
Characteristics of Chromophoric Dissolved Organic Matter (CDOM) Produced by Heterotrophic Bacteria Isolated from Aquaculture Systems

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Bacteria
2.2. Amplification of the 16S Ribosomal RNA Gene and Phenotypic Characterization
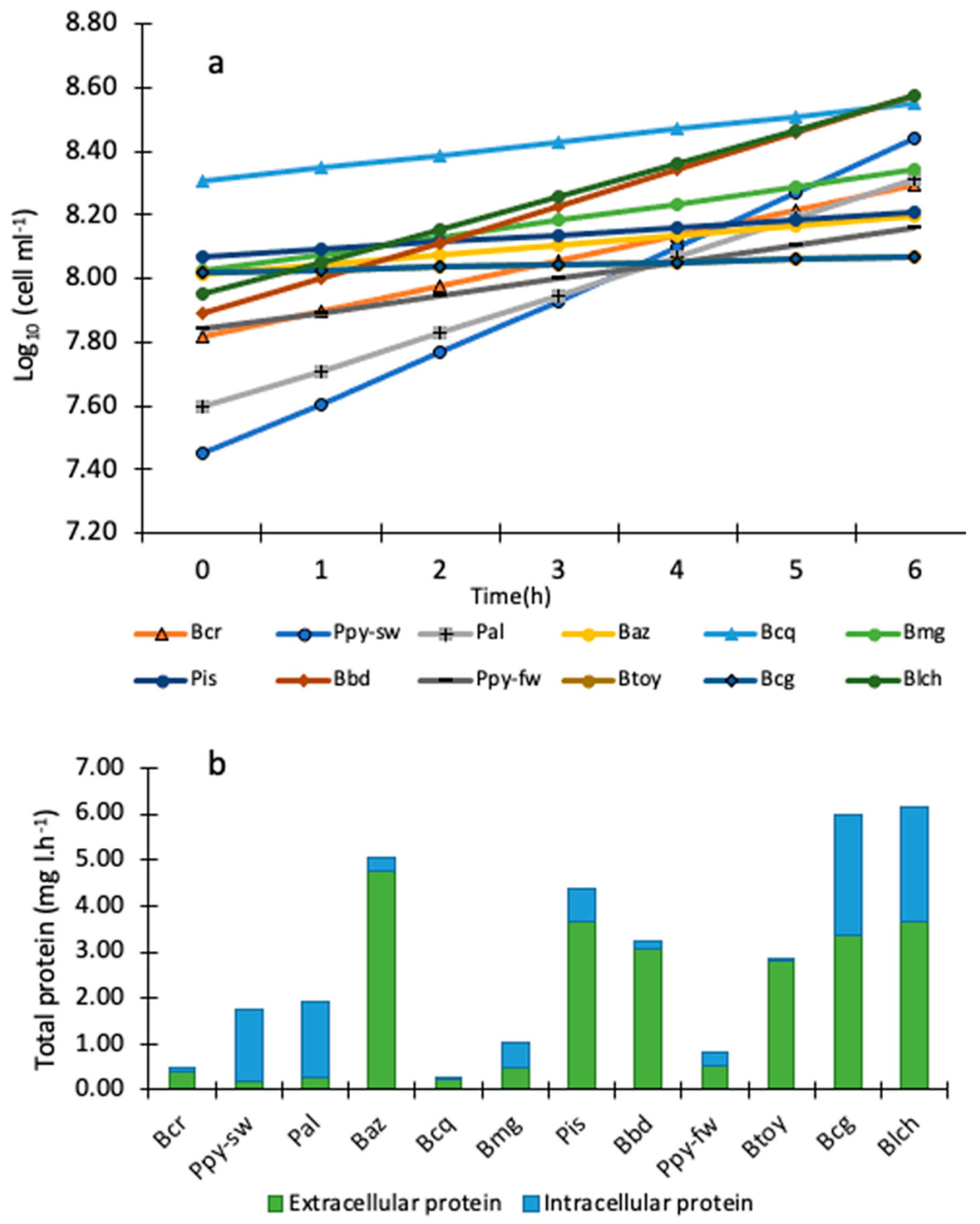
2.3. Bacterial Growth Curves
2.4. Separation of Extracellular (EC) and Intracellular (IC) Bacterial Compounds
2.5. Fluorescence Measurements
2.6. Data Analysis
3. Results
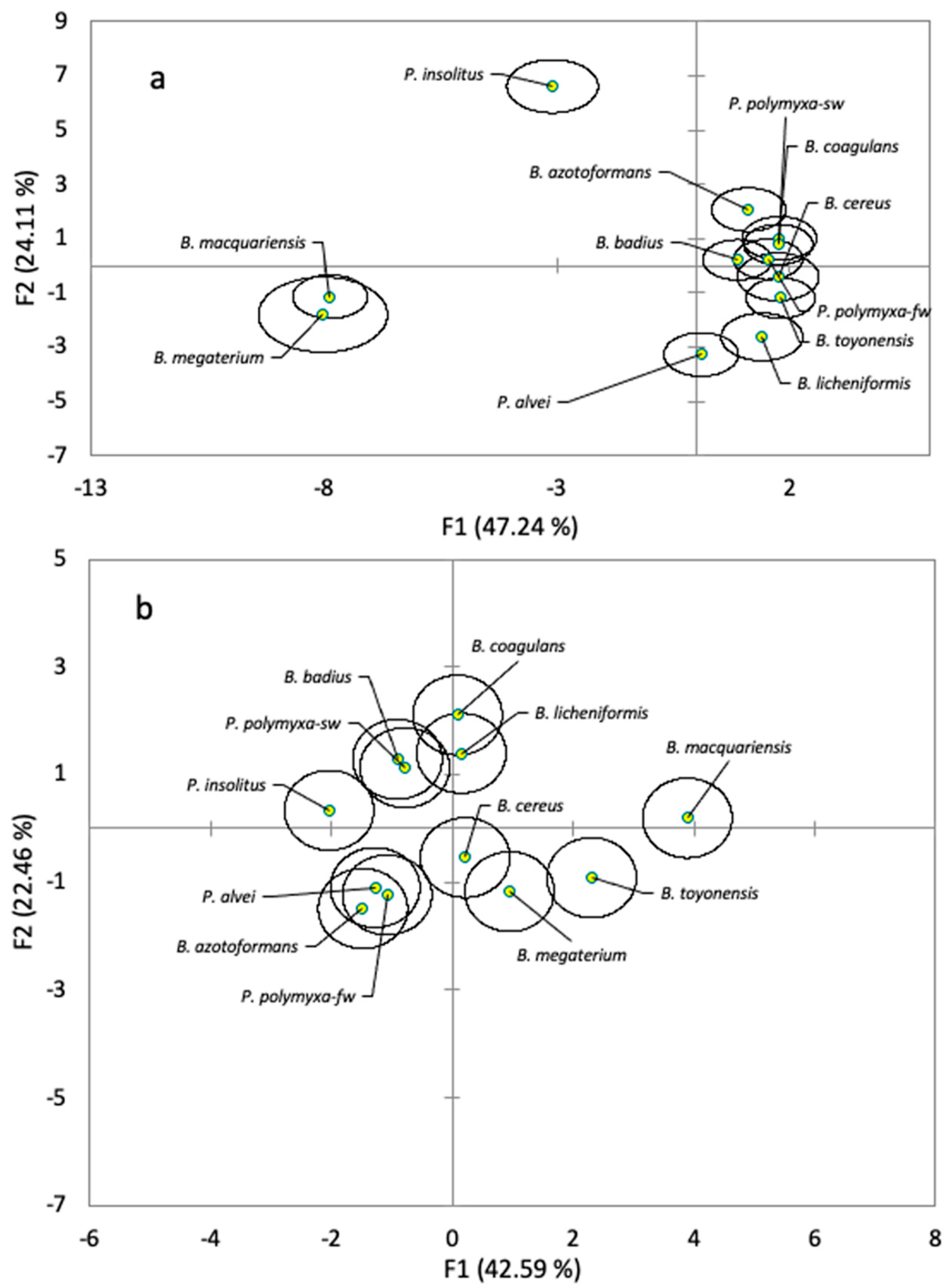
3.1. Phenotypic and Genotypic Identification of the Strains
3.2. Exponential Bacterial Growth
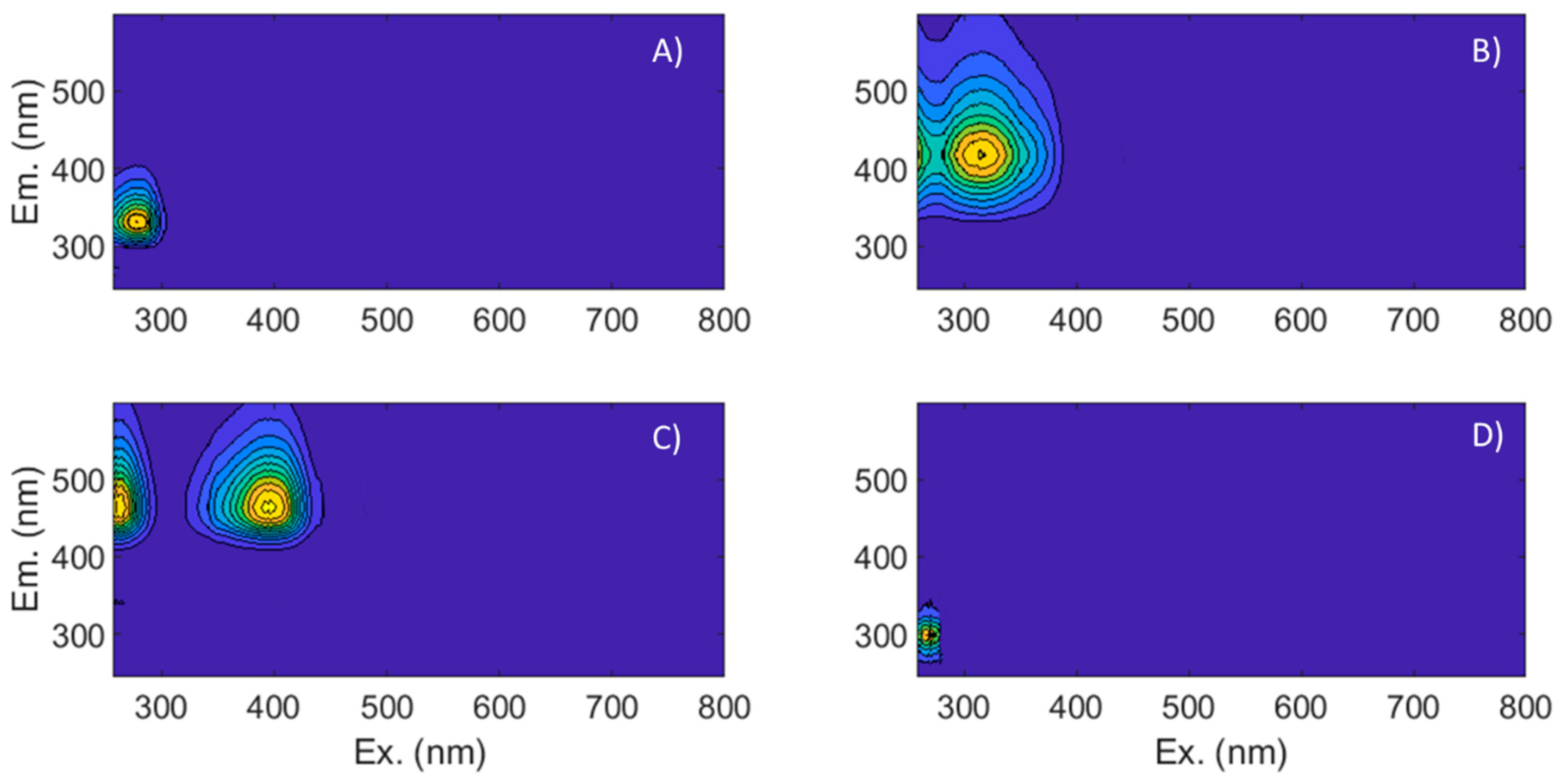
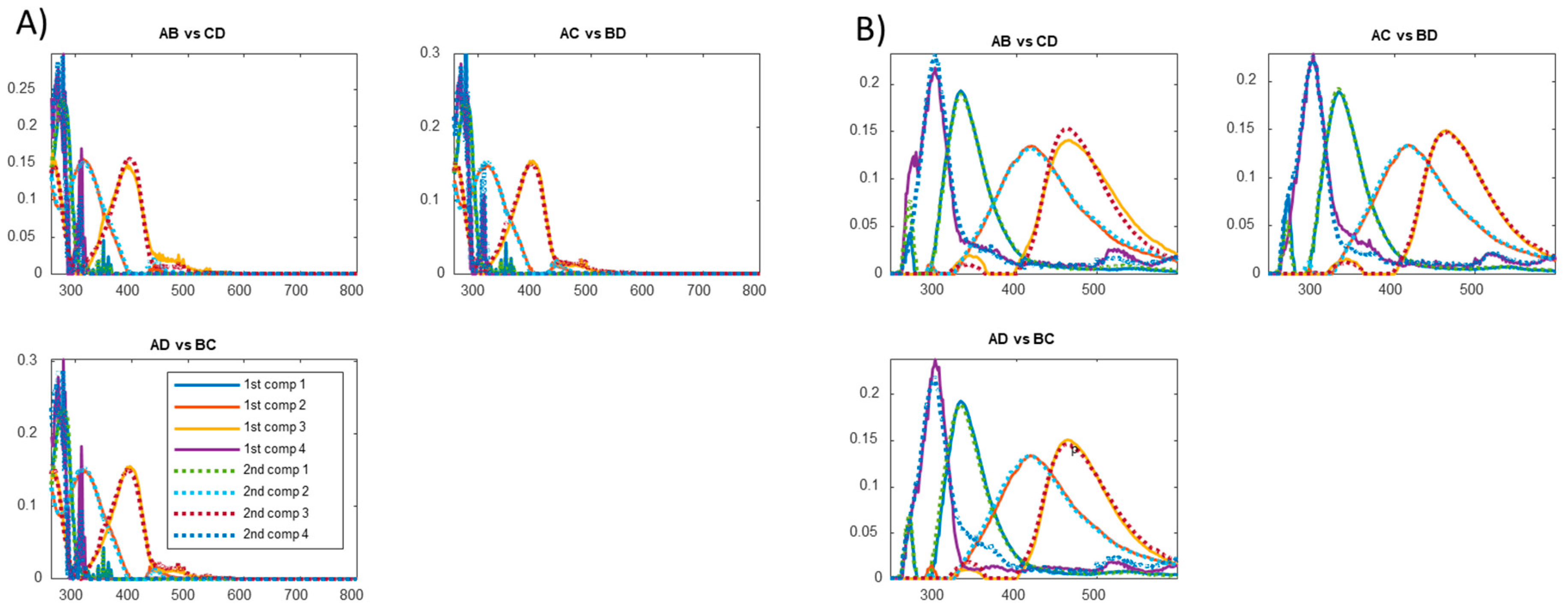
3.3. Fluorescence Characteristics and Identified Components
3.4. Biological Activity: Extracellular (EC) and Intracellular (IC) Bacterial Compounds
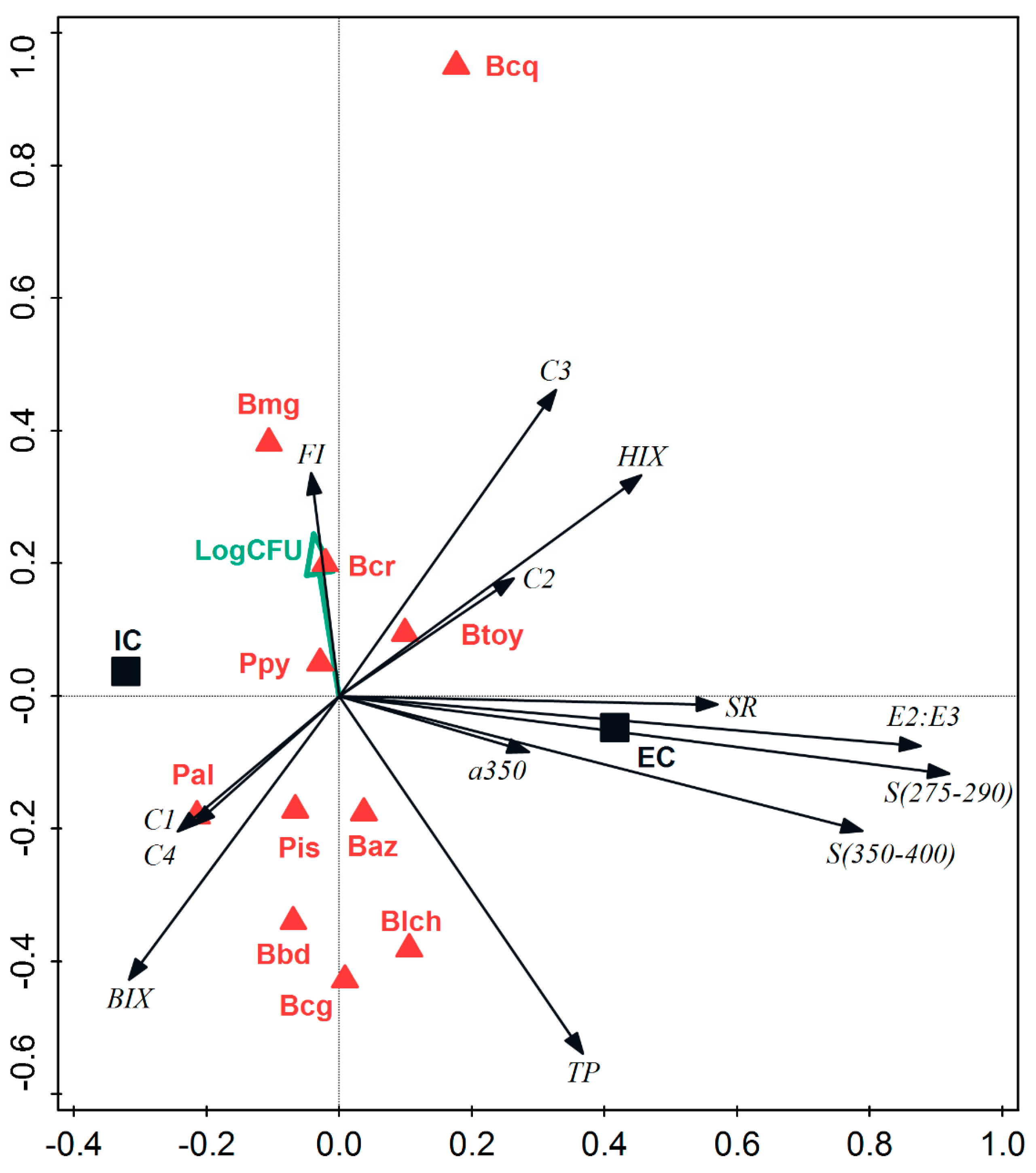
3.5. Differences between Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Buchan, A.; LeCleir, G.R.; Gulvik, C.A.; González, J.M. Master recyclers: Features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 2014, 12, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.A.; Giorgio, P.A.D.; Herndl, G.J. Microbes and the dissipation of energy and respiration: From cells to ecosystems. Oceanography 2007, 20, 89–100. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- Gram, L.; Grossart, H.P.; Schlingloff, A.; Kiorboe, T. Possible quorum sensing in marine snow bacteria: Production of acylated homoserine lactones by Roseobacter strains isolated from marine snow. Appl. Environ. Microbiol. 2002, 68, 4111–4116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.C.; Simon, M.; Alldredge, A.L.; Azam, F. Intense hydrolytic enzyme-activity on marine aggregates and implications for rapid particle dissolution. Nature 1992, 359, 139–142. [Google Scholar] [CrossRef]
- De Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F. Bergey’s Manual of Systematic Bacteriology. In The Firmicutes; Springer Science & Business Media: Berlin, Germany, 2011; Volume 3. [Google Scholar]
- Shimotori, K.; Watanabe, K.; Hama, T. Fluorescence characteristics of humic-like fluorescent dissolved organic matter produced by various taxa of marine bacteria. Aquat. Microb. Ecol. 2012, 65, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Noriega-Ortega, B.E.; Wienhausen, G.; Mentges, A.; Dittmar, T.; Simon, M.; Niggemann, J. Does the chemodiversity of bacterial exometabolomes sustain the chemodiversity of marine dissolved organic matter? Front. Microbiol. 2019, 10, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemette, F.; del Giorgio, P.A. Simultaneous consumption, and production of fluorescent dissolved organic matter by lake bacterioplankton. Environ. Microbiol. 2012, 14, 1432–1443. [Google Scholar] [CrossRef]
- Thompson, S.K.; Cotner, J.B. P-limitation drives changes in DOM production by aquatic bacteria. Aquat. Microb. Ecol. 2020, 85, 35–46. [Google Scholar] [CrossRef]
- Avnimelech, Y. Carbon/nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Milstein, A.; Avnimelech, Y.; Zoran, M.; Joseph, D. Growth performance of hybrid bass and hybrid tilapia in conventional and active suspension intensive ponds. Isr. J. Aquac. Bamidgeh 2001, 53, 147–157. [Google Scholar] [CrossRef]
- Suárez-Puerto, B.; Delgadillo-Díaz, M.; Sánchez-Solís, M.J.; Gullian-Klanian, M. Analysis of the cost-effectiveness and growth of Nile tilapia (Oreochromis niloticus) in biofloc and green water technologies during two seasons. Aquaculture 2021, 538, 736534. [Google Scholar] [CrossRef]
- De Schryver, P.; Crab, R.; Defoirdt, T.; Boon, N.; Verstraete, W. The basics of bio-flocs technology: The added value for aquaculture. Aquaculture 2008, 277, 125–137. [Google Scholar] [CrossRef]
- Gullian Klanian, M.; Delgadillo Díaz, M.; Sánchez Solís, M.J.; Aranda, J.; Moreno Moral, P. Effect of the content of microbial proteins and the poly-β-hydroxybutyric acid in biofloc on the performance and health of Nile tilapia (Oreochromis niloticus) fingerlings fed on a protein-restricted diet. Aquaculture 2020, 519, 602–612. [Google Scholar] [CrossRef]
- Attermeyer, K.; Hornick, T.; Kayler, Z.E.; Bahr, A.; Zwirnmann, E.; Grossart, H.-P.; Premke, K. Enhanced bacterial decomposition with increasing addition of autochthonous to allochthonous carbon without any effect on bacterial community composition. Biogeosciences 2014, 11, 1479–1489. [Google Scholar] [CrossRef] [Green Version]
- Stedmon, C.A.; Markager, S.; Kaas, H. Optical properties and signatures of chromophoric dissolved organic matter (CDOM) in Danish coastal waters. Estuar. Coast. Shelf Sci. 2000, 51, 267–278. [Google Scholar] [CrossRef]
- Coble, P.G. Characterization of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy. Mar. Chem. 1996, 51, 325–346. [Google Scholar] [CrossRef]
- Coble, P.G. Marine optical biogeochemistry: The chemistry of ocean color. Chem. Rev. 2007, 107, 402–418. [Google Scholar] [CrossRef]
- Kramer, G.; Herndl, G. Photo and bioreactivity of chromophoric dissolved organic matter produced by marine bacterioplankton. Aquat. Microb. Ecol. 2004, 36, 239–246. [Google Scholar] [CrossRef]
- Moran, M.A.; Sheldon, W.M.; Zepp, R.G. Carbon Loss and Optical Property Changes during Long-Term Photochemical and Biological Degradation of Estuarine Dissolved Organic Matter. Limnol. Oceanogr. 2010, 45, 1254–1264. [Google Scholar] [CrossRef]
- Shimotori, K.; Omori, Y.; Hama, T. Bacterial production of marine humic-like fluorescent dissolved organic matter and its biogeochemical importance. Aquat. Microb. Ecol. 2010, 58, 55–66. [Google Scholar] [CrossRef]
- Arai, K.; Wada, S.; Shimotori, K.; Omori, Y.; Hama, T. Production and degradation of fluorescent dissolved organic matter derived from bacteria. J. Oceanogr. 2018, 74, 39–52. [Google Scholar] [CrossRef]
- Landa, M.; Blain, S.; Christaki, U.; Monchy, S.; Obernosterer, I. Shifts in bacterial community composition associated with increased carbon cycling in a mosaic of phytoplankton blooms. ISMEJ 2016, 10, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Fox, B.G.; Thorn, R.M.S.; Anesio, A.M.; Reynolds, D.M. The in situ bacterial production of fluorescent organic matter; an investigation at a species level. Water Res. 2017, 125, 350–359. [Google Scholar] [CrossRef]
- Ni, B.J.; Fang, F.; Xie, W.M.; Sun, M.; Sheng, G.P.; Li, W.H.; Yu, H.Q. Characterization of extracellular polymeric substances produced by mixed microorganisms in activated sludge with gel-permeating chromatography, excitation-emission matrix fluorescence spectroscopy measurement and kinetic modeling. Water Res. 2009, 43, 1350–1358. [Google Scholar] [CrossRef]
- Miao, L.; Zhang, Q.; Wang, S.; Li, B.; Wang, Z.; Zhang, S.; Zhang, M.; Peng, Y. Characterization of EPS compositions and microbial community in an Anammox SBBR system treating landfill leachate. Bioresour. Technol. 2018, 249, 108–116. [Google Scholar] [CrossRef]
- Bai, Y.; Su, R.; Yao, Q.; Zhang, C.; Shi, X. Characterization of chromophoric dissolved organic matter (CDOM) in the Bohai Sea and the Yellow Sea using Excitation-Emission Matrix Spectroscopy (EEMs) and Parallel Factor Analysis (PARAFAC). Estuaries Coasts. 2017, 40, 1325–1345. [Google Scholar] [CrossRef]
- Hou, X.; Liu, S.; Feng, Y. The autofluorescence characteristics of bacterial intracellular and extracellular substances during the operation of anammox reactor. Sci. Rep. 2017, 7, 39289. [Google Scholar] [CrossRef]
- Parlanti, E.; Wörz, K.; Geoffroy, L.; Lamotte, M. Dissolved organic matter fluorescence spectroscopy as a tool to estimate biological activity in a coastal zone submitted to anthropogenic inputs. Org. Geochem. 2000, 31, 1765–1781. [Google Scholar] [CrossRef]
- Hudson, N.; Baker, A.; Reynolds, D. Fluorescence analysis of dissolved organic matter in natural, waste and polluted waters—A review. River Res. Appl. 2007, 23, 631–649. [Google Scholar] [CrossRef]
- Murphy, K.R.; Stedmon, C.A.; Waite, T.D.; Ruiz, G.M. Distinguishing between terrestrial and autochthonous organic matter sources in marine environments using fluorescence spectroscopy. Mar. Chem. 2008, 108, 40–58. [Google Scholar] [CrossRef]
- Stedmon, C.A.; Bro, R. Characterizing dissolved organic matter fluorescence with parallel factor analysis: A tutorial. Limnol. Oceanogr. Methods 2008, 6, 572–579. [Google Scholar] [CrossRef]
- Hambly, A.C.; Arvin, E.; Pedersen, L.F.; Pedersen, P.B.; Seredyńska-Sobecka, B.; Stedmon, C.A. Characterising organic matter in recirculating aquaculture systems with fluorescence EEM spectroscopy. Water Res. 2015, 83, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Gullian-Klanian, M.; Gold-Bouchot, G.; Delgadillo-Díaz, M.; Aranda, J.; Sánchez-Solís, M.J. Effect of the use of Bacillus spp. on the characteristics of dissolved fluorescent organic matter and the phytochemical quality of Stevia rebaudiana grown in a recirculating aquaponic system. Environ. Sci. Pollut. Res. 2021, 28, 26326–36343. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.; Tada, Y.; Suzuki, K.; Yamashita, Y. Production and reutilization of fluorescent dissolved organic matter by a marine bacterial strain, Alteromonas macleodii. Front. Microbiol. 2017, 8, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, S.; Tada, Y.; Suzuki, K.; Yamashita, Y. Evaluation of the production of dissolved organic matter by three marine bacterial strains. Front. Microbiol. 2020, 11, 584419. [Google Scholar] [CrossRef] [PubMed]
- Logan, N.A.; De Vos, P. Bacillus. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–163. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basical Local Alignment Search Tool (BLAST). J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Lane, D. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Liu, L.; Ji, M.; Wang, F.; Wang, S.; Qin, G. Insight into the influence of microbial aggregate types on nitrogen removal performance and microbial community in the anammox process—A review and meta-analysis. Sci. Total Environ. 2020, 714, 136571. [Google Scholar] [CrossRef]
- Ranganayaki, S.; Mohan, C. Effect of Sodium molybdate on microbial fixation of nitrogen. Zeitschrift für Allgemeine Mikrobiologie 1981, 21, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Lawaetz, A.J.; Stedmon, C.A. Fluorescence Intensity Calibration Using the Raman Scatter Peak of Water. Appl. Spectrosc. 2009, 63, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Pucher, M.; Wünsch, U.; Weigelhofer, G.; Murphy, K.; Hein, T.; Graeber, D. staRdom: Versatile Software for analyzing spectroscopic data of dissolved organic matter in R. Water 2019, 11, 2366. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Murphy, K.R.; Stedmon, C.A.; Graeber, D.; Bro, R. Fluorescence spectroscopy and multi-way techniques. PARAFAC. Anal. Methods 2013, 5, 6557–6566. [Google Scholar] [CrossRef] [Green Version]
- Stedmon, C.A.; Markager, S. Tracing the production and degradation of autochthonous fractions of dissolved organic matter by fluorescence analysis. Limnol Oceanog. 2005, 50, 1415–1426. [Google Scholar] [CrossRef]
- Massicotte, P.; Asmala, E.; Stedmon, C.; Markager, S. Global distribution of dissolved organic matter along the aquatic continuum: Across rivers, lakes and oceans. Sci. Total Environ. 2017, 609, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Loiselle, S.A.; Bracchini, L.; Cózar, A.; Dattilo, A.M.; Tognazzi, A.; Rossi, C. Variability in photobleaching yields and their related impacts on optical conditions in subtropical lakes. J. Photochem. Photobiol. B Biol. 2009, 95, 129–137. [Google Scholar] [CrossRef]
- Helms, J.; Stubbins, A.; Ritchie, J.D.; Minor, E.C.; Kieber, D.J.; Mopper, K. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol. Oceanogr. 2009, 53, 955–969. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Jin, H. Utilization of UV-Vis Spectroscopy and Related Data Analyses for Dissolved Organic Matter (DOM) Studies: A Review. Crit. Rev. Environ. Sci. Technol. 2017, 47, 131–154. [Google Scholar] [CrossRef]
- Massicotte, P.; Markager, S. Using a Gaussian decomposition approach to model absorption spectra of chromophoric dissolved organic matter. Mar. Chem. 2016, 180, 24–32. [Google Scholar] [CrossRef]
- Del Vecchio, R.; Blough, N.V. Photobleaching of chromophoric dissolved organic matter in natural waters: Kinetics and modeling. Mar. Chem. 2002, 78, 231–253. [Google Scholar] [CrossRef]
- Huguet, A.; Vacher, L.; Relexans, S.; Saubusse, S.; Froidefond, J.M.; Parlanti, E. Properties of fluorescent dissolved organic matter in the Gironde Estuary. Org. Geochem. 2009, 40, 706–719. [Google Scholar] [CrossRef]
- McKnight, D.M.; Boyer, E.W.; Westerhoff, P.K.; Doran, P.T.; Kulbe, T.; Andersen, D.T. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol. Oceanogr. 2001, 46, 38–48. [Google Scholar] [CrossRef]
- Massicotte, P. GitHube.com. Cánada. 2019. Available online: www.pmassicotte.com/eemr/ (accessed on 12 December 2021).
- ter Braak, C.J.F. Canonical community ordination. Part I: Basic theory and linear methods. Ecoscience 1994, 1, 127–140. [Google Scholar] [CrossRef]
- Murphy, K.R.; Stedmon, C.A.; Wenig, P.; Bro, R. OpenFluor- An online spectral library of auto-fluorescence by organic compounds in the environment. Anal. Methods 2014, 6, 658–661. [Google Scholar] [CrossRef] [Green Version]
- Cory, R.M.; McKnight, D.M. Fluorescence Spectroscopy Reveals Ubiquitous Presence of Oxidized and Reduced Quinones in Dissolved Organic Matter. Environ. Sci. Technol. 2005, 39, 8142–8149. [Google Scholar] [CrossRef]
- Cawley, K.M.; Ding, Y.; Fourqurean, J.; Jaffé, R. Characterising the sources and fate of dissolved organic matter in Shark Bay, Australia: A preliminary study using optical properties and stable carbon isotopes. Mar. Freshw. Res. 2012, 63, 1098–1107. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, Y.; Maie, N.; Briceño, H.; Jaffé, R. Optical characterization of dissolved organic matter in tropical rivers of the Guayana Shield, Venezuela. J. Geophys. Res. Biogeosci. 2010, 115, G1. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, Y.; Panton, A.; Mahaffey, C.; Jaffé, R. Assessing the spatial and temporal variability of dissolved organic matter in Liverpool Bay using excitation-emission matrix fluorescence and parallel factor analysis. Ocean. Dyn. 2011, 61, 569–579. [Google Scholar] [CrossRef]
- Cohen, E.; Levy, G.J.; Borisover, M. Fluorescent components of organic matter in wastewater: Efficacy and selectivity of the water treatment. Water Res. 2014, 55, 323–334. [Google Scholar] [CrossRef]
- Shakil, S.; Tank, S.E.; Kokelj, S.V.; Vonk, J.E.; Zolkos, S. Particulate dominance of organic carbon mobilization from thaw slumps on the Peel Plateau, NT: Quantification and implications for stream systems and permafrost carbon release. Environ. Res. Lett. 2020, 15, 114019. [Google Scholar] [CrossRef]
- Lee, S.A.; Kim, T.H.; Kim, G. Tracing terrestrial versus marine sources of dissolved organic carbon in a coastal bay using stable carbon isotopes. Biogeosciences 2020, 17, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Kowalczuk, P.; Tilstone, G.H.; Zabłocka, M.; Röttgers, R.; Thomas, R. Composition of dissolved organic matter along an Atlantic Meridional Transect from fluorescence spectroscopy and Parallel Factor Analysis. Mar. Chem. 2013, 157, 170–184. [Google Scholar] [CrossRef] [Green Version]
- Murphy, K.R.; Ruiz, G.M.; Dunsmuir, W.T.M.; Waite, T.D. Optimized Parameters for Fluorescence-Based Verification of Ballast Water Exchange by Ships. Environ. Sci. Technol. 2006, 40, 2357–2362. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef] [PubMed]
- Findlay, S.E.; Sinsabaugh, R.L.; Sobczak, W.V.; Hoostal, M. Metabolic and structural response of hyporheic microbial communities to variations in supply of dissolved organic matter. Limnol. Oceanogr. 2003, 48, 1608–1617. [Google Scholar] [CrossRef] [Green Version]
- Fellman, J.B.; Hood, E.; Spencer, R.G. Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: A review. Limnol. Oceanogr. 2010, 55, 2452–2462. [Google Scholar] [CrossRef]
- Hoshino, T.; Takehashi, S.; Fujiwara, M.; Kasuya, T. Typhula maritima, a new species of Typhula collected from coastal dunes in Hokkaido, northern Japan. Mycoscience 2009, 50, 430–437. [Google Scholar] [CrossRef]
- Sharma, M.; Kumar, A. Optimization of xylanase secretion from Paenibacillus macquariensis. Curr. Trends Biotechnol. Pharm. 2012, 6, 190–195. [Google Scholar]
- Reddy, C.S.K.; Ghai, R.; Kalia, V.C. Polyhydroxyalkanoates: An overview. Bioresour. Technol. 2003, 87, 137–146. [Google Scholar] [CrossRef]
- Hansen, A.M.; Kraus, T.E.C.; Pellerin, B.A.; Fleck, J.A.; Downing, B.D.; Bergamaschi, D.A. Optical properties of dissolved organic matter (DOM): Effects of biological and photolytic degradation. Limnol. Oceanogr. 2016, 61, 1015–1032. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Fuentes, J.A.; Pérez-Rostro, C.I.; Hernández-Vergara, M.P.; Monroy-Dosta, M.D.C. Variation of the bacterial composition of biofloc and the intestine of Nile tilapia Oreochromis niloticus, cultivated using biofloc technology, supplied different feed rations. Aquac. Res. 2018, 49, 3658–3668. [Google Scholar] [CrossRef]
- Veith, B.; Herzberg, C.; Steckel, S.; Feesche, J.; Maurer, K.H.; Ehrenreich, P.; Bäumer, S.; Henne, A.; Liesegang, H.; Merkl, R.; et al. The complete genome sequence of Bacillus licheniformis DSM13, an organism with great industrial potential. Microb. Physiol. 2004, 7, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Rey, M.W.; Ramaiya, P.; Nelson, B.A.; Brody-Karpin, S.D.; Zaretsky, E.J.; Tang, M.; de Leon, A.L.; Xiang, H.; Gusti, V.; Clausen, I.G.; et al. Complete genome sequence of the industrial bacterium Bacillus licheniformis and comparisons with closely related Bacillus species. Genome Biol. 2004, 5, r77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anandaraj, B.; Vellaichamy, A.; Kachman, M.; Selvamanikandan, A.; Pegu, S.; Murugan, V. Co-production of two new peptide antibiotics by a bacterial isolate Paenibacillus alvei NP75. Biochem. Biophys. Res. Commun. 2009, 379, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, S.M.; Hamed, H.A.; Mouafi, F.E.; Abdelwahed, N.A. Extracellular metabolites produced by a novel strain, Bacillus alvei NRC-14: 3. Synthesis of a bioflocculant that has chitosan-like structure. Life Sci. J. 2011, 8, 883–890. [Google Scholar] [CrossRef]
- Guéguen, C.; Cuss, C.W. Characterization of aquatic dissolved organic matter by asymmetrical flow field-flow fractionation coupled to UV–Visible diode array and excitation emission matrix fluorescence. J. Chromatog. A 2011, 1218, 4188–4198. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Lee, C.M.; Kuan, C.H. Removal of 2, 4-dichlorophenol by suspended and immobilized Bacillus insolitus. Chemosphere 2000, 41, 447–452. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Code | Closest Phylogenetic Neighbor | NCBI:Txid | Source | Identity (%) |
|---|---|---|---|---|
| Baz | Bacillus azotoformans | 1131731 | SW | 100 |
| Bbd | Bacillus badius | 1455 | FW | 99 |
| Bcg | Bacillus coagulans | 1398 | FW | 99 |
| Bcq | Bacillus macquariensis | 1468 | FW | 100 |
| Bcr | Bacillus cereus | 226900 | SW | 100 |
| Blch | Bacillus licheniformis | 279010 | FW | 98 |
| Bmg | Bacillus megaterium | 1348623 | FW | 100 |
| Btoy | Bacillus toyonensis | 121761 | FW | 100 |
| Pal | Paenibacillus alvei | 1206781 | SW | 99 |
| Pis | Psychrobacillus insolitus | 1461 | FW | 99 |
| Ppy | Paenibacillus polymyxa | 886882 | FW | 100 |
| Ppy | Paenibacillus polymyxa | 886882 | SW | 99 |
| Test Name | B. macquariensis | B. badius | B. coagulans | B. licheniformis | P. insolitus | B. megaterium | B. cereus | P. alvei | P. polymyxa | P. toyonensis | B. azotoformans |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gram | + | + | + | + | + | + | + | + | + | + | + |
| Oxidase | + | - | + | - | - | - | - | + | + | ||
| Catalase | + | + | + | + | + | + | - | ||||
| Citrate | + | + | - | - | - | + | - | ||||
| Swollen cell | + | - | - | - | + | - | - | - | - | ||
| Voges–Proskauer | - | - | + | + | - | - | + | - | + | + | - |
| β-galactosidase | - | - | - | - | - | ||||||
| Lysine decarboxylase | - | + | - | - | - | + | - | + | + | + | |
| Ornithine decarboxylase | + | + | - | - | - | + | + | - | - | + | |
| Arginine decarboxylase | + | + | + | + | + | + | + | + | + | + | |
| Amygdalin | - | - | - | - | + | ||||||
| D-arabinose | - | - | + | - | + | + | - | - | - | - | - |
| Lactose | - | - | + | + | - | + | |||||
| Glucose | - | - | + | - | + | - | + | + | + | + | + |
| Inositol | - | - | - | - | - | + | - | - | - | - | - |
| Mannitol | - | - | - | + | - | - | - | - | - | - | - |
| Melibious | - | - | + | + | - | - | - | - | - | ||
| Rhamnose | - | - | - | - | + | - | - | - | - | - | |
| Sorbitol | - | - | - | - | + | - | - | - | - | + | |
| Urea | - | - | - | - | - | - | + | - | - | - | + |
| Starch | + | + | + | + | + | + | + | + | + | + | - |
| Growth in NaCl 6.5% | + | + | - | + | + | + | + | + | + | + | + |
| Bacteria | Cf (Cell mL−1) | Co (Cell mL−1) | K | R2 |
|---|---|---|---|---|
| B. azotoformans | 8.196 | 8.013 | 0.0037 | 0.88 |
| B. badious | 8.573 | 7.892 | 0.0138 | 0.89 |
| B. coagulans | 8.066 | 8.253 | 0.0035 | 0.90 |
| B. macquariensis | 8.551 | 8.307 | 0.0048 | 0.96 |
| B. cereus | 8.295 | 7.817 | 0.0098 | 0.91 |
| B. licheniformis | 8.571 | 7.841 | 0.0066 | 0.90 |
| B. megaterium | 8.342 | 8.022 | 0.0065 | 0.85 |
| B. toyonensis | 8.066 | 8.019 | 0.0009 | 0.97 |
| P. alvei | 8.311 | 7.595 | 0.0151 | 0.98 |
| P. insolitus | 8.206 | 8.068 | 0.0028 | 0.90 |
| P. polymyxa-FW | 8.158 | 7.841 | 0.0065 | 0.89 |
| P. polymyxa-SW | 8.441 | 7.448 | 0.0213 | 0.90 |
| Component | Interpretation | Reference |
|---|---|---|
| C1 | Protein-like | Cawley et al., [61] |
| Tryptophan-like, polyphenols | Yamashita et al., [62] | |
| Tryptophan-like, polyphenols | Yamashita et al., [63] | |
| Peak T, microbial | Hambly et al., [34] | |
| C2 | Terrestrial humic-like | Cawley et al., [61] |
| Marine humic-like, peak M | Yamashita et al., [62] | |
| Humic-like | Yamashita et al., [63] | |
| C3 | Humic-like | Cohen et al., [64] |
| Terrestrial humic/fulvic-like | Shakil et al., [65] | |
| Terrestrial humic-like | Lee et al., [66] | |
| C4 | Tryptophan and tyrosine-like | Cawley et al., [61] |
| Peak T, tryptophan | Kowalczuk et al., [67] | |
| Protein-like | Murphy et al., [68] |
| C1 | C2 | C3 | C4 | FI | HIX | BIX | |
|---|---|---|---|---|---|---|---|
| Extracellular compounds | |||||||
| Baz | 0.47 ± 0.10 | 0.52 ± 0.15 | 0.22 ± 0.06 | 0.39 ± 0.10 | 1.67 ± 0.03 | 1.32 ± 0.23 | 0.82 ± 0.01 |
| Bbd | 0.67 ± 0.15 | 0.49 ± 0.09 | 0.18 ± 0.05 | 2.79 ± 1.49 | 1.39 ± 0.03 | 0.78 ± 0.16 | 1.04 ± 0.02 |
| Bcg | 0.50 ± 0.05 | 0.51 ± 0.02 | 0.18 ± 0.01 | 0.88 ± 0.61 | 1.55 ± 0.02 | 1.21 ± 0.14 | 0.91 ± 0.01 |
| Bcq | 1.64 ± 0.36 | 0.33 ± 0.09 | 5.48 ± 1.15 | 0.49 ± 0.10 | 1.52 ± 0.01 | 3.50 ± 0.90 | 0.63 ± 0.09 |
| Bcr | 0.32 ± 0.05 | 0.29 ± 0.06 | 0.13 ± 0.03 | 0.14 ± 0.02 | 1.45 ± 0.05 | 1.06 ± 0.13 | 0.96 ± 0.03 |
| Blch | 0.29 ± 0.03 | 0.31 ± 0.05 | 0.19 ± 0.04 | 0.36 ± 0.13 | 1.30 ± 0.01 | 1.79 ± 0.17 | 0.79 ± 0.02 |
| Bmg | 4.37 ± 1.09 | 2.21 ± 0.48 | 2.49 ± 0.67 | 1.73 ± 0.44 | 1.45 ± 0.04 | 1.75 ± 0.33 | 0.91 ± 0.04 |
| Btoy | 0.37 ± 0.02 | 0.31 ± 0.03 | 0.14 ± 0.02 | 0.25 ± 0.05 | 1.40 ± 0.07 | 1.01 ± 0.10 | 0.94 ± 0.04 |
| Pal | 13.97 ± 4.26 | 0.28 ± 0.12 | 0.11 ± 0.04 | 3.54 ± 1.24 | 1.40 ± 0.05 | 1.53 ± 0.50 | 0.86 ± 0.06 |
| Pis | 0.63 ± 0.24 | 0.53 ± 0.27 | 0.28 ± 0.12 | 0.42 ± 0.20 | 1.39 ± 0.02 | 0.94 ± 0.10 | 0.95 ± 0.02 |
| Ppy-fw | 0.48 ± 0.06 | 0.46 ± 0.05 | 0.23 ± 0.04 | 0.37 ± 0.11 | 1.40 ± 0.06 | 1.13 ± 0.11 | 0.90 ± 0.01 |
| Ppy-sw | 0.40 ± 0.06 | 0.43 ± 0.07 | 0.28 ± 0.09 | 0.57 ± 0.21 | 1.35 ± 0.03 | 1.63 ± 0.15 | 1.00 ± 0.07 |
| Intracellular compounds | |||||||
| Baz | 3.05 ± 0.81 | 0.17 ± 0.04 | 0.14 ± 0.03 | 2.18 ± 0.62 | 1.33 ± 0.02 | 0.30 ± 0.02 | 1.14 ± 0.07 |
| Bbd | 17.81 ± 8.20 | 0.26 ± 0.08 | 0.25 ± 0.05 | 5.91 ± 2.67 | 1.38 ± 0.07 | 0.60 ± 0.25 | 1.25 ± 0.11 |
| Bcg | 13.97 ± 4.26 | 0.17 ± 0.10 | 0.11 ± 0.04 | 3.54 ± 1.24 | 1.65 ± 0.07 | 0.13 ± 0.02 | 1.53 ± 0.11 |
| Bcq | 3.59 ± 1.43 | 0.83 ± 0.34 | 0.47 ± 0.14 | 1.79 ± 0.56 | 1.97 ± 0.16 | 1.75 ± 0.95 | 0.73 ± 0.11 |
| Bcr | 0.89 ± 0.24 | 0.28 ± 0.08 | 0.08 ± 0.01 | 0.42 ± 0.07 | 1.61 ± 0.10 | 0.89 ± 0.09 | 0.90 ± 0.11 |
| Blch | 4.71 ± 1.26 | 0.20 ± 0.02 | 0.10 ± 0.01 | 1.05 ± 0.29 | 1.62 ± 0.04 | 0.25 ± 0.05 | 1.39 ± 0.06 |
| Bmg | 1.80 ± 0.62 | 0.35 ± 0.11 | 0.19 ± 0.08 | 1.25 ± 0.31 | 1.65 ± 0.08 | 0.78 ± 0.08 | 0.87 ± 0.08 |
| Btoy | 1.27 ± 0.33 | 0.32 ± 0.10 | 0.20 ± 0.06 | 0.79 ± 0.18 | 1.84 ± 0.14 | 1.08 ± 0.33 | 0.75 ± 0.10 |
| Pal | 3.14 ± 1.61 | 0.12 ± 0.03 | 0.07 ± 0.02 | 1.12 ± 0.45 | 1.30 ± 0.10 | 0.42 ± 0.11 | 1.15 ± 0.19 |
| Pis | 2.77 ± 0.68 | 0.24 ± 0.07 | 0.09 ± 0.02 | 2.17 ± 0.54 | 1.16 ± 0.02 | 0.31 ± 0.01 | 1.07 ± 0.09 |
| Ppy-fw | 5.56 ± 1.58 | 0.18 ± 0.04 | 0.15 ± 0.02 | 1.30 ± 0.38 | 1.22 ± 0.23 | 0.94 ± 0.10 | 0.95 ± 0.02 |
| Ppy-sw | 1.71 ± 0.51 | 0.14 ± 0.03 | 0.06 ± 0.01 | 0.46 ± 0.11 | 1.41 ± 0.08 | 0.49 ± 0.15 | 1.16 ± 0.10 |
| a350 | E2/E3 | S275–295 | S350–400 | SR | |
|---|---|---|---|---|---|
| Extracellular compounds | |||||
| Baz | 52.47 ± 18.84 | 121.4 ± 19.66 | 0.75 ± 0.06 | 0.39 ± 0.07 | 40.89 ± 9.35 |
| Bbd | 182.66 ± 69.46 | 67.53 ± 11.90 | 0.57 ± 0.12 | 0.18 ± 0.06 | 99.54 ± 17.39 |
| Bcg | 34.67 ± 2.25 | 149.58 ± 6.55 | 0.80 ± 0.02 | 0.57 ± 0.02 | 34.43 ± 2.00 |
| Bcq | 51.17 ± 15.78 | 134.29 ± 26.06 | 0.63 ± 0.07 | 0.21 ± 0.05 | 72.33 ± 8.75 |
| Bcr | 24.90 ± 7.92 | 133.6 ± 14.49 | 0.77 ± 0.04 | 0.56 ± 0.05 | 22.45 ± 5.63 |
| Blch | 14.84 ± 3.15 | 202.35 ± 11.83 | 0.98 ± 0.16 | 0.77 ± 0.18 | 16.64 ± 3.57 |
| Bmg | 136.67 ± 85.94 | 138.99 ± 11.04 | 0.76 ± 0.04 | 0.33 ± 0.02 | 53.84 ± 3.43 |
| Btoy | 30.43 ± 5.42 | 145.29 ± 19.07 | 0.73 ± 0.03 | 0.62 ± 0.06 | 27.38 ± 1.30 |
| Pal | 47.77 ± 10.76 | 118.15 ± 13.27 | 0.63 ± 0.05 | 0.72 ± 0.04 | 42.38 ± 6.66 |
| Pis | 40.29 ± 4.97 | 153.77 ± 19.47 | 1.04 ± 0.06 | 0.15 ± 0.02 | 113.6 ± 0.01 |
| Ppy-fw | 153.23 ± 91.66 | 89.33 ± 14.69 | 0.59 ± 0.08 | 0.38 ± 0.08 | 50.88 ± 12.71 |
| Ppy-sw | 189.06 ± 96.47 | 63.91 ± 10.97 | 0.48 ± 0.09 | 0.21 ± 0.06 | 93.33 ± 32.45 |
| Intracellular compounds | |||||
| Baz | 21.27 ± 2.71 | 6.62 ± 0.73 | 0.06 ± 0.01 | 0.01 ± 0.00 | 17.26 ± 1.48 |
| Bbd | 55.17 ± 21.81 | 10.44 ± 2.88 | 0.06 ± 0.01 | 0.03 ± 0.02 | 9.86 ± 1.89 |
| Bcg | 29.43 ± 5.92 | 6.39 ± 0.08 | 0.04 ± 0.00 | 0.01 ± 0.00 | 8.27 ± 0.40 |
| Bcq | 13.17 ± 3.26 | 10.28 ± 1.25 | 0.07 ± 0.01 | 0.01 ± 0.00 | 10.44 ± 1.38 |
| Bcr | 5.40 ± 1.57 | 6.02 ± 0.62 | 0.05 ± 0.01 | 0.01 ± 0.00 | 11.15 ± 1.37 |
| Blch | 13.72 ± 3.53 | 7.40 ± 0.36 | 0.05 ± 0.00 | 0.01 ± 0.00 | 7.67 ± 0.36 |
| Bmg | 3.39 ± 0.53 | 11.70 ± 2.58 | 0.08 ± 0.01 | 0.02 ± 0.01 | 12.01 ± 1.64 |
| Btoy | 8.33 ± 1.82 | 15.50 ± 3.85 | 0.08 ± 0.01 | 0.02 ± 0.00 | 10.20 ± 1.33 |
| Pal | 14.65 ± 5.39 | 8.97 ± 1.09 | 0.07 ± 0.01 | 0.01 ± 0.00 | 13.45 ± 1.80 |
| Pis | 2.77 ± 0.68 | 0.24 ± 0.07 | 0.09 ± 0.02 | 2.17 ± 0.54 | 1.16 ± 0.02 |
| Ppy-fw | 22.76 ± 4.14 | 6.38 ± 0.53 | 0.04 ± 0.00 | 0.01 ± 0.00 | 10.80 ± 0.79 |
| Ppy-sw | 9.97 ± 2.25 | 5.86 ± 0.46 | 0.04 ± 0.00 | 0.02 ± 0.01 | 5.39 ± 0.98 |
| Variables | R2 Partial | F | Pr > F | ʎ Wilks | Var (%) |
|---|---|---|---|---|---|
| Extracellular Fraction | |||||
| C3 | 0.759 | 15.998 | <0.0001 | 0.241 | 65.78 |
| S 350–400 | 0.684 | 10.813 | <0.0001 | 0.076 | 20.80 |
| E2/E3 | 0.641 | 8.759 | <0.0001 | 0.027 | 7.47 |
| C1 | 0.624 | 8.001 | <0.0001 | 0.010 | 2.81 |
| FI | 0.497 | 4.670 | <0.0001 | 0.005 | 1.41 |
| S275–295 | 0.420 | 3.354 | 0.002 | 0.003 | 0.82 |
| C2 | 0.401 | 3.039 | 0.004 | 0.002 | 0.49 |
| HIX | 0.474 | 4.010 | 0.000 | 0.001 | 0.26 |
| C4 | 0.389 | 2.782 | 0.007 | 0.001 | 0.16 |
| SR | 0.522 | 4.674 | <0.0001 | 0.000 | 0.08 |
| BIX | 0.426 | 3.102 | 0.003 | 0.000 | 0.04 |
| Intracellular Fraction | |||||
| FI | 0.500 | 6.445 | <0.0001 | 0.441 | 44.23 |
| BIX | 0.457 | 5.361 | <0.0001 | 0.220 | 22.13 |
| SR | 0.378 | 3.813 | 0.000 | 0.120 | 12.01 |
| a350 | 0.375 | 3.704 | 0.000 | 0.074 | 7.47 |
| S 275–295 | 0.338 | 3.112 | 0.002 | 0.047 | 4.67 |
| C3 | 0.294 | 2.504 | 0.011 | 0.031 | 3.09 |
| HIX | 0.266 | 2.140 | 0.029 | 0.022 | 2.18 |
| S 350–400 | 0.256 | 1.998 | 0.043 | 0.016 | 1.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gullian-Klanian, M.; Gold-Bouchot, G.; Sánchez-Solís, M.J. Characteristics of Chromophoric Dissolved Organic Matter (CDOM) Produced by Heterotrophic Bacteria Isolated from Aquaculture Systems. J. Mar. Sci. Eng. 2022, 10, 672. https://doi.org/10.3390/jmse10050672
Gullian-Klanian M, Gold-Bouchot G, Sánchez-Solís MJ. Characteristics of Chromophoric Dissolved Organic Matter (CDOM) Produced by Heterotrophic Bacteria Isolated from Aquaculture Systems. Journal of Marine Science and Engineering. 2022; 10(5):672. https://doi.org/10.3390/jmse10050672
Chicago/Turabian StyleGullian-Klanian, Mariel, Gerardo Gold-Bouchot, and María José Sánchez-Solís. 2022. "Characteristics of Chromophoric Dissolved Organic Matter (CDOM) Produced by Heterotrophic Bacteria Isolated from Aquaculture Systems" Journal of Marine Science and Engineering 10, no. 5: 672. https://doi.org/10.3390/jmse10050672