
Baroreceptors in the Aortic Arch and Their Potential Role in Aortic Dissection and Aneurysms

 , , and
, , and 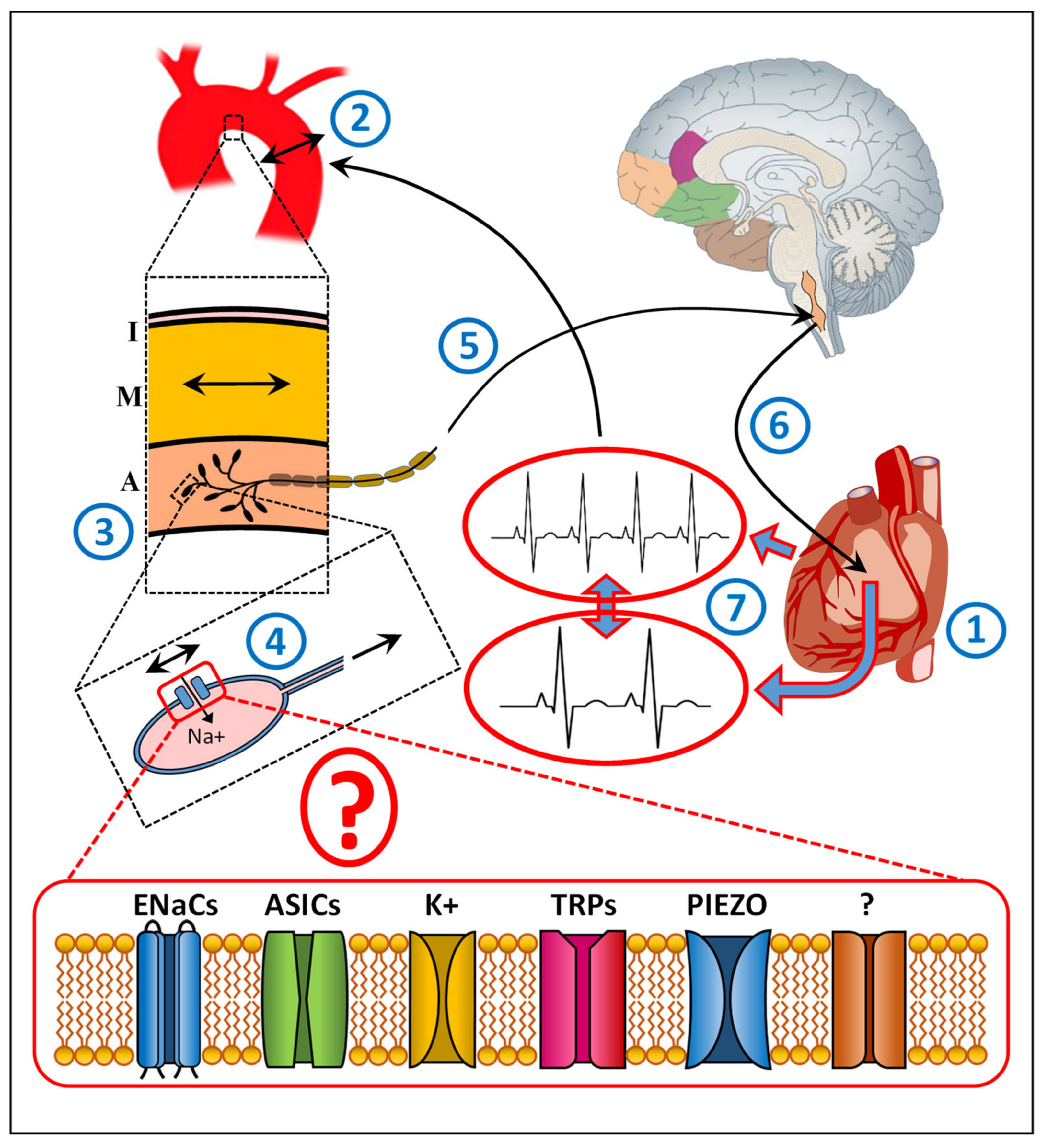{kind=link}
Abstract
:1. Introduction
2. Baroreceptor Research in Animals
3. Baroreceptor Research in Humans
4. Baroreceptors and Aortic Dissection
5. Baroreceptors and Aortic Aneurysms
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Levy, M.N. Neural control of cardiac function. Baillieres Clin. Neurol. 1997, 6, 227–244. [Google Scholar] [PubMed]
- Talman, W.T.; Kelkar, P. Neural control of the Heart Central and peripheral. Neurol. Clin. 1993, 11, 239–256. [Google Scholar] [CrossRef]
- Okada, Y.; Galbreath, M.M.; Shibata, S.; Jarvis, S.S.; VanGundy, T.B.; Meier, R.L.; Vongpatanasin, W.; Levine, B.D.; Fu, Q. Relationship between sympathetic baroreflex sensitivity and arterial stiffness in elderly men and women. Hypertension 2012, 59, 98–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Li, D.P.; Chen, S.R.; Hittelman, W.N.; Pan, H.L. Sensing of blood pressure increase by transient receptor potential vanilloid 1 receptors on baroreceptors. J. Pharmacol. Exp. Ther. 2009, 331, 851–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hainsworth, R. Cardiovascular control from cardiac and pulmonary vascular receptors. Exp. Physiol. 2014, 99, 312–319. [Google Scholar] [CrossRef]
- Lau, O.C.; Shen, B.; Wong, C.O.; Tjong, Y.W.; Lo, C.Y.; Wang, H.C.; Huang, Y.; Yung, W.H.; Chen, Y.C.; Fung, M.L.; et al. TRPC5 channels participate in pressure–sensing in aortic baroreceptors. Nat. Commun. 2016, 7, 11947. [Google Scholar] [CrossRef] [Green Version]
- Kellenberger, S.; Schild, L. Epithelial sodium channel/degenerin family of ion channels, a variety of functions for a shared structure. Physiol. Rev. 2002, 82, 735–767. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shahar, Y. Sensory functions for degenerin/epithelial sodium channels (DEG/ENaC). Adv. Genet. 2011, 76, 1–26. [Google Scholar]
- Corey, D.P.; Garcia-Anoveros, J. Mechanosensation and the DEG/ENaC ion channels. Science 1996, 273, 323–324. [Google Scholar] [CrossRef]
- Horisberger, J.D. Amiloride-sensitive Na channels. Curr. Opin. Cell. Biol. 1998, 10, 443–449. [Google Scholar] [CrossRef]
- Drummond, H.A.; Welsh, M.J.; Abboud, F.M. ENaC subunits are molecular components of the arterial baroreceptor complex. Ann. N. Y. Acad. Sci. 2001, 940, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Fronius, M.; Clauss, W.G. Mechano–sensitivity of ENaC, may the (shear) force be with you. Pflugers Arch. 2008, 455, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Hirooka, Y.; Sunagawa, K. Cardiac sympathetic afferent stimulation induces salt–sensitive sympathoexcitation through hypothalamic epithelial Na+ channel activation. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H530–H539. [Google Scholar] [CrossRef] [Green Version]
- Krueger, B.; Schlotzer-Schrehardt, U.; Haerteis, S.; Zenkel, M.; Chankiewitz, V.E.; Amann, K.U.; Kruse, F.E.; Korbmacher, C. Four subunits (alpha; beta; gamma; delta) of the epithelial sodium channel (ENaC) are expressed in the human eye in various locations. Investig. Ophthalmol. Vis. Sci. 2012, 53, 596–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loffing, J.; Schild, L. Functional domains of the epithelial sodium channel. J. Am. Soc. Nephrol. 2005, 16, 3175–3181. [Google Scholar] [CrossRef] [Green Version]
- Hanukoglu, I. ASIC and ENaC type sodium channels, conformational states and the structures of the ion selectivity filters. FEBS J. 2017, 284, 525–545. [Google Scholar] [CrossRef] [Green Version]
- Hanukoglu, I.; Hanukoglu, A. Epithelial sodium channel (ENaC) family, Phylogeny; structure–function; tissue distribution; and associated inherited diseases. Gene 2016, 579, 95–132. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Wong, C.W. Neurosensory mechanotransd through acid–sensing ion channels. J. Cell. Mol. Med. 2013, 17, 337–349. [Google Scholar] [CrossRef]
- Page, A.J.; Brierley, S.M.; Martin, C.M.; Price, M.P.; Symonds, E.; Butler, R.; Wemmie, J.A.; Blackshaw, L.A. Different contributions of ASIC channels 1a; 2; and 3 in gastrointestinal mechanosensory function. Gut 2005, 54, 1408–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielefeldt, K.; Davis, B.M. Differential effects of ASIC3 and TRPV1 deletion on gastroesophageal sensation in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G130–G138. [Google Scholar] [CrossRef] [Green Version]
- Drew, L.J.; Rohrer, D.K.; Price, M.P.; Blaver, K.E.; Cockayne, D.A.; Cesare, P.; Wood, J.N. Acid–sensing ion channels ASIC2 and ASIC3 do not contribute to mechanically activated currents in mammalian sensory neurones. J. Physiol. 2004, 556, 691–710. [Google Scholar] [CrossRef]
- Kang, S.; Jang, J.H.; Price, M.P.; Gautam, M.; Benson, C.J.; Gong, H.; Welsh, M.J.; Brennan, T.J. Simultaneous disruption of mouse ASIC1a; ASIC2 and ASIC3 genes enhances cutaneous mechanosensitivity. PLoS ONE 2012, 7, e35225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.H.; Cheng, Y.R.; Banks, R.W.; Min, M.Y.; Bewick, G.S.; Chen, C.C. Evidence for the involvement of ASIC3 in sensory mechanotransduction in proprioceptors. Nat. Commun. 2016, 7, 11460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Ma, X.; Sabharwal, R.; Snitsarev, V.; Morgan, D.; Rahmouni, K.; Drummond, H.A.; Whiteis, C.A.; Costa, V.; Price, M.; et al. The ion channel ASIC2 is required for baroreceptor and autonomic control of the circulation. Neuron 2009, 64, 885–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Naruse, K. Stretch–activated BK channel and heart function. Prog. Biophys. Mol. Biol. 2012, 110, 239–244. [Google Scholar] [CrossRef]
- Sackin, H. Stretch–activated ion channels. Kidney Int. 1995, 48, 1134–1147. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef]
- Desai, B.N.; Clapham, D.E. TRP channels and mice deficient in TRP channels. Pflugers Arch. 2005, 451, 11–18. [Google Scholar] [CrossRef]
- Liedtke, W.; Kim, C. Functionality of the TRPV subfamily of TRP ion channels, add mechano–TRP and osmo–TRP to the lexicon. Cell. Mol. Life Sci. 2005, 62, 2985–3001. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 218. [Google Scholar] [CrossRef] [Green Version]
- Glazebrook, P.A.; Schilling, W.P.; Kunze, D.L. TRPC channels as signal transducers. Pflugers Arch. 2005, 451, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranade, S.S.; Woo, S.H.; Dubin, A.E.; Moshourab, R.A.; Wetzel, C.; Petrus, M.; Mathur, J.; Bégay, V.; Coste, B.; Mainquist, J.; et al. Piezo2 is the major transducer of mech. forces for touch sensation in mice. Nature 2014, 516, 121–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonomura, K.; Woo, S.H.; Chang, R.B.; Gillich, A.; Qiu, Z.; Francisco, A.G.; Ranade, A.G.; Liberles, S.D.; Patapoutian, A. Piezo2 senses airway stretch and mediates lung inflation–induced apnoea. Nature 2017, 541, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Timmers, H.J.; Wieling, W.; Karemaker, J.M.; Lenders, J.W. Denervation of carotid baro– and chemoreceptors in humans. J. Physiol. 2003, 553 Pt 1, 3–11. [Google Scholar] [CrossRef]
- Kimani, J.K.; Mungai, J.M. Observations on the structure and innervation of the presumptive carotid sinus area in the giraffe (Giraffa camelopardalis). Acta Anat. 1983, 15, 117–133. [Google Scholar] [CrossRef] [Green Version]
- Kimani, J.K. Elastin and mechanoreceptor mechanisms with special reference to the mammalian carotid sinus. Ciba Found. Symp. 1995, 192, 215–230. [Google Scholar]
- Rees, P.M. Observations on the fine structure and distribution of presumptive baroreceptor nerves at the carotid sinus. J. Comp. Neurol. 1967, 131, 517–548. [Google Scholar] [CrossRef]
- Bagshaw, R.J.; Fischer, G.M. Morphology of the carotid sinus in the dog. J. Appl. Physiol. 1971, 31, 198–202. [Google Scholar] [CrossRef]
- Drummond, H.A.; Price, M.P.; Welsh, M.J.; Abboud, F.M. A molecular component of the arterial baroreceptor mechanotransducer. Neuron 1998, 21, 1435–1441. [Google Scholar] [CrossRef] [Green Version]
- Tu, H.; Zhang, D.; Li, Y.L. Cellular and Molecular Mechanisms Underlying Arterial Baroreceptor Remodeling in Cardiovascular Diseases and Diabetes. Neurosci. Bull. 2019, 35, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Coulson, W.F.; Weissman, N.; Carnes, W.H. Cardiovascular studies on copper–deficient swine. VII. Mechanical properties of aortic and dermal collagen. Lab. Investig. 1965, 14, 303–309. [Google Scholar]
- Li, Y.L.; Zhang, D.; Tu, H.; Muelleman, R.L. Altered ENaC is associated with aortic baroreceptor dysfunction in chronic heart failure. Am. J. Hypertens. 2016, 29, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Drummond, H.A.; Gebremedhin, D.; Harder, D.R. Degenerin/epithelial Na+ channel proteins, components of a vascular mechanosensor. Hypertension 2004, 44, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Honore, E. Sensing pressure with ion channels. Trends Neurosci. 2012, 35, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Sharif-Naeini, R.; Dedman, A.; Folgering, J.H.; Duprat, F.; Patel, A.; Nilius, B.; Honoré, E. TRP channels and mechanosensory transduction, insights into the arterial myogenic response. Pflugers Arch. 2008, 456, 529–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.Z.; Marshall, K.L.; Min, S.; Daou, I.; Chapleau, M.W.; Abboud, F.M.; Liberles, S.D.; Patapoutian, A. PIEZOs mediate neuronal sensing of blood pressure and the baroreceptor reflex. Science 2018, 362, 464–467. [Google Scholar] [CrossRef] [Green Version]
- Mancia, G.; Ferrari, A.; Gregorini, L.; Valentini, R.; Ludbrook, J.; Zanchetti, A. Circulatory reflexes from carotid and extracarotid baroreceptor areas in man. Circ. Res. 1977, 41, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, D.W.; Abboud, F.M.; Mark, A.L. Relative contribution of aortic and carotid baroreflexes to heart rate control in man during steady state and dynamic increases in arterial pressure. J. Clin. Investig. 1985, 76, 2265–2274. [Google Scholar] [CrossRef] [Green Version]
- Hutchings, C.J.; Colussi, P.; Clark, T.G. Ion channels as therapeutic antibody targets. MAbs 2019, 11, 265–296. [Google Scholar] [CrossRef]
- Koivisto, A.P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP channel drug discovery: From target validation to clinical studies. Nat. Rev. Drug Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A. Über die Struktur und die Endigungen der Aorticusfasern im Aortenbogen des Menschen mit Berücksichtigung der Cholinesterase-Aktivität der Pressorreceptoren. Z. Mikroskop. Anat. Forsch. 1956, 62, 194–228. [Google Scholar]
- Grigoreva, T.A. The Innervation of Blood Vessels; Pergamon: New York, NY, USA, 1962. [Google Scholar]
- Hilz, M.J.; Stemper, B.; Neundörfer, B. Physiology and methods for studying the baroreceptor reflex. Fortschr. Neurol. Psychiatr. 2000, 68, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M. Cardiovascular sensory receptors and their regulatory mechanisms. Ind. J. Physiol. Pharmacol. 2003, 47, 124–146. [Google Scholar]
- Laitinen, T.; Hartikainen, J.; Vanninen, E.; Niskanen, L.; Geelen, G.; Länsimies, E. Age and gender dependency of baroreflex sensitivity in healthy subjects. J. Appl. Physiol. 1998, 84, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Monahan, K.D.; Tanaka, H.; Dinenno, F.A.; Seals, D.R. Central arterial compliance is associated with age and habitual exercise–related differences in cardiovagal baroreflex sensitivity. Circulation 2001, 104, 1627–1632. [Google Scholar] [CrossRef] [Green Version]
- Lage, S.G.; Polak, J.F.; O’Leary, D.H.; Creager, M.A. Relationship of arterial compliance to baroreflex function in hypertensive patients. Am. J. Physiol. 1993, 265, H232–H237. [Google Scholar] [CrossRef]
- Conley, B.R.; Doux, J.D.; Lee, P.Y.; Bazar, K.A.; Daniel, S.M.; Yun, A.J. Integrating the theories of Darwin and Bernoulli, maladaptive baroreceptor network dysfunction may explain the pathogenesis of aortic aneurysms. Med. Hypotheses 2005, 65, 266–272. [Google Scholar] [CrossRef]
- Burrus, T.M.; Mokri, B.; Rabinstein, A.A.; Benarroch, E.E. A PRESsing dissection. Neurocrit. Care 2010, 13, 411–423. [Google Scholar] [CrossRef]
- Lénárd, Z.; Studinger, P.; Kováts, Z.; Reneman, R.; Kollai, M. Comparison of aortic arch and carotid sinus distensibility in humans--relation to baroreflex sensitivity. Auton. Neurosci. 2001, 92, 92–99. [Google Scholar] [CrossRef]
- Mancia, G.; Parati, G.; Pomidossi, G.; Casadei, R.; Di Rienzo, M.; Zanchetti, A. Arterial baroreflexes and blood pressure and heart rate variabilities in humans. Hypertension 1986, 8, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadel, P.J.; Stromstad, M.; Wray, D.W.; Smith, S.A.; Raven, P.B.; Secher, N.H. New insights into differential baroreflex control of heart rate in humans. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H735–H743. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, L.; Leone, A.; Murana, G.; Castelli, A.; Alfonsi, J.; Di Bartolomeo, R.; Pacici, D. Acute type A aortic dissection, Rationale and outcomes of extensive repair of the arch and distal aorta. Int. J. Cardiol. 2018, 267, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Gawinecka, J.; Schönrath, F.; von Eckardstein, A. Acute aortic dissection, pathogenesis; risk factors and diagnosis. Swiss. Med. Wkly. 2017, 147, w14489. [Google Scholar] [CrossRef] [PubMed]
- Hagan, P.G.; Nienaber, C.A.; Isselbacher, E.M.; Bruckman, D.; Karavite, D.J.; Russman, P.L.; Evangelista, A.; Fattori, R.; Suzuki, T.; Oh, J.K.; et al. The International Registry of Acute Aortic Dissection (IRAD), new insights into an old disease. JAMA 2000, 283, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Vilacosta, I.; Aragoncillo, P.; Cañadas, V.; San Román, J.A.; Ferreirós, J.; Rodríguez, E. Acute aortic syndrome, a new look at an old conundrum. Postgrad. Med. J. 2010, 86, 52–61. [Google Scholar] [CrossRef]
- White, A.; Broder, J.; Mando-Vandrick, J.; Wendell, J.; Crowe, J. Acute aortic emergencies—Part 2, aortic dissections. Adv. Emerg. Nurs. J. 2013, 35, 28–52. [Google Scholar] [CrossRef]
- Reutersberg, B.; Salvermoser, M.; Trenner, M.; Geisbüsch, S.; Zimmermann, A.; Eckstein, H.H.; Kuehnl, A. Hospital Incidence and In-Hospital Mortality of Surgically and Interventionally Treated Aortic Dissections: Secondary Data Analysis of the Nationwide German Diagnosis-Related Group Statistics From 2006 to 2014. J. Am. Heart Assoc. 2019, 8, e011402. [Google Scholar] [CrossRef]
- Ishii, T.; Asuwa, N. Collagen and elastin degradation by matrix metalloproteinases and tissue inhibitors of matrix metalloproteinase in aortic dissection. Hum. Pathol. 2000, 31, 640–646. [Google Scholar] [CrossRef]
- Januzzi, J.L.; Isselbacher, E.M.; Fattori, R.; Cooper, J.V.; Smith, D.E.; Fang, J.; Eagle, K.A.; Mehta, R.H.; Nienaber, C.A.; Pape, L.A.; et al. Characterizing the young patient with aortic dissection, results from the International Registry of Aortic Dissection (IRAD). J. Am. Coll. Cardiol. 2004, 43, 665–669. [Google Scholar] [CrossRef] [Green Version]
- Chae, C.U.; Lee, R.T.; Rifai, N.; Ridker, P.M. Blood pressure and inflammation in apparently healthy men. Hypertension 2001, 38, 399–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elefteriades, J.A. Natural history of thoracic aortic aneurysms, indications for surgery; and surgical versus nonsurgical risks. Ann. Thorac. Surg. 2002, 74, S1877–S1898. [Google Scholar] [CrossRef]
- Tsai, T.T.; Trimarchi, S.; Nienaber, C.A. Acute aortic dissection, perspectives from the International Registry of Acute Aortic Dissection (IRAD). Eur. J. Vasc. Endovasc. Surg. 2009, 37, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggebrecht, H.; Schmermund, A.; von Birgelen, C.; Naber, C.K.; Bartel, T.; Wenzel, R.R.; Erbel, R. Resistant hypertension in patients with chronic aortic dissection. J. Hum. Hypertens. 2005, 19, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Januzzi, J.L.; Sabatine, M.S.; Choi, J.C.; Abernethy, W.B.; Isselbacher, E.M. Refractory systemic hypertension following type B aortic dissection. Am. J. Cardiol. 2001, 88, 686–688. [Google Scholar] [CrossRef]
- MacGillivray, T.E.; Gleason, T.G.; Patel, H.J.; Aldea, G.S.; Bavaria, J.E.; Beaver, T.M.; Chen, E.P.; Czerny, M.; Estrera, A.L.; Firestone, S.; et al. The Society of Thoracic Surgeons/American Association for Thoracic Surgery Clinical Practice Guidelines on the Management of Type B Aortic Dissection. Ann. Thorac. Surg. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Lewis, A.H.; Grandl, J. Mechanical sensitivity of Piezo1 ion channels can be tuned by cellular membrane tension. Elife 2015, 4, e12088. [Google Scholar] [CrossRef] [Green Version]
- Sherifova, S.; Holzapfel, G.A. Biomechanics of aortic wall failure with a focus on dissection and aneurysm: A review. Acta Biomater. 2019, 99, 1–17. [Google Scholar] [CrossRef]
- Reeps, C.; Pelisek, J.; Seidl, S.; Schuster, T.; Zimmermann, A.; Kuehnl, A.; Eckstein, H.H. Inflammatory infiltrates and neovessels are relevant sources of MMPs in abdominal aortic aneurysm wall. Pathobiology 2009, 76, 243–252. [Google Scholar] [CrossRef]
- Reeps, C.; Kehl, S.; Tanios, F.; Biehler, J.; Pelisek, J.; Wall, W.A.; Eckstein, H.H.; Gee, M.W. Biomechanics and gene expression in abdominal aortic aneurysm. J. Vasc. Surg. 2014, 60, 1640–1647.e2. [Google Scholar] [CrossRef] [Green Version]
- Tanios, F.; Gee, M.W.; Pelisek, J.; Kehl, S.; Biehler, J.; Grabher-Meier, V.; Wall, W.A.; Eckstein, H.H.; Reeps, C. Interaction of biomechanics with extracellular matrix components in abdominal aortic aneurysm wall. Eur. J. Vasc. Endovasc. Surg. 2015, 50, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keisler, B.; Carter, C. Abdominal aortic aneurysm. Am. Fam. Physician 2015, 91, 538–543. [Google Scholar] [PubMed]
- Klassen, S.A.; Chirico, D.; Dempster, K.S.; Shoemaker, J.K.; O’Leary, D.D. Role of aortic arch vascular mechanics in cardiovagal baroreflex sensitivity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R24–R32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lord, R.N.; Wakeham, D.J.; Pugh, C.J.A.; Simpson, L.L.; Talbot, J.S.; Lodge, F.M.; Curry, B.A.; Dawkins, T.G.; Shave, R.E.; Moore, J.P. The influence of barosensory vessel mechanics on the vascular sympathetic baroreflex: Insights into aging and blood pressure homeostasis. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H370–H376. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reutersberg, B.; Pelisek, J.; Ouda, A.; de Rougemont, O.; Rössler, F.; Zimmermann, A. Baroreceptors in the Aortic Arch and Their Potential Role in Aortic Dissection and Aneurysms. J. Clin. Med. 2022, 11, 1161. https://doi.org/10.3390/jcm11051161
Reutersberg B, Pelisek J, Ouda A, de Rougemont O, Rössler F, Zimmermann A. Baroreceptors in the Aortic Arch and Their Potential Role in Aortic Dissection and Aneurysms. Journal of Clinical Medicine. 2022; 11(5):1161. https://doi.org/10.3390/jcm11051161
Chicago/Turabian StyleReutersberg, Benedikt, Jaroslav Pelisek, Ahmed Ouda, Olivier de Rougemont, Fabian Rössler, and Alexander Zimmermann. 2022. "Baroreceptors in the Aortic Arch and Their Potential Role in Aortic Dissection and Aneurysms" Journal of Clinical Medicine 11, no. 5: 1161. https://doi.org/10.3390/jcm11051161