. [Google Scholar] [CrossRef] [Green Version]
- Chapple, T.K.; Gleiss, A.C.; Jewell, O.J.D.; Wikelski, M.; Block, B.A. Tracking sharks without teeth: A non-invasive rigid tag attachment for large predatory sharks. Anim. Biotelemet. 2015, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Cade, D.E.; Friedlaender, A.S.; Calambokidis, J.; Goldbogen, J.A. Kinematic Diversity in Rorqual Whale Feeding Mechanisms. Curr. Biol. 2016, 26, 2617–2624. [Google Scholar] [CrossRef] [Green Version]
- Caruso, F.; Hickmott, L.; Warren, J.D.; Segre, P.; Chiang, G.; Bahamonde, P.; ESPAÑOL-JIMÉNEZ, S.; Li, S.; Bocconcelli, A. Diel differences in blue whale (Balaenoptera musculus) dive behavior increase nighttime risk of ship strikes in northern Chilean Patagonia. Integr. Zool. 2020, 16, 594–611. [Google Scholar] [CrossRef]
- Meynecke, J.O.; Liebsch, N. Asset Tracking Whales—First Deployment of a Custom-Made GPS/GSM Suction Cup Tag on Migrating Humpback Whales. J. Mar. Sci. Eng. 2021, 9, 597. [Google Scholar] [CrossRef]
- Nelms, S.E.; Alfaro-Shigueto, J.; Arnould, J.P.; Avila, I.C.; Nash, S.B.; Campbell, E.; Carter, M.I.; Collins, T.; Currey, R.J.; Domit, C. Marine mammal conservation: Over the horizon. Endang. Species Res. 2021, 44, 291–325. [Google Scholar] [CrossRef]
- Kato, H.; Perrin, W.F. Bryde’s Whales: Balaenoptera edeni/brydei, in Encyclopedia of Marine Mammals; Elsevier: Burlington, MA, USA, 2009; pp. 158–163. [Google Scholar]
- Tezanos-Pinto, G.; Hupman, K.; Wiseman, N.; Dwyer, S.; Baker, C.; Brooks, L.; Outhwaite, B.; Lea, C.; Stockin, K. Local abundance, apparent survival and site fidelity of Bryde’s whales in the Hauraki Gulf (New Zealand) inferred from long-term photo-identification. Endang. Species Res. 2017, 34, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Kershaw, F.; Leslie, M.S.; Collins, T.; Mansur, R.M.; Smith, B.D.; Minton, G.; Baldwin, R.; LeDuc, R.G.; Anderson, R.C.; Brownell Jr, R.L. Population differentiation of 2 forms of Bryde’s whales in the Indian and Pacific Oceans. J. Hered. 2013, 104, 755–764. [Google Scholar] [CrossRef]
- Cooke, J.G.; Brownell, R.L., Jr. Balaenoptera edeni. The IUCN Red List of Threatened Species 2018: ET2476A50349178. 2018. Available online: https://www.iucnredlist.org/species/2476/50349178 (accessed on 1 September 2022).
- Corkeron, P.; Reeves, R.; Rosel, P. Balaenoptera edeni (Gulf of MEXICO Subpopulation). The IUCN Red List of Threatened Species 2017; e.T117636167A117636174. 2017. Available online: https://www.iucnredlist.org/species/117636167/117636174 (accessed on 1 September 2022).
- Constantine, R.; Iwata, T.; Nieukirk, S.L.; Penry, G.S. Future directions in research on Bryde’s whales. Front. Mar. Sci. 2018, 5, 333. [Google Scholar] [CrossRef] [Green Version]
- Wang, P. Chinese Cetaceans; Chemical Industrial Press: Beijing, China, 2012. [Google Scholar]
- Parsons, E.; Felley, M.L.; Porter, L.J. An annotated checklist of cetaceans recorded from Hong Kong’s territorial waters. Asian Mar. Biol. 1995, 12, 79–100. [Google Scholar]
- Kato, H.; Shinohara, E.; Kishiro, T.; Noji, S. Distribution of Bryde’s whales off Kochi, southwest Japan, from the 1994/95 sighting survey. Rep. Int. Whaling. Comm. 1996, 46, 429–436. [Google Scholar]
- Jefferson, T.A.; Hung, S.K. An updated, annotated checklist of the marine mammals of Hong Kong. Mammalia 2007, 71, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Lin, M.; Li, S. Species diversity and spatiotemporal patterns based on cetacean stranding records in China, 1950–2018. Sci. Total Environ. 2022, 822, 153651. [Google Scholar] [CrossRef]
- Wada, S.; Oishi, M.; Yamada, T.K. A newly discovered species of living baleen whale. Nature 2003, 426, 278–281. [Google Scholar] [CrossRef]
- Chen, B.; Zhu, L.; Jefferson, T.; Zhou, K.; Yang, G. Coastal Bryde’s Whales’ (Balaenoptera edeni) Foraging Area Near Weizhou Island in the Beibu Gulf. Aquat. Mamm. 2019, 45, 274–279. [Google Scholar] [CrossRef]
- Chen, M.; Huang, S.-L.; Wu, H.; Mei, Z.; Chen, Y.; Zheng, J.; Cheng, W.; Deng, K.; Yang, C.; Mao, C.; et al. Occurrence of Bryde’s whales, Balaenoptera edeni, in the northern Beibu Gulf, China. Mar. Mamm. Sci. 2019, 35, 1643–1652. [Google Scholar] [CrossRef]
- Chantrapornsyl, S.; Adulyanukosol, K.; Kittiwathanawong, K. Records of cetaceans in Thailand. Phuket Mar. Biol. Res. Bull. 1996, 61, 39–63. [Google Scholar]
- Iwata, T.; Akamatsu, T.; Thongsukdee, S.; Cherdsukjai, P.; Adulyanukosol, K.; Sato, K. Tread-water feeding of Bryde’s whales. Curr. Biol. 2017, 27, R1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, M.; Mao, C.; Han, Y.; Hao, Y.; Wang, K.; Mei, Z.; Wang, D. Community-based population monitoring for large baleen whales: The case study of Bryde’s whale in Beibu Gulf of China. Integr. Zool. 2021, 16, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.J. Biotelemetry and biologging in endangered species research and animal conservation: Relevance to regional, national, and IUCN Red List threat assessments. Endang. Species Res. 2008, 4, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Izadi, S.; Johnson, M.; de Soto, N.A.; Constantine, R. Night-life of Bryde’s whales: Ecological implications of resting in a baleen whale. Behav. Ecol. Sociobiol. 2018, 72, 78. [Google Scholar] [CrossRef]
- Croll, D.; Acevedo-Gutierrez, A.; Tershy, B. The diving behavior of blue and fin whales: Is dive duration shorter than expected based on oxygen stores? Comp. Biochem. Physiol. A 2001, 129, 797–809. [Google Scholar] [CrossRef]
- Akamatsu, T.; Wang, D.; Wang, K.; Wei, Z.; Zhao, Q.; Naito, Y. Diving behaviour of freshwater finless porpoises (Neophocaena phocaenoides) in an oxbow of the Yangtze River, China. ICES J. Mar. Sci. 2002, 59, 438–443. [Google Scholar] [CrossRef]
- Panigada, S.; Zanardelli, M.; Canese, S.; Jahoda, M. How deep can baleen whales dive? Mar. Ecol. Prog. Ser. 1999, 187, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Alves, F.; Dinis, A.; Cascão, I.; Freitas, L. Bryde’s whale (Balaenoptera brydei) stable associations and dive profiles: New insights into foraging behavior. Mar. Mamm. Sci. 2010, 26, 202–212. [Google Scholar] [CrossRef]
- Soldevilla, M.S.; Hildebrand, J.A.; Frasier, K.E.; Dias, L.A.; Martinez, A.; Mullin, K.D.; Rosel, P.E.; Garrison, L.P. Spatial distribution and dive behavior of Gulf of Mexico Bryde’s whales: Potential risk of vessel strikes and fisheries interactions. Endang. Species Res. 2017, 32, 533–550. [Google Scholar] [CrossRef] [Green Version]
- Goldbogen, J.A.; Friedlaender, A.S.; Calambokidis, J.; McKenna, M.F.; Simon, M.; Nowacek, D.P. Integrative approaches to the study of baleen whale diving behavior, feeding performance, and foraging ecology. Bioscience 2013, 63, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Friedlaender, A.; Goldbogen, J.; Nowacek, D.; Read, A.; Johnston, D.; Gales, N. Feeding rates and under-ice foraging strategies of the smallest lunge filter feeder, the Antarctic minke whale (Balaenoptera bonaerensis). J. Exp. Biol. 2014, 217, 2851–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cade, D.E.; Seakamela, S.M.; Findlay, K.P.; Fukunaga, J.; Kahane-Rapport, S.R.; Warren, J.D.; Calambokidis, J.; Fahlbusch, J.A.; Friedlaender, A.S.; Hazen, E.L. Predator-scale spatial analysis of intra-patch prey distribution reveals the energetic drivers of rorqual whale super-group formation. Funct. Ecol. 2021, 35, 894–908. [Google Scholar] [CrossRef]
- Villagra, D.; García-Cegarra, A.; Gallardo, D.I.; Pacheco, A.S. Energetic effects of whale-watching boats on humpback whales on a breeding ground. Front. Mar. Sci. 2021, 7, 600508. [Google Scholar] [CrossRef]
- Schaffar, A.; Madon, B.; Garrigue, C.; Constantine, R. Behavioural effects of whale-watching activities on an Endangered population of humpback whales wintering in New Caledonia. Endang. Species Res. 2013, 19, 245–254. [Google Scholar] [CrossRef]
- Di Clemente, J.; Christiansen, F.; Pirotta, E.; Steckler, D.; Wahlberg, M.; Pearson, H.C. Effects of whale watching on the activity budgets of humpback whales, Megaptera novaeangliae (Borowski, 1781), on a feeding ground. Aquat. Conserv. Mar. Freshwat. Ecosyst. 2018, 28, 810–820. [Google Scholar] [CrossRef]
- Sprogis, K.R.; Bejder, L.; Hanf, D.; Christiansen, F. Behavioural responses of migrating humpback whales to swim-with-whale activities in the Ningaloo Marine Park, Western Australia. J. Exp. Mar. Biol. Ecol. 2020, 522, 151254. [Google Scholar] [CrossRef]
- Hooker, S.K.; Baird, R.W. Diving and ranging behaviour of odontocetes: A methodological review and critique. Mamm. Rev. 2001, 31, 81–105. [Google Scholar] [CrossRef]
- Cade, D.E.; Barr, K.R.; Calambokidis, J.; Friedlaender, A.S.; Goldbogen, J.A. Determining forward speed from accelerometer jiggle in aquatic environments. J. Exp. Biol. 2018, 221, jeb170449. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.J.; Johnson, M.P.; Tyack, P.L.; Terray, E.A. Swimming gaits, passive drag and buoyancy of diving sperm whales Physeter macrocephalus. J. Exp. Biol. 2004, 207, 1953–1967. [Google Scholar] [CrossRef] [Green Version]
- Stimpert, A.; Mattila, D.; Nosal, E.-M.; Au, W. Tagging young humpback whale calves: Methodology and diving behavior. Endang. Species Res. 2012, 19, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Keen, E.; Falcone, E.; Andrews, R.; Schorr, G. Diel Dive Behavior of Fin Whales (Balaenoptera physalus) in the Southern California Bight. Aquat. Mamm. 2019, 45, 233–243. [Google Scholar] [CrossRef]
- Irvine, B. Behavioral changes in dolphins in a strange environment. Q. J. Fla. Acad. Sci. 1971, 34, 206–212. [Google Scholar]
- Rheingold, H.L.; Eckerman, C.O. Familiar social and nonsocial stimuli and the kitten’s response to a strange environment. Dev. Psychobiol.: J. Int. Soc. Dev. Psychobiol. 1971, 4, 71–89. [Google Scholar] [CrossRef]
- Kummrow, M. Diagnostic and therapeutic guidelines to abnormal behavior in captive nonhuman primates. Vet. Clin. Exot. Anim. 2021, 24, 253–266. [Google Scholar] [CrossRef]
- Mate, B.R.; Lagerquist, B.A.; Calambokidis, J. Movements of North Pacific Blue whales during the feeding season off Southern California and their southern fall migration. Mar. Mamm. Sci. 1999, 15, 1246–1257. [Google Scholar] [CrossRef]
- Murase, H.; Tamura, T.; Otani, S.; Nishiwaki, S. Satellite tracking of Bryde’s whales Balaenoptera edeni in the offshore western North Pacific in summer 2006 and 2008. Fish. Sci. 2016, 82, 35–45. [Google Scholar] [CrossRef]
- Bordes, F.; Morand, S. The impact of multiple infections on wild animal hosts: A review. Infect. Ecol. Epidemiol. 2011, 1, 7346. [Google Scholar] [CrossRef] [Green Version]
- Ghai, R.R.; Fugere, V.; Chapman, C.A.; Goldberg, T.L.; Davies, T.J. Sickness behaviour associated with non-lethal infections in wild primates. Proc. R. Soc. B 2015, 282, 20151436. [Google Scholar] [CrossRef] [Green Version]
- Bohn, S.J.; Turner, J.M.; Warnecke, L.; Mayo, C.; McGuire, L.; Misra, V.; Bollinger, T.K.; Willis, C.K. Evidence of ‘sickness behaviour’in bats with white-nose syndrome. Behaviour 2016, 153, 981–1003. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, D.G.; Jones, M.E.; Cameron, E.Z.; Kerlin, D.H.; McCallum, H.; Storfer, A.; Hohenlohe, P.A.; Hamede, R.K. Infectious disease and sickness behaviour: Tumour progression affects interaction patterns and social network structure in wild Tasmanian devils. Proc. R. Soc. B 2020, 287, 20202454. [Google Scholar] [CrossRef]
- Blersch, R.; Bonnell, T.R.; Ganswindt, A.; Young, C.; Barrett, L.; Henzi, S.P. Sick and tired: Sickness behaviour, polyparasitism and food stress in a gregarious mammal. Behav. Ecol. Sociobiol. 2021, 75, 169. [Google Scholar] [CrossRef]
- Mayer, J. Use of behavior analysis to recognize pain in small mammals. Lab. Anim. 2007, 36, 43–48. [Google Scholar] [CrossRef]
- Long, X.; Chen, P.; Yuan, H.; Feng, X.; Yu, J.; Shu, L.; Li, X.; Chen, W.; Chen, Z. Evaluation on carrying capacity of fishery resources in coastal waters of Dapeng Peninsula, Shenzhen. South China Fish. Sci. 2019, 15, 56–64. [Google Scholar]
- Zhang, J.; Qiu, J.-W.; Chen, C.-L.; Zeng, J.-W.; Wang, X.-F. Analysis of the Fish Community Structure and Its Relationship with Environmental Factors in Autumn and Spring in Northern Dapeng Bay, Guangdong. J. Guangdong Ocean. Univ. 2020, 40, 43–52. [Google Scholar]
- Penry, G.S.; Cockcroft, V.G.; Hammond, P.S. Seasonal fluctuations in occurrence of inshore Bryde’s whales in Plettenberg Bay, South Africa, with notes on feeding and multispecies associations. Afr. J. Mar. Sci. 2011, 33, 403–414. [Google Scholar] [CrossRef]
- Wiseman, N.; Parsons, S.; Stockin, K.A.; Baker, C.S. Seasonal occurrence and distribution of Bryde’s whales in the Hauraki Gulf, New Zealand. Mar. Mamm. Sci. 2011, 27, E253–E267. [Google Scholar] [CrossRef]
- Maciel, I.S.; Tardin, R.H.; Simão, S.M. Occurrence and habitat use of Bryde’s whales (Balaenoptera edeni) in the Cabo Frio region, South-eastern Brazil. J. Mar. Biol. Assoc. UK 2018, 98, 1081–1086. [Google Scholar] [CrossRef]
- Jia, C.; Zhuang, S. Current situation and evaluation of fishery resources in the east coast of Shenzhen city. China Fish. 2009, 8, 21–23. [Google Scholar]
- Yoshida, H.; Kato, H. Phylogenetic relationships of Bryde’s whales in theWestern North Pacific and adjacent waters inferred from mitochondrial DNA sequences. Mar. Mamm. Sci. 1999, 15, 1269–1286. [Google Scholar] [CrossRef]
- Kanda, N.; Goto, M.; Kato, H.; McPhee, M.V.; Pastene, L.A. Population genetic structure of Bryde’s whales (Balaenoptera brydei) at the inter-oceanic and trans-equatorial levels. Conserv. Genet. 2007, 8, 853–864. [Google Scholar] [CrossRef]
- Erbe, C. Underwater Acoustics: Noise and the Effects on Marine Mammal—A Pocket Handbook, 3rd ed.; JASCO Applied Sciences: Capalaba, Australia, 2011; Volume 164, p. 64. [Google Scholar]
- Popper, A.N.; Hawkins, A. The Effects of Noise on Aquatic Life; Springer Science & Business Media: Heidelberg, Germany, 2012; Volume 730. [Google Scholar]
- Finneran, J.J. Noise-induced hearing loss in marine mammals: A review of temporary threshold shift studies from 1996 to 2015. J. Acoust. Soc. Am. 2015, 138, 1702–1726. [Google Scholar] [CrossRef]
- Christiansen, F.; Rasmussen, M.; Lusseau, D. Whale watching disrupts feeding activities of minke whales on a feeding ground. Mar. Ecol. Prog. Ser. 2013, 478, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Enrico, P.; Booth, C.G.; Costa, D.P.; Fleishman, E.; Kraus, S.D.; Lusseau, D.; Moretti, D.; New, L.F.; Schick, R.S.; Schwarz, L.K.; et al. Understanding the population consequences of disturbance. Ecol. Evol. 2018, 8, 9934–9946. [Google Scholar]
- Li, S.; Wu, H.; Xu, Y.; Peng, C.; Fang, L.; Lin, M.; Xing, L.; Zhang, P. Mid- to high-frequency noise from high-speed boats and its potential impacts on humpback dolphins. J. Acoust. Soc. Am. 2015, 138, 942–952. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Tag Deployment Time (GMT + 8) | Tag Retrieve Time (GMT + 8) | Deployment Location | Retrieve Location | Track Duration (min) | Total Numbers of Dives | Total Numbers of Feeding |
|---|---|---|---|---|---|---|---|
| 2 July 2021 | 12:51:16 | 16:32:14 | 22°35.681’ N 114°25.279’ E | 22°35.686’ N 114°25.551’ E | 220.97 | 278 | 45 |
| 4 July 2021 | 6:38:37 | 9:32:10 | 22°35.740’ N 114°25.473’ E | 22°35.478’ N 114°25.830’ E | 173.55 | 177 | 32 |
| Total | —— | —— | —— | —— | 394.52 | 455 | 77 |
| All Dives (n = 455) | Deep Dives (n = 238) | Shallow Dives (n = 217) | |
|---|---|---|---|
| Max. depth (m) | 4.03 ± 3.17 | 6.49 ± 2.48 | 1.33 ± 0.61 |
| Dive duration (s) | 46.52 ± 39.89 | 72.20 ± 38.71 | 18.36 ± 13.30 |
| Bottom duration (s) | 24.57 ± 30.67 | 38.11 ± 34.45 | 9.73 ± 15.90 |
| Max speed (m/s) | 2.40 ± 1.30 | 2.84 ± 1.52 | 1.91 ± 0.74 |
| Mean speed (m/s) | 0.61 ± 0.18 | 0.67 ± 0.15 | 0.53 ± 0.17 |
| Degree of descent angle | 29.35 ± 12.61 | 34.42 ± 13.60 | 23.77 ± 8.46 |
| Degree of ascent angle | 18.77 ± 14.33 | 25.50 ± 15.73 | 11.36 ± 6.97 |
| Descending rate (m/s) | 0.37 ± 0.19 | 0.44 ± 0.19 | 0.29 ± 0.15 |
| Ascending rate (m/s) | 0.49 ± 0.37 | 0.45 ± 0.20 | 0.53 ± 0.49 |
| Number of Tread-water feeding events | 77 | 77 | 0 |
| Statistics | With Feeding Behaviours (n = 77) | Without Feeding Behaviours (n = 161) | p Value | |
|---|---|---|---|---|
| Dive duration (s) | 66.59 ± 39.51 | 76.19 ± 37.78 | 0.015 * | |
| Descent | Duration (s) | 15.67 ± 7.88 | 18.16 ± 13.24 | 0.100 |
| Averaged rate (m/s) | 0.50 ± 0.21 | 0.42 ± 0.17 | 0.005 * | |
| Bottom | Duration (s) | 40.47 ± 38.84 | 36.98 ± 32.20 | 0.810 |
| Maximum depth (m) | 6.71 ± 2.19 | 6.39 ± 2.61 | 0.041 * | |
| Ascent | Duration (s) | 10.45 ± 3.37 | 21.05 ± 16.98 | 0.000 * |
| Averaged rate (m/s) | 0.55 ± 0.15 | 0.40 ± 0.21 | 0.000 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Liu, M.; Lin, W.; Li, S. First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours. J. Mar. Sci. Eng. 2022, 10, 1422. https://doi.org/10.3390/jmse10101422
Dong L, Liu M, Lin W, Li S. First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours. Journal of Marine Science and Engineering. 2022; 10(10):1422. https://doi.org/10.3390/jmse10101422
Chicago/Turabian StyleDong, Lijun, Mingming Liu, Wenzhi Lin, and Songhai Li. 2022. "First Suction Cup Tagging on a Small and Coastal Form Bryde’s Whale (Balaenoptera edeni edeni) in China to Investigate Its Dive Profiles and Foraging Behaviours" Journal of Marine Science and Engineering 10, no. 10: 1422. https://doi.org/10.3390/jmse10101422