
An Approach to Identify HLA Class II Immunogenic Epitopes in the Greek Population through Machine Learning Algorithms
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Anti-HLA Antibodies Measurement
2.3. Descriptive Statistics
3. Results
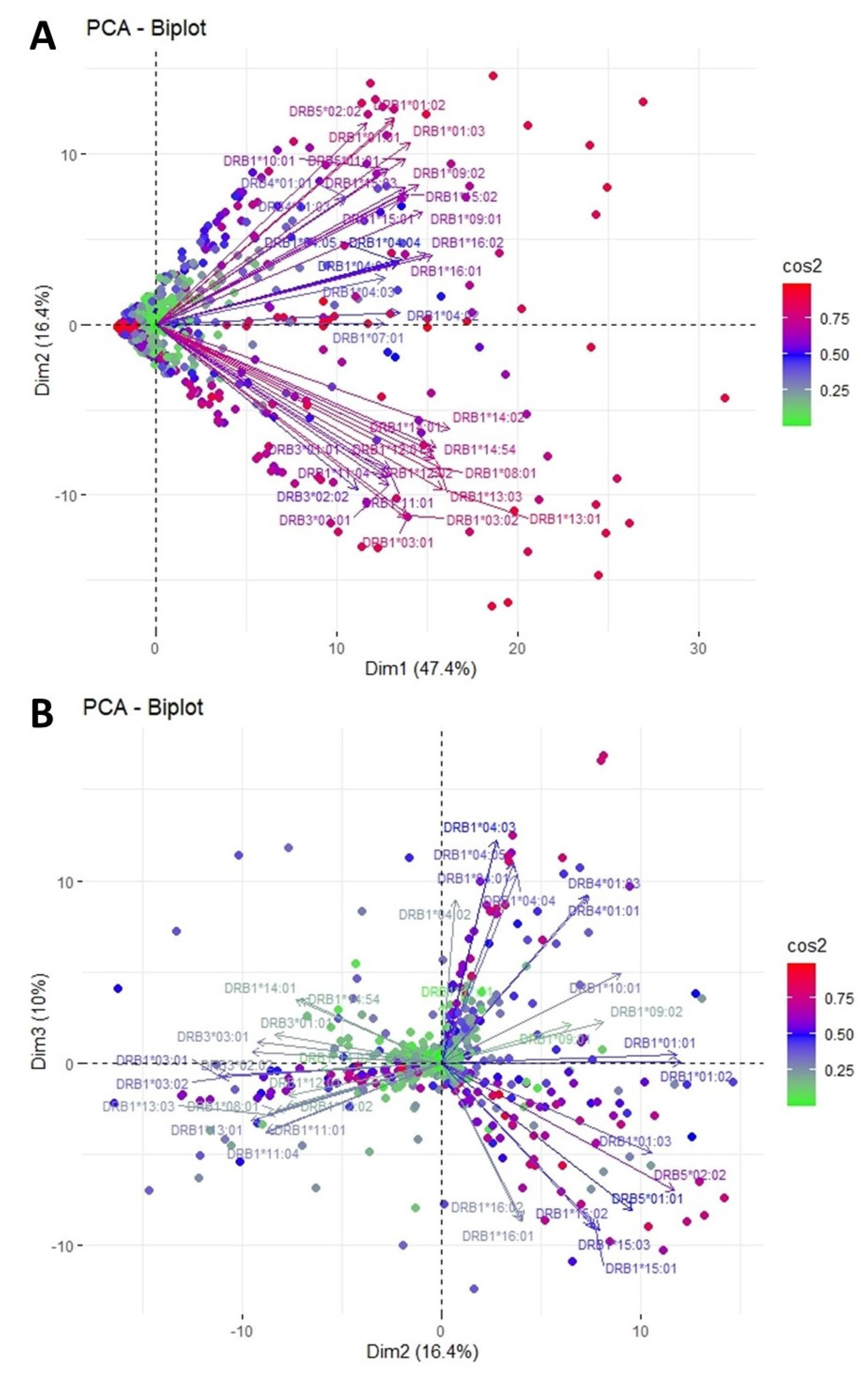
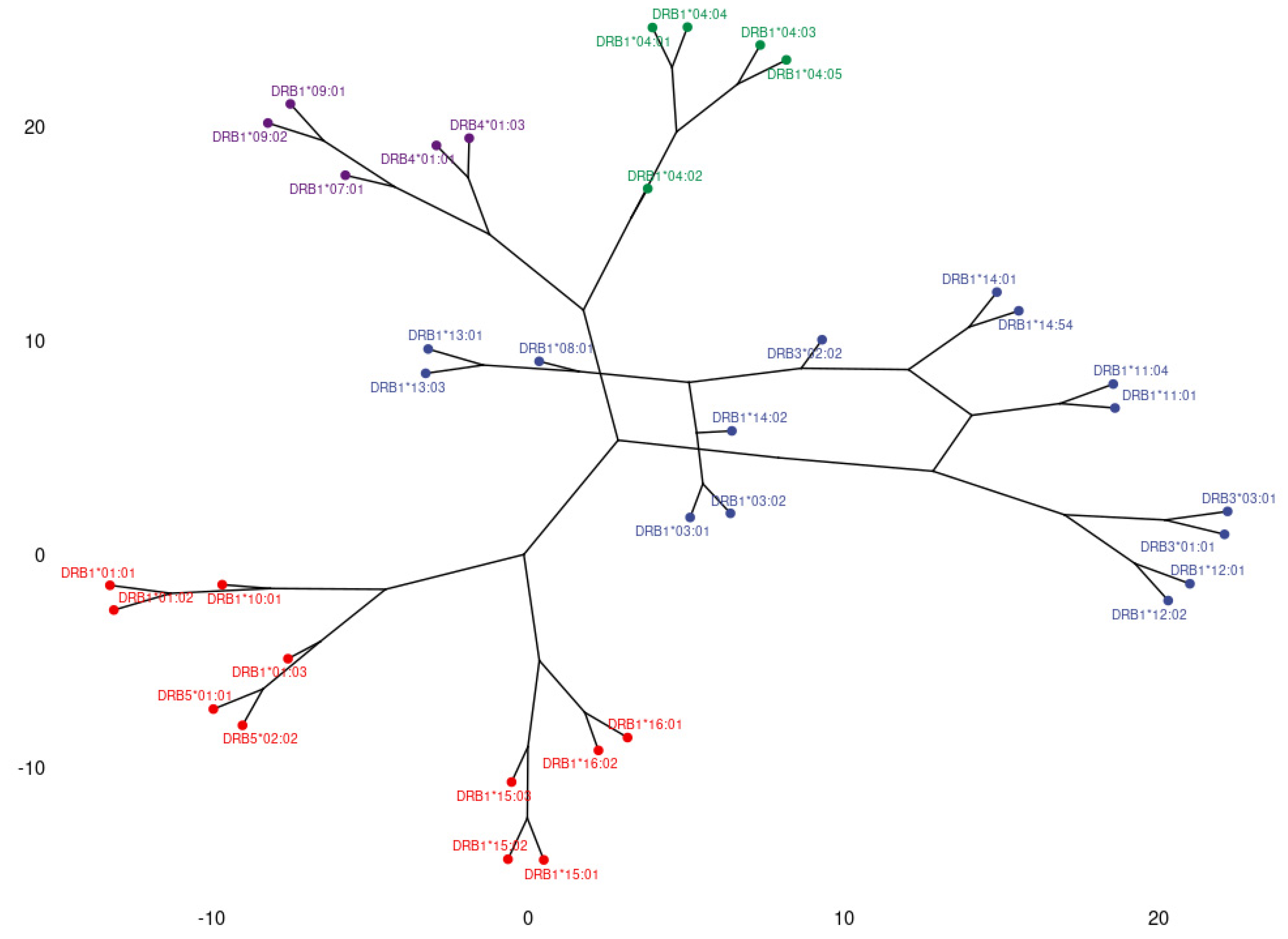
3.1. Clustering and Epitopic Analysis of Anti-HLA-DR Immune Response
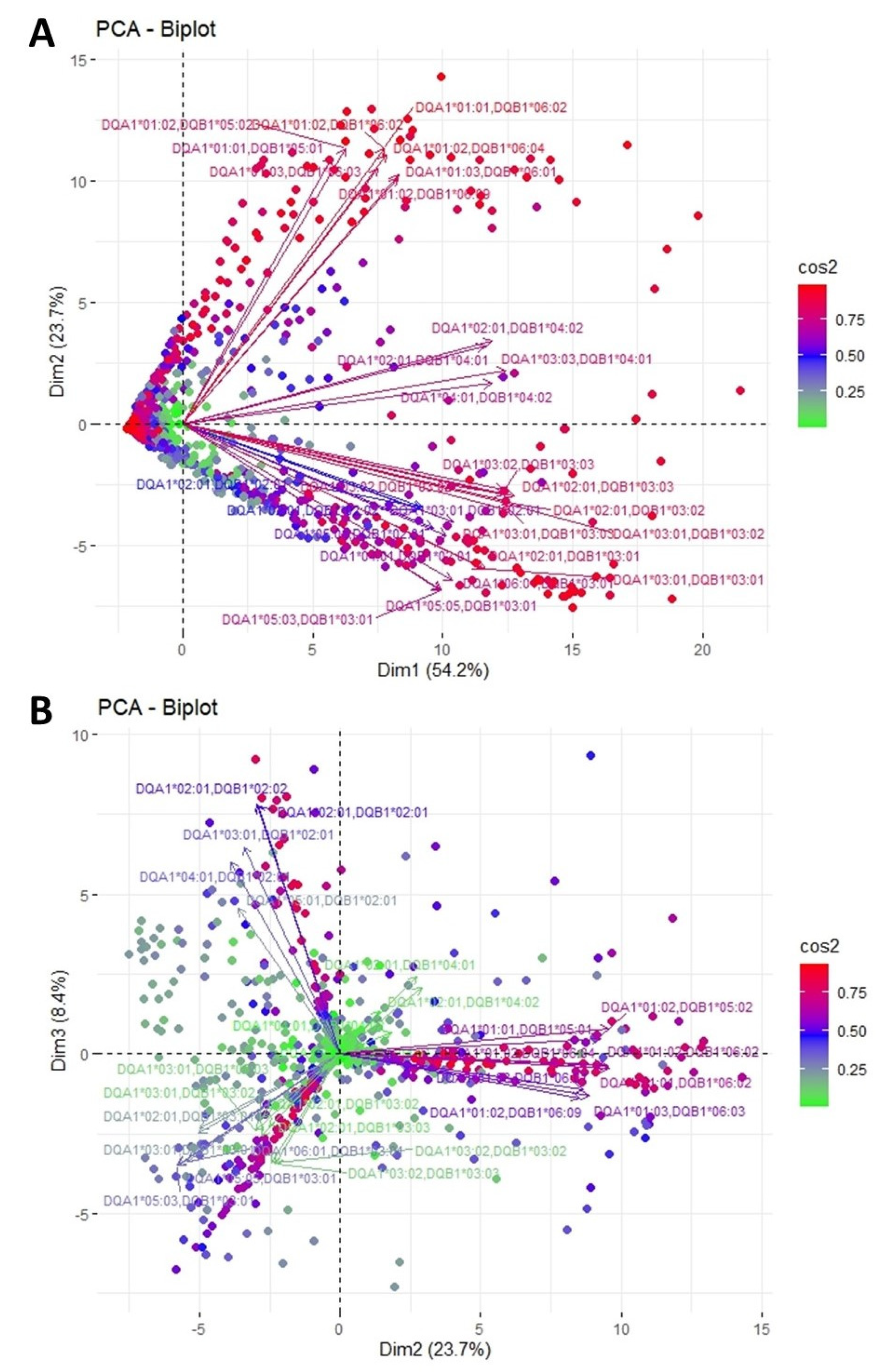
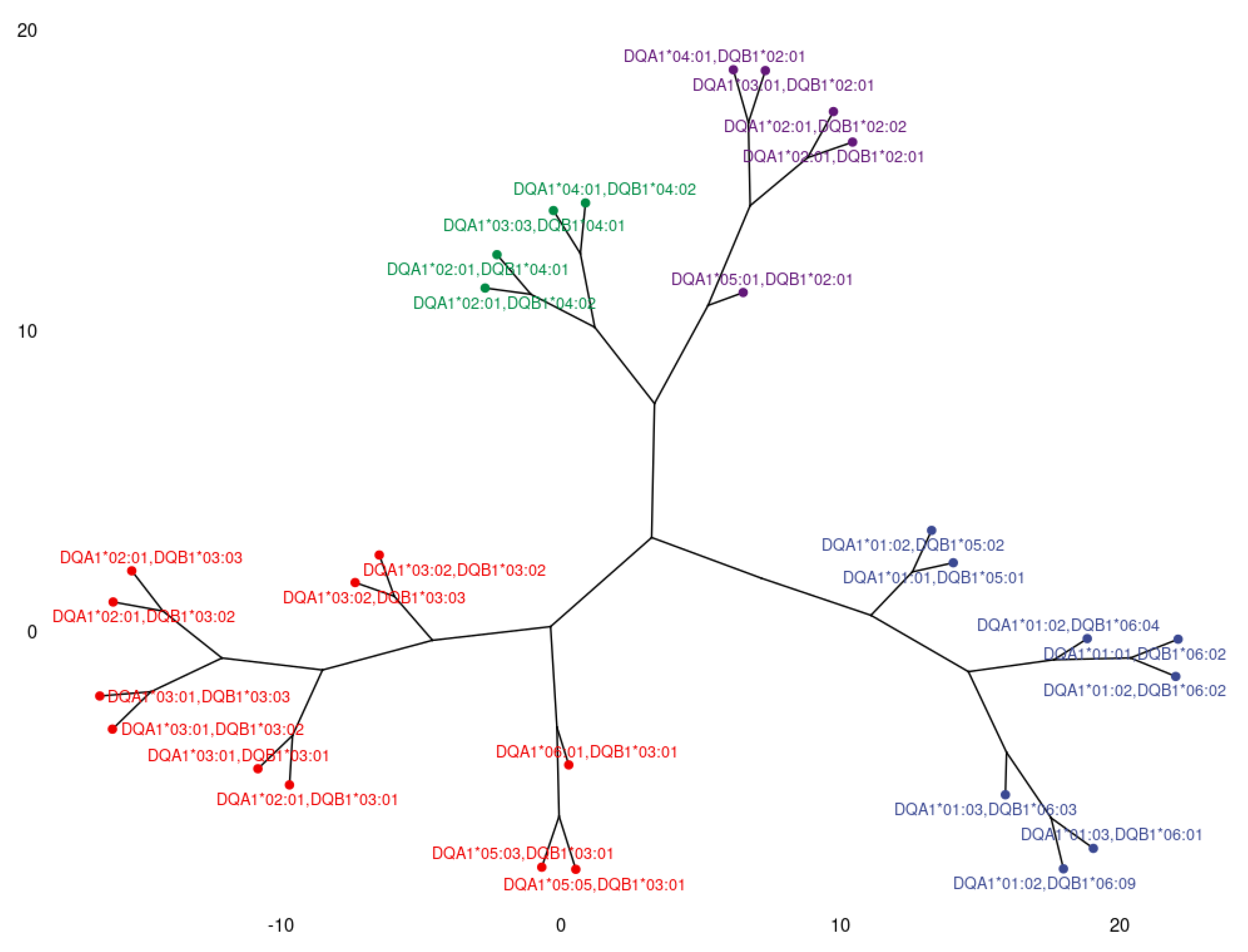
3.2. Clustering and Epitopic Analysis of Anti-HLA-DQ Immune Response
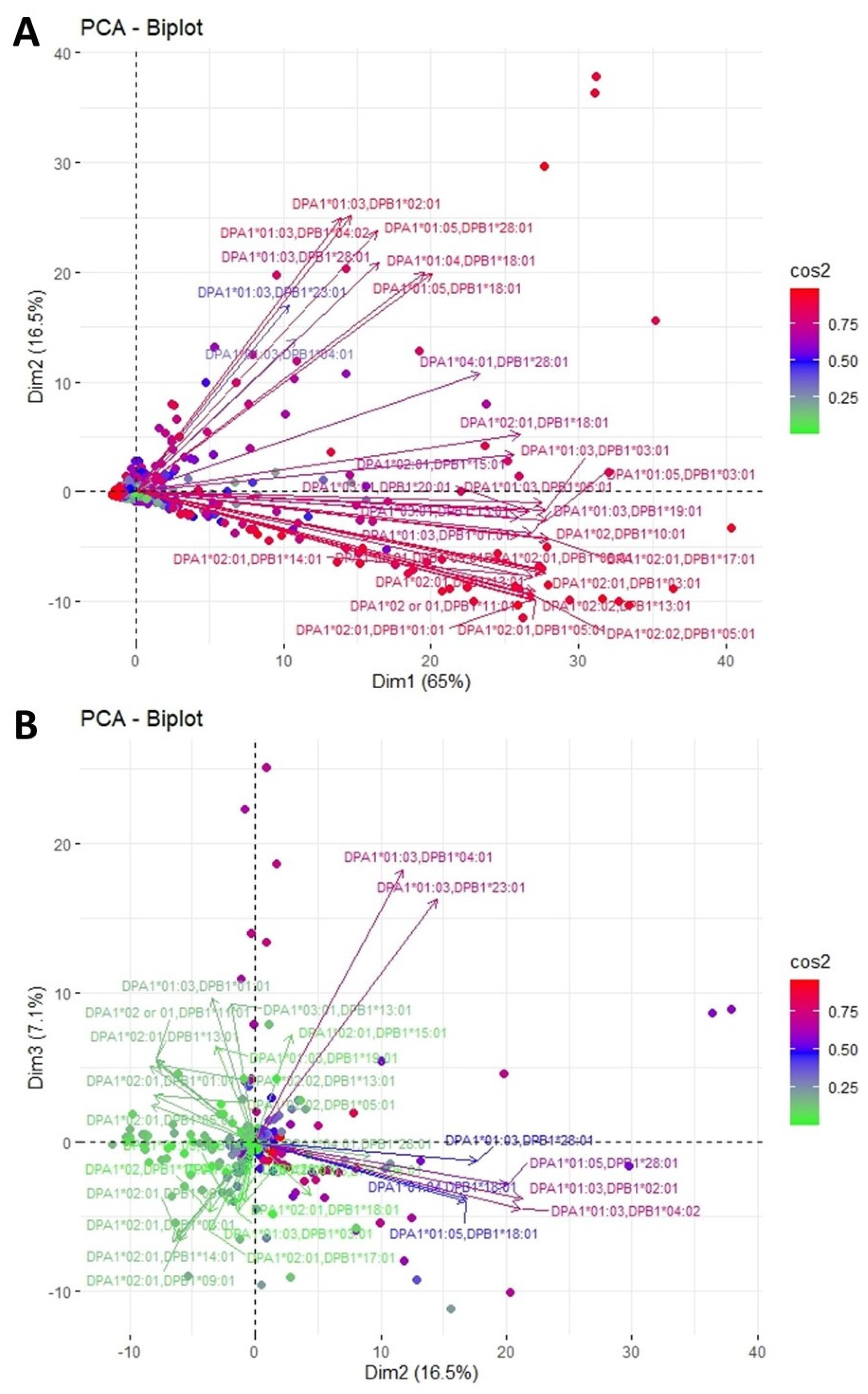
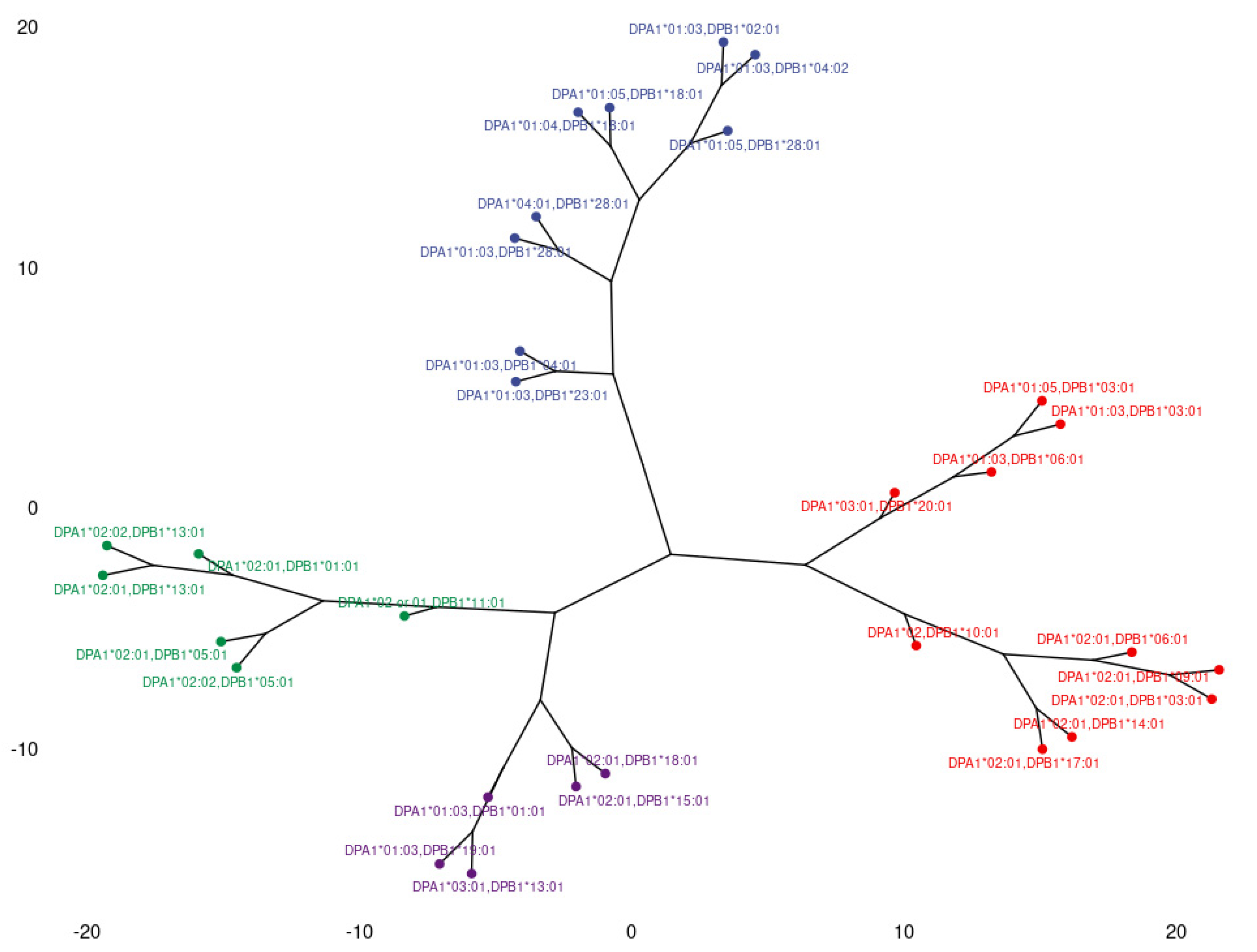
3.3. Clustering and Epitopic Analysis of Anti-HLA-DP Immune Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheldon, S.; Poulton, K. HLA typing and its influence on organ transplantation. Methods Mol. Biol. 2006, 333, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, B.M. A glow of HLA typing in organ transplantation. Clin. Transl. Med. 2013, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muczynski, K.A.; Cotner, T.; Anderson, S.K. Unusual expression of human lymphocyte antigen class II in normal renal microvascular endothelium. Kidney Int. 2001, 59, 488–497. [Google Scholar] [CrossRef] [Green Version]
- Muczynski, K.A.; Ekle, D.M.; Coder, D.M.; Anderson, S.K. Normal Human Kidney HLA-DR–Expressing Renal Microvascular Endothelial Cells: Characterization, Isolation, and Regulation of MHC Class II Expression. J. Am. Soc. Nephrol. 2003, 14, 1336–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matern, B.M.; Olieslagers, T.I.; Voorter, C.E.M.; Groeneweg, M.; Tilanus, M.G.J. Insights into the polymorphism in HLA-DRA and its evolutionary relationship with HLA haplotypes. HLA 2020, 95, 117–127. [Google Scholar] [CrossRef]
- Choo, S.Y. The HLA System: Genetics, Immunology, Clinical Testing, and Clinical Implications. Yonsei Med. J. 2007, 48, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Tambur, A.R. Auto- and allo-epitopes in DQ alloreactive antibodies. Curr. Opin. Organ Transplant. 2016, 21, 355–361. [Google Scholar] [CrossRef]
- Tambur, A.R.; Kosmoliaptsis, V.; Claas, F.H.; Mannon, R.B.; Nickerson, P.; Naesens, M. Significance of HLA-DQ in kidney transplantation: Time to reevaluate human leukocyte antigen–matching priorities to improve transplant outcomes? An expert review and recommendations. Kidney Int. 2021, 100, 1012–1022. [Google Scholar] [CrossRef]
- Cross, A.R.; Lion, J.; Poussin, K.; Glotz, D.; Mooney, N. Inflammation Determines the Capacity of Allogenic Endothelial Cells to Regulate Human Treg Expansion. Front. Immunol. 2021, 12, 666531. [Google Scholar] [CrossRef]
- Lion, J.; Taflin, C.; Cross, A.R.; Robledo-Sarmiento, M.; Mariotto, E.; Savenay, A.; Carmagnat, M.; Suberbielle, C.; Charron, D.; Haziot, A.; et al. HLA Class II Antibody Activation of Endothelial Cells Promotes Th17 and Disrupts Regulatory T Lymphocyte Expansion. Am. J. Transplant. 2016, 16, 1408–1420. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.R.; Lion, J.; Poussin, K.; Assayag, M.; Taupin, J.-L.; Glotz, D.; Mooney, N. HLA-DQ alloantibodies directly activate the endothelium and compromise differentiation of FoxP3high regulatory T lymphocytes. Kidney Int. 2019, 96, 689–698. [Google Scholar] [CrossRef]
- Vittoraki, A.G.; Fylaktou, A.; Tarassi, K.; Tsinaris, Z.; Siorenta, A.; Petasis, G.C.; Gerogiannis, D.; Lehmann, C.; Carmagnat, M.; Doxiadis, I.; et al. Hidden Patterns of Anti-HLA Class I Alloreactivity Revealed Through Machine Learning. Front. Immunol. 2021, 12, 670956. [Google Scholar] [CrossRef]
- Vittoraki, A.G.; Fylaktou, A.; Tarassi, K.; Tsinaris, Z.; Petasis, G.C.; Gerogiannis, D.; Kheav, V.-D.; Carmagnat, M.; Lehmann, C.; Doxiadis, I.; et al. Patterns of 1,748 Unique Human Alloimmune Responses Seen by Simple Machine Learning Algorithms. Front. Immunol. 2020, 11, 1667. [Google Scholar] [CrossRef]
- Schnaidt, M.; Weinstock, C.; Jurisic, M.; Schmid-Horch, B.; Ender, A.; Wernet, D. HLA Antibody Specification Using Single-Antigen Beads—A Technical Solution for the Prozone Effect. Transplantation 2011, 92, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, A. The application of principal component analysis to drug discovery and biomedical data. Drug Discov. Today 2017, 22, 1069–1076. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.H., Jr. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- De Amorim, R.C.; Makarenkov, V.; Mirkin, B. A-Ward pβ: Effective hierarchical clustering using the Minkowski metric and a fast k -means initialisation. Inf. Sci. 2016, 370–371, 343–354. [Google Scholar] [CrossRef] [Green Version]
- HLA Epitope Registry. Available online: https://www.epregistry.com.br/ (accessed on 1 October 2022).
- Cross, A.R.; Lion, J.; Loiseau, P.; Charron, D.; Taupin, J.-L.; Glotz, D.; Mooney, N. Donor Specific Antibodies are not only directed against HLA-DR: Minding your Ps and Qs. Hum. Immunol. 2016, 77, 1092–1100. [Google Scholar] [CrossRef]
- Ntokou, I.-S.A.; Iniotaki, A.G.; Kontou, E.N.; Darema, M.N.; Apostolaki, M.D.; Kostakis, A.G.; Boletis, J.N. Long-term follow up for anti-HLA donor specific antibodies postrenal transplantation: High immunogenicity of HLA class II graft molecules. Transpl. Int. 2011, 24, 1084–1093. [Google Scholar] [CrossRef]
- Tambur, A.R. HLA-Epitope Matching or Eplet Risk Stratification: The Devil Is in the Details. Front. Immunol. 2018, 9, 2010. [Google Scholar] [CrossRef] [PubMed]
- Duquesnoy, R.J.; Askar, M. HLAMatchmaker: A Molecularly Based Algorithm for Histocompatibility Determination. V. Eplet Matching for HLA-DR, HLA-DQ, and HLA-DP. Hum. Immunol. 2007, 68, 12–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geneugelijk, K.; Thus, K.A.; Spierings, E. Predicting Alloreactivity in Transplantation. J. Immunol. Res. 2014, 2014, 159479. [Google Scholar] [CrossRef] [PubMed]
- Kamburova, E.G.; Hoitsma, A.; Claas, F.H.; Otten, H.G. On behalf of the PROCARE Consortium Results and reflections from the PROfiling Consortium on Antibody Repertoire and Effector functions in kidney transplantation: A mini-review. HLA 2019, 94, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Gniazdowski, Z. New Interpretation of Principal Components Analysis. Zesz. Nauk. WWSI 2017, 11, 43–65. [Google Scholar]
- Ponomarenko, J.V.; Bui, H.-H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Tambur, A.R.; Leventhal, J.R.; Friedewald, J.; Ramon, D.S. The Complexity of Human Leukocyte Antigen (HLA)-DQ Antibodies and Its Effect on Virtual Crossmatching. Transplantation 2010, 90, 1117–1124. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anti-HLA-DRB Group Responses in PCA | HLA-DR Eplets Recognized by Antibody Groups in PCA | Location of Dendrogram Antibody Groups that Appear to Recognize the Corresponding Eplets | HLA-DR Alleles where the Eplets are Expressed | ElliPro Score |
|---|---|---|---|---|
| Group C | ||||
| *01:01, *01:02, *01:03, *09:01, *09:02, *10:01, *15:01, *15:02, *15:03, *16:01, *16:02, DRB5*01:01 DRB5*02:02 | 6C | upper | DRB5*02:02 | High |
| 13FE | upper | DRB1*01:01, 01:02, 01:03 DRB1*09:01, 09:02 | Very Low | |
| 28H | upper | DRB1*09:01, 09:02 DRB5*01:01 DRB5*02:02 | Low | |
| 30C | upper | DRB1*01:01, 01:02, 01:03 | Intermediate | |
| 30D | upper | DRB5*01:01 | Intermediate | |
| 30G | upper | DRB1*09:01, 09:02 DRB5*02:02 | Intermediate | |
| 30RV | upper | DRB1*10 | Intermediate | |
| 37S | both | DRB1*01:01, 01:02, 01:03 DRB1*15:01, 15:02, 15:03 DRB1*16:01, 16:02 | High | |
| 71A | both | DRB1*15:01, 15:02, 15:03 DRB5*02:02 | Intermediate | |
| 96EV | upper | DRB1*01:01, 01:02, 01:03 DRB5*01:01 DRB5*02:02 | High | |
| 108T | upper | DRB5*01:01 DRB5*02:02 | High | |
| 142M | lower | DRB1*15:01, 15:02, 15:03 DRB1*16:01, 16:02 | High | |
| Group B | ||||
| *04:01 *04:02 *04:03 *04:04 *04:05 *07:01 DRB4*01:01 DRB4*01:03 | 25Q | left | DRB1*07:01 | Intermediate |
| 48Q | left | DRB4*01:01, 01:03 | High | |
| 96Y | right | DRB1*04:01, 04:02, 04:03, 04:04, 04:05 | High | |
| Group A | ||||
| *03:01 *03:02 *08:01 *11:01 *11:04 *12:01 *12:02 *13:01 *13:03 *14:01 *14:02 *14:54 DRB3*01:01 DRB3*02:02 DRB3*03:01 | 11STS | upper | DRB1*03:01, 03:02 DRB1*11:01, 11:04 DRB1*13:01, 13:03 DRB1*14:01, 14:02, 14:54 | Intermediate |
| 13SE | both | DRB1*03:01, 03:02 DRB1*11:01, 11:04 DRB1*13:01, 13:03 DRB1*14:01, 14:02, 14:54 DRB3*01:01 DRB3*02:02 DRB3*03:01 | Very low | |
| 16Y | both | DRB1*08:01, DRB1*12:01, 12:02 | Low | |
| 31FH | both | DRB1*03:01, 03:02 DRB1*12:01, 12:02 DRB1*13:01 DRB1*14:01, 14:02, 14:54 DRB3*01:01 DRB3*02:02 DRB3*03:01 | High | |
| 37FL | lower | DRB3*01:01 | High | |
| 37L | lower | DRB1*12:01, 12:02 | High | |
| 51R | lower | DRB3*02:02 | High | |
| 57A | lower | DRB1*14:01, 14:54 | High | |
| 57DE | lower | DRB1*11:01, 11:04 | High | |
| 70QQ | both | DRB3*02:02 DRB3*03:01 | High | |
| 74L | upper | DRB1*08:01 | High | |
| 74R | both | DRB1*03:01, 03:02 DRB3*01:01 | High | |
| 77N | both | DRB1*03:01, 03:02 DRB3*01:01 DRB3*02:02 DRB3*03:01 | High | |
| 96HK | both | DRB1*03:01, 03:02 DRB1*08:01 DRB1*11:01, 11:04 DRB1*12:01, 12:02 DRB1*13:01 DRB1*14:01, 14:02, 14:54 | ||
| 98Q | both | DRB3*01:01 DRB3*02:02 DRB3*03:01 | Intermediate | |
| 112Y | upper | DRB1*14:01 | High | |
| 149H | both | DRB1*03:01, 03:02 DRB1*08:01 DRB1*11:01, 11:04 DRB1*12:01, 12:02 DRB1*13:01 DRB1*14:01, 14:02, 14:54 DRB3*03:01 | Very low |
| HLA-DQ Eplets Recognized by Antibody Groups in PCA | HLA-DQ Alleles where the Eplets are Expressed | ElliPro Score | |
|---|---|---|---|
| DQA1*01:01, DQB1*05:01 DQA1*01:01, DQB1*06:02 DQA1*01:02, DQB1*05:02 DQA1*01:02, DQB1*06:02 DQA1*01:02, DQB1*06:04 DQA1*01:02, DQB1*06:09 DQA1*01:03, DQB1*06:01 DQA1*01:03, DQB1*06:03 | 3P | DQB1*06:01 | High |
| 30H | DQB1*05:01, 05:02 DQB1*06:04, 06:03 | Intermediate | |
| 37YV | DQB1*05:01, 05:02 | High | |
| 52PQ | Whole DQB1 group | High | |
| 55RPD | DQB1*06:01, 06:02, 06:03 | High | |
| 56PS | DQB1*05:02 | High | |
| 57V | DQB1*05:01 DQB1*06:04, 06:09 | High | |
| 67VG | DQB1*05:01, 05:02 DQB1*06:02, 06:03 | High | |
| 70GT | DQB1*06:02, 06:03 | High | |
| 86A | DQB1*05:01, 05:02 DQB1*06:01, 06:02, 06:03 | Intermediate | |
| 87F | DQB1*06:01, 06:02, 06:03 | High | |
| 87Y | DQB1*05:01, 05:02 DQB1*06:04, 06:09 | Intermediate | |
| 116I | DQB1*05:01, 05:02 | High | |
| 125G | DQB1*06:01, 06:02, 06:03, 06:04, 06:09 | Very low | |
| 125SQ | DQB1*05:01 | Low | |
| 130Q | DQB1*06:04, 06:09 | Intermediate | |
| 52SK | Whole DQA1 group | High | |
| 129QS | DQA1*01:01, 01:02 | High | |
| 130A | DQA1*01:03 | High | |
| DQA1*02:01, DQB1*04:01 DQA1*02:01, DQB1*04:02 DQA1*03:03, DQB1*04:01 DQA1*04:01, DQB1*04:02 | 23L | DQB1*04:01 | High |
| 56L | Whole DQB1 group | High | |
| DQA1*02:01, DQB1*03:01 DQA1*02:01, DQB1*03:02 DQA1*02:01, DQB1*03:03 DQA1*03:01, DQB1*03:01 DQA1*03:01, DQB1*03:02 DQA1*03:01, DQB1*03:03 DQA1*03:02, DQB1*03:02 DQA1*03:02, DQB1*03:03 DQA1*05:03, DQB1*03:01 DQA1*05:05, DQB1*03:01 DQA1*06:01, DQB1*03:01 | 45EV | DQB1*03:01 | High |
| 55PP | Whole DQB1 group | High | |
| 55PPA | DQB1*03:02 | High | |
| 55PPD | DQB1*03:01, 03:03 | High | |
| 160S | DQA1*05:03 | High | |
| DQA1*02:01, DQB1*02:01 DQA1*02:01, DQB1*02:02 DQA1*03:01, DQB1*02:01 DQA1*04:01, DQB1*02:01 DQA1*05:01, DQB1*02:01 | 52LL | Whole DQB1 group | High |
| 135G | DQB1*02:02 | High |
| HLA-DP Eplets Recognized by Antibody Groups in PCA | Location of Dendrogram Antibody Groups that Appear to Recognize the Corresponding Eplets | HLA-DP Alleles where the Eplets are Expressed | ElliPro Score | |
|---|---|---|---|---|
| DPA1*01:03, DPB1*02:01 DPA1*01:03, DPB1*04:02 DPA1*01:03, DPB1*28:01 DPA1*01:04, DPB1*18:01 DPA1*01:05, DPB1*18:01 DPA1*01:05, DPB1*28:01 DPA1*01:03, DPB1*04:01 DPA1*01:03, DPB1*23:01 | 84GGPM | Upper | DPB1*02:01 DPB1*04:01 DPB1*04:02 DPB1*23:01 | High |
| 178M | Upper | DPB1*04:02 | High | |
| DPA1*01:03, DPB1*01:01 DPA1*01:03, DPB1*03:01 DPA1*01:03, DPB1*06:01 DPA1*01:03, DPB1*11:01 DPA1*01:03, DPB1*19:01 DPA1*01:05, DPB1*03:01 DPA1*02:01, DPB1*01:01 DPA1*02:01, DPB1*03:01 DPA1*02:01, DPB1*05:01 DPA1*02:01, DPB1*06:01 DPA1*02:01, DPB1*09:01 DPA1*02:01, DPB1*10:01 DPA1*02:01, DPB1*13:01 DPA1*02:01, DPB1*14:01 DPA1*02:01, DPB1*15:01* DPA1*02:01, DPB1*18:01* DPA1*02:01, DPB1*17:01 DPA1*02:02, DPB1*05:01 DPA1*02:02, DPB1*10:01 DPA1*02:02, DPB1*11:01 DPA1*02:02, DPB1*13:01 DPA1*03:01, DPB1*13:01 DPA1*03:01, DPB1*20:01 DPA1*04:01, DPB1*28:01* | 9H | Left | DPB1*09:01 DPB1*10:01 DPB1*14:01 DPB1*17:01 | Low |
| 9YL | Both | DPB1*03:01 DPB1*06:01 DPB1*11:01 DPB1*13:01 DPB1*20:01 | Low | |
| 11L | Both | DPB1*03:01 DPB1*06:01 DPB1*09:01 DPB1*10:01 DPB1*11:01 DPB1*13:01 DPB1*14:01 DPB1*17:01 DPB1*20:01 | Very Low | |
| 33EYA | Right | DPB1*01:01 DPB1*13:01 | High | |
| 35LV | Right | DPB1*05:01 | High | |
| 35YA | Right | DPB1*01:01 DPB1*11:01 DPB1*13:01 DPB1*15:01 | High | |
| 55EAE | Right | DPB1*05:01 DPB1*19:01 | High | |
| 57D | Left | DPB1*03:01 DPB1*06:01 DPB1*09:01 DPB1*14:01 DPB1*17:01 DPB1*20:01 | High | |
| 65LE | Left | DPB1*06:01 | High | |
| 69R | Right | DPB1*11:01 DPB1*15:01 | High | |
| 76I | Right | DPB1*13:01 DPB1*19:01 | Intermediate | |
| 76V | Both | DPB1*01:01 DPB1*03:01 DPB1*09:01 DPB1*10:01 DPB1*14:01 | Intermediate | |
| 84DEAV | Both | DPB1*01:01 DPB1*03:01 DPB1*05:01 DPB1*06:01 DPB1*09:01 DPB1*10:01 DPB1*11:01 DPB1*13:01 DPB1*14:01 DPB1*17:01 DPB1*20:01 | High | |
| 11M | Both | DPA1*02:02 DPA1*03:01 | Very low | |
| 31Q | Both | DPA1*02:01 DPA1*02:02 | Very low | |
| 50R | Both | DPA1*02:01 DPA1*02:02 DPA1*04:01 | High | |
| 66S | Both | DPA1*03:01 | High | |
| 111R | Both | DPA1*02:01 DPA1*02:02 | Low | |
| 127P | Both | DPA1*02:01 DPA1*02:02 DPA1*04:01 | High | |
| 160V | Both | DPA1*02:01 DPA1*02:02 DPA1*04:01 | High | |
| 190A | Both | DPA1*04:01 | High |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fylaktou, A.; Lioulios, G.; Tarassi, K.; Siorenta, A.; Petasis, G.C.; Gerogiannis, D.; Theodorou, I.; Iniotaki, A.G.; Vittoraki, A.G. An Approach to Identify HLA Class II Immunogenic Epitopes in the Greek Population through Machine Learning Algorithms. J. Clin. Med. 2022, 11, 7046. https://doi.org/10.3390/jcm11237046
Fylaktou A, Lioulios G, Tarassi K, Siorenta A, Petasis GC, Gerogiannis D, Theodorou I, Iniotaki AG, Vittoraki AG. An Approach to Identify HLA Class II Immunogenic Epitopes in the Greek Population through Machine Learning Algorithms. Journal of Clinical Medicine. 2022; 11(23):7046. https://doi.org/10.3390/jcm11237046
Chicago/Turabian StyleFylaktou, Asimina, Georgios Lioulios, Katerina Tarassi, Alexandra Siorenta, George Ch Petasis, Demetris Gerogiannis, Ioannis Theodorou, Aliki G. Iniotaki, and Angeliki G. Vittoraki. 2022. "An Approach to Identify HLA Class II Immunogenic Epitopes in the Greek Population through Machine Learning Algorithms" Journal of Clinical Medicine 11, no. 23: 7046. https://doi.org/10.3390/jcm11237046