
miRNA Regulatory Networks Associated with Peripheral Vascular Diseases

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. miRNA Expression Datasets
2.3. Data Analysis
3. Results
3.1. The Study Group Characteristics
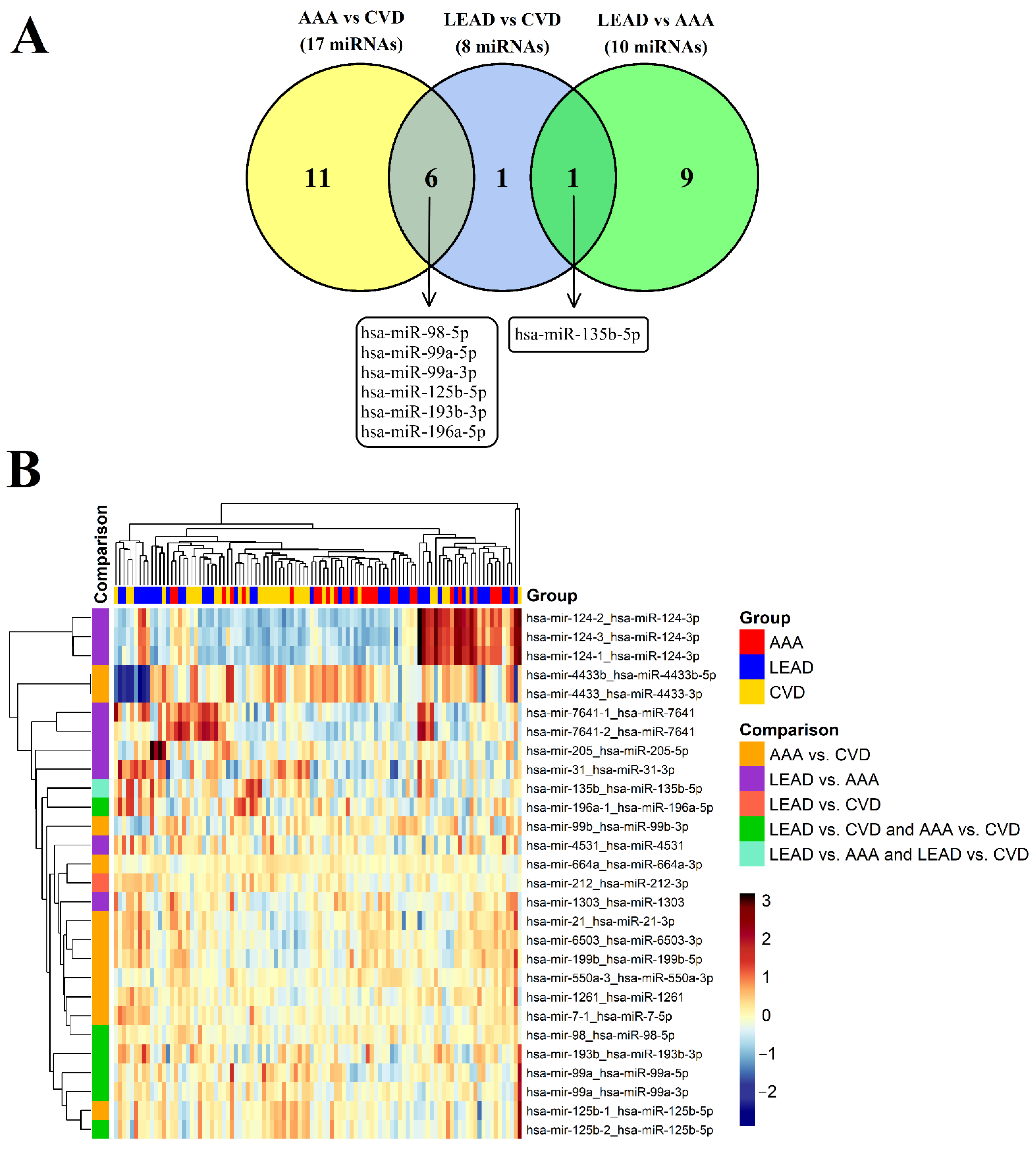
3.2. Identification of Differentially Expressed miRNAs between the LEAD, AAA, and CVD Groups
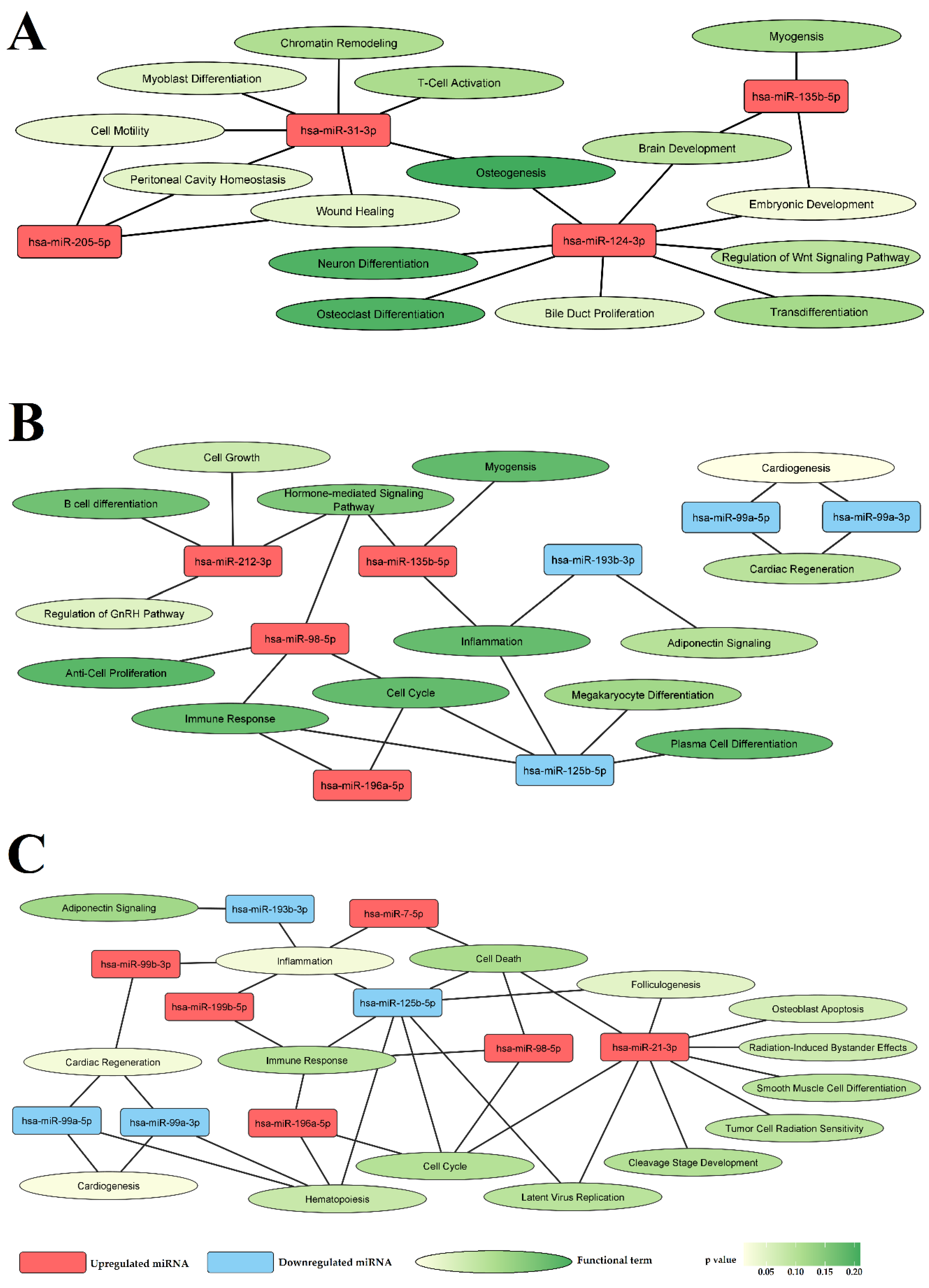
3.3. Selected miRNAs Are Closely Related to Vascular Pathology
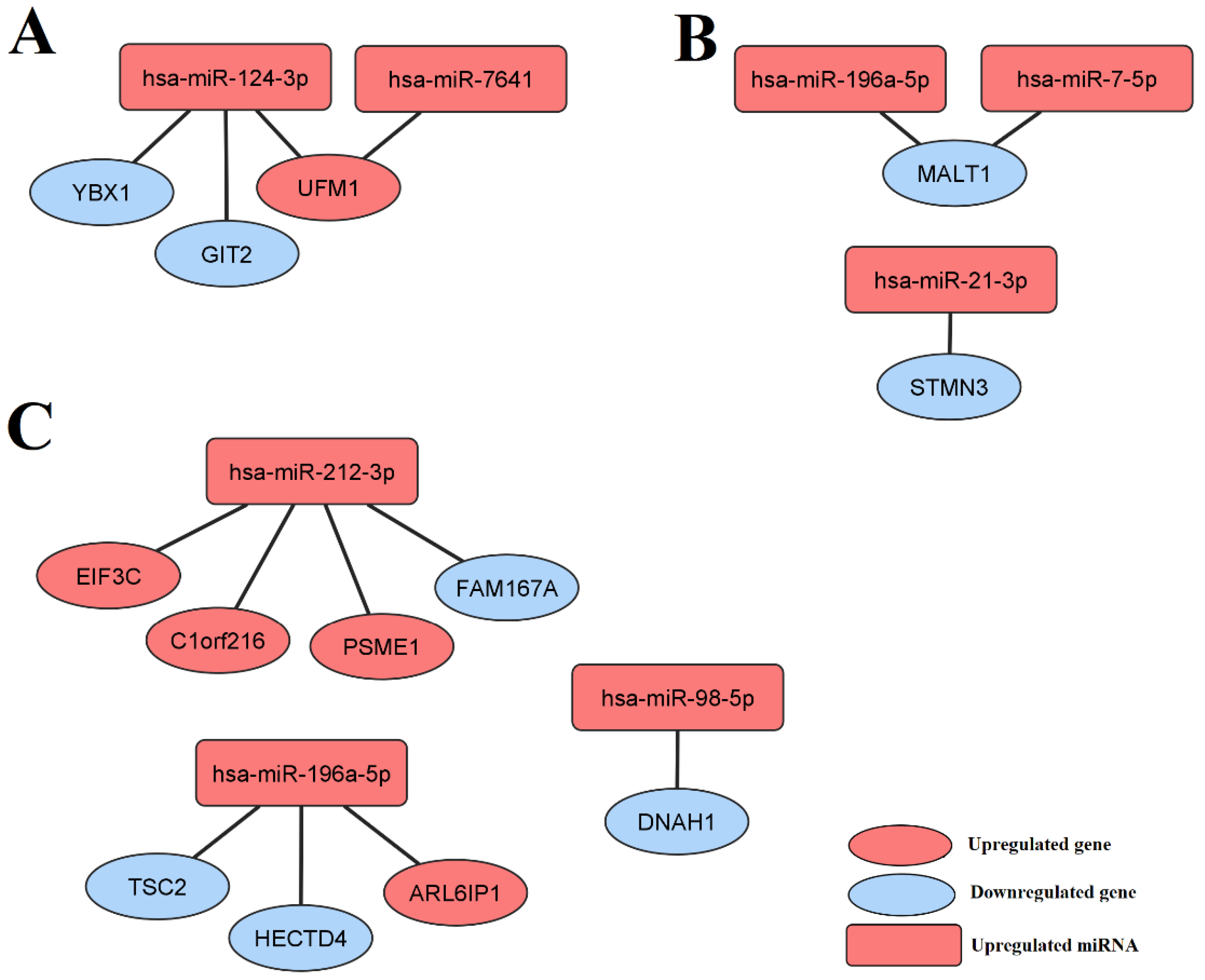
3.4. Selected miRNAs Regulate Genes Differentially Expressed between the LEAD, AAA, and CVD Groups
3.5. Some of the Selected miRNAs Are Associated with the Clinical and Demographic Characteristics of the Study Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, D.; Sung, Y.M.; Park, J.; Kim, S.; Kim, J.; Park, J.; Ha, H.; Bae, J.Y.; Kim, S.; Baek, D. General Rules for Functional MicroRNA Targeting. Nat. Genet. 2016, 48, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorshid, H.R.K.; Fard, S.S.; Ghaffari, S.H. An Overview of MicroRNAs: Biology, Functions, Therapeutics, and Analysis Methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA Therapeutics: Towards a New Era for the Management of Cancer and Other Diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Kalayinia, S.; Arjmand, F.; Maleki, M.; Malakootian, M.; Singh, C.P. MicroRNAs: Roles in Cardiovascular Development and Disease. Cardiovasc. Pathol. 2021, 50, 107296. [Google Scholar] [CrossRef] [PubMed]
- Munjas, J.; Sopić, M.; Stefanović, A.; Košir, R.; Ninić, A.; Joksić, I.; Antonić, T.; Spasojević-Kalimanovska, V.; Prosenc Zmrzljak, U. Non-Coding RNAs in Preeclampsia—Molecular Mechanisms and Diagnostic Potential. Int. J. Mol. Sci. 2021, 22, 10652. [Google Scholar] [CrossRef] [PubMed]
- Aboyans, V.; Ricco, J.-B.; Bartelink, M.-L.E.L.; Björck, M.; Brodmann, M.; Cohnert, T.; Collet, J.-P.; Czerny, M.; De Carlo, M.; Debus, S.; et al. 2017 ESC Guidelines on the Diagnosis and Treatment of Peripheral Arterial Diseases, in Collaboration with the European Society for Vascular Surgery (ESVS). Eur. Heart J. 2018, 39, 763–816. [Google Scholar] [CrossRef] [Green Version]
- Hamburg, N.M.; Creager, M.A. Pathophysiology of Intermittent Claudication in Peripheral Artery Disease. Circ. J. 2017, 81, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Song, P.; Rudan, D.; Zhu, Y.; Fowkes, F.J.I.; Rahimi, K.; Fowkes, F.G.R.; Rudan, I. Global, Regional, and National Prevalence and Risk Factors for Peripheral Artery Disease in 2015: An Updated Systematic Review and Analysis. Lancet Glob. Health 2019, 7, e1020–e1030. [Google Scholar] [CrossRef] [Green Version]
- Shu, J.; Santulli, G. Update on Peripheral Artery Disease: Epidemiology and Evidence-Based Facts. Atherosclerosis 2018, 275, 379–381. [Google Scholar] [CrossRef]
- Nehler, M.R.; Duval, S.; Diao, L.; Annex, B.H.; Hiatt, W.R.; Rogers, K.; Zakharyan, A.; Hirsch, A.T. Epidemiology of Peripheral Arterial Disease and Critical Limb Ischemia in an Insured National Population. J. Vasc. Surg. 2014, 60, 686–695.e2. [Google Scholar] [CrossRef] [Green Version]
- Chaikof, E.L.; Dalman, R.L.; Eskandari, M.K.; Jackson, B.M.; Lee, W.A.; Mansour, M.A.; Mastracci, T.M.; Mell, M.; Murad, M.H.; Nguyen, L.L.; et al. The Society for Vascular Surgery Practice Guidelines on the Care of Patients with an Abdominal Aortic Aneurysm. J. Vasc. Surg. 2018, 67, 2–77.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattanzi, S. Abdominal Aortic Aneurysms: Pathophysiology and Clinical Issues. J. Intern. Med. 2020, 288, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Kent, K.C.; Zwolak, R.M.; Egorova, N.N.; Riles, T.S.; Manganaro, A.; Moskowitz, A.J.; Gelijns, A.C.; Greco, G. Analysis of Risk Factors for Abdominal Aortic Aneurysm in a Cohort of More than 3 Million Individuals. J. Vasc. Surg. 2010, 52, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, R.; Ghoorah, K.; Kunadian, V. Abdominal Aortic Aneurysms and Risk Factors for Adverse Events. Cardiol. Rev. 2016, 24, 88–93. [Google Scholar] [CrossRef]
- Murali Krishna, S.; Morton, S.K.; Li, J.; Golledge, J. Risk Factors and Mouse Models of Abdominal Aortic Aneurysm Rupture. Int. J. Mol. Sci. 2020, 21, 7250. [Google Scholar] [CrossRef]
- Eberhardt, R.T.; Raffetto, J.D. Chronic Venous Insufficiency. Circulation 2014, 130, 333–346. [Google Scholar] [CrossRef]
- Ligi, D.; Croce, L.; Mannello, F. Chronic Venous Disorders: The Dangerous, the Good, and the Diverse. Int. J. Mol. Sci. 2018, 19, 2544. [Google Scholar] [CrossRef] [Green Version]
- Nicolaides, A.; Kakkos, S.; Baekgaard, N.; Comerota, A.; de Maeseneer, M.; Eklof, B.; Giannoukas, A.D.; Lugli, M.; Maleti, O.; Myers, K.; et al. Management of Chronic Venous Disorders of the Lower Limbs. Guidelines According to Scientific Evidence. Part I. Int. Angiol. 2018, 37, 181–254. [Google Scholar] [CrossRef]
- Nicolaides, A.; Kakkos, S.; Baekgaard, N.; Comerota, A.; de Maeseneer, M.; Eklof, B.; Giannoukas, A.; Lugli, M.; Maleti, O.; Mansilha, A.; et al. Management of Chronic Venous Disorders of the Lower Limbs. Guidelines According to Scientific Evidence. Part II. Int. Angiol. 2020, 39, 175–240. [Google Scholar] [CrossRef]
- Mansilha, A.; Sousa, J. Pathophysiological Mechanisms of Chronic Venous Disease and Implications for Venoactive Drug Therapy. Int. J. Mol. Sci. 2018, 19, 1669. [Google Scholar] [CrossRef] [Green Version]
- Lurie, F.; Passman, M.; Meisner, M.; Dalsing, M.; Masuda, E.; Welch, H.; Bush, R.L.; Blebea, J.; Carpentier, P.H.; De Maeseneer, M.; et al. The 2020 Update of the CEAP Classification System and Reporting Standards. J. Vasc. Surg. Venous Lymphat. Disord. 2020, 8, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Vuylsteke, M.E.; Colman, R.; Thomis, S.; Guillaume, G.; Van Quickenborne, D.; Staelens, I. An Epidemiological Survey of Venous Disease Among General Practitioner Attendees in Different Geographical Regions on the Globe: The Final Results of the Vein Consult Program. Angiology 2018, 69, 779–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conte, M.S.; Bradbury, A.W.; Kolh, P.; White, J.V.; Dick, F.; Fitridge, R.; Mills, J.L.; Ricco, J.-B.; Suresh, K.R.; Murad, M.H.; et al. Global Vascular Guidelines on the Management of Chronic Limb-Threatening Ischemia. Eur. J. Vasc. Endovasc. Surg. 2019, 58, S1–S109.e33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittens, C.; Davies, A.H.; Bækgaard, N.; Broholm, R.; Cavezzi, A.; Chastanet, S.; de Wolf, M.; Eggen, C.; Giannoukas, A.; Gohel, M.; et al. Editor’s Choice—Management of Chronic Venous Disease: Clinical Practice Guidelines of the European Society for Vascular Surgery (ESVS). Eur. J. Vasc. Endovasc. Surg. 2015, 49, 678–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Zhang, C. MicroRNAs in Vascular Disease. J. Cardiovasc. Pharmacol. 2011, 57, 8–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Hernando, C.; Suárez, Y. MicroRNAs in Endothelial Cell Homeostasis and Vascular Disease. Curr. Opin. Hematol. 2018, 25, 227–236. [Google Scholar] [CrossRef]
- Ono, K.; Horie, T.; Baba, O.; Kimura, M.; Tsuji, S.; Rodriguez, R.R.; Miyagawa, S.; Kimura, T. Functional Non-Coding RNAs in Vascular Diseases. FEBS J. 2020, 288, 6315–6330. [Google Scholar] [CrossRef]
- Kumar, S.; Williams, D.; Sur, S.; Wang, J.-Y.; Jo, H. Role of Flow-Sensitive MicroRNAs and Long Noncoding RNAs in Vascular Dysfunction and Atherosclerosis. Vascul. Pharmacol. 2019, 114, 76–92. [Google Scholar] [CrossRef]
- Vogiatzi, G.; Oikonomou, E.; Deftereos, S.; Siasos, G.; Tousoulis, D. Peripheral Artery Disease: A Micro-RNA-Related Condition? Curr. Opin. Pharmacol. 2018, 39, 105–112. [Google Scholar] [CrossRef]
- Hamburg, N.M.; Leeper, N.J. Therapeutic Potential of Modulating MicroRNA in Peripheral Artery Disease. Curr. Vasc. Pharmacol. 2015, 13, 316–323. [Google Scholar] [CrossRef]
- Iyer, V.; Rowbotham, S.; Biros, E.; Bingley, J.; Golledge, J. A Systematic Review Investigating the Association of MicroRNAs with Human Abdominal Aortic Aneurysms. Atherosclerosis 2017, 261, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Liu, G.; Huang, Y.; Lu, X.; Lu, M.; Huang, X.; Li, W.; Jiang, M. MicroRNA Profiling in Great Saphenous Vein Tissues of Patients with Chronic Venous Insufficiency. Tohoku J. Exp. Med. 2012, 228, 341–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Liu, Z.; Shen, L.; Jin, Y.; Xu, G.; Zhang, Z.; Fang, C.; Guan, W.; Liu, C. Augmentation of MiR-202 in Varicose Veins Modulates Phenotypic Transition of Vascular Smooth Muscle Cells by Targeting Proliferator–Activated Receptor-γ Coactivator-1α. J. Cell. Biochem. 2019, 120, 10031–10042. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.A.; Adesina-Georgiadis, K.N.; Spagou, K.; Vorkas, P.A.; Li, J.V.; Shalhoub, J.; Holmes, E.; Davies, A.H. A Comprehensive Characterisation of the Metabolic Profile of Varicose Veins; Implications in Elaborating Plausible Cellular Pathways for Disease Pathogenesis. Sci. Rep. 2017, 7, 2989. [Google Scholar] [CrossRef]
- Saenz-Pipaon, G.; Martinez-Aguilar, E.; Orbe, J.; González Miqueo, A.; Fernandez-Alonso, L.; Paramo, J.A.; Roncal, C. The Role of Circulating Biomarkers in Peripheral Arterial Disease. Int. J. Mol. Sci. 2021, 22, 3601. [Google Scholar] [CrossRef]
- Barbalata, T.; Moraru, O.E.; Stancu, C.S.; Devaux, Y.; Simionescu, M.; Sima, A.V.; Niculescu, L.S. Increased MiR-142 Levels in Plasma and Atherosclerotic Plaques from Peripheral Artery Disease Patients with Post-Surgery Cardiovascular Events. Int. J. Mol. Sci. 2020, 21, 9600. [Google Scholar] [CrossRef]
- Plana, E.; Gálvez, L.; Medina, P.; Navarro, S.; Fornés-Ferrer, V.; Panadero, J.; Miralles, M. Identification of Novel MicroRNA Profiles Dysregulated in Plasma and Tissue of Abdominal Aortic Aneurysm Patients. Int. J. Mol. Sci. 2020, 21, 4600. [Google Scholar] [CrossRef]
- Torres-Do Rego, A.; Barrientos, M.; Ortega-Hernández, A.; Modrego, J.; Gómez-Gordo, R.; Álvarez-Sala, L.A.; Cachofeiro, V.; Gómez-Garre, D. Identification of a Plasma Microrna Signature as Biomarker of Subaneurysmal Aortic Dilation in Patients with High Cardiovascular Risk. J. Clin. Med. 2020, 9, 2783. [Google Scholar] [CrossRef]
- Nana, P.; Dakis, K.; Brodis, A.; Spanos, K.; Kouvelos, G. Circulating Biomarkers for the Prediction of Abdominal Aortic Aneurysm Growth. J. Clin. Med. 2021, 10, 1718. [Google Scholar] [CrossRef]
- Zalewski, D.P.; Ruszel, K.P.; Stępniewski, A.; Gałkowski, D.; Bogucki, J.; Komsta, Ł.; Kołodziej, P.; Chmiel, P.; Zubilewicz, T.; Feldo, M.; et al. Dysregulations of MicroRNA and Gene Expression in Chronic Venous Disease. J. Clin. Med. 2020, 9, 1251. [Google Scholar] [CrossRef]
- Bogucka-Kocka, A.; Zalewski, D.P.; Ruszel, K.P.; Stępniewski, A.; Gałkowski, D.; Bogucki, J.; Komsta, Ł.; Kołodziej, P.; Zubilewicz, T.; Feldo, M.; et al. Dysregulation of MicroRNA Regulatory Network in Lower Extremities Arterial Disease. Front. Genet. 2019, 10, 1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalewski, D.P.; Ruszel, K.P.; Stępniewski, A.; Gałkowski, D.; Bogucki, J.; Komsta, Ł.; Kołodziej, P.; Chmiel, P.; Zubilewicz, T.; Feldo, M.; et al. Dysregulation of MicroRNA Modulatory Network in Abdominal Aortic Aneurysm. J. Clin. Med. 2020, 9, 1974. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.-C.; Müller, M. PROC: An Open-Source Package for R and S+ to Analyze and Compare ROC Curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. MiRNet 2.0: Network-Based Visual Analytics for MiRNA Functional Analysis and Systems Biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Zalewski, D.P.; Ruszel, K.P.; Stępniewski, A.; Gałkowski, D.; Bogucki, J.; Kołodziej, P.; Szymańska, J.; Płachno, B.J.; Zubilewicz, T.; Feldo, M.; et al. Identification of Transcriptomic Differences between Lower Extremities Arterial Disease, Abdominal Aortic Aneurysm and Chronic Venous Disease in Peripheral Blood Mononuclear Cells Specimens. Int. J. Mol. Sci. 2021, 22, 3200. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Asadi-Yousefabad, S.-L.; Nammian, P.; Tabei, S.M.B.; Daneshi, S.; Fallahi, J.; Razban, V.; Sheikhha, M.H. Angiogenesis in Diabetic Mouse Model with Critical Limb Ischemia; Cell and Gene Therapy. Microvasc. Res. 2022, 141, 104339. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Lin, S.-J.; Wu, T.-C. MiR-548j-5p Regulates Angiogenesis in Peripheral Artery Disease. Sci. Rep. 2022, 12, 838. [Google Scholar] [CrossRef]
- Lamin, V.; Verry, J.; Eigner-Bybee, I.; Fuqua, J.D.; Wong, T.; Lira, V.A.; Dokun, A.O. Modulation of MiR-29a and ADAM12 Reduces Post-Ischemic Skeletal Muscle Injury and Improves Perfusion Recovery and Skeletal Muscle Function in a Mouse Model of Type 2 Diabetes and Peripheral Artery Disease. Int. J. Mol. Sci. 2022, 23, 429. [Google Scholar] [CrossRef] [PubMed]
- Theofilis, P.; Oikonomou, E.; Vogiatzi, G.; Antonopoulos, A.S.; Siasos, G.; Iliopoulos, D.C.; Perrea, D.; Tsioufis, C.; Tousoulis, D. The Impact of Proangiogenic MicroRNA Modulation on Blood Flow Recovery Following Hind Limb Ischemia. A Systematic Review and Meta-Analysis of Animal Studies. Vascul. Pharmacol. 2021, 141, 106906. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, W.; Zhang, L.; Wang, L.; Li, J.; Shu, C.; Li, X. Roles of MicroRNAs in Peripheral Artery In-Stent Restenosis after Endovascular Treatment. BioMed Res. Int. 2021, 2021, e9935671. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ni, Y.-Q.; Liu, Y.-S. Mechanisms of Action of MiRNAs and LncRNAs in Extracellular Vesicle in Atherosclerosis. Front. Cardiovasc. Med. 2021, 8, 733985. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, L.; Liu, Y.; Zeng, W.; Wang, K. Potency of MiR-144-3p in Promoting Abdominal Aortic Aneurysm Progression in Mice Correlates with Apoptosis of Smooth Muscle Cells. Vascul. Pharmacol. 2022, 142, 106901. [Google Scholar] [CrossRef] [PubMed]
- Si, X.; Chen, Q.; Zhang, J.; Zhou, W.; Chen, L.; Chen, J.; Deng, N.; Li, W.; Liu, D.; Wang, L.; et al. MicroRNA-23b Prevents Aortic Aneurysm Formation by Inhibiting Smooth Muscle Cell Phenotypic Switching via FoxO4 Suppression. Life Sci. 2022, 288, 119092. [Google Scholar] [CrossRef]
- Huang, T.; Liu, S.; Liu, R.; Pan, B.; Wang, W. Inhibition of MiR-188-5p Suppresses Progression of Experimental Abdominal Aortic Aneurysms. J. Cardiovasc. Pharmacol. 2021, 77, 107–114. [Google Scholar] [CrossRef]
- Cao, Y.; Cao, Z.; Wang, W.; Jie, X.; Li, L.; Cao, Y.; Cao, Z.; Wang, W.; Jie, X.; Li, L. MicroRNA-199a-5p Regulates FOXC2 to Control Human Vascular Smooth Muscle Cell Phenotypic Switch. Mol. Med. Rep. 2021, 24, 627. [Google Scholar] [CrossRef]
- Wu, J.; Li, X.; Li, D.; Ren, X.; Li, Y.; Herter, E.K.; Qian, M.; Toma, M.-A.; Wintler, A.-M.; Sérézal, I.G.; et al. MicroRNA-34 Family Enhances Wound Inflammation by Targeting LGR4. J. Investig. Dermatol. 2020, 140, 465–476.e11. [Google Scholar] [CrossRef]
- Kansakar, U.; Varzideh, F.; Mone, P.; Jankauskas, S.S.; Santulli, G. Functional Role of MicroRNAs in Regulating Cardiomyocyte Death. Cells 2022, 11, 983. [Google Scholar] [CrossRef]
- Xu, Z.; Han, Y.; Liu, J.; Jiang, F.; Hu, H.; Wang, Y.; Liu, Q.; Gong, Y.; Li, X. MiR-135b-5p and MiR-499a-3p Promote Cell Proliferation and Migration in Atherosclerosis by Directly Targeting MEF2C. Sci. Rep. 2015, 5, 12276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Guan, Y.; Liu, B.; Lin, Y.; Yan, Y.; Wang, H.; Wang, H.; Jing, B. MicroRNA-99a-5p Alleviates Atherosclerosis via Regulating Homeobox A1. Life Sci. 2019, 232, 116664. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.-G.; Guo, W.-P.; Tang, Z.-Y.; Li, S.-H.; Wu, H.-Y.; Zhang, L.-C. Expression Level and Prospective Mechanism of MiRNA-99a-3p in Head and Neck Squamous Cell Carcinoma Based on MiRNA-Chip and MiRNA-Sequencing Data in 1, 167 Cases. Pathol. Res. Pract. 2019, 215, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Shu, X.; Sun, L.; Yu, L.; Sun, L.; Yang, Z.; Ran, Y. MiR-196a-5p Modulates Gastric Cancer Stem Cell Characteristics by Targeting Smad4. Int. J. Oncol. 2017, 50, 1965–1976. [Google Scholar] [CrossRef] [Green Version]
- Xin, H.; Wang, C.; Liu, Z. MiR-196a-5p Promotes Metastasis of Colorectal Cancer via Targeting IκBα. BMC Cancer 2019, 19, 30. [Google Scholar] [CrossRef]
- Li, X.; Chen, Z.; Ni, Y.; Bian, C.; Huang, J.; Chen, L.; Xie, X.; Wang, J. Tumor-Associated Macrophages Secret Exosomal MiR-155 and MiR-196a-5p to Promote Metastasis of Non-Small-Cell Lung Cancer. Transl Lung Cancer Res. 2021, 10, 1338–1354. [Google Scholar] [CrossRef]
- Huang, J.; Manning, B.D. A Complex Interplay between Akt, TSC2 and the Two MTOR Complexes. Biochem. Soc. Trans. 2009, 37, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Motoshima, H.; Goldstein, B.J.; Igata, M.; Araki, E. AMPK and Cell Proliferation—AMPK as a Therapeutic Target for Atherosclerosis and Cancer. J. Physiol. 2006, 574, 63–71. [Google Scholar] [CrossRef]
- McAllister-Lucas, L.M.; Jin, X.; Gu, S.; Siu, K.; McDonnell, S.; Ruland, J.; Delekta, P.C.; Van Beek, M.; Lucas, P.C. The CARMA3-Bcl10-MALT1 Signalosome Promotes Angiotensin II-Dependent Vascular Inflammation and Atherogenesis. J. Biol. Chem. 2010, 285, 25880–25884. [Google Scholar] [CrossRef] [Green Version]
- Birch, C.A.; Molinar-Inglis, O.; Trejo, J. Subcellular Hot Spots of GPCR Signaling Promote Vascular Inflammation. Curr. Opin. Endocr. Metab. Res. 2021, 16, 37–42. [Google Scholar] [CrossRef]
- Lee, H.-Y.; Chen, C.-K.; Ho, C.-M.; Lee, S.-S.; Chang, C.-Y.; Chen, K.-J.; Jou, Y.-S. EIF3C-Enhanced Exosome Secretion Promotes Angiogenesis and Tumorigenesis of Human Hepatocellular Carcinoma. Oncotarget 2018, 9, 13193–13205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Li, S.; Chen, D.; Chen, B.; Yu, T.; Zhao, F.; Wang, Q.; Yao, M.; Huang, S.; Chen, Z.; et al. Transcriptomic Analyses of RNA-Binding Proteins Reveal EIF3c Promotes Cell Proliferation in Hepatocellular Carcinoma. Cancer Sci. 2017, 108, 877–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Xu, X.; Luan, P.; Kou, W.; Li, M.; Yu, Q.; Zhuang, J.; Xu, Y.; Peng, W.; Jian, W. MiR-124-3p Regulates Angiogenesis in Peripheral Arterial Disease by Targeting STAT3. Mol. Med. Rep. 2020, 22, 4890–4898. [Google Scholar] [CrossRef] [PubMed]
- Van Gastel, J.; Boddaert, J.; Jushaj, A.; Premont, R.T.; Luttrell, L.M.; Janssens, J.; Martin, B.; Maudsley, S. GIT2—A Keystone in Ageing and Age-Related Disease. Ageing Res. Rev. 2018, 43, 46–63. [Google Scholar] [CrossRef]
- He, X.; Wang, S.; Li, M.; Zhong, L.; Zheng, H.; Sun, Y.; Lai, Y.; Chen, X.; Wei, G.; Si, X.; et al. Long Noncoding RNA GAS5 Induces Abdominal Aortic Aneurysm Formation by Promoting Smooth Muscle Apoptosis. Theranostics 2019, 9, 5558–5576. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Patients with LEAD (n = 40) | Patients with AAA (n = 28) | Patients with CVD (n = 34) | p1 |
|---|---|---|---|---|
| Age | 57.6 ± 9.82 56.5 (43–71) | 66.4 ± 4.52 67 (57–76) | 44.1 ± 10.07 42 (27–78) | 9.966 × 10−12 |
| Sex males/females | 35 (87.5%)/5 (12.5%) | 25 (89.3%)/ 3 (10.7%) | 17 (50%)/ 17 (50%) | 2.180 × 10−4 |
| Body mass index (BMI) | 27.2 ± 2.62 27.3 (21.9–31.6) | 25.1 ± 3.30 25.2 (18.0–31.3) | 23.9 ± 2.35 24.0 (20.1–28.8) | 1.670 × 10−5 |
| Risk factors and cardiovascular comorbidities | ||||
| Smoking never/former/current | 0 (0%)/18 (45%)/ 22 (55%) | 9 (32.1%)/ 9 (32.1%)/10 (35.7%) | 16 (47%)/13 (38%)/ 5 (14.7%) | 2.369 × 10−5 |
| Diabetes type 2 | 5 (12.5%) | 6 (21.4%) | 0 (0%) | 9.309 × 10−3 |
| Hypertension | 36 (90%) | 19 (67.9%) | 0 (0%) | 1.066 × 10−17 |
| Coronary artery disease (CAD) | 11 (27.5%) | 7 (25%) | 0 (0%) | 8.508 × 10−4 |
| Myocardial infarction | 8 (20%) | 3 (10.7%) | 0 (0%) | 0.011 |
| Stroke/transient ischemic attack | 2 (5%) | 1 (3.6%) | 0 (0%) | 0.493 |
| Hematological and biochemical blood parameters | ||||
| Red blood cells (M/µL) | 4.74 ± 0.30 4.75 (4.11–5.18) | 4.94 ± 0.21 4.99 (4.56–5.50) | 4.78 ± 0.34 4.83 (3.78–5.50) | 0.017 |
| White blood cells (K/µL) | 5.49 ± 0.69 5.27 (4.45–6.89) | 5.66 ± 0.70 5.78 (4.44–6.90) | 5.76 ± 0.71 5.78 (4.67–6.90) | 0.361 |
| Lymphocytes (K/µL) | 3.04 ± 0.54 3.03 (2.01–3.99) | 2.99 ± 0.44 3.02 (2.05–3.78) | 2.99 ± 0.45 3.00 (2.04–3.99) | 0.771 |
| Monocytes (K/µL) | 0.47 ± 0.15 0.41 (0.22–0.87) | 0.39 ± 0.12 0.33 (0.26–0.67) | 0.33 ± 0.07 0.32 (0.19–0.56) | 9.727 × 10−6 |
| Neutrophils (K/µL) | 4.21 ± 0.47 4.07 (3.51–5.21) | 4.17 ± 0.43 4.20 (3.33–4.78) | 4.17 ± 0.62 4.31 (2.12–4.99) | 0.809 |
| Eosinophils (K/µL) | 0.21 ± 0.09 0.20 (0.10–0.56) | 0.18 ± 0.05 0.18 (0.11–0.31) | 0.16 ± 0.04 0.17 (0.08–0.23) | 6.072 × 10−3 |
| Basophils (K/µL) | 0.10 ± 0.03 0.1 (0.07–0.19) | 0.11 ± 0.02 0.11 (0.08–0.20) | 0.10 ± 0.03 0.10 (0.02–0.20) | 3.955 × 10−3 |
| Platelets (K/µL) | 309.3 ± 75.7 295.5 (179–561) | 419.9 ± 124.0 404.5 (211–756) | 368.26 ± 104.15 365.5 (211–756) | 4.643 × 10−5 |
| Hemoglobin (g/dL) | 14.12 ± 0.52 14.01 (12.99–14.99) | 14.02 ± 0.51 13.99 (13.34–15.00) | 14.08 ± 0.48 14.06 (13.30–14.99) | 0.692 |
| Hematocrit (%) | 41.3 ± 1.42 41.5 (38.4–43.8) | 40.8 ± 1.30 41 (38–43) | 39.9 ± 1.63 40 (37–43) | 6.775 × 10−4 |
| Creatinine (mmol/L) | 78.7 ± 12.6 79 (56–99) | 54.2 ± 11.5 51.5 (39–87) | 58.2 ± 14.20 56 (37–93) | 2.086 × 10−10 |
| Urea (mmol/L) | 4.69 ± 0.83 4.79 (2.99–6.02) | 4.66 ± 0.67 4.69 (3.45–5.88) | 5.09 ± 0.86 4.94 (3.45–6.87) | 0.119 |
| Medication | ||||
| Statins | 34 (85%) | 13 (46.4%) | 0 (0%) | 2.090 × 10−13 |
| Acetylsalicylic acid | 40 (100%) | 27 (96.4%) | 0 (0%) | 7.961 × 10−26 |
| Clopidogrel | 8 (20%) | 3 (10.7%) | 0 (0%) | 6.382 × 10−3 |
| Beta-adrenergic blockers | 27 (67.5%) | 16 (57.1%) | 0 (0%) | 4.427 × 10−12 |
| Angiotensin-converting enzyme inhibitor | 20 (50%) | 4 (14.3%) | 0 (0%) | 6.266 × 10−7 |
| Ca2+ channel blockers | 11 (27.5%) | 2 (7.14%) | 0 (0%) | 5.431 × 10−4 |
| Fibrates | 5 (12.5%) | 2 (7.14%) | 0 (0%) | 0.092 |
| Metformin | 2 (5%) | 3 (10.7%) | 0 (0%) | 0.148 |
| Gliclazide | 4 (10%) | 4 (14.3%) | 0 (0%) | 0.055 |
| Micronized diosmin | 0 (0%) | 0 (0%) | 24 (70.6%) | 1.012 × 10−15 |
| Preparation with hesperidin, Ruscus aculeatus, and vitamin C | 0 (0%) | 0 (0%) | 15 (44.1%) | 4.550 × 10−8 |
| No. | miRNA Transcript | miRNA ID 1 | p | Fold Change | ROC-AUC |
|---|---|---|---|---|---|
| LEAD vs. AAA—Upregulated miRNAs | |||||
| 1. | hsa-mir-124-2_hsa-miR-124-3p | hsa-miR-124-3p | 6.715 × 10−4 | 7.441 | 0.585 |
| 2. | hsa-mir-124-1_hsa-miR-124-3p | hsa-miR-124-3p | 1.665 × 10−3 | 6.015 | 0.550 |
| 3. | hsa-mir-124-3_hsa-miR-124-3p | hsa-miR-124-3p | 1.946 × 10−3 | 6.455 | 0.562 |
| 4. | hsa-mir-7641-2_hsa-miR-7641 | hsa-miR-7641 | 7.019 × 10−3 | 3.398 | 0.646 |
| 5. | hsa-mir-205_hsa-miR-205-5p | hsa-miR-205-5p | 9.943 × 10−3 | 3.414 | 0.558 |
| 6. | hsa-mir-31_hsa-miR-31-3p | hsa-miR-31-3p | 1.066 × 10−2 | 2.040 | 0.749 |
| 7. | hsa-mir-135b_hsa-miR-135b-5p | hsa-miR-135b-5p | 1.121 × 10−2 | 2.565 | 0.656 |
| 8. | hsa-mir-7641-1_hsa-miR-7641 | hsa-miR-7641 | 2.886 × 10−2 | 2.231 | 0.629 |
| LEAD vs. AAA—Downregulated miRNAs | |||||
| 9. | hsa-mir-4531_hsa-miR-4531 | hsa-miR-4531 | 8.039 × 10−4 | 0.439 | 0.521 |
| 10. | hsa-mir-1303_hsa-miR-1303 | hsa-miR-1303 | 9.764 × 10−4 | 0.386 | 0.629 |
| LEAD vs. CVD—Upregulated miRNAs | |||||
| 1. | hsa-mir-196a-1_hsa-miR-196a-5p | hsa-miR-196a-5p | 7.766 × 10−3 | 14.106 | 0.603 |
| 2. | hsa-mir-98_hsa-miR-98-5p | hsa-miR-98-5p | 1.599 × 10−2 | 2.086 | 0.646 |
| 3. | hsa-mir-212_hsa-miR-212-3p | hsa-miR-212-3p | 3.163 × 10−2 | 2.339 | 0.558 |
| 4. | hsa-mir-135b_hsa-miR-135b-5p | hsa-miR-135b-5p | 3.597 × 10−2 | 7.939 | 0.524 |
| LEAD vs. CVD—Downregulated miRNAs | |||||
| 5. | hsa-mir-99a_hsa-miR-99a-5p | hsa-miR-99a-5p | 5.074 × 10−3 | 0.211 | 0.626 |
| 6. | hsa-mir-99a_hsa-miR-99a-3p | hsa-miR-99a-3p | 5.534 × 10−3 | 0.113 | 0.607 |
| 7. | hsa-mir-125b-2_hsa-miR-125b-5p | hsa-miR-125b-5p | 1.317 × 10−2 | 0.259 | 0.659 |
| 8. | hsa-mir-193b_hsa-miR-193b-3p | hsa-miR-193b-3p | 4.348 × 10−2 | 0.263 | 0.527 |
| AAA vs. CVD—Upregulated miRNAs | |||||
| 1. | hsa-mir-98_hsa-miR-98-5p | hsa-miR-98-5p | 5.135 × 10−3 | 2.227 | 0.674 |
| 2. | hsa-mir-196a-1_hsa-miR-196a-5p | hsa-miR-196a-5p | 8.853 × 10−3 | 11.438 | 0.516 |
| 3. | hsa-mir-199b_hsa-miR-199b-5p | hsa-miR-199b-5p | 1.133 × 10−2 | 2.608 | 0.750 |
| 4. | hsa-mir-7-1_hsa-miR-7-5p | hsa-miR-7-5p | 1.496 × 10−2 | 2.079 | 0.601 |
| 5. | hsa-mir-1261_hsa-miR-1261 | hsa-miR-1261 | 1.766 × 10−2 | 2.256 | 0.611 |
| 6. | hsa-mir-21_hsa-miR-21-3p | hsa-miR-21-3p | 3.115 × 10−2 | 2.567 | 0.798 |
| 7. | hsa-mir-99b_hsa-miR-99b-3p | hsa-miR-99b-3p | 3.642 × 10−2 | 3.652 | 0.564 |
| 8. | hsa-mir-6503_hsa-miR-6503-3p | hsa-miR-6503-3p | 4.350 × 10−2 | 2.313 | 0.733 |
| 9. | hsa-mir-550a-3_hsa-miR-550a-3p | hsa-miR-550a-3p | 4.373 × 10−2 | 2.026 | 0.693 |
| AAA vs. CVD—Downregulated miRNAs | |||||
| 10. | hsa-mir-664a_hsa-miR-664a-3p | hsa-miR-664a-3p | 1.218 × 10−4 | 0.499 | 0.824 |
| 11. | hsa-mir-125b-2_hsa-miR-125b-5p | hsa-miR-125b-5p | 8.773 × 10−4 | 0.182 | 0.619 |
| 12. | hsa-mir-99a_hsa-miR-99a-5p | hsa-miR-99a-5p | 1.189 × 10−3 | 0.185 | 0.608 |
| 13. | hsa-mir-99a_hsa-miR-99a-3p | hsa-miR-99a-3p | 1.251 × 10−3 | 0.092 | 0.640 |
| 14. | hsa-mir-125b-1_hsa-miR-125b-5p | hsa-miR-125b-5p | 6.648 × 10−3 | 0.247 | 0.598 |
| 15. | hsa-mir-4433b_hsa-miR-4433b-5p | hsa-miR-4433b-5p | 2.297 × 10−2 | 0.197 | 0.544 |
| 16. | hsa-mir-4433_hsa-miR-4433-3p | hsa-miR-4433a-3p | 2.297 × 10−2 | 0.197 | 0.544 |
| 17. | hsa-mir-193b_hsa-miR-193b-3p | hsa-miR-193b-3p | 4.728 × 10−2 | 0.292 | 0.450 |
| Characteristic | miRNA Transcript | miRNA ID 1 | p |
|---|---|---|---|
| Sex | hsa-mir-6503_hsa-miR-6503-3p | hsa-miR-6503-3p | 3.02 × 10−3 |
| hsa-mir-4433b_hsa-miR-4433b-5p | hsa-miR-4433b-5p | 1.17 × 10−2 | |
| hsa-mir-4433_hsa-miR-4433-3p | hsa-miR-4433a-3p | 1.17 × 10−2 | |
| hsa-mir-199b_hsa-miR-199b-5p | hsa-miR-199b-5p | 1.17 × 10−2 | |
| hsa-mir-21_hsa-miR-21-3p | hsa-miR-21-3p | 1.48 × 10−2 | |
| hsa-mir-1261_hsa-miR-1261 | hsa-miR-1261 | 2.21 × 10−2 | |
| Acetylsalicylic acid medication | hsa-mir-664a_hsa-miR-664a-3p | hsa-miR-664a-3p | 3.02 × 10−3 |
| hsa-mir-199b_hsa-miR-199b-5p | hsa-miR-199b-5p | 9.42 × 10−3 | |
| hsa-mir-6503_hsa-miR-6503-3p | hsa-miR-6503-3p | 1.17 × 10−2 | |
| hsa-mir-21_hsa-miR-21-3p | hsa-miR-21-3p | 1.48 × 10−2 | |
| Diosmin medication | hsa-mir-664a_hsa-miR-664a-3p | hsa-miR-664a-3p | 3.42 × 10−2 |
| hsa-mir-6503_hsa-miR-6503-3p | hsa-miR-6503-3p | 4.04 × 10−2 |
| Characteristic | miRNA Transcript | miRNA ID 1 | Regression Coefficient | p |
|---|---|---|---|---|
| Monocyte counts | hsa-mir-99b_hsa-miR-99b-3p 4 | hsa-miR-99b-3p | −1.38 | 6.07 × 10−3 |
| hsa-mir-4433b_hsa-miR-4433b-5p 4 | hsa-miR-4433b-5p | −2.12 | 6.03 × 10−3 | |
| hsa-mir-4433_hsa-miR-4433-3p 4 | hsa-miR-4433a-3p | −2.12 | 6.03 × 10−3 | |
| hsa-mir-31_hsa-miR-31-3p 2 | hsa-miR-31-3p | 2.23 | 2.01 × 10−3 | |
| hsa-mir-7-1_hsa-miR-7-5p 4 | hsa-miR-7-5p | 0.92 | 1.44 × 10−3 | |
| Creatinine | hsa-mir-125b-1_hsa-miR-125b-5p 4 | hsa-miR-125b-5p | −0.01 | 2.02 × 10−3 |
| hsa-mir-125b-2_hsa-miR-125b-5p 3,4 | hsa-miR-125b-5p | −0.01 | 6.17 × 10−3 | |
| Urea | hsa-mir-125b-1_hsa-miR-125b-5p 4 | hsa-miR-125b-5p | 0.20 | 5.24 × 10−3 |
| hsa-mir-125b-2_hsa-miR-125b-5p 3,4 | hsa-miR-125b-5p | 0.21 | 4.55 × 10−3 | |
| Sex | hsa-mir-7641-2_hsa-miR-7641 2 | hsa-miR-7641 | 0.72 | 4.40 × 10−3 |
| Myocardial infarction | hsa-mir-7-1_hsa-miR-7-5p 4 | hsa-miR-7-5p | −0.57 | 5.99 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalewski, D.P.; Ruszel, K.P.; Stępniewski, A.; Gałkowski, D.; Feldo, M.; Kocki, J.; Bogucka-Kocka, A. miRNA Regulatory Networks Associated with Peripheral Vascular Diseases. J. Clin. Med. 2022, 11, 3470. https://doi.org/10.3390/jcm11123470
Zalewski DP, Ruszel KP, Stępniewski A, Gałkowski D, Feldo M, Kocki J, Bogucka-Kocka A. miRNA Regulatory Networks Associated with Peripheral Vascular Diseases. Journal of Clinical Medicine. 2022; 11(12):3470. https://doi.org/10.3390/jcm11123470
Chicago/Turabian StyleZalewski, Daniel P., Karol P. Ruszel, Andrzej Stępniewski, Dariusz Gałkowski, Marcin Feldo, Janusz Kocki, and Anna Bogucka-Kocka. 2022. "miRNA Regulatory Networks Associated with Peripheral Vascular Diseases" Journal of Clinical Medicine 11, no. 12: 3470. https://doi.org/10.3390/jcm11123470