Modulating Visuomotor Sequence Learning by Repetitive Transcranial Magnetic Stimulation: What Do We Know So Far?

 ,
,
Abstract
:1. Introduction
1.1. Different Variations of the SRTT
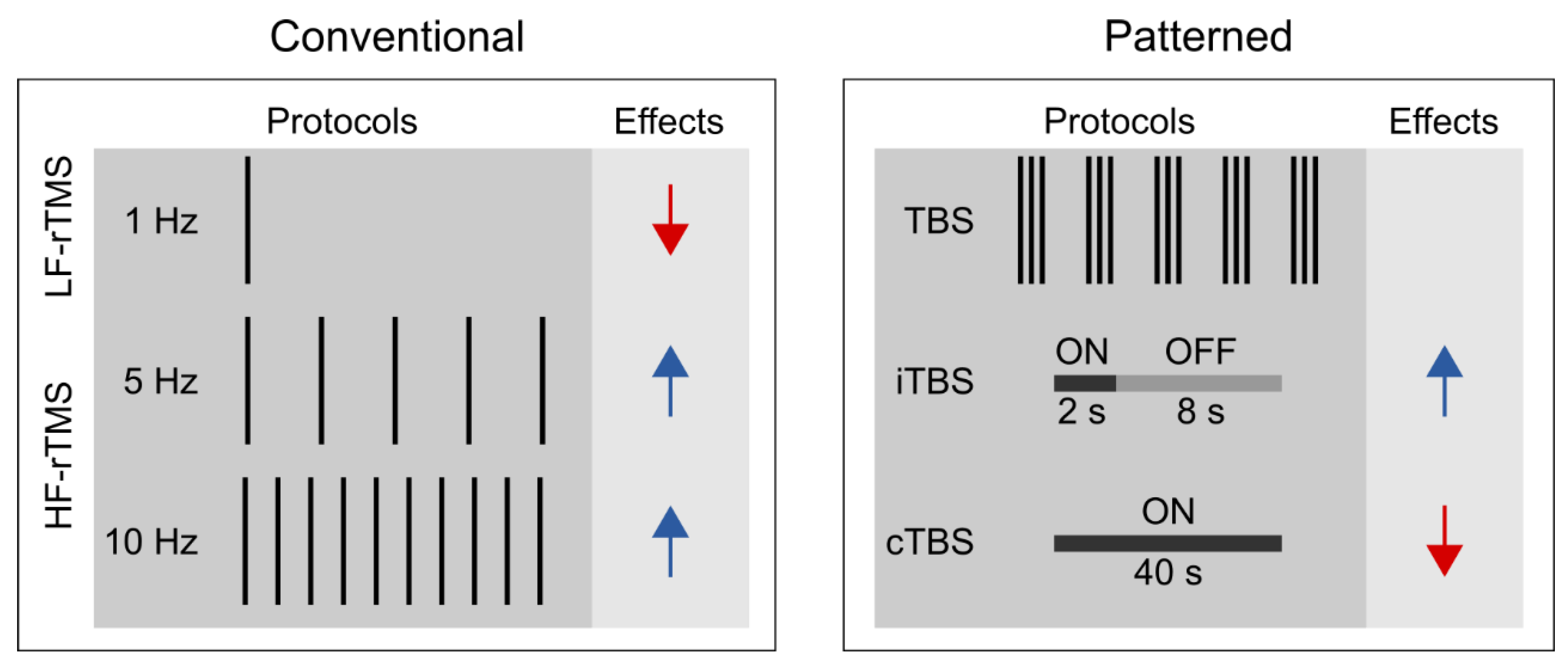
1.2. Different rTMS Protocols
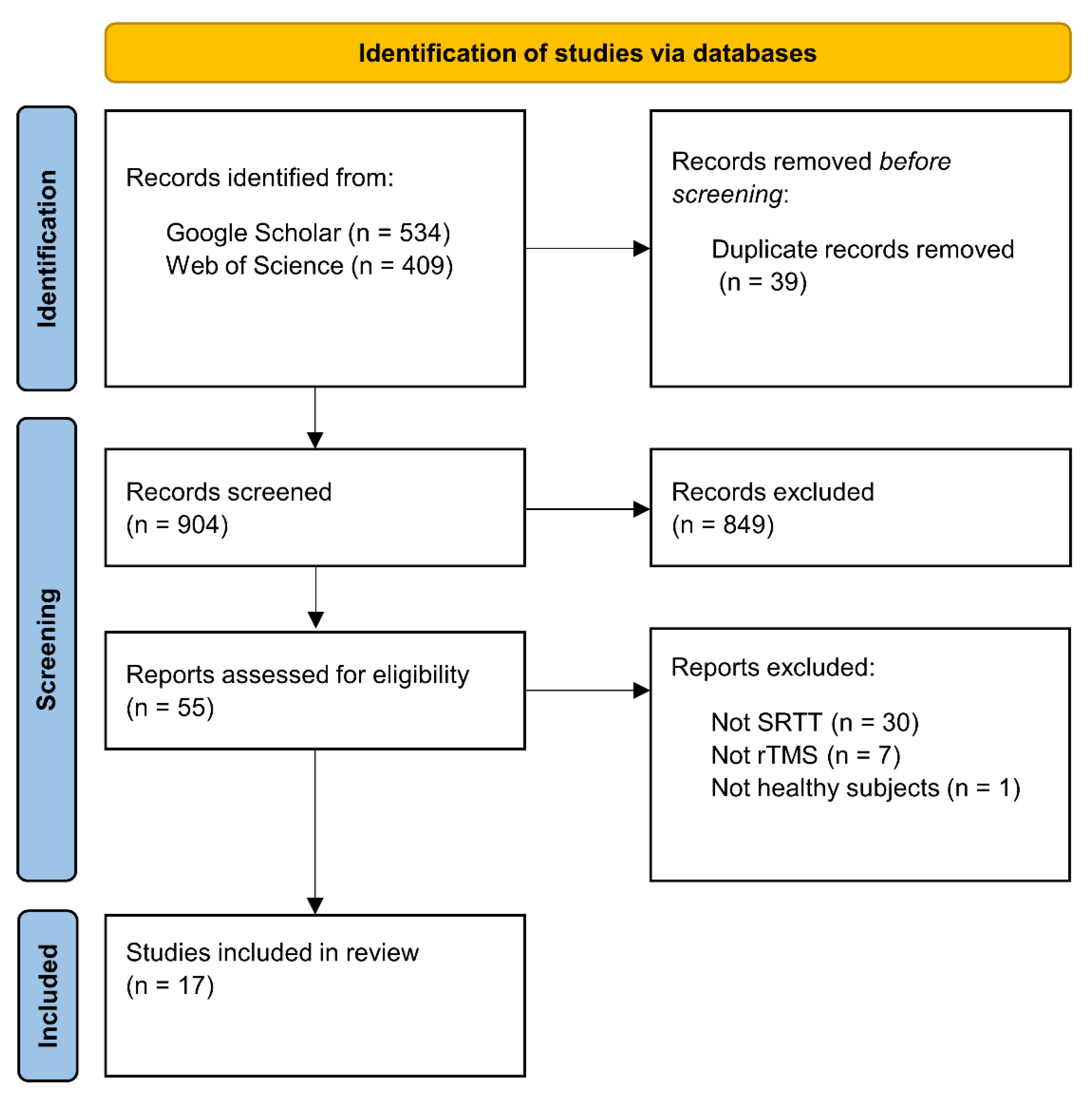
2. Literature Search and Study Selection
3. What Factors Determine the Effect of rTMS on Sequence Learning?
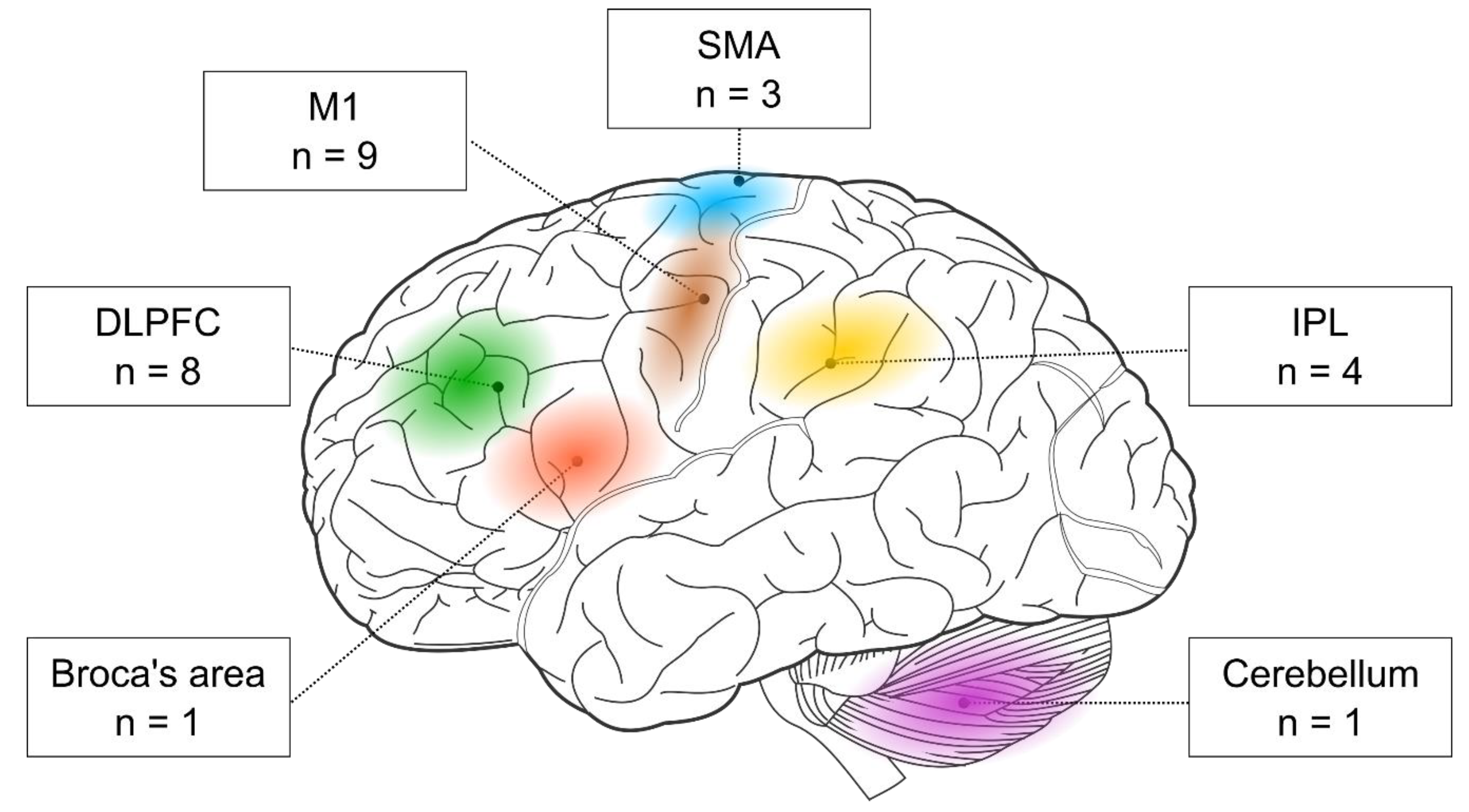
3.1. Stimulated Brain Regions
3.2. ‘Inhibitory’ and ‘Facilitatory’ rTMS Protocols
3.3. Stimulated Hemisphere(s)
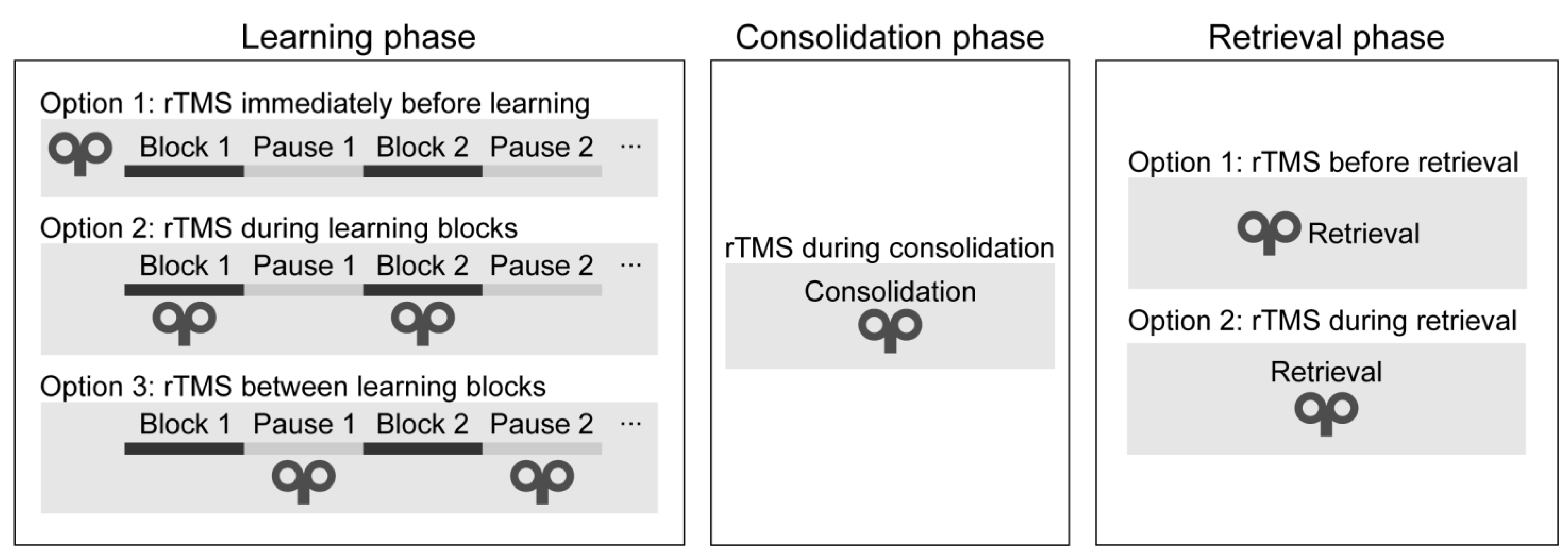
3.4. Timing of the Stimulation
3.5. Type of the SRTT Sequence
3.6. Methodological Features
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aizenstein, Howard J., Meryl A. Butters, Kristi A. Clark, Jennifer L. Figurski, V. Andrew Stenger, Robert D. Nebes, Charles F. Reynolds, and Cameron S. Carter. 2006. Prefrontal and striatal activation in elderly subjects during concurrent implicit and explicit sequence learning. Neurobiology of Aging 27: 741–51. [Google Scholar] [CrossRef]
- Ambrus, Géza Gergely, Teodóra Vékony, Karolina Janacsek, Anna B. C. Trimborn, Gyula Kovács, and Dezso Nemeth. 2020. When less is more: Enhanced statistical learning of non-adjacent dependencies after disruption of bilateral DLPFC. Journal of Memory and Language 114: 104144. [Google Scholar] [CrossRef]
- Andoh, Jamila, and Jean-Luc Martinot. 2008. Interhemispheric compensation: A hypothesis of TMS-induced effects on language-related areas. European Psychiatry 23: 281–88. [Google Scholar] [CrossRef]
- Ashe, James, Ovidiu V. Lungu, Alexandra T. Basford, and Xiaofeng Lu. 2006. Cortical control of motor sequences. Current Opinion in Neurobiology 16: 213–21. [Google Scholar] [CrossRef]
- Baetens, Kris, Mahyar Firouzi, Frank Van Overwalle, and Natacha Deroost. 2020. Involvement of the cerebellum in the serial reaction time task (SRT) (Response to Janacsek et al.). NeuroImage 220: 117114. [Google Scholar] [CrossRef] [PubMed]
- Bagnato, Sergio, Antonio Curra, Nicola Modugno, Francesca Gilio, Angelo Quartarone, Victor Rizzo, Paolo Girlanda, Maurizio Inghilleri, and Alfredo Berardelli. 2005. One-hertz subthreshold rTMS increases the threshold for evoking inhibition in the human motor cortex. Experimental Brain Research 160: 368–74. [Google Scholar] [CrossRef] [PubMed]
- Beldarrain, Marian Gomez, Jordan Gafman, Ibone Ruiz de Velasco, Alvaro Pascual-Leone, and Juan Garcia-Monco. 2002. Prefrontal lesions impair the implicit and explicit learning of sequences on visuomotor tasks. Experimental Brain Research 142: 529–38. [Google Scholar] [CrossRef] [PubMed]
- Beldarrain, Marian Gómez, Jordan Grafman, Alvaro Pascual-Leone, and Juan C. Garcia-Monco. 1999. Procedural learning is impaired in patients with prefrontal lesions. Neurology 52: 1853–53. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, Til Ole, and Gesa Hartwigsen. 2020. Inferring Causality from Noninvasive Brain Stimulation in Cognitive Neuroscience. Journal of Cognitive Neuroscience 33: 195–225. [Google Scholar] [CrossRef]
- Bergstrom, Jennifer C. Romano, James H. Howard, and Darlene V. Howard. 2012. Enhanced Implicit Sequence Learning in College-age Video Game Players and Musicians. Applied Cognitive Psychology 26: 91–96. [Google Scholar] [CrossRef]
- Berlot, Eva, Nicola J. Popp, and Jörn Diedrichsen. 2020. A critical re-evaluation of fMRI signatures of motor sequence learning. eLife 9: 419–30. [Google Scholar] [CrossRef] [PubMed]
- Beynel, Lysianne, John Paul Powers, and Lawrence Gregory Appelbaum. 2020. Effects of repetitive transcranial magnetic stimulation on resting-state connectivity: A systematic review. NeuroImage 211: 116596. [Google Scholar] [CrossRef] [PubMed]
- Boayue, Nya Mehnwolo, Gábor Csifcsák, Per Aslaksen, Zsolt Turi, Andrea Antal, Josephine Groot, Guy E. Hawkins, Birte Forstmann, Alexander Opitz, Axel Thielscher, and et al. 2020. Increasing propensity to mind-wander by transcranial direct current stimulation? A registered report. European Journal of Neuroscience 51: 755–80. [Google Scholar] [CrossRef] [PubMed]
- Boyd, Lara A., and Meghan A. Linsdell. 2009. Excitatory repetitive transcranial magnetic stimulation to left dorsal premotor cortex enhances motor consolidation of new skills. BMC Neuroscience 10: 72. [Google Scholar] [CrossRef] [PubMed]
- Breton, Jocelyn, and Edwin M. Robertson. 2017. Dual enhancement mechanisms for overnight motor memory consolidation. Nature Human Behaviour 1: 0111. [Google Scholar] [CrossRef] [PubMed]
- Buetefisch, Cathrin M., Cortney Howard, Christina Korb, Marc W. Haut, Linda Shuster, Paola Pergami, Cheryl Smith, and Gerald Hobbs. 2015. Conditions for enhancing the encoding of an elementary motor memory by rTMS. Clinical Neurophysiology 126: 581–93. [Google Scholar] [CrossRef]
- Bütefisch, Cathrin M., Vikram Khurana, Leonid Kopylev, and Leonardo G. Cohen. 2004. Enhancing Encoding of a Motor Memory in the Primary Motor Cortex By Cortical Stimulation. Journal of Neurophysiology 91: 2110–16. [Google Scholar] [CrossRef]
- Cao, Na, Yanling Pi, Fanghui Qiu, Yanqiu Wang, Xue Xia, Yu Liu, and Jian Zhang. 2022. Plasticity changes in dorsolateral prefrontal cortex associated with procedural sequence learning are hemisphere-specific. NeuroImage 259: 119406. [Google Scholar] [CrossRef]
- Cao, Na, Yanling Pi, Ke Liu, Haijiang Meng, Yanqiu Wang, Jian Zhang, Yin Wu, and Xiaoying Tan. 2018. Inhibitory and facilitatory connections from dorsolateral prefrontal to primary motor cortex in healthy humans at rest—An rTMS study. Neuroscience Letters 687: 82–87. [Google Scholar] [CrossRef]
- Cardenas, Valerie A., Jyoti V. Bhat, Andrea M. Horwege, Tobin J. Ehrlich, James Lavacot, Daniel H. Mathalon, Gary H. Glover, Brian J. Roach, Bashar W. Badran, Steven D. Forman, and et al. 2022. Anatomical and fMRI-network comparison of multiple DLPFC targeting strategies for repetitive transcranial magnetic stimulation treatment of depression. Brain Stimulation 15: 63–72. [Google Scholar] [CrossRef]
- Cárdenas-Morales, Lizbeth, Dennis A. Nowak, Thomas Kammer, Robert C. Wolf, and Carlos Schönfeldt-Lecuona. 2010. Mechanisms and Applications of Theta-burst rTMS on the Human Motor Cortex. Brain Topography 22: 294–306. [Google Scholar] [CrossRef]
- Cash, Robin F. H., Luca Cocchi, Jinglei Lv, Yumeng Wu, Paul B. Fitzgerald, and Andrew Zalesky. 2021. Personalized connectivity-guided DLPFC-TMS for depression: Advancing computational feasibility, precision and reproducibility. Human Brain Mapping 42: 4155–72. [Google Scholar] [CrossRef] [PubMed]
- Chouinard, Philippe A., Gabriel Leonard, and Tomáš Paus. 2005. Role of the Primary Motor and Dorsal Premotor Cortices in the Anticipation of Forces during Object Lifting. Journal of Neuroscience 25: 2277–84. [Google Scholar] [CrossRef] [PubMed]
- Clark, Gillian M., Michael P. Barham, Anna T. Ware, James M. A. Plumridge, Bernadette O’Sullivan, Kristie Lyons, Tegan Fitzgibbon, Bree Buck, George J. Youssef, Michael T. Ullman, and et al. 2019. Dissociable implicit sequence learning mechanisms revealed by continuous theta-burst stimulation. Behavioral Neuroscience 133: 341–49. [Google Scholar] [CrossRef] [PubMed]
- Clerget, Emeline, William Poncin, Luciano Fadiga, and Etienne Olivier. 2012. Role of Broca’s Area in Implicit Motor Skill Learning: Evidence from Continuous Theta-burst Magnetic Stimulation. Journal of Cognitive Neuroscience 24: 80–92. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Daniel A., Alvaro Pascual-Leone, Daniel Z. Press, and Edwin M. Robertson. 2005. Off-line learning of motor skill memory: A double dissociation of goal and movement. Proceedings of The National Academy of Sciences 102: 18237–241. [Google Scholar] [CrossRef]
- Cohen, Daniel A., and Edwin M. Robertson. 2011. Preventing interference between different memory tasks. Nature Neuroscience 14: 953–55. [Google Scholar] [CrossRef]
- Dahms, Christiane, Stefan Brodoehl, Otto W. Witte, and Carsten M. Klingner. 2020. The importance of different learning stages for motor sequence learning after stroke. Human Brain Mapping 41: 270–86. [Google Scholar] [CrossRef] [PubMed]
- Daselaar, Sander M., Serge A. R. B. Rombouts, Dick J. Veltman, Jeroen G. W. Raaijmakers, and Cees Jonker. 2003. Similar network activated by young and old adults during the acquisition of a motor sequence. Neurobiology of Aging 24: 1013–19. [Google Scholar] [CrossRef]
- Daw, Nathaniel D., Yael Niv, and Peter Dayan. 2005. Uncertainty-based competition between prefrontal and dorsolateral striatal systems for behavioral control. Nature Neuroscience 8: 1704–11. [Google Scholar] [CrossRef]
- Dayan, Eran, Nitzan Censor, Ethan R. Buch, Marco Sandrini, and Leonardo G. Cohen. 2013. Noninvasive brain stimulation: From physiology to network dynamics and back. Nature Neuroscience 16: 838–44. [Google Scholar] [CrossRef]
- de Jesus, Danilo R., Gabriela Pereira de Souza Favalli, Sylco S. Hoppenbrouwers, Mera S. Barr, Robert Chen, Paul B. Fitzgerald, and Zafiris J. Daskalakis. 2014. Determining optimal rTMS parameters through changes in cortical inhibition. Clinical Neurophysiology 125: 755–62. [Google Scholar] [CrossRef] [PubMed]
- De Vries, Meinou H., Andre C. R. Barth, Sandra Maiworm, Stefan Knecht, Pienie Zwitserlood, and Agnes Flöel. 2010. Electrical Stimulation of Broca’s Area Enhances Implicit Learning of an Artificial Grammar. Journal of Cognitive Neuroscience 22: 2427–36. [Google Scholar] [CrossRef] [PubMed]
- De Witte, Sara, Debby Klooster, Josefien Dedoncker, Romain Duprat, Jonathan Remue, and Chris Baeken. 2018. Left prefrontal neuronavigated electrode localization in tDCS: 10–20 EEG system versus MRI-guided neuronavigation. Psychiatry Research: Neuroimaging 274: 1–6. [Google Scholar] [CrossRef]
- Dharani, Krishnagopal. 2015. The Biology of Thought: A Neuronal Mechanism in the Generation of Thought—A New Molecular Model. Amsterdam: Elsevier, pp. 53–74. [Google Scholar] [CrossRef]
- Dirnberger, Georg, Judith Novak, and Christian Nasel. 2013. Perceptual Sequence Learning Is More Severely Impaired than Motor Sequence Learning in Patients with Chronic Cerebellar Stroke. Journal of Cognitive Neuroscience 25: 2207–15. [Google Scholar] [CrossRef]
- Doppelmayr, Michael, Nils Henrik Pixa, and Fabian Steinberg. 2016. Cerebellar, but not Motor or Parietal, High-Density Anodal Transcranial Direct Current Stimulation Facilitates Motor Adaptation. Journal of the International Neuropsychological Society 22: 928–36. [Google Scholar] [CrossRef]
- Doyon, Julien, Danielle Gaudreau, Robert Laforce Jr., Martin Castonguay, Paul J. Bedard, Francois Bedard, and Jean-Pierre Bouchard. 1997. Role of the Striatum, Cerebellum, and Frontal Lobes in the Learning of a Visuomotor Sequence. Brain and Cognition 34: 218–45. [Google Scholar] [CrossRef]
- Duecker, Felix, and Alexander T. Sack. 2015. Rethinking the role of sham TMS. Frontiers in Psychology 6: 210. [Google Scholar] [CrossRef]
- Ferrari, Chiara, Zaira Cattaneo, Viola Oldrati, Letizia Casiraghi, Francesco Castelli, Egidio D’angelo, and Tomaso Vecchi. 2018. TMS Over the Cerebellum Interferes with Short-term Memory of Visual Sequences. Scientific Reports 8: 6722. [Google Scholar] [CrossRef]
- Fitzgerald, Paul B., Sarah Fountain, and Zafiris J. Daskalakis. 2006. A comprehensive review of the effects of rTMS on motor cortical excitability and inhibition. Clinical Neurophysiology 117: 2584–96. [Google Scholar] [CrossRef] [PubMed]
- Foerde, Karin, and Russ A. Poldrack. 2009. Procedural Learning in Humans. Encyclopedia of Neuroscience 2009: 1083–91. [Google Scholar] [CrossRef]
- Galea, Joseph M., Alejandro Vazquez, Neel Pasricha, Jean-Jacques Orban de Xivry, and Pablo Celnik. 2011. Dissociating the Roles of the Cerebellum and Motor Cortex during Adaptive Learning: The Motor Cortex Retains What the Cerebellum Learns. Cerebral Cortex 21: 1761–70. [Google Scholar] [CrossRef]
- Galea, Joseph M., Neil B. Albert, Thomas Ditye, and R. Chris Miall. 2010. Disruption of the Dorsolateral Prefrontal Cortex Facilitates the Consolidation of Procedural Skills. Journal of Cognitive Neuroscience 22: 1158–64. [Google Scholar] [CrossRef] [PubMed]
- Gann, Mareike A., Bradley R. King, Nina Dolfen, Menno P. Veldman, Kimberly L. Chan, Nicolaas A. J. Puts, Richard A. E. Edden, Marco Davare, Stephan P. Swinnen, Dante Mantini, and et al. 2021. Hippocampal and striatal responses during motor learning are modulated by prefrontal cortex stimulation. NeuroImage 237: 118158. [Google Scholar] [CrossRef]
- Gann, Mareike A., Nina Dolfen, Bradley R. King, Edwin M. Robertson, and Geneviève Albouy. 2023. Prefrontal stimulation as a tool to disrupt hippocampal and striatal reactivations underlying fast motor memory consolidation. Brain Stimulation 16: 1336–45. [Google Scholar] [CrossRef] [PubMed]
- Gentner, Reinhard, Katharina Wankerl, Claus Reinsberger, Daniel Zeller, and Joseph Classen. 2008. Depression of Human Corticospinal Excitability Induced by Magnetic Theta-burst Stimulation: Evidence of Rapid Polarity-Reversing Metaplasticity. Cerebral Cortex 18: 2046–53. [Google Scholar] [CrossRef]
- Glinski, Benedikt. 2021. Effects of Different Inhibitory Non-Invasive Brain Stimulation Protocols on Performance in a Motor Sequence Learning Task. Master’s Thesis, University of Twente, Enschede, The Netherlands. [Google Scholar]
- Goldsworthy, Mitchell R., Brenton Hordacre, John C. Rothwell, and Michael C. Ridding. 2021. Effects of rTMS on the brain: Is there value in variability? Cortex 139: 43–59. [Google Scholar] [CrossRef]
- Gomez-Beldarrain, Marian, Juan C. Garcia-Monco, Berta Rubio, and Alvaro Pascual-Leone. 1998. Effect of focal cerebellar lesions on procedural learning in the serial reaction time task. Experimental Brain Research 120: 25–30. [Google Scholar] [CrossRef]
- Grafton, Scott T., Eliot Hazeltine, and Richard B. Ivry. 1998. Abstract and Effector-Specific Representations of Motor Sequences Identified with PET. Journal of Neuroscience 18: 9420–28. [Google Scholar] [CrossRef]
- Grafton, Scott T., Eliot Hazeltine, and Richard B. Ivry. 2002. Motor sequence learning with the nondominant left hand. Experimental Brain Research 146: 369–78. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Masashi, Nagako Murase, Alkomiet Hasan, Michelle Balaratnam, and John C. Rothwell. 2013. The Role of Interneuron Networks in Driving Human Motor Cortical Plasticity. Cerebral Cortex 23: 1593–605. [Google Scholar] [CrossRef] [PubMed]
- Hazeltine, Eliot, Scott T. Grafton, and Richard Ivry. 1997. Attention and stimulus characteristics determine the locus of motor- sequence encoding. A PET study. Brain 120: 123–40. [Google Scholar] [CrossRef] [PubMed]
- Hermsdorf, Franz, Christopher Fricke, Anika Stockert, Joseph Classen, and Jost-Julian Rumpf. 2020. Motor Performance But Neither Motor Learning Nor Motor Consolidation Are Impaired in Chronic Cerebellar Stroke Patients. The Cerebellum 19: 275–85. [Google Scholar] [CrossRef]
- Hikosaka, Okihide, Hiroyuki Nakahara, Miya K. Rand, Katsuyuki Sakai, Xiaofeng Lu, Kae Nakamura, Shigehiro Miyachi, and Kenji Doya. 1999. Parallel neural networks for learning sequential procedures. Trends in Neurosciences 22: 464–71. [Google Scholar] [CrossRef]
- Hikosaka, Okihide, Kuniyoshi Sakai, Satoru Miyauchi, Ryousuke Takino, Yuka Sasaki, and Benno Putz. 1996. Activation of human presupplementary motor area in learning of sequential procedures: A functional MRI study. Journal of Neurophysiology 76: 617–21. [Google Scholar] [CrossRef]
- Honda, Manabu, Marie-Pierre Deiber, Vicente Ibánez, Alvaro Pascual-Leone, Ping Zhuang, and Mark Hallett. 1998. Dynamic cortical involvement in implicit and explicit motor sequence learning. A PET study. Brain 121: 2159–73. [Google Scholar] [CrossRef]
- Hotermans, Christophe, Philippe Peigneux, Alain Maertens de Noordhout, Gustave Moonen, and Pierre Maquet. 2008. Repetitive transcranial magnetic stimulation over the primary motor cortex disrupts early boost but not delayed gains in performance in motor sequence learning. European Journal of Neuroscience 28: 1216–21. [Google Scholar] [CrossRef]
- Howard, Darlene V., James H. Howard, Karin Japikse, Cara DiYanni, Amanda Thompson, and Rachel Somberg. 2004. Implicit Sequence Learning: Effects of Level of Structure, Adult Age, and Extended Practice. Psychology and Aging 19: 79–92. [Google Scholar] [CrossRef]
- Huang, Ying-Zu, Mark J. Edwards, Elisabeth Rounis, Kailash P. Bhatia, and John C. Rothwell. 2005. Theta Burst Stimulation of the Human Motor Cortex. Neuron 45: 201–6. [Google Scholar] [CrossRef]
- Huang, Ying-Zu, Ming-Kue Lu, Andrea Antal, Joseph Classen, Michael Nitsche, Ulf Ziemann, Michael Ridding, Masashi Hamada, Yoshikazu Ugawa, Shapour Jaberzadeh, and et al. 2017. Plasticity induced by non-invasive transcranial brain stimulation: A position paper. Clinical Neurophysiology 128: 2318–29. [Google Scholar] [CrossRef] [PubMed]
- Janacsek, Karolina, and Dezso Nemeth. 2013. Implicit sequence learning and working memory: Correlated or complicated? Cortex 49: 2001–6. [Google Scholar] [CrossRef]
- Janacsek, Karolina, and Dezso Nemeth. 2015. The puzzle is complicated: When should working memory be related to implicit sequence learning, and when should it not? (Response to Martini et al.). Cortex 64: 411–12. [Google Scholar] [CrossRef] [PubMed]
- Janacsek, Karolina, and Dezso Nemeth. 2022. Procedural Memory. In The Cognitive Unconscious. New York: Oxford University Press, p. 22-C2.P104. [Google Scholar] [CrossRef]
- Janacsek, Karolina, József Fiser, and Dezso Nemeth. 2012. The best time to acquire new skills: Age-related differences in implicit sequence learning across the human lifespan. Developmental Science 15: 496–505. [Google Scholar] [CrossRef] [PubMed]
- Janacsek, Karolina, Kyle F. Shattuck, Kaitlyn M. Tagarelli, Jarrad A. G. Lum, Peter E. Turkeltaub, and Michael T. Ullman. 2020. Sequence learning in the human brain: A functional neuroanatomical meta-analysis of serial reaction time studies. NeuroImage 207: 116387. [Google Scholar] [CrossRef]
- Japikse, Karin C., Selam Negash, James H. Howard, and Darlene V. Howard. 2003. Intermanual transfer of procedural learning after extended practice of probabilistic sequences. Experimental Brain Research 148: 38–49. [Google Scholar] [CrossRef]
- Jayaram, Gowri, Byron Tang, Rani Pallegadda, Erin V. L. Vasudevan, Pablo Celnik, Jason Bouffard, Sauro E. Salomoni, Catherine Mercier, Kylie Tucker, Jean-Sébastien Roy, and et al. 2012. Modulating locomotor adaptation with cerebellar stimulation. Journal of Neurophysiology 107: 2950–57. [Google Scholar] [CrossRef]
- Ji, Gong-Jun, Fengqiong Yu, Wei Liao, and Kai Wang. 2017. Dynamic aftereffects in supplementary motor network following inhibitory transcranial magnetic stimulation protocols. NeuroImage 149: 285–94. [Google Scholar] [CrossRef]
- Juhasz, Dora, Dezso Nemeth, and Karolina Janacsek. 2019. Is there more room to improve? The lifespan trajectory of procedural learning and its relationship to the between- and within-group differences in average response times. PLoS ONE 14: e0215116. [Google Scholar] [CrossRef]
- Jung, Patrick, and Ulf Ziemann. 2009. Homeostatic and Nonhomeostatic Modulation of Learning in Human Motor Cortex. Journal of Neuroscience 29: 5597–604. [Google Scholar] [CrossRef]
- Keele, Steven W., Richard Ivry, Ulrich Mayr, Eliot Hazeltine, and Herbert Heuer. 2003. The cognitive and neural architecture of sequence representation. Psychological Review 110: 316–39. [Google Scholar] [CrossRef]
- Keramati, Mehdi, Amir Dezfouli, and Payam Piray. 2011. Speed/Accuracy Trade-Off between the Habitual and the Goal-Directed Processes. PLoS Computational Biology 7: e1002055. [Google Scholar] [CrossRef]
- Khatibi, Ali, Shahabeddin Vahdat, Ovidiu Lungu, Jurgen Finsterbusch, Christian Büchel, Julien Cohen-Adad, Veronique Marchand-Pauvert, and Julien Doyon. 2022. Brain-spinal cord interaction in long-term motor sequence learning in human: An fMRI study. NeuroImage 253: 119111. [Google Scholar] [CrossRef] [PubMed]
- Kim, Yun-Hee, Ji-Won Park, Myoung-Hwan Ko, Sung Ho Jang, and Peter K. W. Lee. 2004. Facilitative effect of high frequency subthreshold repetitive transcranial magnetic stimulation on complex sequential motor learning in humans. Neuroscience Letters 367: 181–85. [Google Scholar] [CrossRef]
- Klomjai, Wanalee, Rose Katz, and Alexandra Lackmy-Vallée. 2015. Basic principles of transcranial magnetic stimulation (TMS) and repetitive TMS (rTMS). Annals of Physical and Rehabilitation Medicine 58: 208–13. [Google Scholar] [CrossRef]
- Kóbor, Andrea, Karolina Janacsek, Petra Hermann, Zzófia Zavecz, Virág Varga, Valéria Csépe, Zoltán Vidnyánszki, Gyula Kovacs, and Dezso Nemeth. 2022. Finding pattern in the noise: Persistent implicit statistical knowledge impacts the processing of unpredictable stimuli. PsyArXiv. [Google Scholar] [CrossRef]
- Lang, Nicolas, Jochen Harms, Thomas Weyh, Roger N. Lemon, Walter Paulus, John C. Rothwell, and Hartwig R. Siebner. 2006. Stimulus intensity and coil characteristics influence the efficacy of rTMS to suppress cortical excitability. Clinical Neurophysiology 117: 2292–301. [Google Scholar] [CrossRef]
- Lee, Sang Wan, Shinsuke Shimojo, and John P. O’doherty. 2014. Neural Computations Underlying Arbitration between Model-Based and Model-free Learning. Neuron 81: 687–99. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, Matthew D. 2000. Intuition: A social cognitive neuroscience approach. Psychological Bulletin 126: 109–37. [Google Scholar] [CrossRef]
- Lum, Jarrad A. G., Andrea Mills, James M. A. Plumridge, Nicole P. Sloan, Gillian M. Clark, Martina Hedenius, and Peter G. Enticott. 2018. Transcranial direct current stimulation enhances retention of a second (but not first) order conditional visuo-motor sequence. Brain and Cognition 127: 34–41. [Google Scholar] [CrossRef] [PubMed]
- Lynch, Charles J., Immanuel G. Elbau, Tommy H. Ng, Danielle Wolk, Shasha Zhu, Aliza Ayaz, Jonathan D. Power, Benjamin Zebley, Faith M. Gunning, and Conor Liston. 2022. Automated optimization of TMS coil placement for personalized functional network engagement. Neuron 110: 3263–3277.e4. [Google Scholar] [CrossRef]
- Maeda, Fumiko, Julian P. Keenan, Jose M. Tormos, Helge Topka, and Alvaro Pascual-Leone. 2000. Interindividual variability of the modulatory effects of repetitive transcranial magnetic stimulation on cortical excitability. Experimental Brain Research 133: 425–30. [Google Scholar] [CrossRef]
- McCalley, Daniel M., Daniel H. Lench, Jade D. Doolittle, Julia P. Imperatore, Michaela Hoffman, and Colleen A. Hanlon. 2021. Determining the optimal pulse number for theta burst induced change in cortical excitability. Scientific Reports 11: 8726. [Google Scholar] [CrossRef]
- Meehan, Sean K., Jeanie R. Zabukovec, Elizabeth Dao, Katharine L. Cheung, Meghan A. Linsdell, and Lara A. Boyd. 2013. One hertz repetitive transcranial magnetic stimulation over dorsal premotor cortex enhances offline motor memory consolidation for sequence-specific implicit learning. European Journal of Neuroscience 38: 3071–79. [Google Scholar] [CrossRef] [PubMed]
- Miller, Earl K., and Jonathan D. Cohen. 2001. An Integrative Theory of Prefrontal Cortex Function. Annual Review of Neuroscience 24: 167–202. [Google Scholar] [CrossRef] [PubMed]
- Muellbacher, Wolf, Ulf Ziemann, Joerg Wissel, Nguyet Dang, Markus Kofler, Stefano Facchini, Babak Boroojerdi, Werner Poewe, and Mark Hallett. 2002. Early consolidation in human primary motor cortex. Nature 415: 640–44. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, Dezso, Emese Hallgató, Karolina Janacsek, Timea Sándor, and Zsuzsa Londe. 2009. Perceptual and motor factors of implicit skill learning. NeuroReport 20: 1654–58. [Google Scholar] [CrossRef]
- Nemeth, Dezso, Karolina Janacsek, Bertalan Polner, and Zoltan Ambrus Kovacs. 2013. Boosting Human Learning by Hypnosis. Cerebral Cortex 23: 801–5. [Google Scholar] [CrossRef]
- Nemeth, Dezso, Karolina Janacsek, Gabor Csifcsak, Gabor Szvoboda, James H. Howard, and Darlene V. Howard. 2011. Interference between Sentence Processing and Probabilistic Implicit Sequence Learning. PLoS ONE 6: e17577. [Google Scholar] [CrossRef]
- Nissen, Mary Jo, and Peter Bullemer. 1987. Attentional requirements of learning: Evidence from performance measures. Cognitive Psychology 19: 1–32. [Google Scholar] [CrossRef]
- Ozdemir, Recep A., Pierre Boucher, Peter J. Fried, Davide Momi, Ali Jannati, Alvaro Pascual-Leone, Emiliano Santarnecchi, and Mouhsin M. Shafi. 2021. Reproducibility of cortical response modulation induced by intermittent and continuous theta-burst stimulation of the human motor cortex. Brain Stimulation 14: 949–64. [Google Scholar] [CrossRef]
- Page, Matthew J., Joanne E. McKenzie, Patrick M. Bossuyt, Isabelle Boutron, Tammy C. Hoffmann, Cynthia D. Mulrow, Larissa Shamseer, Jennifer M. Tetzlaff, Elie A. Akl, Sue E. Brennan, and et al. 2021. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 372: n71. [Google Scholar] [CrossRef] [PubMed]
- Park, Jungtak, Karolina Janacsek, Dezso Nemeth, and Hyeon-Ae Jeon. 2022. Reduced functional connectivity supports statistical learning of temporally distributed regularities. NeuroImage 260: 119459. [Google Scholar] [CrossRef]
- Pascual-Leone, Alvaro, John R. Gates, and Anil Dhuna. 1991. Induction of speech arrest and counting errors with rapid-rate transcranial magnetic stimulation. Neurology 41: 697–702. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Leone, Alvaro, Eric M. Wassermann, Jordan Grafman, and Mark Hallett. 1996. The role of the dorsolateral prefrontal cortex in implicit procedural learning. Experimental Brain Research 107: 479–85. [Google Scholar] [CrossRef]
- Pedraza, Felipe, Bence C. Farkas, Teodóra Vékony, Frederic Haesebaert, Romane Phelipon, Imola Mihalecz, Karolina Janacsek, Royce Anders, Barbara Tillmann, Gaën Plancher, and et al. 2023. Evidence for a competitive relationship between executive functions and statistical learning. BioRxiv. [Google Scholar] [CrossRef]
- Peigneux, Philippe, Steven Laureys, Xavier Delbeuck, and Pierre Maquet. 2001. Sleeping brain, learning brain. The role of sleep for memory systems. NeuroReport 12: A111–24. [Google Scholar] [CrossRef]
- Pell, Gaby S., Yiftach Roth, and Abraham Zangen. 2011. Modulation of cortical excitability induced by repetitive transcranial magnetic stimulation: Influence of timing and geometrical parameters and underlying mechanisms. Progress in Neurobiology 93: 59–98. [Google Scholar] [CrossRef]
- Perez, Monica A., Satoshi Tanaka, Steven P. Wise, Daniel T. Willingham, and Leonardo G. Cohen. 2008. Time-Specific Contribution of the Supplementary Motor Area to Intermanual Transfer of Procedural Knowledge. Journal of Neuroscience 28: 9664–69. [Google Scholar] [CrossRef]
- Polanía, Rafael, Michael A. Nitsche, and Christian C. Ruff. 2018. Studying and modifying brain function with non-invasive brain stimulation. Nature Neuroscience 21: 174–87. [Google Scholar] [CrossRef]
- Poldrack, Russell A., Fred W. Sabb, Karin Foerde, Sabrina M. Tom, Robert F. Asarnow, Susan Y. Bookheimer, and Barbara J. Knowlton. 2005. The Neural Correlates of Motor Skill Automaticity. Journal of Neuroscience 25: 5356–64. [Google Scholar] [CrossRef]
- Prashad, Shikha, Yue Du, and Jane E. Clark. 2021. Sequence Structure Has a Differential Effect on Underlying Motor Learning Processes. Journal of Motor Learning and Development 9: 38–57. [Google Scholar] [CrossRef]
- Reber, Arthur S. 1967. Implicit learning of artificial grammars. Journal of Verbal Learning and Verbal Behavior 6: 855–63. [Google Scholar] [CrossRef]
- Reber, Arthur S. 1989. Implicit learning and tacit knowledge. Journal of Experimental Psychology: General 118: 219–35. [Google Scholar] [CrossRef]
- Robertson, Edwin M. 2007. The Serial Reaction Time Task: Implicit Motor Skill Learning? Journal of Neuroscience 27: 10073–75. [Google Scholar] [CrossRef] [PubMed]
- Robertson, Edwin M., Alvaro Pascual-Leone, and R. Chris Miall. 2004. Current concepts in procedural consolidation. Nature Reviews Neuroscience 5: 576–82. [Google Scholar] [CrossRef]
- Robertson, Edwin M., Daniel Z. Press, and Alvaro Pascual-Leone. 2005. Off-Line Learning and the Primary Motor Cortex. Journal of Neuroscience 25: 6372–78. [Google Scholar] [CrossRef]
- Robertson, Edwin M., Jose M. Tormos, Fumiko Maeda, and Alvaro Pascual-Leone. 2001. The Role of the Dorsolateral Prefrontal Cortex during Sequence Learning is Specific for Spatial Information. Cerebral Cortex 11: 628–35. [Google Scholar] [CrossRef]
- Rosenthal, Clive R., Emma E. Roche-Kelly, Masud Husain, and Christopher Kennard. 2009. Response-Dependent Contributions of Human Primary Motor Cortex and Angular Gyrus to Manual and Perceptual Sequence Learning. Journal of Neuroscience 29: 15115–25. [Google Scholar] [CrossRef]
- Rothkegel, Holger, Martin Sommer, and Walter Paulus. 2010. Breaks during 5Hz rTMS are essential for facilitatory after effects. Clinical Neurophysiology 121: 426–30. [Google Scholar] [CrossRef]
- Ruitenberg, Marit F. L., Willem B. Verwey, Dennis J. L. G. Schutter, and Elger L. Abrahamse. 2014. Cognitive and neural foundations of discrete sequence skill: A TMS study. Neuropsychologia 56: 229–38. [Google Scholar] [CrossRef]
- Sack, Alexander T., Joan A. Camprodon, Alvaro Pascual-Leone, and Rainer Goebel. 2005. The Dynamics of Interhemispheric Compensatory Processes in Mental Imagery. Science 308: 702–4. [Google Scholar] [CrossRef] [PubMed]
- Sack, Alexander T., Roi Cohen Kadosh, Teresa Schuhmann, Michelle Moerel, Vincent Walsh, and Rainer Goebel. 2009. Optimizing Functional Accuracy of TMS in Cognitive Studies: A Comparison of Methods. Journal of Cognitive Neuroscience 21: 207–21. [Google Scholar] [CrossRef]
- Seidler, Raphael D., Arnie Purushotham, Seong-Gi Kim, Kaamil Ugurbil, Daniel Willingham, and James Ashe. 2002. Cerebellum Activation Associated with Performance Change but Not Motor Learning. Science 296: 2043–46. [Google Scholar] [CrossRef]
- Seidler, Raphael D., Arnie Purushotham, Seong-Gi Kim, Kaamil Ugurbil, Daniel Willingham, and James Ashe. 2005. Neural correlates of encoding and expression in implicit sequence learning. Experimental Brain Research 165: 114–24. [Google Scholar] [CrossRef]
- Shanks, David R. 2005. Implicit learning. In Handbook of Cognition. Thousand Oaks: SAGE Publications Ltd. [Google Scholar] [CrossRef]
- Shimizu, Takahiro, Ritsuko Hanajima, Yuichiro Shirota, Ryosuke Tsutsumi, Nobuyuki Tanaka, Yasuo Terao, Masashi Hamada, and Yoshikazu Ugawa. 2020. Plasticity induction in the pre-supplementary motor area (pre-SMA) and SMA-proper differentially affects visuomotor sequence learning. Brain Stimulation 13: 229–38. [Google Scholar] [CrossRef]
- Shin, Jacqueline C., and Richard B. Ivry. 2003. Spatial and Temporal Sequence Learning in Patients with Parkinson’s Disease or Cerebellar Lesions. Journal of Cognitive Neuroscience 15: 1232–43. [Google Scholar] [CrossRef]
- Simeoni, Sara, Ricci Hannah, Daisuke Sato, Michiyuki Kawakami, John Rothwell, and Gian Luigi Gigli. 2016. Effects of Quadripulse Stimulation on Human Motor Cortex Excitability: A Replication Study. Brain Stimulation 9: 148–50. [Google Scholar] [CrossRef]
- Smalle, Eleonore H. M., and Riikka Möttönen. 2023. Cognitive Development as a Piece of the Language Learning Puzzle. Cognitive Science 47: e13296. [Google Scholar] [CrossRef] [PubMed]
- Smalle, Eleonore H. M., Muriel Panouilleres, Arnaud Szmalec, and Riikka Möttönen. 2017. Language learning in the adult brain: Disrupting the dorsolateral prefrontal cortex facilitates word-form learning. Scientific Reports 7: 13966. [Google Scholar] [CrossRef] [PubMed]
- Smalle, Eleonore H. M., Tatsuya Daikoku, Arnaud Szmalec, Wouter Duyck, and Riikka Möttönen. 2022. Unlocking adults’ implicit statistical learning by cognitive depletion. Proceedings of the National Academy of Sciences of the United States of America 119: e2026011119. [Google Scholar] [CrossRef]
- Smittenaar, Peter, Thomas H. B. FitzGerald, Vincenzo Romei, Nicholas D. Wright, and Raymond J. Dolan. 2013. Disruption of Dorsolateral Prefrontal Cortex Decreases Model-Based in Favor of Model-free Control in Humans. Neuron 80: 914–19. [Google Scholar] [CrossRef]
- Song, Sunbin, James H. Howard, and Darlene V. Howard. 2007. Sleep Does Not Benefit Probabilistic Motor Sequence Learning. Journal of Neuroscience 27: 12475–83. [Google Scholar] [CrossRef]
- Song, Sunbin, James H. Howard, and Darlene V. Howard. 2008. Perceptual sequence learning in a serial reaction time task. Experimental Brain Research 189: 145–58. [Google Scholar] [CrossRef]
- Steel, Adam, Sunbin Song, Devin Bageac, Kristine M. Knutson, Aysha Keisler, Ziad S. Saad, Stephen J. Gotts, Eric M. Wassermann, and Leonora Wilkinson. 2016. Shifts in connectivity during procedural learning after motor cortex stimulation: A combined transcranial magnetic stimulation/functional magnetic resonance imaging study. Cortex 74: 134–48. [Google Scholar] [CrossRef]
- Thut, Gregor, and Alvaro Pascual-Leone. 2010. A Review of Combined TMS-EEG Studies to Characterize Lasting Effects of Repetitive TMS and Assess Their Usefulness in Cognitive and Clinical Neuroscience. Brain Topography 22: 219–32. [Google Scholar] [CrossRef]
- Tiksnadi, Amanda, Takenobu Murakami, Winnugroho Wiratman, Hideyuki Matsumoto, and Yoshikazu Ugawa. 2020. Direct comparison of efficacy of the motor cortical plasticity induction and the interindividual variability between TBS and QPS. Brain Stimulation 13: 1824–33. [Google Scholar] [CrossRef] [PubMed]
- Torriero, Sara, Massimiliano Oliveri, Giacomo Koch, Carlo Caltagirone, and Laura Petrosini. 2004. Interference of Left and Right Cerebellar rTMS with Procedural Learning. Journal of Cognitive Neuroscience 16: 1605–11. [Google Scholar] [CrossRef] [PubMed]
- Tóth-Fáber, Eszter, Dezso Nemeth, and Karolina Janacsek. 2023. Lifespan developmental invariance in memory consolidation: Evidence from procedural memory. PNAS Nexus 2: pgad037. [Google Scholar] [CrossRef] [PubMed]
- Tunovic, Sanjin, Daniel Z. Press, and Edwin M. Robertson. 2014. A Physiological Signal That Prevents Motor Skill Improvements during Consolidation. Journal of Neuroscience 34: 5302–10. [Google Scholar] [CrossRef]
- Turi, Zsolt, Maximilian Lenz, Walter Paulus, Matthias Mittner, and Andreas Vlachos. 2021. Selecting stimulation intensity in repetitive transcranial magnetic stimulation studies: A systematic review between 1991 and 2020. European Journal of Neuroscience 53: 3404–15. [Google Scholar] [CrossRef]
- Turrigiano, Gina. 2012. Homeostatic Synaptic Plasticity: Local and Global Mechanisms for Stabilizing Neuronal Function. Cold Spring Harbor Perspectives in Biology 4: a005736. [Google Scholar] [CrossRef]
- Uddén, Julia, Martin Ingvar, Peter Hagoort, and Karl Magnus Petersson. 2017. Broca’s region: A causal role in implicit processing of grammars with crossed non-adjacent dependencies. Cognition 164: 188–98. [Google Scholar] [CrossRef]
- van der Graaf, Ferdinand H. C. E., R. Paul Maguire, Klaus L. Leenders, and Bauke M. de Jong. 2006. Cerebral activation related to implicit sequence learning in a Double Serial Reaction Time task. Brain Research 1081: 179–90. [Google Scholar] [CrossRef] [PubMed]
- Veldman, Menno P., Natasha M. Maurits, M. A. M. Nijland, Nicholas E. Wolters, John C. Mizelle, and Tibor Hortobágyi. 2018. Spectral and temporal electroencephalography measures reveal distinct neural networks for the acquisition, consolidation, and interlimb transfer of motor skills in healthy young adults. Clinical Neurophysiology 129: 419–30. [Google Scholar] [CrossRef] [PubMed]
- Verwey, Willem B., Benedikt Glinski, Min-Fang Kuo, Mohammad Ali Salehinejad, and Michael A. Nitsche. 2022. Consolidation of motor sequence learning eliminates susceptibility of SMAproper to TMS: A combined rTMS and cTBS study. Experimental Brain Research 240: 1743–55. [Google Scholar] [CrossRef]
- Verwey, Willem B., Robin Lammens, and Jack van Honk. 2002. On the role of the SMA in the discrete sequence production task: A TMS study. Neuropsychologia 40: 1268–76. [Google Scholar] [CrossRef]
- Walker, Matthew P., and Robert Stickgold. 2005. It’s Practice, with Sleep, that Makes Perfect: Implications of Sleep-Dependent Learning and Plasticity for Skill Performance. Clinics in Sports Medicine 24: 301–17. [Google Scholar] [CrossRef]
- Wilkinson, Leonora, Adam Steel, Eric Mooshagian, Trelawny Zimmermann, Aysha Keisler, Jeffrey D. Lewis, and Eric M. Wassermann. 2015. Online feedback enhances early consolidation of motor sequence learning and reverses recall deficit from transcranial stimulation of motor cortex. Cortex 71: 134–47. [Google Scholar] [CrossRef]
- Wilkinson, Leonora, James T. Teo, Ignacio Obeso, John C. Rothwell, and Marjan Jahanshahi. 2010. The Contribution of Primary Motor Cortex is Essential for Probabilistic Implicit Sequence Learning: Evidence from Theta Burst Magnetic Stimulation. Journal of Cognitive Neuroscience 22: 427–36. [Google Scholar] [CrossRef]
- Willingham, Daniel B., Joanna Salidis, John D. E. Gabrieli, Brian Greeley, Rachael D. Seidler, Heidi M. Schambra, Mitsunari Abe, David A. Luckenbaugh, Janine Reis, John W. Krakauer, and et al. 2002. Direct Comparison of Neural Systems Mediating Conscious and Unconscious Skill Learning. Journal of Neurophysiology 88: 1451–60. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, Atsushi, Spencer A. Arbuckle, and Jörn Diedrichsen. 2018. The Role of Human Primary Motor Cortex in the Production of Skilled Finger Sequences. Journal of Neuroscience 38: 1430–42. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Peng, and Naftali Raz. 2014. Prefrontal cortex and executive functions in healthy adults: A meta-analysis of structural neuroimaging studies. Neuroscience & Biobehavioral Reviews 42: 180–92. [Google Scholar] [CrossRef]
- Zolnai, Tamás, Dominika Réka Dávid, Orsolya Pesthy, Marton Nemeth, Mariann Kiss, Márton Nagy, and Dezso Nemeth. 2022. Measuring statistical learning by eye-tracking. Experimental Results 3: e10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Area | Authors and Year | rTMS Protocol | Hemisphere | Timing of Stimulation | Type of Sequence | Outcome |
|---|---|---|---|---|---|---|
| M1 | ||||||
| (Robertson et al. 2005) | 1 Hz rTMS | Left only | After learning | 12-item implicit deterministic | Blocked offline improvements over the day, but not overnight | |
| (Breton and Robertson 2017) | 1 Hz rTMS | Left only | After learning | 12-item implicit/explicit deterministic | Blocked offline improvements in explicit, but not in implicit task | |
| (Tunovic et al. 2014) (Experiment 3) | cTBS | Right only | After learning | 12-item explicit deterministic | Offline improvements after cTBS | |
| (Cohen and Robertson 2011) (Experiment 2) | 1 Hz rTMS | Right only | After learning | 12-item implicit deterministic | Increased learning after 12 h consolidation by preventing interference with a declarative task | |
| (Wilkinson et al. 2010) | cTBS, iTBS | Left only | Before learning | 12-item implicit probabilistic | Learning was prevented by cTBS | |
| (Wilkinson et al. 2015) | cTBS | Left only | Before learning | 12-item implicit probabilistic | Decreased initial sequence learning and recall | |
| (Steel et al. 2016) | cTBS | Left only | Before learning | 12-item implicit probabilistic | Learning was disrupted | |
| (Rosenthal et al. 2009) (Experiment 1, 2) | cTBS | Contralateral to dominant hand | Before learning | 12-item implicit probabilistic | Learning was disrupted in manual, but not in perceptual task | |
| (Clark et al. 2019) | cTBS | Left only | Before learning | 12-item implicit deterministic | Decreased learning in simple, but not in a more complex sequence | |
| DLPFC | ||||||
| (Pascual-Leone et al. 1996) | 5 Hz rTMS | Left or right in separate conditions | During learning | 12-item implicit deterministic | Learning was disrupted | |
| (Robertson et al. 2001) | 1 Hz rTMS | Contralateral to dominant hand | Before learning | 10-item implicit deterministic | Learning was prevented in spatial, but not in color cue guided task | |
| (Wilkinson et al. 2010) | cTBS | Left only | Before learning | 12-item implicit probabilistic | No effect on learning | |
| (Gann et al. 2021) | cTBS, iTBS | Left only | Before learning | 8-item explicit deterministic | No effect on learning | |
| (Galea et al. 2010) | cTBS | Left or right in separate groups | After learning | 12-item explicit deterministic | Improved learning after 8 h consolidation | |
| (Tunovic et al. 2014) (Experiment 2) | cTBS | Right only | After learning | 12-item explicit deterministic | Offline improvements after cTBS | |
| (Cohen and Robertson 2011) (Experiment 2) | 1 Hz rTMS | Right only | After learning | 12-item implicit deterministic | Decreased learning after 12 h consolidation by failing to prevent interference with a declarative task | |
| (Ambrus et al. 2020) | 1 Hz rTMS | Bilaterally | Between learning blocks | 8-item implicit probabilistic | Improved learning after 24 h consolidation | |
| Broca’s area | ||||||
| (Clerget et al. 2012) | cTBS | Left only | Before learning | 20-item implicit deterministic | Learning was prevented | |
| SMA | ||||||
| (Pascual-Leone et al. 1996) | 5 Hz rTMS | Not applicable | During learning | 12-item implicit deterministic | No effect on learning | |
| (Wilkinson et al. 2010) | cTBS | Not applicable | Before learning | 12-item implicit probabilistic | No effect on learning | |
| (Perez et al. 2008) | 1 Hz rTMS | Not applicable | During learning | 12-item implicit deterministic | Blocked intermanual transfer of the skill | |
| IPL | ||||||
| (Robertson et al. 2001) | 1 Hz rTMS | Contralateral to the dominant hand | Before learning | 10-item implicit deterministic | No effect on learning | |
| (Rosenthal et al. 2009) (Experiment 1, 2) | cTBS | Right only | Before learning | 12-item implicit probabilistic | Learning was disrupted in perceptual, but not in manual task | |
| (Breton and Robertson 2017) | 1 Hz rTMS | Left only | After learning | 12-item implicit, explicit deterministic | Blocked offline improvements in implicit, but not in explicit task | |
| (Clark et al. 2019) | cTBS | Left only | Before learning | 12-item implicit, deterministic | No effect on learning | |
| Cerebellum | ||||||
| (Torriero et al. 2004) | 1 Hz rTMS | Left or right in separate groups | Before learning | 12-item implicit, deterministic | Learning was disrupted |
| Design | Study | Type of Control Condition | N | N/Group |
|---|---|---|---|---|
| Within-subjects | ||||
| (Pascual-Leone et al. 1996) | Absence of stimulation | 7 | ||
| (Robertson et al. 2001) | Active control (parietal cortex) | 6 | ||
| (Gann et al. 2021) | No control | 19 | ||
| (Steel et al. 2016) | Sham stimulation with placebo coil | 22 | ||
| Between-subjects | ||||
| (Robertson et al. 2005) | Sham stimulation with placebo coil, time, and site control | 36 | 6 | |
| (Breton and Robertson 2017) | Sham stimulation with placebo coil | 67 | 12–16 | |
| (Tunovic et al. 2014) (Experiment 2) | Intermediate TBS, localization control | 24 | 12 | |
| (Tunovic et al. 2014) (Experiment 3) | Intermediate TBS | 24 | 12 | |
| (Cohen and Robertson 2011) (Experiment 2) | Sham stimulation over vertex with placebo coil | 40 | 10 | |
| (Wilkinson et al. 2010) | Sham stimulation with 90° rotation of the coil | 40 | 8 | |
| (Rosenthal et al. 2009) (Experiment 1, 2) | Sham stimulation over vertex with placebo coil | 24, 24 | 8, 8 | |
| (Galea et al. 2010) | Active control (occipital cortex) | 30 | 10 | |
| (Ambrus et al. 2020) | Sham stimulation with 90° rotation of the coil | 31 | 15 and 16 | |
| (Clerget et al. 2012) | Active control (vertex) | 17 | 8 and 9 | |
| (Perez et al. 2008) | Sham stimulation with a second coil discharged in the air | 33 | 11 | |
| (Torriero et al. 2004) | Absence of stimulation | 36 | 5–7 | |
| Mixed design | ||||
| (Wilkinson et al. 2015) | Sham stimulation with placebo coil | 40 | 20 | |
| (Clark et al. 2019) | Sham stimulation with placebo coil | 48 | 16 |
| Study | Localization Technique | Intensity | Total Number of Pulses | Data Type of the Result |
|---|---|---|---|---|
| (Pascual-Leone et al. 1996) |
| 115% of MT | not available | RT |
| (Robertson et al. 2001) |
| 115% of MT | 900 | RT |
| (Robertson et al. 2005) |
| 90% of MT | 600 | Learning score based on RT |
| (Breton and Robertson 2017) |
* Note that the origin of the exact coordinate cannot be traced with high certainty (one of the two cited articles used different coordinates and the other did not involve neuroimaging) | 90% of MT | 600 | Learning improvement based on RT |
| (Tunovic et al. 2014) (Experiment 2 and 3) |
* Note that all of the cited prior studies in this reference reported coordinates at the left hemisphere (x = −40, y = 32, z = 30), and not at the right hemisphere | 80% of AMT | 600 | Learning score based on RT |
| (Cohen and Robertson 2011) (Experiment 2) |
* Note that all of the cited prior studies in this reference reported coordinates at the left hemisphere (x = −40, y = 32, z = 30), and not at the right hemisphere | 90% of MT | 600 | Learning score based on RT |
| (Rosenthal et al. 2009) (Experiment 1, 2) |
| 70% of RMT | 300 | Learning score based on RT |
| (Wilkinson et al. 2010) |
| 80% of AMT | 600 | Learning score based on RT |
| (Wilkinson et al. 2015) |
| 80% of AMT | 600 | Learning score based on RT and accuracy |
| (Steel et al. 2016) |
| 80% of AMT | 600 | Learning score based on RT |
| (Torriero et al. 2004) |
| 90% of MT | 600 | RT |
| (Perez et al. 2008) |
| 80% of RMT | 1200 | RT |
| (Galea et al. 2010) |
| 80% of AMT | 600 | Learning improvement based on RT |
| (Clerget et al. 2012) |
| 80% of RMT | 600 | RT |
| (Clark et al. 2019) |
| 70% of RMT | 600 | RT |
| (Gann et al. 2021) |
| 80% of AMT | 600 | RT and accuracy |
| (Ambrus et al. 2020) |
| 55% of MSO | 3000 (1500/hemisphere) | Learning score based on RT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szücs-Bencze, L.; Vékony, T.; Pesthy, O.; Szabó, N.; Kincses, T.Z.; Turi, Z.; Nemeth, D. Modulating Visuomotor Sequence Learning by Repetitive Transcranial Magnetic Stimulation: What Do We Know So Far? J. Intell. 2023, 11, 201. https://doi.org/10.3390/jintelligence11100201
Szücs-Bencze L, Vékony T, Pesthy O, Szabó N, Kincses TZ, Turi Z, Nemeth D. Modulating Visuomotor Sequence Learning by Repetitive Transcranial Magnetic Stimulation: What Do We Know So Far? Journal of Intelligence. 2023; 11(10):201. https://doi.org/10.3390/jintelligence11100201
Chicago/Turabian StyleSzücs-Bencze, Laura, Teodóra Vékony, Orsolya Pesthy, Nikoletta Szabó, Tamás Zsigmond Kincses, Zsolt Turi, and Dezso Nemeth. 2023. "Modulating Visuomotor Sequence Learning by Repetitive Transcranial Magnetic Stimulation: What Do We Know So Far?" Journal of Intelligence 11, no. 10: 201. https://doi.org/10.3390/jintelligence11100201