Cadherin 11 Inhibition Downregulates β-catenin, Deactivates the Canonical WNT Signalling Pathway and Suppresses the Cancer Stem Cell-Like Phenotype of Triple Negative Breast Cancer

 ,
,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Public Dataset Analyses
2.2. Immunohistochemistry
2.3. Cell Culture
2.4. CDH11 Knockdown
2.5. Western Blotting
2.6. Transwell Matrigel Invasion and Migration Assay
2.7. Immunofluorescence Assay
2.8. Mammosphere Formation Assay
2.9. In Vivo Study
2.10. Statistical Analysis
3. Results
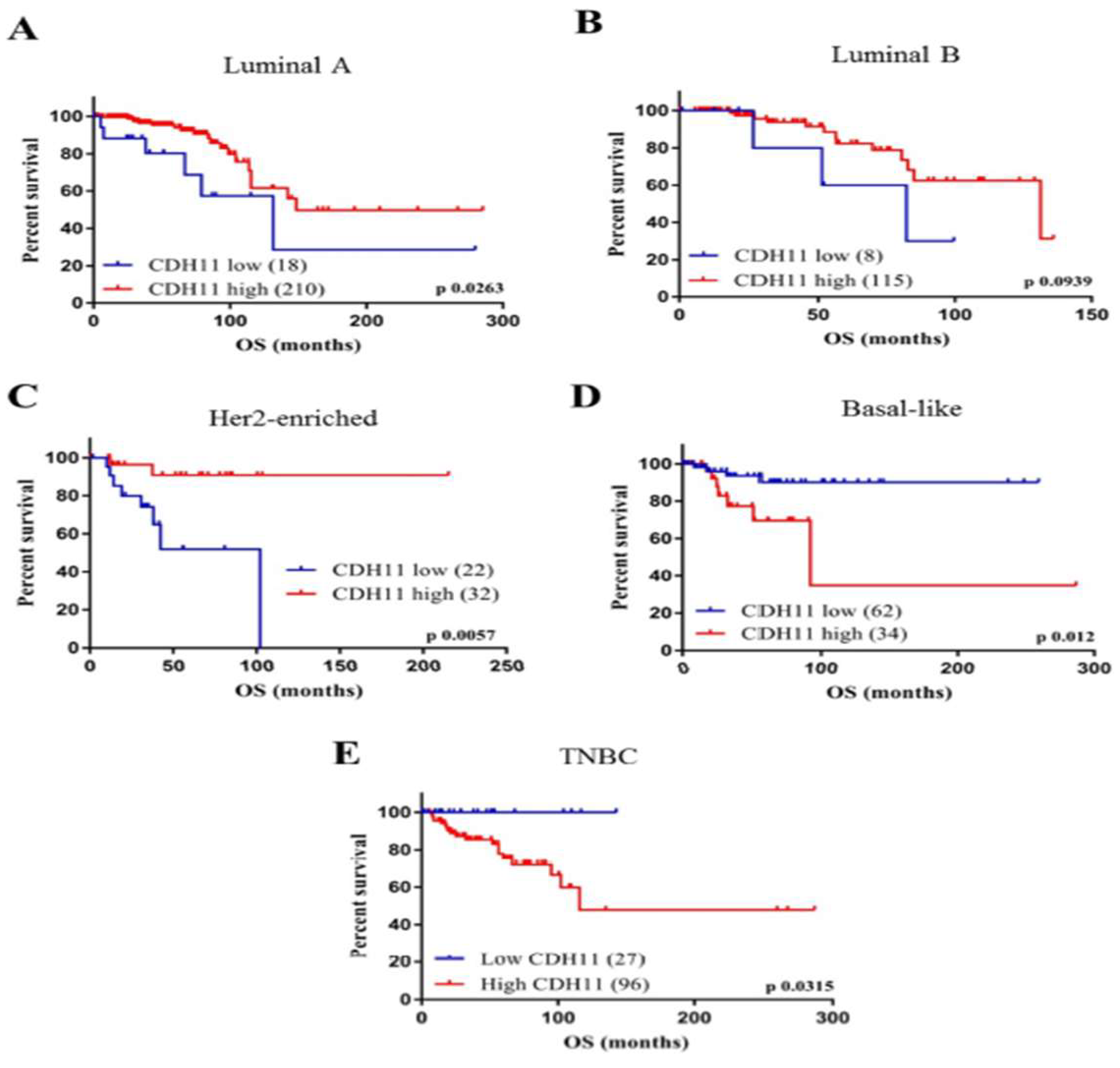
3.1. High CDH11 Expression has Significant Positive Correlation with Poor Overall Survival in Patients with Basal-Like and Triple Negative Breast Cancer
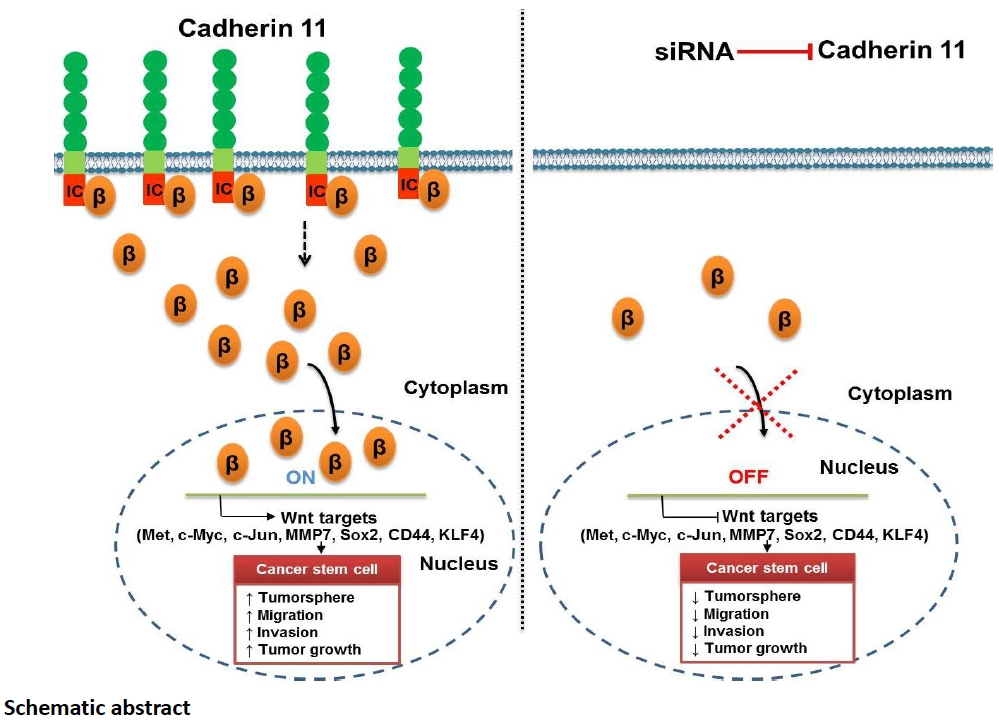
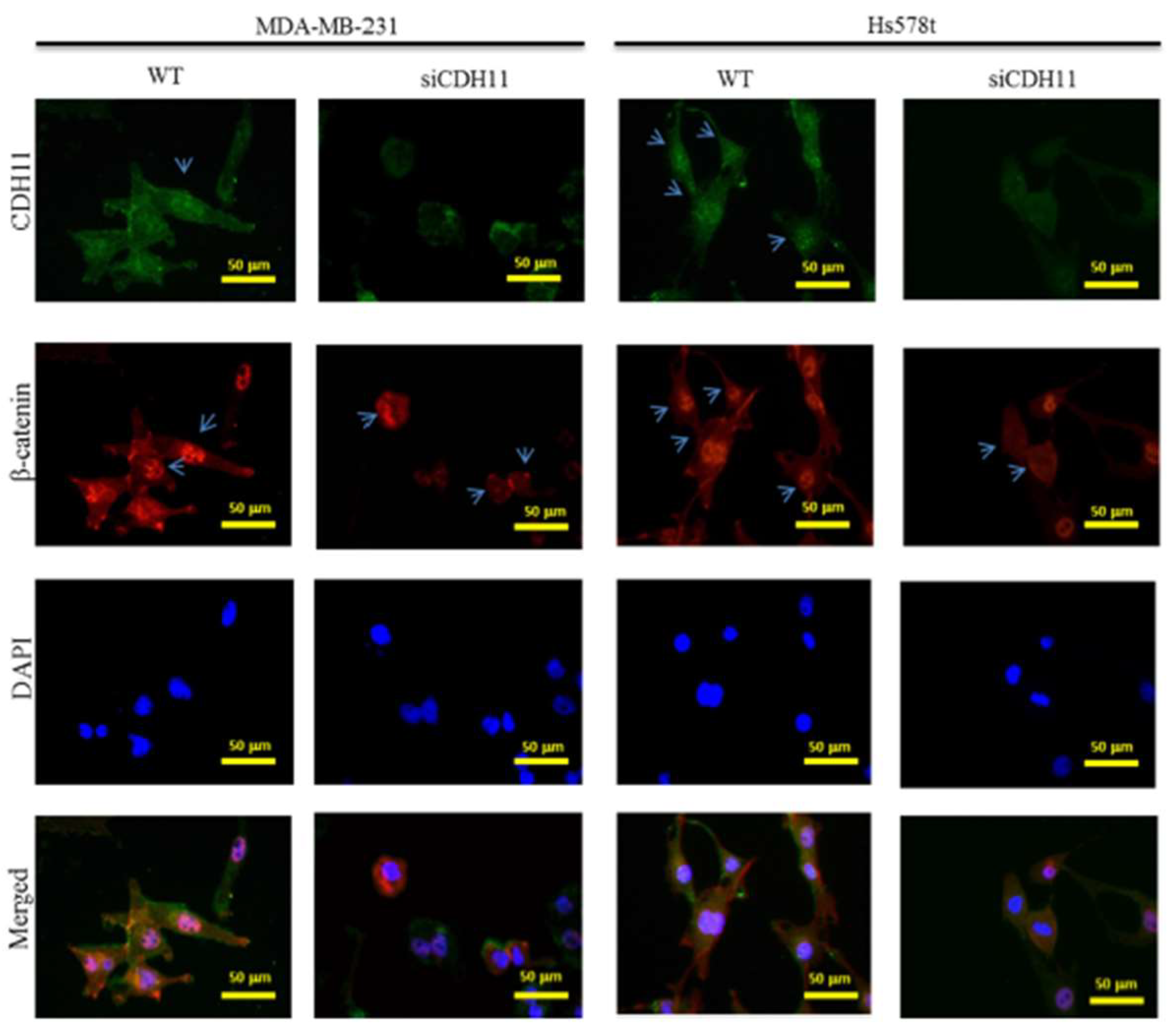
3.2. CDH11 Positively Modulates β-Catenin Expression and Is Associated with Activation of the Canonical WNT Signalling Pathway
3.3. CDH11 Inhibition Suppresses the Stem Cell-Like Phenotype of TNBC Cells
3.4. CDH11 Inhibition Markedly Attenuate the Migration And invasion of TNBC Cells
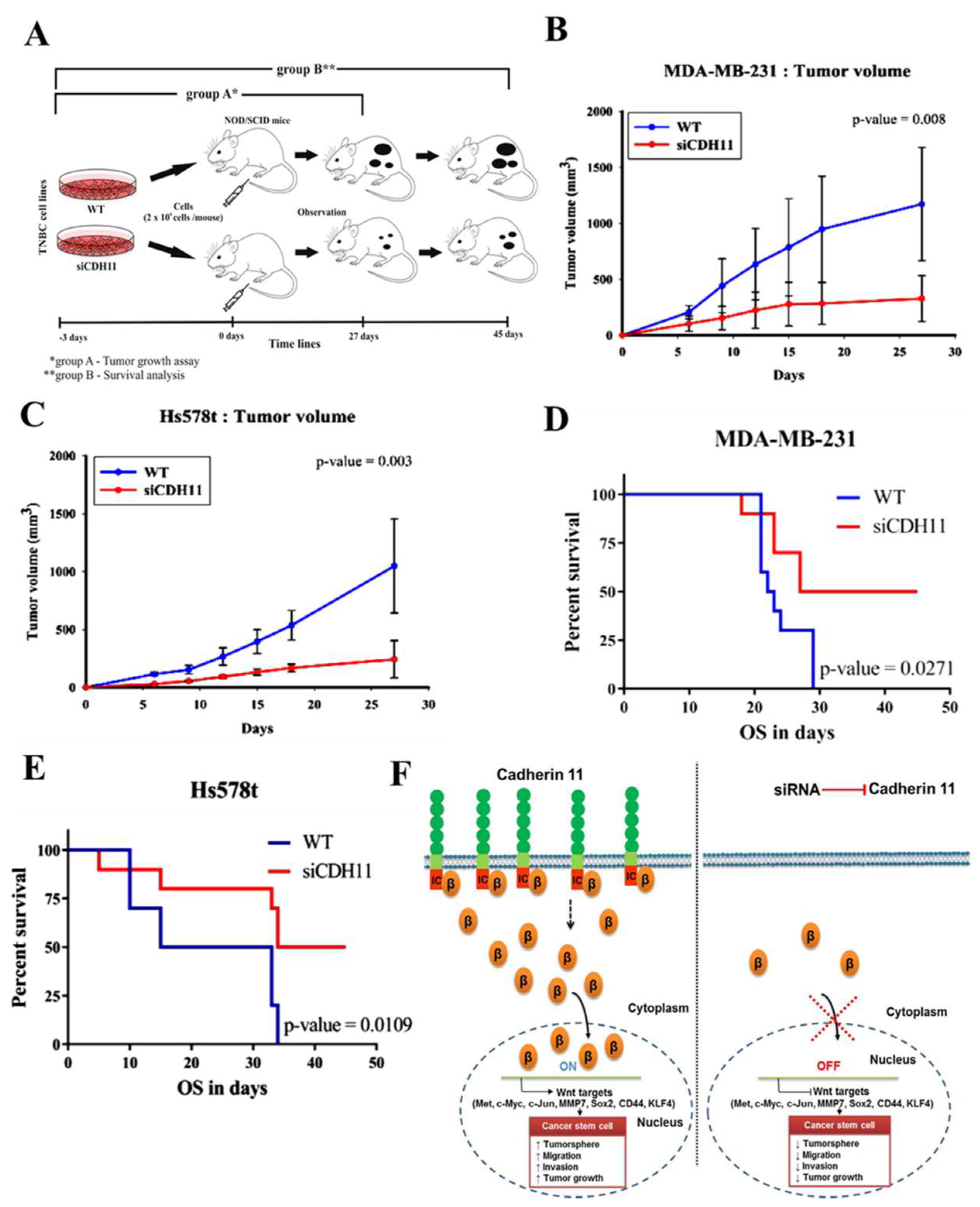
3.5. Silencing CDH11 Significantly Attenuates Tumorigenicity and Tumor Growth of TNBC Cells, In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
Abbreviations
| CDH11 | cadherin 11 |
| CSC | cancer stem cell |
| TNBC | triple negative breast cancer |
| WNT | wingless-type MMTV integration site |
| siRNA | small interfering RNA |
| NOD/SCID | non-obese diabetic/severe combined immunodeficiency |
References
- Rakha, E.A.; Chan, S. Metastatic Triple-negative Breast Cancer. Clin. Oncol. 2011, 23, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Balko, J.M.; Schwarz, L.J.; Luo, N.; Estrada, M.V.; Giltnane, J.M.; Davila-Gonzalez, D.; Wang, K.; Sanchez, V.; Dean, P.T.; Combs, S.E.; et al. Triple-negative breast cancers with amplification of JAK2 at the 9p24 locus demonstrate JAK2-specific dependence. Sci. Transl. Med. 2016, 8, 334ra53. [Google Scholar] [CrossRef] [PubMed]
- Largillier, R.; Ferrero, J.M.; Doyen, J.; Barriere, J.; Namer, M.; Mari, V.; Courdi, A.; Hannoun-Levi, J.M.; Ettore, F.; Birtwisle-Peyrottes, I.; et al. Prognostic factors in 1038 women with metastatic breast cancer. Ann. Oncol. 2008, 19, 2012–2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koury, J.; Zhong, L.; Hao, J. Targeting Signalling Pathways in Cancer Stem Cells for Cancer Treatment. Stem Cells Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, L.N.; Chow, E.K.-H. Mechanisms of chemoresistance in cancer stem cells. Clin. Transl. Med. 2013, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, L.; Nurcombe, V.; Cool, S.M. Wnt signalling controls the fate of mesenchymal stem cells. Gene 2009, 433, 1–7. [Google Scholar] [CrossRef]
- Pohl, S.-G.; Brook, N.; Agostino, M.; Arfuso, F.; Kumar, A.P.; Dharmarajan, A. Wnt signalling in triple-negative breast cancer. Oncogenesis 2017, 6, e310. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signalling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Xu, J.; Prosperi, J.R.; Choudhury, N.; Olopade, O.I.; Goss, K.H. Beta-Catenin Is Required for the Tumorigenic Behavior of Triple-Negative Breast Cancer Cells. PLoS ONE 2015, 10, 1–11. [Google Scholar]
- Hendriksen, J.; Jansen, M.; Brown, C.M.; van der Velde, H.; van Ham, M.; Galjart, N.; Offerhaus, G.J.; Fagotto, F.; Fornerod, M. Plasma membrane recruitment of dephosphorylated -catenin upon activation of the Wnt pathway. J. Cell Sci. 2008, 121, 1793–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsulic, S.; Huber, O.; Kemler, R. E-cadherin binding prevents β-catenin nuclear localization and β-catenin/LEF-1-mediated transactivation. J. Cell Sci. 1999, 112, 1237–1245. [Google Scholar] [PubMed]
- Howard, S.; Deroo, T.; Fujita, Y.; Itasaki, N. A positive role of cadherin in wnt/β-catenin signalling during epithelial-mesenchymal transition. PLoS ONE 2011, 6, e23899. [Google Scholar] [CrossRef] [PubMed]
- Pohlodek, K.; Tan, Y.Y.; Singer, C.F.; Gschwantler-Kaulich, D. Cadherin-11 expression is upregulated in invasive human breast cancer. Oncol. Lett. 2016, 12, 4393–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satcher, R.L.; Pan, T.; Cheng, C.J.; Lee, Y.C.; Lin, S.C.; Yu, G.; Li, X.; Hoang, A.G.; Tamboli, P.; Jonasch, E.; et al. Cadherin-11 in Renal Cell Carcinoma Bone Metastasis. PLoS ONE 2014, 9, e89880. [Google Scholar] [CrossRef]
- Chu, K.; Cheng, C.-J.; Ye, X.; Lee, Y.-C.; Zurita, A.J.; Chen, D.-T.; Yu-Lee, L.-Y.; Zhang, S.; Yeh, E.T.; Hu, M.C.-T.; et al. Cadherin-11 Promotes the Metastasis of Prostate Cancer Cells to Bone. Mol. Cancer Res. 2008, 6, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Anders, C.K.; Carey, L.A.; Frazier, D.P.; Kendig, R.D. Biology, Metastatic Patterns and Treatment of Patients with Triple-Negtive Breast Cancer. Clin. Breast Cancer 2010, 9, S73–S81. [Google Scholar] [CrossRef]
- Altaner, C.; Altanerova, V. Stem cell based glioblastoma gene therapy. Neoplasma 2012, 59, 622–630. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Assefnia, S.; Dakshanamurthy, S.; Guidry Auvil, J.M.; Hampel, C.; Anastasiadis, P.Z.; Kallakury, B.; Uren, A.; Foley, D.W.; Brown, M.L.; Shapiro, L.; et al. Cadherin-11 in poor prognosis malignancies and rheumatoid arthritis: Common target, common therapies. Oncotarget 2014, 5, 1458–1474. [Google Scholar] [CrossRef]
- Fedchenko, N.; Reifenrath, J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue—A review. Diagn. Pathol. 2014, 9, 221. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.S.; Chanana, P.; Jhamb, S. Biomarkers in triple negative breast cancer: A review. World J. Clin. Oncol. 2015, 6, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Prat, A.; Adamo, B.; Cheang, M.C.U.; Anders, C.K.; Carey, L.A.; Perou, C.M. Molecular Characterization of Basal-Like and Non-Basal-Like Triple-Negative Breast Cancer. Oncologist 2013, 18, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.B.; Kim, J.Y.; Cho, S.D.; Park, K.S.; Jung, J.Y.; Lee, H.Y.; Hong, I.S.; Nam, J.S. Blockade of Wnt/β-catenin signalling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci. Rep. 2015, 5, 12465. [Google Scholar] [CrossRef] [PubMed]
- Birtolo, C.; Pham, H.; Morvaridi, S.; Chheda, C.; Go, V.L.W.; Ptasznik, A.; Edderkaoui, M.; Weisman, M.H.; Noss, E.; Brenner, M.B.; et al. Cadherin-11 Is a Cell Surface Marker Up-Regulated in Activated Pancreatic Stellate Cells and Is Involved in Pancreatic Cancer Cell Migration. Am. J. Pathol. 2017, 187, 146–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Ying, J.; Li, H.; Zhang, Y.; Shu, X.; Fan, Y.; Tan, J.; Cao, Y.; Tsao, S.W.; Srivastava, G.; et al. The human cadherin 11 is a pro-apoptotic tumor suppressor modulating cell stemness through Wnt/β-catenin signalling and silenced in common carcinomas. Oncogene 2012, 31, 3901–3912. [Google Scholar] [CrossRef] [PubMed]
- VON Bülow, C.; Oliveira-Ferrer, L.; Loning, T.; Trillsch, F.; Manher, S.; Milde-Langosch, K. Cadherin-11 mRNA and protein expression in ovarian tumors of different malignancy: No evidence of oncogenic or tumor-suppressive function. Mol. Clin. Oncol. 2015, 3, 1067–1072. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Wang, Z.; Zhou, L.; Ren, Y.; Gong, Y.; Qin, W.; Bai, L.; Hu, J.; Wang, T. The role of cadherin-11 in microcystin-LR-induced migration and invasion in colorectal carcinoma cells. Oncol. Lett. 2018, 15, 1417–1422. [Google Scholar] [CrossRef]
- Marchong, M.N.; Yurkowski, C.; Ma, C.; Spencer, C.; Pajovic, S.; Gallie, B.L. Cdh11 acts as a tumor suppressor in a murine retinoblastoma model by facilitating tumor cell death. PLoS Genet. 2010, 6, e1000923. [Google Scholar] [CrossRef]
- Pishvaian, M.J.; Feltes, C.M.; Thompson, P.; Bussemakers, M.J.; Schalken, J.A.; Byers, S.W. Cadherin-11 is expressed in invasive breast cancer cell lines. Cancer Res. 1999, 59, 947–952. [Google Scholar]
- Ding, F.; Wang, M.; Du, Y.; Du, S.; Zhu, Z.; Yan, Z. BHX Inhibits the Wnt Signalling Pathway by Suppressing β-catenin Transcription in the Nucleus. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Noss, E.H.; Watts, G.F.M.; Zocco, D.; Keller, T.L.; Whitman, M.; Blobel, C.P.; Lee, D.M.; Brenner, M.B. Evidence for cadherin-11 cleavage in the synovium and partial characterization of its mechanism. Arthritis Res. Ther. 2015, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mullooly, M.; McGowan, P.M.; Kennedy, S.A.; Madden, S.F.; Crown, J.; O’Donovan, N.; Duffy, M.J. ADAM10: A new player in breast cancer progression? Br. J. Cancer 2015, 113, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Qin, S.; Schulte, B.A.; Ethier, S.P.; Tew, K.D.; Wang, G.Y. MYC inhibition depletes cancer stem-like cells in triple-negative breast cancer. Cancer Res. 2017, 77, 6641–6650. [Google Scholar] [CrossRef] [PubMed]
- Idowu, M.O.; Kmieciak, M.; Dumur, C.; Burton, R.S.; Grimes, M.M.; Powers, C.N.; Manjili, M.H. CD44(+)/CD24(−/low) cancer stem/progenitor cells are more abundant in triple-negative invasive breast carcinoma phenotype and are associated with poor outcome. Hum. Pathol. 2012, 43, 364–373. [Google Scholar] [CrossRef]
- Van Raay, T.J.; Moore, K.B.; Iordanova, I.; Steele, M.; Jamrich, M.; Harris, W.A.; Vetter, M.L. Frizzled 5 signalling governs the neural potential of progenitors in the developing Xenopus retina. Neuron 2005, 46, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Leis, O.; Eguiara, A.; Lopez-Arribillaga, E.; Alberdi, M.J.; Hernandez-Garcia, S.; Elorriaga, K.; Pandiella, A.; Rezola, R.; Martin, A.G. Sox2 expression in breast tumours and activation in breast cancer stem cells. Oncogene 2012, 31, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Feltes, C.M.; Kudo, A.; Blaschuk, O.; Byers, S.W. An alternatively spliced cadherin-11 enhances human breast cancer cell invasion. Cancer Res. 2002, 62, 6688–6697. [Google Scholar] [PubMed]
- Tomita, K.; Van Bokhoven, A.; Van Leenders, G.J.L.H.; Ruijter, E.T.G.; Jansen, C.F.J.; Bussemakers, M.J.G.; Schalken, J.A. Cadherin switching in human prostate cancer progression. Cancer Res. 2000, 60, 3650–3654. [Google Scholar] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satriyo, P.B.; Bamodu, O.A.; Chen, J.-H.; Aryandono, T.; Haryana, S.M.; Yeh, C.-T.; Chao, T.-Y. Cadherin 11 Inhibition Downregulates β-catenin, Deactivates the Canonical WNT Signalling Pathway and Suppresses the Cancer Stem Cell-Like Phenotype of Triple Negative Breast Cancer. J. Clin. Med. 2019, 8, 148. https://doi.org/10.3390/jcm8020148
Satriyo PB, Bamodu OA, Chen J-H, Aryandono T, Haryana SM, Yeh C-T, Chao T-Y. Cadherin 11 Inhibition Downregulates β-catenin, Deactivates the Canonical WNT Signalling Pathway and Suppresses the Cancer Stem Cell-Like Phenotype of Triple Negative Breast Cancer. Journal of Clinical Medicine. 2019; 8(2):148. https://doi.org/10.3390/jcm8020148
Chicago/Turabian StyleSatriyo, Pamungkas Bagus, Oluwaseun Adebayo Bamodu, Jia-Hong Chen, Teguh Aryandono, Sofia Mubarika Haryana, Chi-Tai Yeh, and Tsu-Yi Chao. 2019. "Cadherin 11 Inhibition Downregulates β-catenin, Deactivates the Canonical WNT Signalling Pathway and Suppresses the Cancer Stem Cell-Like Phenotype of Triple Negative Breast Cancer" Journal of Clinical Medicine 8, no. 2: 148. https://doi.org/10.3390/jcm8020148