
Exploration of the Supraspinal Hypotheses about Spinal Cord Stimulation and Dorsal Root Ganglion Stimulation: A Systematic Review

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protocol and Registration
2.2. Search Strategy
2.3. Eligibility Criteria
2.4. Study Selection
2.5. Data Extraction
2.6. Risk of Bias Assessment and Confidence in the Body of Evidence
3. Results
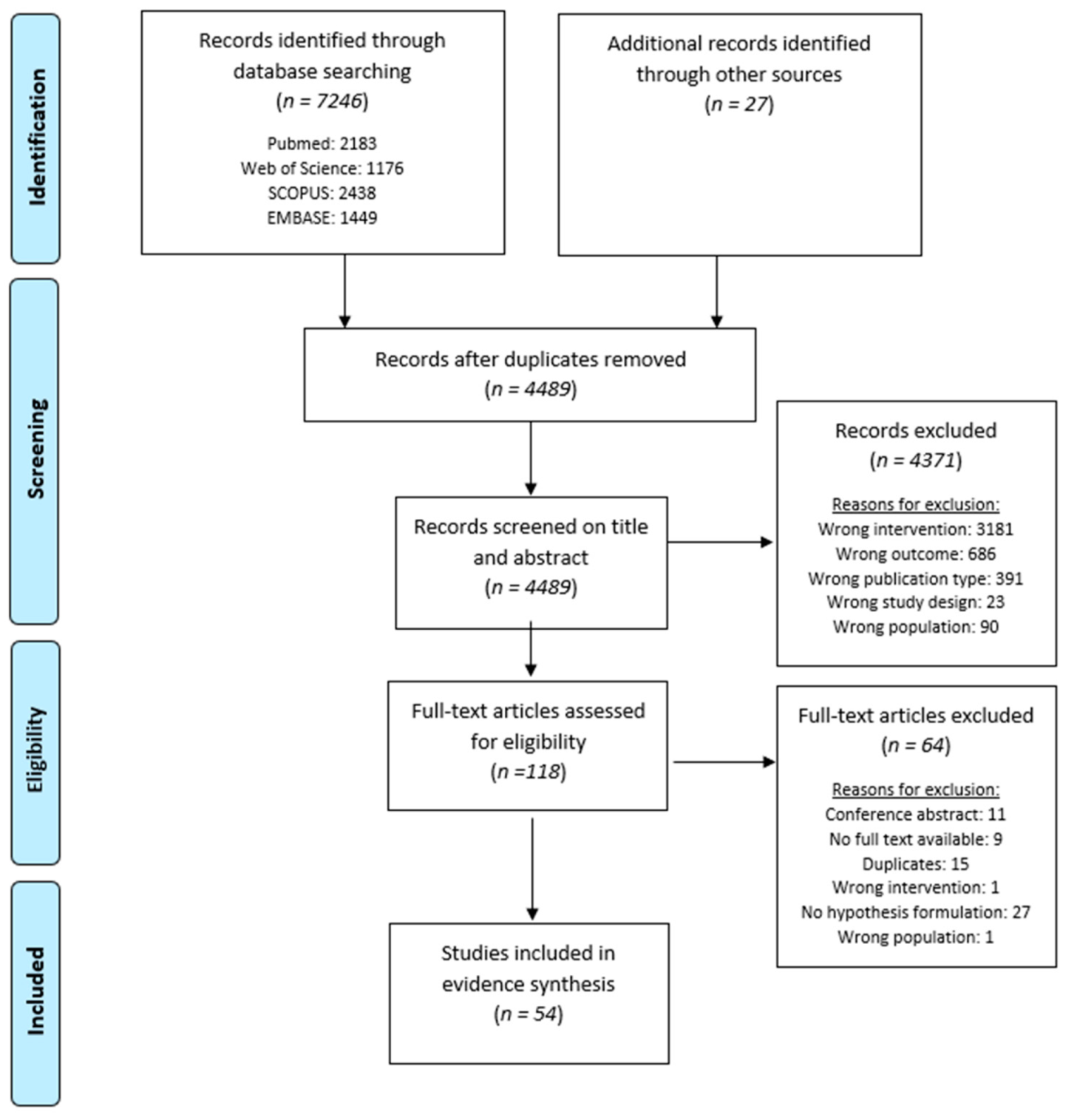
3.1. Study Selection
3.2. Study Characteristics
3.3. Risk of Bias Assessment
3.4. Results of Individual Studies
3.4.1. Ascending Pathways
3.4.2. Ascending Medial Pathway
Scientific Support for This Hypothesis
Confidence in the Body of Evidence for This Hypothesis
3.4.3. Ascending Lateral Pathway
Scientific Support for This Hypothesis
Confidence in the Body of Evidence for This Hypothesis
3.4.4. Descending Pathways
Scientific Support for This Hypothesis
Confidence in the Body of Evidence for This Hypothesis
3.4.5. Spinal–Cerebral (Thalamic)-Loop
Confidence in the Body of Evidence for This Hypothesis
3.4.6. Affective/Motivational Influence
Confidence in the Body of Evidence for This Hypothesis
3.4.7. Miscellaneous
Confidence in the Body of Evidence for This Hypothesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grider, J.S.; Manchikanti, L.; Carayannopoulos, A.; Sharma, M.L.; Balog, C.C.; E Harned, M.; Grami, V.; Justiz, R.; Nouri, K.H.; Hayek, S.M.; et al. Effectiveness of spinal cord stimulation in chronic spinal pain: A systematic review. Pain Physician 2016, 19, 33–54. [Google Scholar] [CrossRef]
- Shealy, C.N.; Mortimer, J.T.; Reswick, J.B. Electrical inhibition of pain by stimulation of the dorsal columns: Preliminary clinical report. Anesth. Analg. 1967, 46, 489–491. [Google Scholar] [CrossRef]
- Meyerson, B.; Herregodts, P.; Linderoth, B.; Ren, B. An experimental animal model of spinal cord stimulation for pain. Ster. Funct. Neurosurg. 1994, 62, 256–262. [Google Scholar] [CrossRef]
- Melzack, R.; Wall, P.D. Pain mechanisms: A new theory. Science 1965, 150, 971–978. [Google Scholar] [CrossRef]
- Barchini, J.; Tchachaghian, S.; Shamaa, F.; Jabbur, S.; Meyerson, B.; Song, Z.; Linderoth, B.; Saadé, N. Spinal segmental and supraspinal mechanisms underlying the pain-relieving effects of spinal cord stimulation: An experimental study in a rat model of neuropathy. Neuroscience 2012, 215, 196–208. [Google Scholar] [CrossRef]
- Bentley, L.D.; Duarte, R.; Furlong, P.L.; Ashford, R.L.; Raphael, J.H. Brain activity modifications following spinal cord stimulation for chronic neuropathic pain: A systematic review. Eur. J. Pain 2015, 20, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarthy, K.; A Fishman, M.; Zuidema, X.; Hunter, C.W.; Levy, R. Mechanism of action in burst spinal cord stimulation: Review and recent advances. Pain Med. 2019, 20, S13–S22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linderoth, B.; Foreman, R.D. Conventional and novel spinal stimulation algorithms: Hypothetical mechanisms of action and comments on outcomes. Neuromodulation Technol. Neural Interface 2017, 20, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Caylor, J.; Reddy, R.; Yin, S.; Cui, C.; Huang, M.; Huang, C.; Rao, R.; Baker, D.G.; Simmons, A.; Souza, D.; et al. Spinal cord stimulation in chronic pain: Evidence and theory for mechanisms of action. Bioelectron. Med. 2019, 5. [Google Scholar] [CrossRef]
- Ahmed, S.; Yearwood, T.; De Ridder, D.; Vanneste, S. Burst and high frequency stimulation: Underlying mechanism of action. Expert Rev. Med. Devices 2017, 15, 61–70. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 62, e1–e34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Lin, J.; Demner-Fushman, D. Evaluation of PICO as a knowledge representation for clinical questions. AMIA Annu. Symp. Proc. 2006, 2006, 359–363. [Google Scholar]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan—A web and mobile app for systematic reviews. Syst. Rev. 2016, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Driessen, S.; Bodewein, L.; Dechent, D.; Graefrath, D.; Schmiedchen, K.; Stunder, D.; Kraus, T.; Petri, A.-K. Biological and health-related effects of weak static magnetic fields (≤1 mT) in humans and vertebrates: A systematic review. PLoS ONE 2020, 15, e0230038. [Google Scholar] [CrossRef]
- National Toxicology Program (NTP). Handbook for Conducting a Literature-Based Health Assessment Using OHAT Approach for Systematic Review and Evidence Integration; U.S. Dept. of Health and Human Services, National Toxicology Program: Research Triangle, NC, USA, 2015. [Google Scholar]
- Rooney, A.A.; Boyles, A.; Wolfe, M.S.; Bucher, J.R.; Thayer, K.A. Systematic review and evidence integration for literature-based environmental health science assessments. Environ. Health Perspect. 2014, 122, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Morgan, R.L.; Thayer, K.A.; Bero, L.; Bruce, N.; Falck-Ytter, Y.; Ghersi, D.; Guyatt, G.; Hooijmans, C.; Langendam, M.; Mandrioli, D.; et al. GRADE: Assessing the quality of evidence in environmental and occupational health. Environ. Int. 2016, 92–93, 611–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, J.; Pulecchi, F.; Dilena, R.; Oliviero, A.; Priori, A.; Foffani, G. Spinal direct current stimulation modulates the activity of gracile nucleus and primary somatosensory cortex in anaesthetized rats. J. Physiol. 2011, 589, 4981–4996. [Google Scholar] [CrossRef]
- Atweh, S.F.; Dajani, B.M.; Saade, N.E.; Jabbur, S.J. Supraspinal inhibition of trigeminal input into subnucleus caudalis by dorsal column stimulation. Brain Res. 1985, 348, 401–404. [Google Scholar] [CrossRef]
- Bantli, H.; Bloedel, J.R.; Thienprasit, P. Supraspinal interactions resulting from experimental dorsal column stimulation. J. Neurosurg. 1975, 42, 296–300. [Google Scholar] [CrossRef]
- Dejongste, M.J.; Hautvast, R.W.; Ruiters, M.H.; Ter Horst, G.J. Spinal cord stimulation and the induction of c-fos and heat shock protein 72 in the central nervous system of rats. Neuromodulation Technol. Neural Interface 1998, 1, 73–84. [Google Scholar] [CrossRef]
- Dembowsky, K.; Czachurski, J.; Seller, H. An intracellular study of the synaptic input to sympathetic preganglionic neurones of the third thoracic segment of the cat. J. Auton. Nerv. Syst. 1985, 13, 201–244. [Google Scholar] [CrossRef]
- El-Khoury, C.; Hawwa, N.; Baliki, M.; Atweh, S.; Jabbur, S.; Saadé, N. Attenuation of neuropathic pain by segmental and supraspinal activation of the dorsal column system in awake rats. Neuroscience 2002, 112, 541–553. [Google Scholar] [CrossRef]
- Linderoth, B.; Stiller, C.-O.; Gunasekera, L.; O’Connor, W.; Franck, J.; Gazelius, B.; Brodin, E. Release of neurotransmitters in the CNS by spinal cord stimulation: Survey of present state of knowledge and recent experimental studies. Ster. Funct. Neurosurg. 1993, 61, 157–170. [Google Scholar] [CrossRef]
- Maeda, Y.; Ikeuchi, M.; Wacnik, P.; Sluka, K.A. Increased c-fos immunoreactivity in the spinal cord and brain following spinal cord stimulation is frequency-dependent. Brain Res. 2009, 1259, 40–50. [Google Scholar] [CrossRef]
- Meuwissen, K.P.; Van Der Toorn, A.; Gu, J.W.; Zhang, T.C.; Dijkhuizen, R.M.; Joosten, E.A. Active recharge burst and tonic spinal cord stimulation engage different supraspinal mechanisms: A functional magnetic resonance imaging study in peripherally injured chronic neuropathic rats. Pain Pr. 2020, 20, 510–521. [Google Scholar] [CrossRef]
- Pawela, C.P.; Kramer, J.M.; Hogan, Q.H. Dorsal root ganglion stimulation attenuates the BOLD signal response to noxious sensory input in specific brain regions: Insights into a possible mechanism for analgesia. NeuroImage 2017, 147, 10–18. [Google Scholar] [CrossRef]
- Quindlen-Hotek, J.C.; Kent, A.R.; De Anda, P.; Kartha, S.; Benison, A.M.; Winkelstein, B.A. Changes in neuronal activity in the anterior cingulate cortex and primary somatosensory cortex with nonlinear burst and tonic spinal cord stimulation. Neuromodulation Technol. Neural Interface 2020, 23, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Saadé, N.E.; Barchini, J.; Tchachaghian, S.; Chamaa, F.; Jabbur, S.J.; Song, Z.; Meyerson, B.A.; Linderoth, B. The role of the dorsolateral funiculi in the pain relieving effect of spinal cord stimulation: A study in a rat model of neuropathic pain. Exp. Brain Res. 2014, 233, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Saadé, N.E.; Atweh, S.F.; Tabet, M.S.; Jabbur, S.J. Inhibition of nociceptive withdrawal flexion reflexes through a dorsal column-brainstem-spinal loop. Brain Res. 1985, 335, 306–308. [Google Scholar] [CrossRef]
- Saade, N.E.; Tabet, M.S.; Banna, N.R.; Atweh, S.F.; Jabbur, S.J. Inhibition of nociceptive evoked activity in spinal neurons through a dorsal column-brainstem-spinal loop. Brain Res. 1985, 339, 115–118. [Google Scholar] [CrossRef]
- Song, Z.; Ansah, O.; Meyerson, B.; Pertovaara, A.; Linderoth, B. The rostroventromedial medulla is engaged in the effects of spinal cord stimulation in a rodent model of neuropathic pain. Neuroscience 2013, 247, 134–144. [Google Scholar] [CrossRef]
- Song, Z.; Ansah, O.; Meyerson, B.; Pertovaara, A.; Linderoth, B. Exploration of supraspinal mechanisms in effects of spinal cord stimulation: Role of the locus coeruleus. Neuroscience 2013, 253, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Ultenius, C.; Meyerson, B.A.; Linderoth, B. Pain relief by spinal cord stimulation involves serotonergic mechanisms: An experimental study in a rat model of mononeuropathy. Pain 2009, 147, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Stiller, C.-O.; Linderoth, B.; O’Connor, W.T.; Franck, J.; Falkenberg, T.; Ungerstedt, U.; Brodin, E. Repeated spinal cord stimulation decreases the extracellular level of γ-aminobutyric acid in the periaqueductal gray matter of freely moving rats. Brain Res. 1995, 699, 231–241. [Google Scholar] [CrossRef]
- Tazawa, T.; Kamiya, Y.; Kobayashi, A.; Saeki, K.; Takiguchi, M.; Nakahashi, Y.; Shinbori, H.; Funakoshi, K.; Goto, T. Spinal cord stimulation modulates supraspinal centers of the descending antinociceptive system in rats with unilateral spinal nerve injury. Mol. Pain 2015, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, R.; Gupta, A.; Kelley, C.A.; Vallejo, A.; Rink, J.; Williams, J.M.; Cass, C.L.; Smith, W.J.; Benyamin, R.; Cedeño, D.L. Effects of phase polarity and charge balance spinal cord stimulation on behavior and gene expression in a rat model of neuropathic pain. Neuromodulation Technol. Neural Interface 2019, 23, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.U.; Zhang, Y.; Chen, L.; Hillary, K.S.; Cohen, A.; Vo, T.; Houghton, M.; Mao, J. Effects of spinal cord stimulation on pain thresholds and sensory perceptions in chronic pain patients. Neuromodulation Technol. Neural Interface 2015, 18, 355–360. [Google Scholar] [CrossRef]
- Blair, R.G.; Lee, R.G.; Vanderlinden, G. Dorsal column stimulation. Its effect on the somatosensory evoked response. Arch. Neurol. 1975, 32, 826–829. [Google Scholar] [CrossRef]
- Bocci, T.; De Carolis, G.; Paroli, M.; Barloscio, D.; Parenti, L.; Tollapi, L.; Valeriani, M.; Sartucci, F. Neurophysiological comparison among tonic, high frequency, and burst spinal cord stimulation: Novel insights into spinal and brain mechanisms of action. Neuromodulation Technol. Neural Interface 2018, 21, 480–488. [Google Scholar] [CrossRef]
- Buentjen, L.; Vicheva, P.; Chander, B.; Beccard, S.; ScD, C.C.; Azañón, E.; Stenner, M.; Deliano, M. spatial filtering of electroencephalography reduces artifacts and enhances signals related to spinal cord stimulation (SCS). Neuromodulation Technol. Neural Interface 2020. [Google Scholar] [CrossRef]
- De Andrade, D.C.; Bendib, B.; Hattou, M.; Keravel, Y.; Nguyen, J.-P.; Lefaucheur, J.-P. Neurophysiological assessment of spinal cord stimulation in failed back surgery syndrome. Pain 2010, 150, 485–491. [Google Scholar] [CrossRef]
- De Groote, S.; Goudman, L.; Linderoth, B.; Buyck, F.; Rigoard, P.; De Jaeger, M.; Van Schuerbeek, P.; Peeters, R.; Sunaert, S.; Moens, M. A regions of interest voxel-based morphometry study of the human brain during high-frequency spinal cord stimulation in patients with failed back surgery syndrome. Pain Pr. 2020, 20, 878–888. [Google Scholar] [CrossRef] [PubMed]
- De Groote, S.; Goudman, L.; Van Schuerbeek, P.; Peeters, R.; Sunaert, S.; Linderoth, B.; De Andrés, J.; Rigoard, P.; De Jaeger, M.; Moens, M. Effects of spinal cord stimulation on voxel-based brain morphometry in patients with failed back surgery syndrome. Clin. Neurophysiol. 2020, 131, 2578–2587. [Google Scholar] [CrossRef] [PubMed]
- De Groote, S.; Goudman, L.; Peeters, R.; Linderoth, B.; Vanschuerbeek, P.; Sunaert, S.; Jaeger, M.D.; De Smedt, A.; Moens, M. Magnetic resonance imaging exploration of the human brain during 10 kHz spinal cord stimulation for failed back surgery syndrome: A resting state functional magnetic resonance imaging study. Neuromodul. Technol. Neural Interface 2019, 23, 46–55. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Vanneste, S. Burst and tonic spinal cord stimulation: Different and common brain mechanisms. Neuromodulation Technol. Neural Interface 2015, 19, 47–59. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Plazier, M.; Kamerling, N.; Menovsky, T.; Vanneste, S. Burst spinal cord stimulation for limb and back pain. World Neurosurg. 2013, 80, 642–649.e1. [Google Scholar] [CrossRef]
- Deogaonkar, M.; Sharma, M.; Oluigbo, C.; Nielson, D.M.; Yang, X.; Vera-Portocarrero, L.; Molnar, G.F.; Abduljalil, A.; Sederberg, P.B.; Knopp, M.; et al. Spinal cord stimulation (SCS) and Functional magnetic resonance imaging (fMRI): Modulation of cortical connectivity with therapeutic SCS. Neuromodulation Technol. Neural Interface 2015, 19, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Gildenberg, P.; Murthy, K. Influence of dorsal column stimulation upon human thalamic somatosensory-evoked potentials. Ster. Funct. Neurosurg. 1980, 43, 8–17. [Google Scholar] [CrossRef]
- Goudman, L.; Brouns, R.; De Groote, S.; De Jaeger, M.; Huysmans, E.; Forget, P.; Moens, M. Association between spinal cord stimulation and top-down nociceptive inhibition in people with failed back surgery syndrome: A cohort study. Phys. Ther. 2019, 99, 915–923. [Google Scholar] [CrossRef]
- Goudman, L.; Linderoth, B.; Nagels, G.; Huysmans, E.; Moens, M. Cortical mapping in conventional and high dose spinal cord stimulation: An exploratory power spectrum and functional connectivity analysis with electroencephalography. Neuromodulation Technol. Neural Interface 2019, 23, 74–81. [Google Scholar] [CrossRef]
- Kishima, H.; Saitoh, Y.; Oshino, S.; Hosomi, K.; Ali, M.; Maruo, T.; Hirata, M.; Goto, T.; Yanagisawa, T.; Sumitani, M.; et al. Modulation of neuronal activity after spinal cord stimulation for neuropathic pain; H215O PET study. NeuroImage 2010, 49, 2564–2569. [Google Scholar] [CrossRef]
- Kunitake, A.; Iwasaki, T.; Hidaka, N.; Nagamachi, S.; Katsuki, H.; Uno, T.; Takasaki, M. The effects of spinal cord stimulation on the neuronal activity of the brain in patients with chronic neuropathic pain. Pain Res. 2005, 20, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Larson, S.J.; Sances, A.; Riegel, D.H.; Meyer, G.A.; Dallmann, D.E.; Swiontek, T. Neurophysiological effects of dorsal column stimulation in man and monkey. J. Neurosurg. 1974, 41, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Lind, A.-L.; Khoonsari, P.E.; Sjödin, M.; Katila, L.; Wetterhall, M.; Gordh, T.; Kultima, K. spinal cord stimulation alters protein levels in the cerebrospinal fluid of neuropathic pain patients: A proteomic mass spectrometric analysis. Neuromodulation Technol. Neural Interface 2016, 19, 549–562. [Google Scholar] [CrossRef]
- Mehta, V.; Bouchareb, Y.; Ramaswamy, S.; Ahmad, A.; Wodehouse, T.; Haroon, A. Metabolic imaging of pain matrix using 18 F Fluoro-deoxyglucose positron emission tomography/computed tomography for patients undergoing L2 dorsal root ganglion stimulation for low back pain. Neuromodulation Technol. Neural Interface 2019, 23, 222–233. [Google Scholar] [CrossRef]
- Modesti, L.M.; Waszak, M. Firing pattern of cells in human thalamus during dorsal column stimulation. Ster. Funct. Neurosurg. 1975, 38, 251–258. [Google Scholar] [CrossRef]
- Moens, M.; Mariën, P.; Brouns, R.; Poelaert, J.; De Smedt, A.; Buyl, R.; Droogmans, S.; Van Schuerbeek, P.; Sunaert, S.; Nuttin, B. Spinal cord stimulation modulates cerebral neurobiology: A proton magnetic resonance spectroscopy study. Neuroradiology 2013, 55, 1039–1047. [Google Scholar] [CrossRef]
- Moens, M.; Sunaert, S.; Mariën, P.; Brouns, R.; De Smedt, A.; Droogmans, S.; Van Schuerbeek, P.; Peeters, R.; Poelaert, J.; Nuttin, B. Spinal cord stimulation modulates cerebral function: An fMRI study. Neuroradiology 2012, 54, 1399–1407. [Google Scholar] [CrossRef]
- Morgalla, M.H.; Filho, M.F.D.B.; Chander, B.S.; Soekadar, S.R.; Tatagiba, M.; Lepski, G. Neurophysiological effects of dorsal root ganglion stimulation (DRGS) in pain processing at the cortical level. Neuromodulation Technol. Neural Interface 2018, 22, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Pahapill, P.A.; Zhang, W. Restoration of altered somatosensory cortical representation with spinal cord stimulation therapy in a patient with complex regional pain syndrome: A magnetoencephalography case study. Neuromodulation Technol. Neural Interface 2014, 17, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Poláček, H.; Kozák, J.; Vrba, I.; Vrána, J.; Stančák, A. Effects of spinal cord stimulation on the cortical somatosensory evoked potentials in failed back surgery syndrome patients. Clin. Neurophysiol. 2007, 118, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Royds, J.; Conroy, M.J.; Dunne, M.R.; Cassidy, H.; Matallanas, D.; Lysaght, J.; McCrory, C. Examination and characterisation of burst spinal cord stimulation on cerebrospinal fluid cellular and protein constituents in patient responders with chronic neuropathic pain—A Pilot Study. J. Neuroimmunol. 2020, 344, 577249. [Google Scholar] [CrossRef] [PubMed]
- Schlaier, J.R.; Eichhammer, P.; Langguth, B.; Doenitz, C.; Binder, H.; Hajak, G.; Brawanski, A. Effects of spinal cord stimulation on cortical excitability in patients with chronic neuropathic pain: A pilot study. Eur. J. Pain 2007, 11, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Shimoji, K.; Shimizu, H.; Maruyama, Y.; Matsuki, M.; Kuribayashi, H.; Fujioka, H. Dorsal column stimulation in man: Facilitation of primary afferent depolarization. Anesth. Analg. 1982, 61, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Schuh-Hofer, S.; Fischer, J.; Unterberg, A.; Treede, R.-D.; Ahmadi, R. Spinal cord stimulation modulates descending pain inhibition and temporal summation of pricking pain in patients with neuropathic pain. Acta Neurochir. 2018, 160, 2509–2519. [Google Scholar] [CrossRef]
- Stančák, A.; Kozák, J.; Vrba, I.; Tintěra, J.; Vrána, J.; Poláček, H.; Stančák, M. Functional magnetic resonance imaging of cerebral activation during spinal cord stimulation in failed back surgery syndrome patients. Eur. J. Pain 2008, 12, 137–148. [Google Scholar] [CrossRef]
- Sufianov, A.A.; Shapkin, A.G.; Sufianova, G.Z.; Elishev, V.G.; Barashin, D.A.; Berdichevskii, V.B.; Churkin, S.V. Functional and metabolic changes in the brain in neuropathic pain syndrome against the background of chronic epidural electrostimulation of the spinal cord. Bull. Exp. Biol. Med. 2014, 157, 462–465. [Google Scholar] [CrossRef]
- Weigel, R.; Capelle, H.H.; Flor, H.; Krauss, J.K. Event-related cortical processing in neuropathic pain under long-term spinal cord stimulation. Pain Physician 2015, 18, 185–194. [Google Scholar]
- Yearwood, T.; De Ridder, D.; Bin Yoo, H.; Falowski, S.; Venkatesan, L.; To, W.T.; Vanneste, S. Comparison of neural activity in chronic pain patients during tonic and burst spinal cord stimulation using fluorodeoxyglucose positron emission tomography. Neuromodulation Technol. Neural Interface 2019, 23, 56–63. [Google Scholar] [CrossRef]
- Yen, C.-T.; Lu, P.-L. Thalamus and pain. Acta Anaesthesiol. Taiwanica 2013, 51, 73–80. [Google Scholar] [CrossRef]
- Ray, J.P.; Price, J.L. The organization of projections from the mediodorsal nucleus of the thalamus to orbital and medial prefrontal cortex in macaque monkeys. J. Comp. Neurol. 1993, 337, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-C.; Shyu, B.-C. Differential projections from the mediodorsal and centrolateral thalamic nuclei to the frontal cortex in rats. Brain Res. 2004, 995, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Valeriani, M.; Rambaud, L.; Mauguière, F. Scalp topography and dipolar source modelling of potentials evoked by CO2 laser stimulation of the hand. Electroencephalogr. Clin. Neurophysiol. Potentials Sect. 1996, 100, 343–353. [Google Scholar] [CrossRef]
- Gauriau, C.; Bernard, J.-F. Posterior triangular thalamic neurons convey nociceptive messages to the secondary somatosensory and insular cortices in the rat. J. Neurosci. 2004, 24, 752–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Larrea, L.; Frot, M.; Valeriani, M. Brain generators of laser-evoked potentials: From dipoles to functional significance. Neurophysiol. Clin. Neurophysiol. 2003, 33, 279–292. [Google Scholar] [CrossRef]
- Valentini, E.; Hu, L.; Chakrabarti, B.; Hu, Y.; Aglioti, S.; Iannetti, G. The primary somatosensory cortex largely contributes to the early part of the cortical response elicited by nociceptive stimuli. NeuroImage 2012, 59, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Basbaum, A.I.; Clanton, C.H.; Fields, H.L. Opiate and stimulus-produced analgesia: Functional anatomy of a medullospinal pathway. Proc. Natl. Acad. Sci. USA 1976, 73, 4685–4688. [Google Scholar] [CrossRef] [Green Version]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised international association for the study of pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef]
- Johnson, M. Transcutaneous electrical nerve stimulation: Mechanisms, clinical application and evidence. Rev. Pain 2007, 1, 7–11. [Google Scholar] [CrossRef]
- Lv, Q.; Wu, F.; Gan, X.; Yang, X.; Zhou, L.; Chen, J.; He, Y.; Zhang, R.; Zhu, B.; Liu, L. The involvement of descending pain inhibitory system in electroacupuncture-induced analgesia. Front. Integr. Neurosci. 2019, 13, 38. [Google Scholar] [CrossRef]
- Van Oosterwijck, J.; Marušič, U.; De Wandele, I.; Paul, L.; Meeus, M.; Moorkens, G.; Lambrecht, L.; Danneels, L.; Nijs, J. The role of autonomic function in exercise-induced endogenous analgesia: A case-control study in myalgic encephalomyelitis/chronic fatigue syndrome and healthy people. Pain Physician 2017, 20, E389–E399. [Google Scholar]
- Gomolka, S.; Vaegter, H.B.; Nijs, J.; Meeus, M.; Gajsar, H.; Hasenbring, M.I.; Titze, C. Assessing endogenous pain inhibition: Test–retest reliability of exercise-induced hypoalgesia in local and remote body parts after aerobic cycling. Pain Med. 2019, 20, 2272–2282. [Google Scholar] [CrossRef]
- Knotkova, H.; Hamani, C.; Sivanesan, E.; Le Beuffe, M.F.E.; Moon, J.Y.; Cohen, S.P.; A Huntoon, M. Neuromodulation for chronic pain. Lancet 2021, 397, 2111–2124. [Google Scholar] [CrossRef]
- Meuwissen, K.P.; De Vries, L.E.; Gu, J.W.; Zhang, T.C.; Joosten, E.A.J. Burst and tonic spinal cord stimulation both activate spinal GABAergic mechanisms to attenuate pain in a rat model of chronic neuropathic pain. Pain Pr. 2019, 20, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Y. Spinal cord stimulation: Neurophysiological and neurochemical mechanisms of action. Curr. Pain Headache Rep. 2012, 16, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Ultenius, C.; Song, Z.; Lin, P.; Meyerson, B.A.; Linderoth, B. Spinal GABAergic mechanisms in the effects of spinal cord stimulation in a rodent model of neuropathic pain: Is GABA synthesis involved? Neuromodulation Technol. Neural Interface 2012, 16, 114–120. [Google Scholar] [CrossRef]
- Linderoth, B.; Herregodts, P.; Meyerson, B.A. Sympathetic mediation of peripheral vasodilation induced by spinal cord stimulation: Animal studies of the role of cholinergic and adrenergic receptor subtypes. Neurosurgery 1994, 35, 711–719. [Google Scholar] [CrossRef]
- Stiller, C.-O.; Cui, J.-G.; O’Connor, W.; Brodin, E.; Meyerson, B.A.; Linderoth, B. Release of γ-aminobutyric acid in the dorsal horn and suppression of tactile allodynia by spinal cord stimulation in mononeuropathic rats. Neurosurgery 1996, 39, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.K.; Vaughan, C.W. Descending modulation of pain: The GABA disinhibition hypothesis of analgesia. Curr. Opin. Neurobiol. 2014, 29, 159–164. [Google Scholar] [CrossRef]
- Zander, H.J.; Graham, R.D.; Anaya, C.J.; Lempka, S.F. Anatomical and technical factors affecting the neural response to epidural spinal cord stimulation. J. Neural Eng. 2020, 17, 036019. [Google Scholar] [CrossRef] [PubMed]
- Anaya, C.J.; Zander, H.J.; Graham, R.D.; Sankarasubramanian, V.; Lempka, S.F. Evoked potentials recorded from the spinal cord during neurostimulation for pain: A computational modeling study. Neuromodulation Technol. Neural Interface 2020, 23, 64–73. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Inclusion | Exclusion |
|---|---|
| Animals/humans treated with SCS or DRG stimulation. Computational studies about SCS/DRG were also allowed. | Other types of neuromodulation. |
| Supraspinal outcome measurements including, but not limited to, experimental pain measurements, brain imaging, histopathology. | No investigation of supraspinal outcome measurements. |
| Hypothesis about the supraspinal working mechanisms was explicitly stated in the article, including, but not limited to, expected theories, suggestions, hypotheses, assumptions or ideas. | No formulation of a possible underlying hypothesis. |
| Full-text (quasi) experimental or observational studies, case reports. | Systematic reviews and meta-analyses, narrative reviews, letters to the editor, conference abstracts, studies without available full-text version. |
| English, French or Dutch written. | Other languages. |
| Author/Year | Randomization Exposure Level | Allocation Concealment | Appropriate Comparison Groups | Confounding and Modifying Variables | Identical Experimental Cconditions | Blinding of Research Personnel | Attrition or Exclusion of Outcome Data | Confidence Exposure Characterization | Confidence Outcome Assessment | All Measured Outcomes Reported | Other Potential Threats | Quality Category (TIER-System) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aguilar J. 2011 [18] | − | − | + | − | ++ | ++ | + | ++ | ++ | 2nd tier | ||
| Atweh S.F. 1985 [19] | −− | NA | NA | NA | ++ | ++ | + | ++ | + | 2nd tier | ||
| Bantli H. 1975 [20] | −− | − | + | − | − | − | + | − | −− | 2nd tier | ||
| Barchini J. 2012 [5] | − | + | + | + | − | ++ | ++ | + | ++ | 1st tier | ||
| Dejongste M. 1998 [21] | + | + | + | + | ++ | ++ | + | + | ++ | 1st tier | ||
| Dembowsky K. 1985 [22] | −− | NA | + | − | ++ | + | + | − | ++ | 2nd tier | ||
| El-Khoury C. 2002 [23] | − | − | + | − | − | ++ | − | ++ | ++ | 2nd tier | ||
| Linderoth B. 1993 [24] | −− | NA | NA | − | − | + | + | ++ | ++ | 2nd tier | ||
| Maeda Y. 2009 [25] | + | − | + | + | − | ++ | ++ | ++ | ++ | 1st tier | ||
| Meuwissen K. 2020 [26] | + | + | + | − | ++ | ++ | + | ++ | ++ | 1st tier | ||
| Pawela C. 2017 [27] | − | − | + | − | − | ++ | − | + | ++ | 2nd tier | ||
| Quindlen-Hotek J. 2020 [28] | − | − | + | − | − | ++ | − | ++ | ++ | 2nd tier | ||
| Saade N. 2015 [29] | − | − | + | − | − | ++ | − | + | ++ | 2nd tier | ||
| Saade N. 1985 [30] | −− | NA | NA | NA | − | ++ | − | + | ++ | 3th tier | ||
| Saade N. 1985 [31] | −− | NA | NA | NA | − | ++ | + | ++ | ++ | 2nd tier | ||
| Song Z. 2013 [32] | −− | − | + | − | − | ++ | − | ++ | ++ | 2nd tier | ||
| Song Z. 2013 [33] | −− | − | + | − | − | ++ | − | ++ | ++ | 2nd tier | ||
| Song Z. 2009 [34] | −− | − | + | − | − | ++ | − | ++ | ++ | 2nd tier | ||
| Stiller C. 1995 [35] | − | − | + | − | − | ++ | + | + | ++ | 2nd tier | ||
| Tazawa T. 2015 [36] | −− | − | + | − | − | ++ | + | ++ | ++ | 2nd tier | ||
| Vallejo R. 2019 [37] | + | − | + | + | + | ++ | ++ | ++ | ++ | 1st tier |
| Author/Year | Design | Randomization Exposure level | Allocation Concealment | Appropriate Comparison Groups | Confounding and Modifying Variables | Identical Experimental Conditions | Blinding of Research Personnel and Subjects | Attrition or Exclusion of Outcome Data | Confidence Exposure Characterization | Confidence Outcome Assessment | All Measured Outcomes Reported | Other Potential Threats | Quality Category (TIER-System) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ahmed S. 2015 [38] | HCT | −− | − | − | ++ | ++ | − | ++ | ++ | 2nd tier | |||
| Blair R.D. 1975 [39] | case series | − | ++ | − | + | + | 2nd tier | ||||||
| Bocci T. 2018 [40] | HCT | + | − | − | − | ++ | + | ++ | ++ | 2nd tier | |||
| Buentjen L. 2020 [41] | HCT | − | −− | − | ++ | ++ | + | + | + | 2nd tier | |||
| de Andrade DC 2010 [42] | HCT | + | + | + | ++ | ++ | + | ++ | ++ | 1st tier | |||
| De Groote S. 2020 [43] | cohort | + | ++ | ++ | ++ | + | ++ | ++ | 1st tier | ||||
| De Groote S. 2020 [44] | cohort | + | ++ | ++ | ++ | + | ++ | ++ | 1st tier | ||||
| De Groote S. 2020 [45] | cohort | + | ++ | ++ | ++ | + | ++ | ++ | 1st tier | ||||
| De Ridder D. 2016 [46] | HCT | + | + | + | ++ | ++ | + | ++ | ++ | 1st tier | |||
| De Ridder D. 2013 [47] | HCT | + | + | + | − | ++ | + | − | ++ | 1st tier | |||
| Deogaonkar M. 2016 [48] | HCT | − | + | + | ++ | ++ | + | ++ | ++ | 1st tier | |||
| Gildenberg P.L. 1980 [49] | case series | − | + | + | −− | + | 2nd tier | ||||||
| Goudman L. 2019 [50] | cohort | + | − | ++ | ++ | −− | ++ | ++ | 2nd tier | ||||
| Goudman L. 2019 [51] | cohort | + | − | ++ | ++ | + | ++ | ++ | 2nd tier | ||||
| Kishima H. 2010 [52] | HCT | −− | NA | + | ++ | ++ | + | ++ | ++ | 1st tier | |||
| Kunitake A. 2005 [53] | case series | ++ | ++ | + | + | ++ | 1st tier | ||||||
| Larson S. 1974 [54] | HCT | −− | NA | − | − | − | ++ | + | + | + | 2nd tier | ||
| Lind A.L. 2016 [55] | HCT | −− | + | + | + | ++ | + | ++ | ++ | 1st tier | |||
| Mehta V. 2019 [56] | cohort | + | − | − | ++ | + | ++ | ++ | 2nd tier | ||||
| Modesti L.M. 1975 [57] | case series | − | ++ | + | + | + | 2nd tier | ||||||
| Moens M. 2013 [58] | HCT | + | + | + | ++ | ++ | + | ++ | ++ | 1st tier | |||
| Moens M. 2012 [59] | HCT | − | + | + | ++ | ++ | + | ++ | ++ | 1st tier | |||
| Morgalla M.H. 2019 [60] | cohort | + | − | − | ++ | + | ++ | ++ | 2nd tier | ||||
| Pahapill P.A. 2014 [61] | case series | − | − | + | + | + | 2nd tier | ||||||
| Polacek H. 2007 [62] | HCT | + | − | − | + | ++ | + | ++ | ++ | 1st tier | |||
| Royds J. 2020 [63] | cohort | + | − | − | ++ | + | ++ | ++ | 2nd tier | ||||
| Schlaier J.R. 2007 [64] | HCT | −− | NA | − | ++ | ++ | + | ++ | ++ | 2nd tier | |||
| Shimoji K. 1982 [65] | case series | − | − | + | ++ | + | 2nd tier | ||||||
| Schuh-Hofer S. 2018 [66] | HCT | − | − | − | ++ | ++ | −− | ++ | ++ | 2nd tier | |||
| Stancak A. 2008 [67] | HCT | + | − | − | ++ | ++ | + | ++ | ++ | 1st tier | |||
| Sufianov A.A. 2014 [68] | cohort | + | − | ++ | ++ | + | ++ | ++ | 2nd tier | ||||
| Weigel R. 2015 [69] | HCT | −− | − | − | + | ++ | − | ++ | ++ | 2nd tier | |||
| Yearwood T. 2019 [70] | HCT | ++ | ++ | + | ++ | ++ | + | ++ | ++ | 1st tier | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goudman, L.; De Groote, S.; Linderoth, B.; De Smedt, A.; Eldabe, S.; Duarte, R.V.; Moens, M. Exploration of the Supraspinal Hypotheses about Spinal Cord Stimulation and Dorsal Root Ganglion Stimulation: A Systematic Review. J. Clin. Med. 2021, 10, 2766. https://doi.org/10.3390/jcm10132766
Goudman L, De Groote S, Linderoth B, De Smedt A, Eldabe S, Duarte RV, Moens M. Exploration of the Supraspinal Hypotheses about Spinal Cord Stimulation and Dorsal Root Ganglion Stimulation: A Systematic Review. Journal of Clinical Medicine. 2021; 10(13):2766. https://doi.org/10.3390/jcm10132766
Chicago/Turabian StyleGoudman, Lisa, Sander De Groote, Bengt Linderoth, Ann De Smedt, Sam Eldabe, Rui V. Duarte, and Maarten Moens. 2021. "Exploration of the Supraspinal Hypotheses about Spinal Cord Stimulation and Dorsal Root Ganglion Stimulation: A Systematic Review" Journal of Clinical Medicine 10, no. 13: 2766. https://doi.org/10.3390/jcm10132766