Alterations in Small Intestine and Liver Morphology, Immunolocalization of Leptin, Ghrelin and Nesfatin-1 as Well as Immunoexpression of Tight Junction Proteins in Intestinal Mucosa after Gastrectomy in Rat Model

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Experimental Section
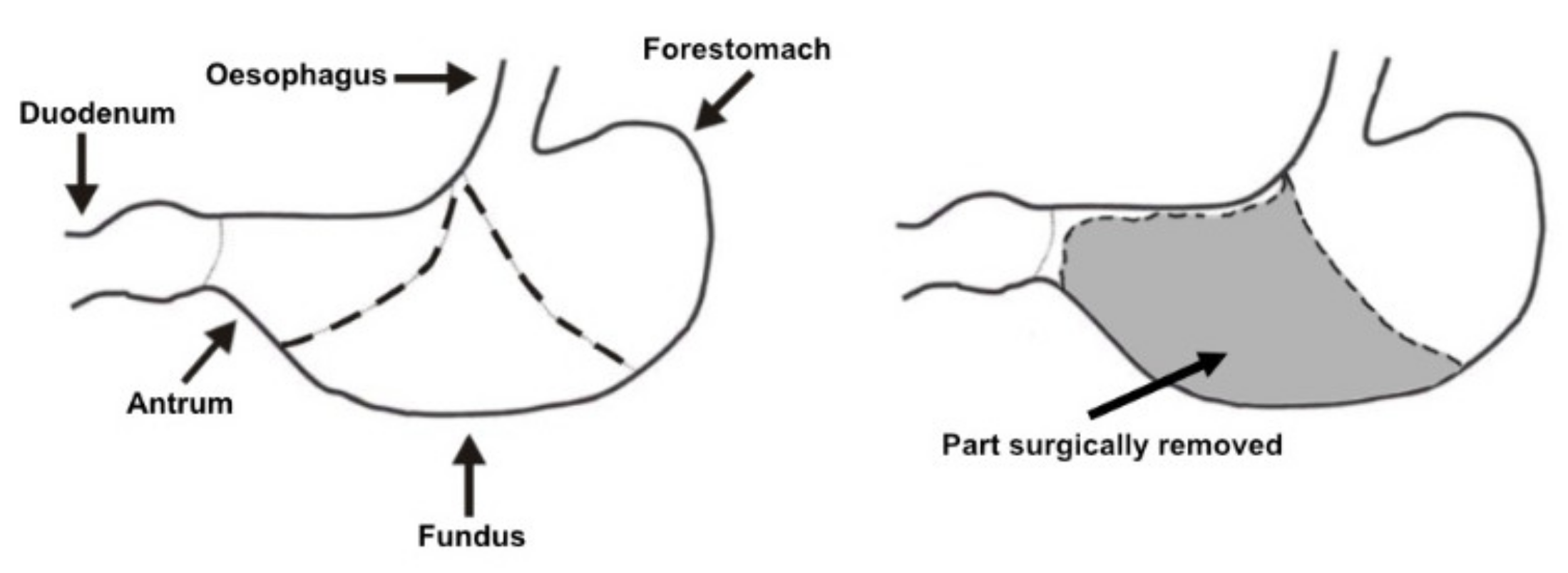
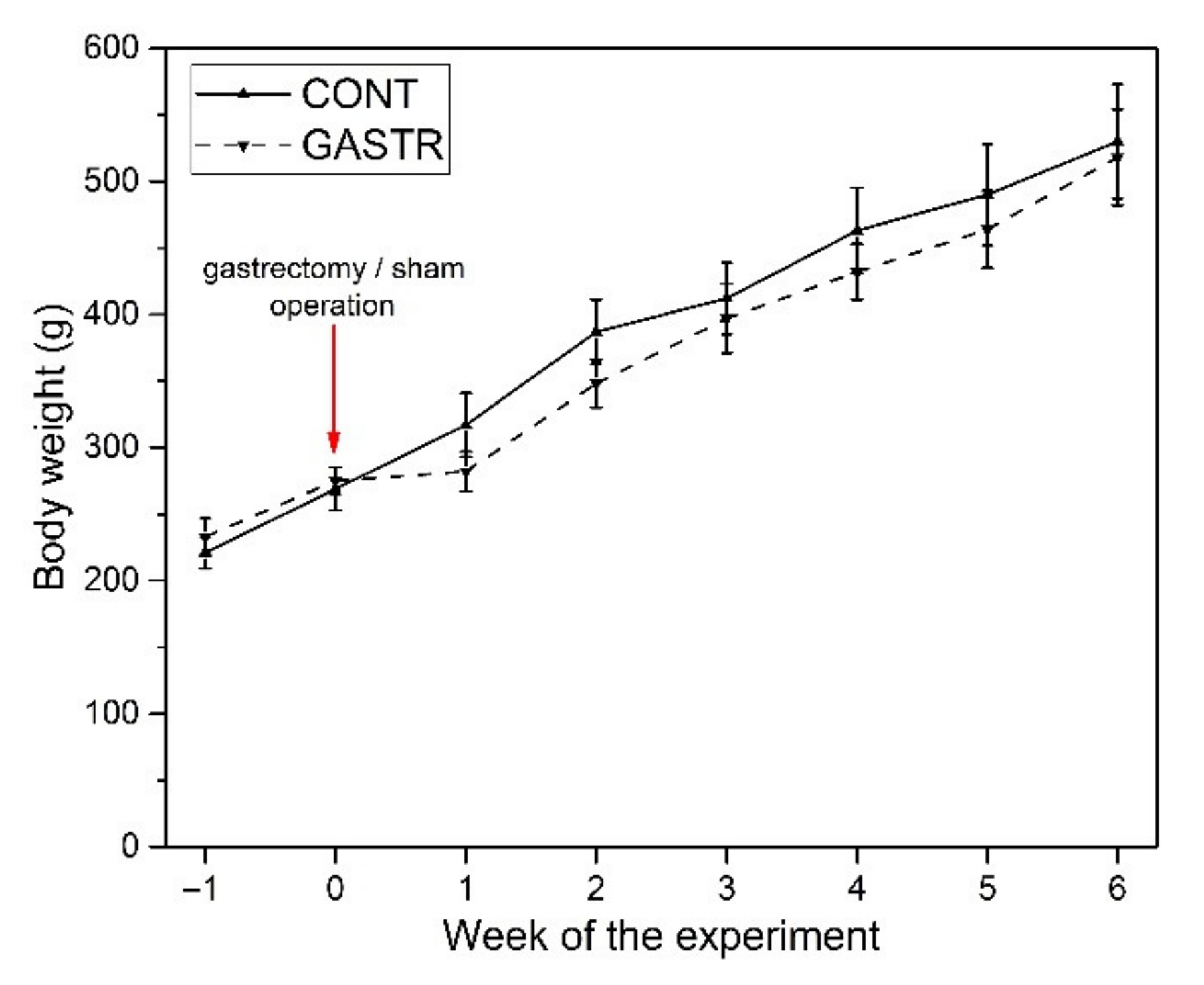
2.1. Experimental Design
2.2. Tissue Collection and Histomorphometry Analysis
2.3. Immunohistochemistry
2.4. Statistical Analysis
3. Results
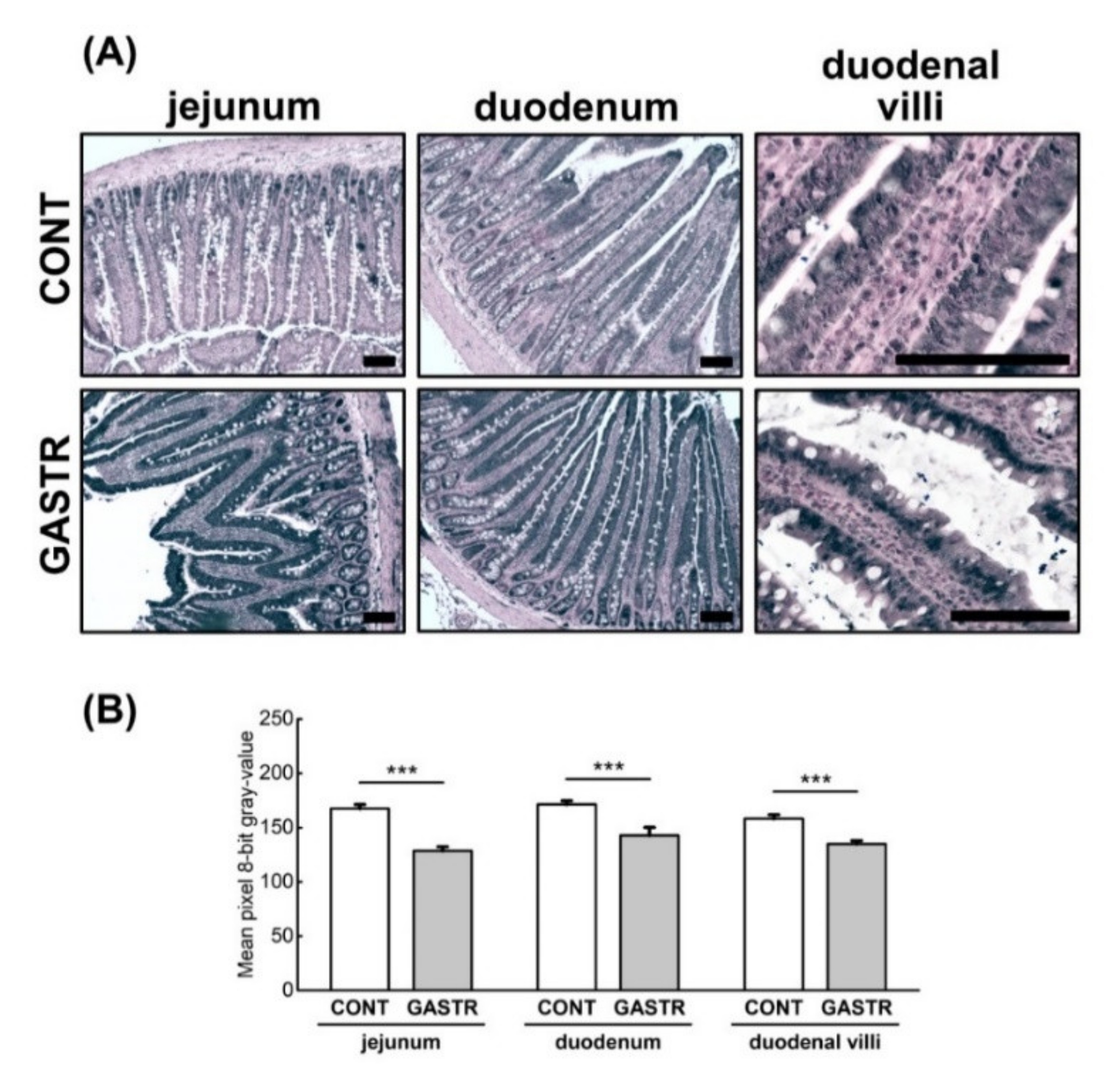
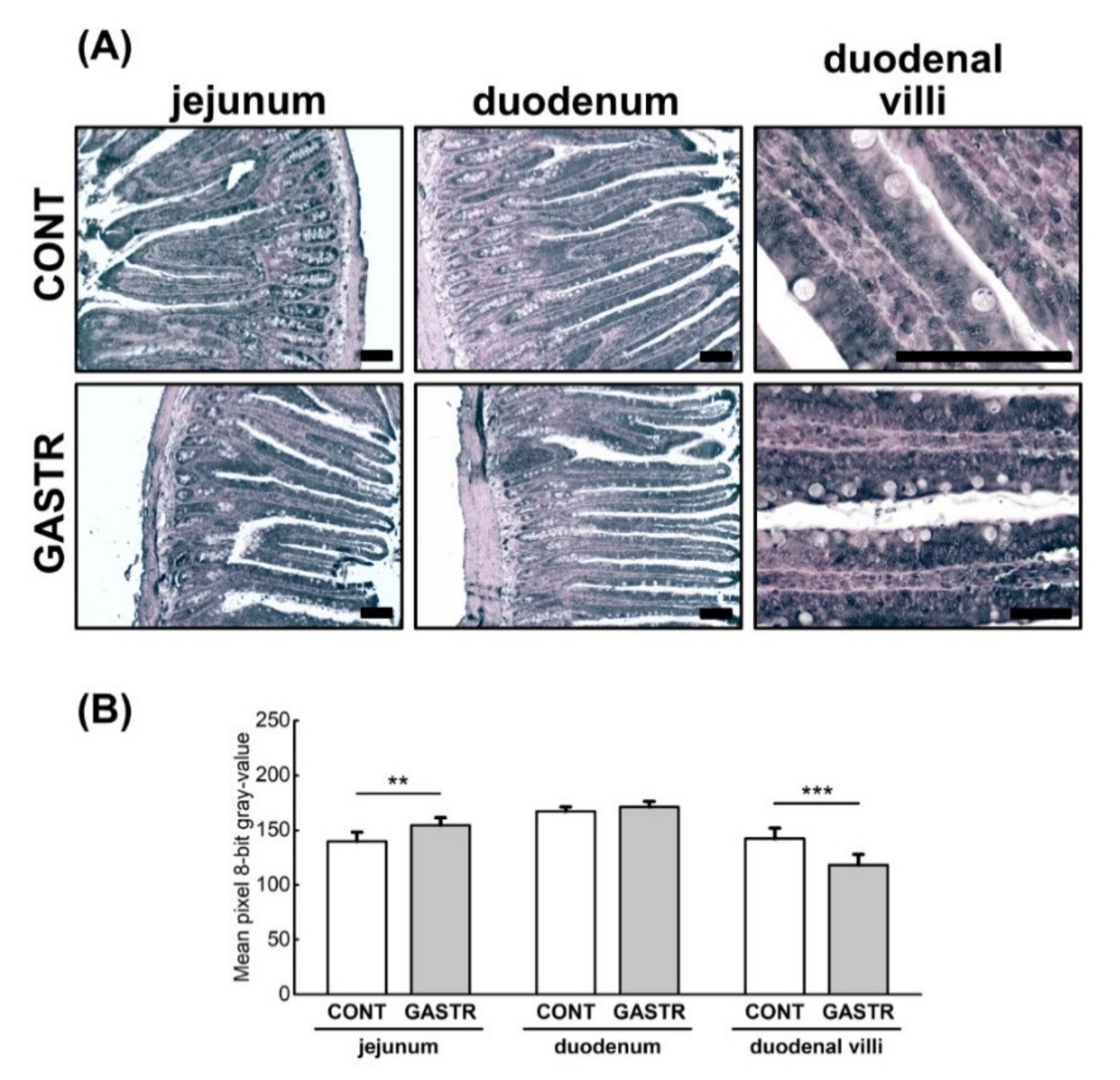
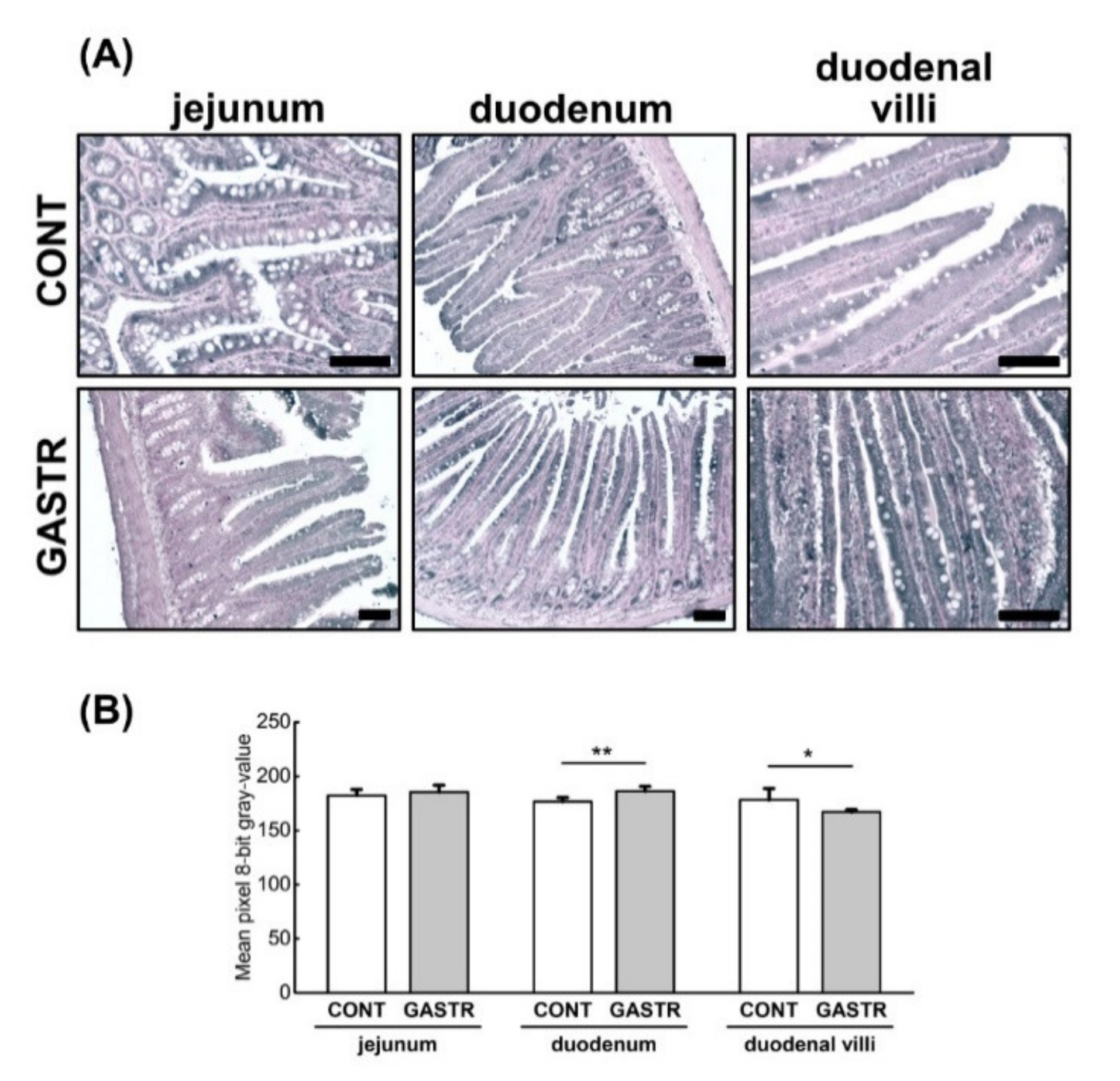
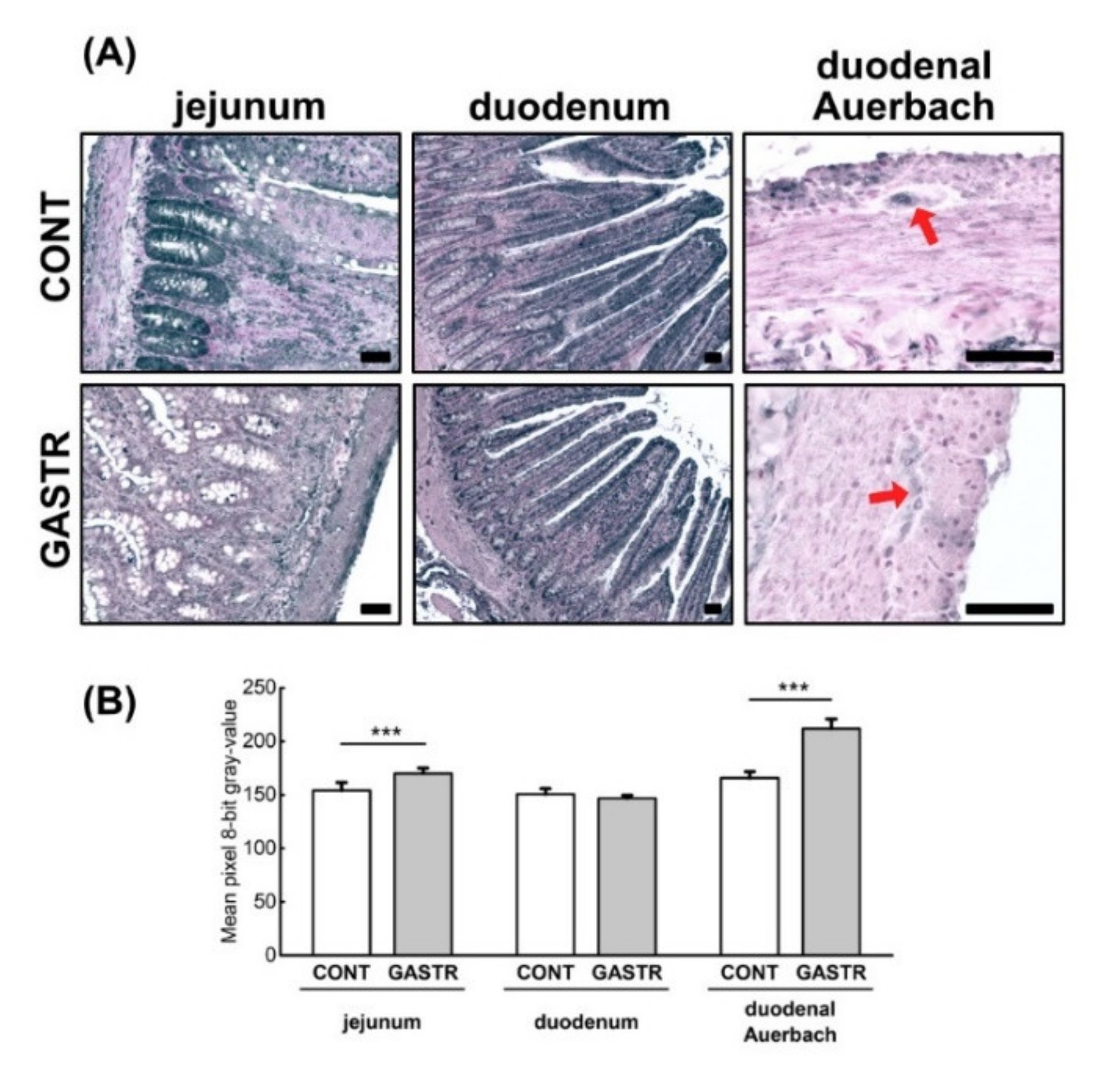
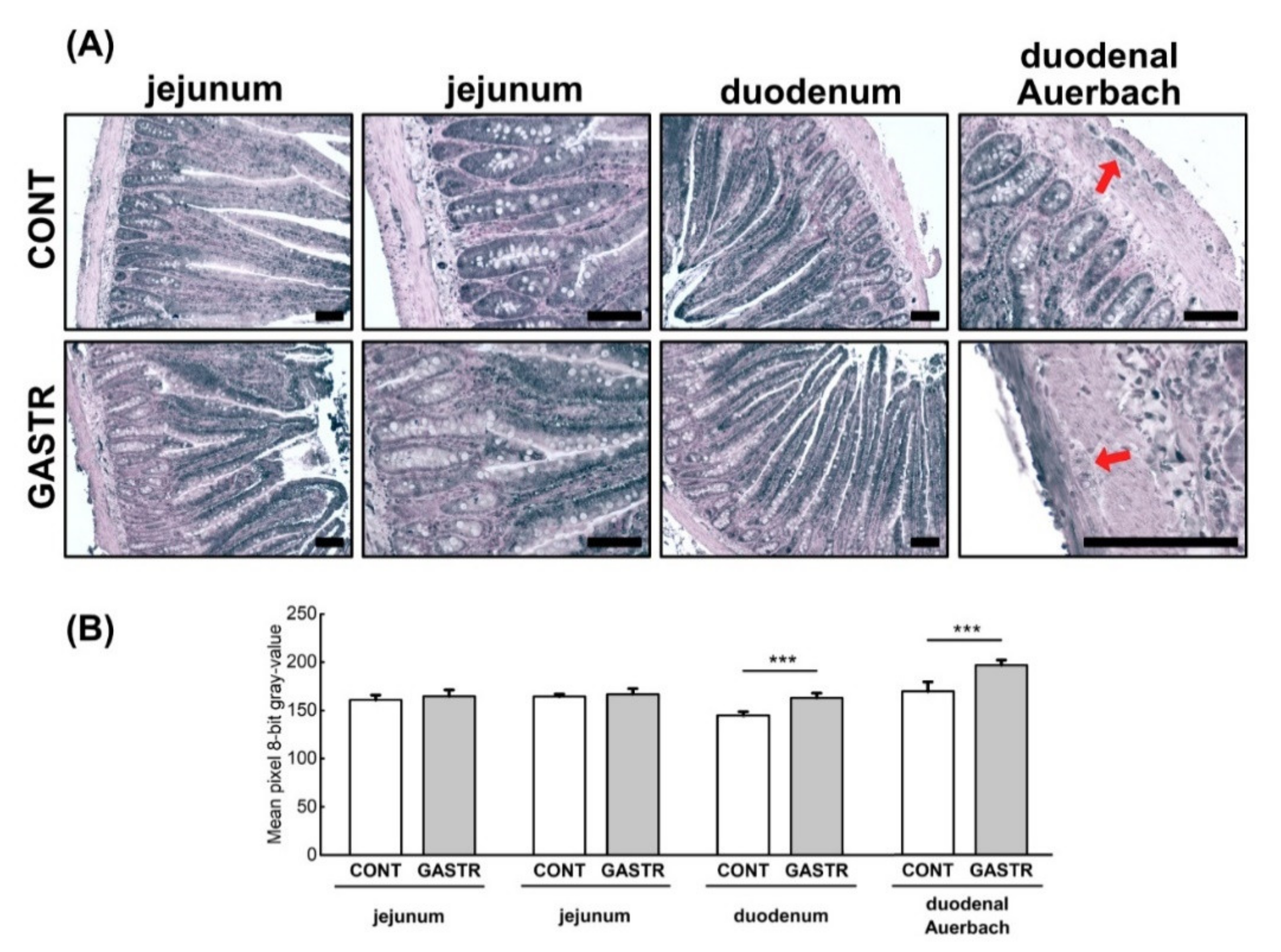
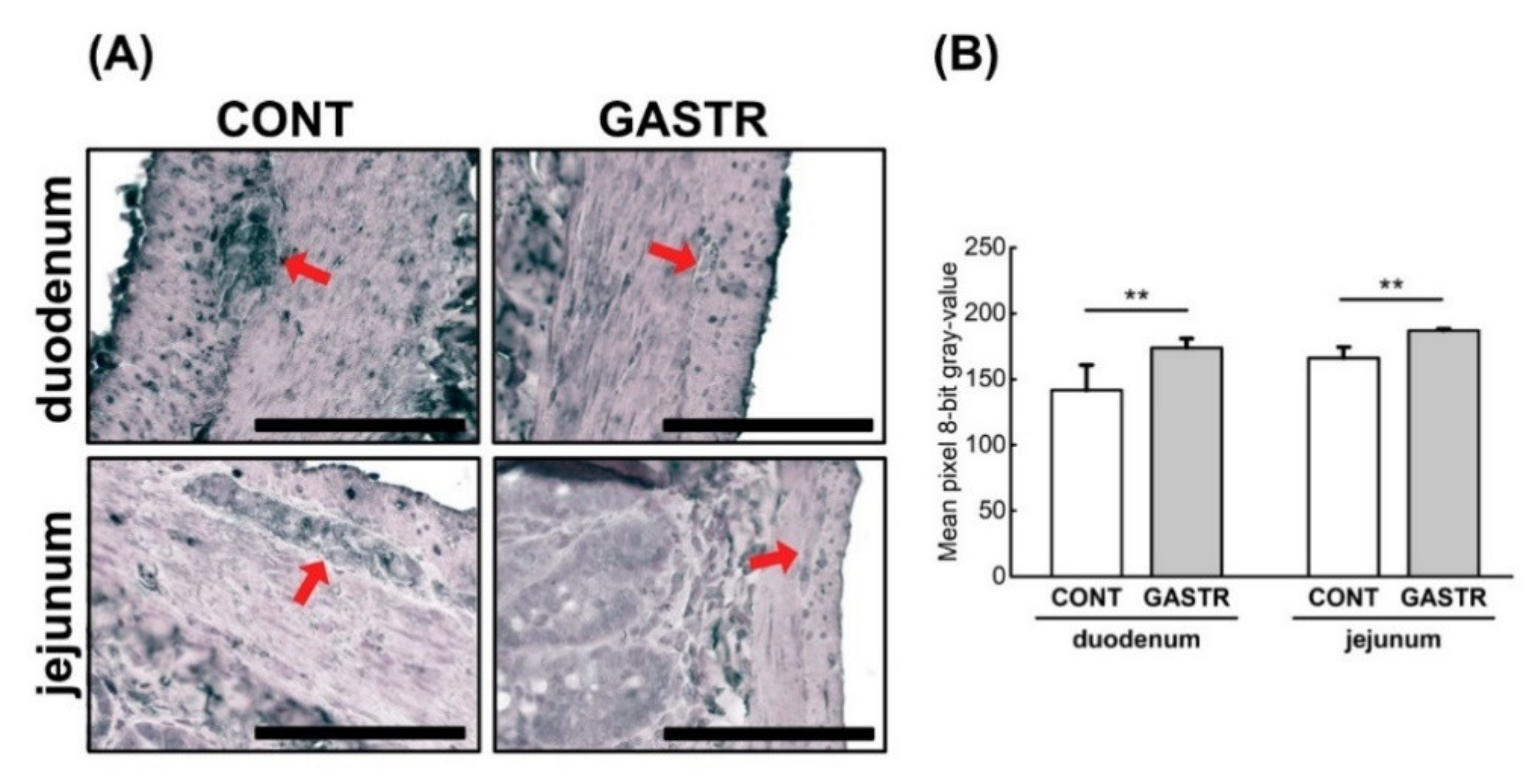
3.1. Morphology
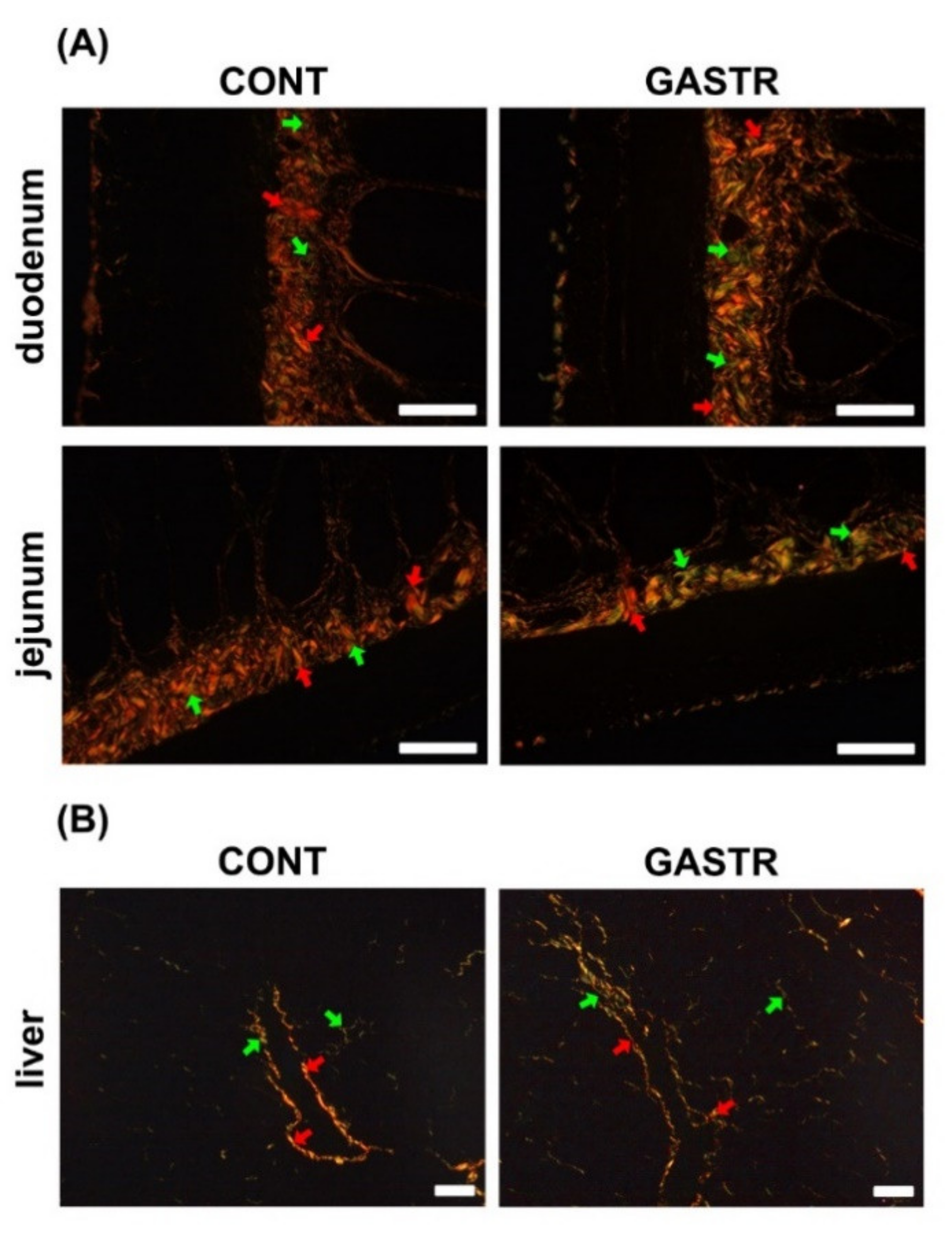
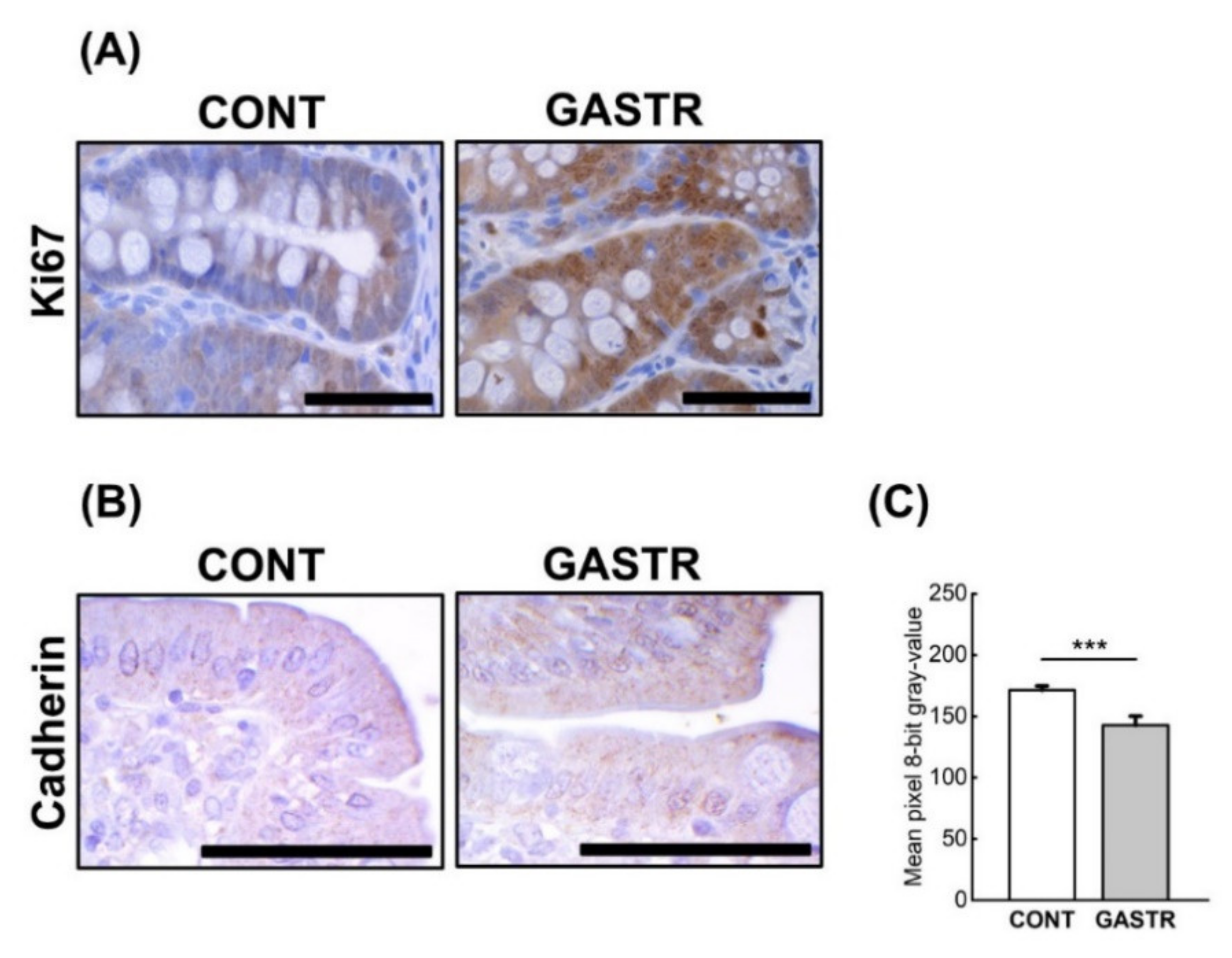
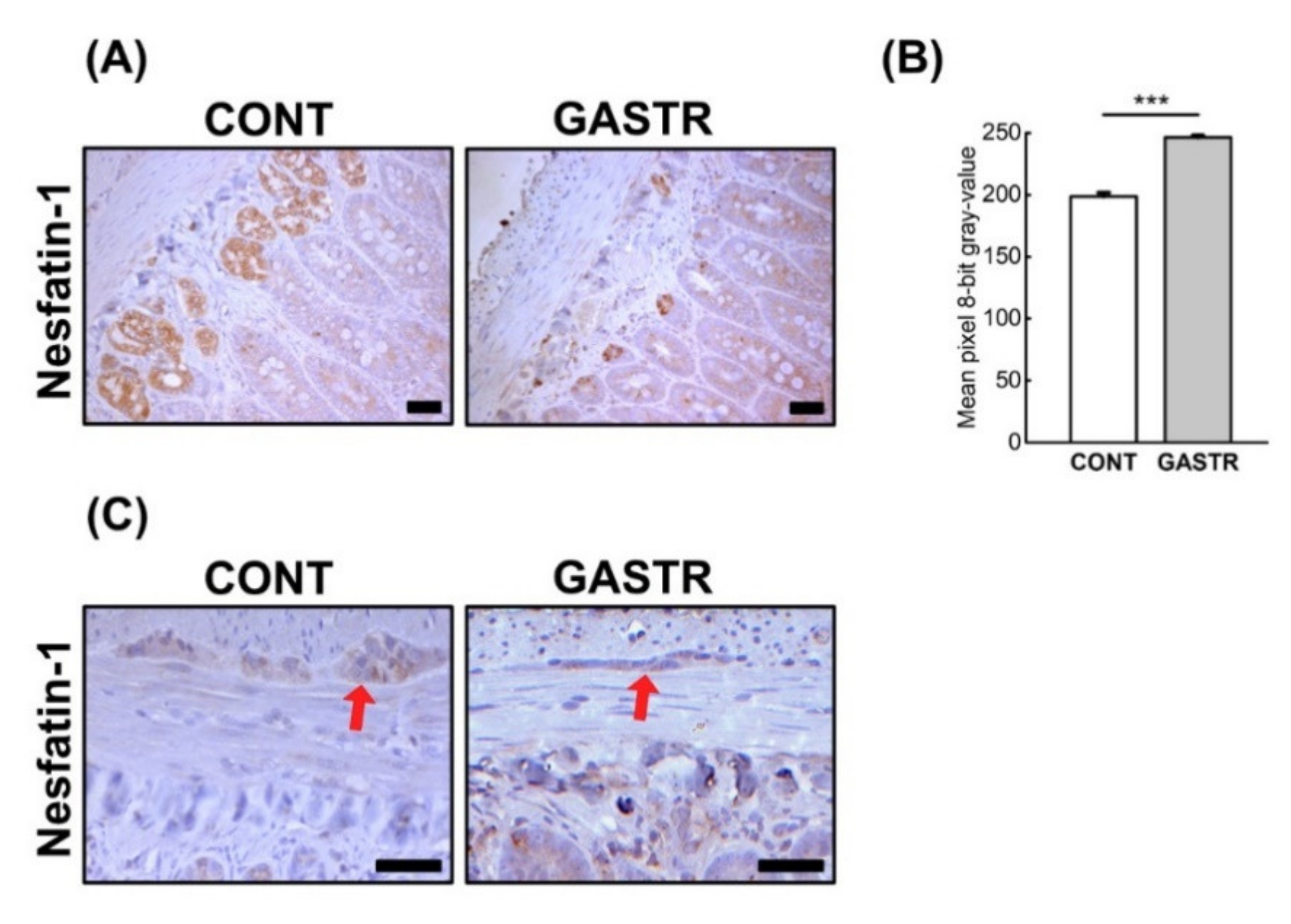
3.2. Immunolocalization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Piessen, G.; Triboulet, J.P.; Mariette, C. Reconstruction after gastrectomy: Which technique is best? J. Visc. Surg. 2010, 147, e273–e283. [Google Scholar] [CrossRef] [PubMed]
- Murawka, D.; Połom, K. Pathophysiological disorders, quality of life evaluation and recommendations after total resection of the stomach. Rep. Pract. Oncol. Radiother. 2008, 13, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Goldenshluger, M.; Goldenshluger, A.; Keinan-Boker, L.; Cohen, M.J.; Ben-Porat, T.; Gerasi, H.; Amun, M.; Abu-Gazala, M.; Khalaileh, A.; Mintz, Y.; et al. Postoperative outcomes, weight loss predictors, and late gastrointestinal symptoms following laparoscopic sleeve gastrectomy. J. Gastrointest. Surg. 2017, 21, 2009–2015. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Lee, H.-J.; Choi, Y.S.; Kim, T.H.; Hun, Y.-J.; Suh, Y.-S.; Kong, S.-H.; Yang, H.-K. Postoperative quality of life after total gastrectomy compared with partial gastrectomy: Longitudinal evaluation by European Organization for Research and Treatment of Cancer-OG25 and STO22. J. Gastric Cancer 2016, 16, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Trung, V.N.; Yamamoto, H.; Furukawa, A.; Yamaguchi, T.; Murata, S.; Yoshimura, M.; Murakami, Y.; Sato, S.; Otani, H.; Ugi, S.; et al. Enhanced intestinal motility during oral glucose tolerance test after laparoscopic sleeve gastrectomy: Preliminary results using cine magnetic resonance imaging. PLoS ONE 2013, 8, e65739. [Google Scholar] [CrossRef] [Green Version]
- Faulconbridge, L.F.; Ruparel, K.; Loughead, J.; Alliosn, K.C.; Hesson, L.A.; Fiabricatore, A.N.; Rochette, A.; Ritter, A.; Hopson, R.D.; Sarwe, D.B.; et al. Changes in neural responsivity to highly palatable foods following roux–en–y gastric bypass, sleeve gastrectomy, or weight stability: An fMRI study. Obesity 2016, 24, 1054–1060. [Google Scholar] [CrossRef]
- Yousseif, A.; Emmanuel, J.; Karra, E.; Millet, Q.; Elkalaawy, M.; Jenkinson, A.D.; Hashemi, M.; Adamo, M.; Finer, N.; Finnes, A.G.; et al. Differential effects of laparoscopic sleeve gastrectomy and laparoscopic gastric bypass on appetite, circulating acyl-ghrelin, peptide YY3-36 and active GLP-1 levels in non–diabetic humans. Obes. Surg. 2014, 24, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Myronovych, A.; Kirby, M.; Ryan, K.K.; Zhang, W.; Jha, P.; Setchell, K.D.; Dexheimer, P.J.; Aronow, B.; Seeley, R.J.; Kohli, R. Vertical sleeve gastrectomy reduces hepatic steatosis while increasing serum bile acids in a weight-loss-independent manner. Obesity 2014, 22, 390–400. [Google Scholar] [CrossRef]
- Damms–Machado, A.; Mitra, S.; Schollenberger, A.E.; Kramer, K.M.; Meile, T.; Königsrainer, A.; Huson, D.H.; Bischoff, S.C. Effects of surgical and dietary weight loss therapy for obesity on gut microbiota composition and nutrient absorption. BioMed Res. Int. 2015, 2015, 806248. [Google Scholar] [CrossRef]
- Dib, N.; Kiciak, A.; Pietrzak, P.; Ferenc, K.; Jaworski, P.; Kapica, M.; Tarnowski, W.; Zabielski, R. Early-effect of bariatric surgery (Scopinaro method) on intestinal hormones and adipokines in insulin resistant Wistar rat. J. Physiol. Pharmacol. 2013, 64, 571–577. [Google Scholar]
- Li, B.; Lu, Y.; Srikant, C.B.; Gao, Z.H.; Liu, J.L. Intestinal adaptation and Reg gene expression induced by antidiabetic duodenal–jejunal bypass surgery in Zucker fatty rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G635–G645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Roux, C.W.; Borg, C.; Wallis, K.; Vincent, R.P.; Bueter, M.; Goodlad, R.; Ghatei, M.A.; Bloom, S.R.; Aylwin, S.J.B. Gut hypertrophy after gastric bypass is associated with increased glucagon–like peptide 2 and intestinal crypt cell proliferation. Ann. Surg. 2010, 252, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Taqi, E.; Wallace, L.E.; de Heuvel, E.; Chelikani, P.K.; Zheng, H.; Berthoud, H.-R.; Holst, J.J.; Sigalet, D.L. The influence of nutrients, biliary-pancreatic secretions, and systemic trophic hormones on intestinal adaptation in a Roux-en-Y bypass model. J. Pediatr. Surg. 2010, 45, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Seeley, R.J.; Chambers, A.P.; Sandoval, D.A. The role of gut adaptation in the potent effects of multiple bariatric surgeries on obesity and diabetes. Cell Metab. 2015, 21, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Holzer, P. Implications of tachykinins and calcitonin gene-related peptide in inflammatory bowel disease. Digestion 1998, 59, 269–283. [Google Scholar] [CrossRef]
- Gonkowski, S. Substance P as a neuronal factor in the enteric nervous system of the porcine descending colon in physiological conditions and during selected pathogenic processes. Biofactors 2013, 39, 542–551. [Google Scholar] [CrossRef]
- Vasina, V.; Barbara, G.; Talamonti, L.; Stanghellini, V.; Corinaldesi, R.; Tonini, M.; De Ponti, F.; De Giorgio, R. Enteric neuroplasticity evoked by inflammation. Auton. Neurosci. 2006, 126–127, 264–272. [Google Scholar] [CrossRef]
- Lehto-Axtelius, D.; Stenström, M.; Johnell, O. Osteopenia after gastrectomy, fundectomy or antrectomy: An experimental study in the rat. Regul. Pept. 1998, 78, 41–50. [Google Scholar] [CrossRef]
- Surve, V.V.; Andersson, N.; Lehto-Axtelius, D.; Hakanson, R. Comparison of osteopenia after gastrectomy, ovariectomy and prednisolone treatment in the young female rat. Acta Orthop. Scand. 2001, 72, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Dobrowolski, P.; Piersiak, T.; Surve, V.V.; Kruszewska, D.; Gawron, A.; Pacuska, P.; Hakanson, R.; Pierzynowski, S.P. Dietary α-ketoglutarate reduces gastrectomy-evoked loss of calvaria and trabecular bone in female rats. Scand. J. Gastroenterol. 2008, 43, 551–558. [Google Scholar] [CrossRef]
- Klinge, B.; Lehto-Axtelius, D.; Akerman, M.; Hakanson, R. Structure of calvaria after gastrectomy: An experimental study in the rat. Scand. J. Gastroenterol. 1995, 30, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Rudyk, H.; Tomaszewska, E.; Arciszewski, M.B.; Muszyński, S.; Tomczyk-Warunek, A.; Dobrowolski, P.; Donaldson, J.; Brezvyn, O.; Kotsyumbas, I. Histomorphometrical changes in intestine structure and innervation following experimental fumonisins intoxication in male Wistar rats. Pol. J. Vet. Sci. 2020, 23, 77–88. [Google Scholar] [PubMed]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques, 7th ed.; Churchill Livingstone: New York, NY, USA, 2013. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Kisielinski, K.; Willis, S.; Prescher, A.; Klosterhalfen, B.; Schumpelick, V. A simple new method to calculate small intestine absorptive surface in the rat. Clin. Exp. Med. 2002, 2, 131–135. [Google Scholar] [CrossRef]
- Kasacka, I.; Piotrowska, Ż.; Niezgoda, M.; Lewnadowska, A.; Łebkowski, W. Ageing-related changes in the levels of β-catenin, CacyBP/SIP, galectin-3 and immunoproteasome subunit LMP7 in the heart of men. PLoS ONE 2020, 15, e0229462. [Google Scholar] [CrossRef] [PubMed]
- Bolton, J.S.; Conway, W.C., II. Postgastrectomy syndromes. Surg. Clin. N. Am. 2011, 91, 1105–1122. [Google Scholar] [CrossRef]
- Shaw, D.; Gohil, K.; Basson, M.D. Intestinal mucosal atrophy and adaptation. World J. Gastroenterol. 2012, 18, 6357–6375. [Google Scholar] [CrossRef]
- Drozdowski, L.A.; Clandinin, M.T.; Thomson, A.B.R. Morphological, kinetic, membrane biochemical and genetic aspects of intestinal enteroplasticity. World J. Gastroenterol. 2009, 15, 774–787. [Google Scholar] [CrossRef]
- Burant, C.F.; Flink, S.; DePaoli, A.M.; Chen, J.; Lee, W.S.; Hediger, M.A.; Buse, J.B.; Chang, E.B. Small intestine hexose transport in experimental diabetes. Increased transporter mRNA and protein expression in enterocytes. J. Clin. Investig. 1994, 93, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Sturm, A.; Layer, P.; Goebell, H.; Dignass, A.U. Short–bowel syndrome: An update on the therapeutic approach. Scand. J. Gastroenterol. 1997, 32, 289–296. [Google Scholar] [CrossRef]
- Cavin, J.B.; Couvelard, A.; Lebtahi, R.; Ducroc, R.; Arapis, K.; Voitellier, E.; Cluzeaud, F.; Gillard, L.; Hourseau, M.; Mikail, N.; et al. Differences in alimentary glucose absorption and intestinal disposal of blood glucose following Roux-en-Y gastric bypass vs sleeve gastrectomy. Gastroenterology 2016, 150, 454–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumphrey, M.B.; Hao, Z.; Townsend, R.L.; Patterson, L.M.; Berthould, H.R. Sleeve gastrectomy does not cause hypertrophy and reprogramming of intestinal glucose metabolism in rats. Obes. Surg. 2015, 25, 1468–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavin, J.B.; Voitellier, E.; Cluzeaud, F.; Kapel, N.; Marmuse, J.-P.; Chevallier, J.-M.; Msika, S.; Bado, A.; Le Gall, M. Malabsorption and intestinal adaptation after one anastomosis gastric bypass compared to Roux–en–Y gastric bypass in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G492–G500. [Google Scholar] [CrossRef] [PubMed]
- Habegger, K.M.; Al-Massadi, O.; Heppner, K.M.; Myronovych, A.; Holland, J.; Berger, J.; Yi, C.X.; Gao, Y.; Lehti, M.; Ottaway, N.; et al. Duodenal nutrient exclusion improves metabolic syndrome and stimulates villus hyperplasia. Gut 2014, 63, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Kohli, R.; Kirby, M.; Setchell, K.D.; Jha, P.; Klustaitis, K.; Woollett, L.A.; Pfluger, P.T.; Balistreri, W.F.; Tso, P.; Jandacek, R.J.; et al. Intestinal adaptation after ileal interposition surgery increases bile acid recycling and protects against obesity-related comorbidities. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G652–G660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauronen, J.; Pakarinen, M.P.; Kuusanmäki, P.; Savilahti, E.; Vento, P.; Paavonen, T.; Halttunen, J. Intestinal adaptation after massive proximal small-bowel resection in the pig. Scand. J. Gastroenterol. 1998, 33, 152–158. [Google Scholar] [PubMed]
- McDuffie, L.A.; Bucher, B.T.; Erwin, C.R.; Wakeman, D.; White, F.V.; Warner, B.W. Intestinal adaptation after small bowel resection in human infants. J. Pediatr. Surg. 2011, 46, 1045–1051. [Google Scholar] [CrossRef] [Green Version]
- Iversen, B.M.; Schjonsby, H.; Skagen, D.W.; Solhaug, J.H. Intestinal adaptation after jejuno-ileal bypass operation for massive obesity. Eur. J. Clin. Investig. 1976, 6, 355–360. [Google Scholar] [CrossRef]
- Kapica, M.; Puzio, I. Effect of fundectomy, antrectomy and gastrectomy on pancreatic and brush border enzyme activity in rats. Med. Weter. 2018, 74, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Lutz, T.A.; Bueter, M. The use of rat and mouse models in bariatric surgery experiments. Front. Nutr. 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautron, L.; Zechner, J.F.; Aguirre, V. Vagal innervation patterns following Roux-en-Y gastric bypass in the mouse. Int. J. Obes. 2013, 37, 1603–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGregor, I.; Parent, J.; Meyer, J.H. Gastric emptying of liquid meals and pancre-atic and biliary secretion after subtotal gastrectomy or truncal vagotomy andpyloroplasty in man. Gastroenterology 1977, 72, 95–205. [Google Scholar]
- Domínguez-Muñoz, J.E. Pancreatic enzyme replacement therapy: Exocrinepancreatic insufficiency after gastrointestinal surgery. HPB 2009, 11 (Suppl. S3), 3–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leth, R.D.; Abrahamsson, H.; Kilander, A.; Lundell, L.R. Malabsorption of fat after partial gastric resection. A study of pathophysiologic mechanisms. Eur. J. Surg. 1991, 157, 205–208. [Google Scholar]
- Hansen, L.; Lampert, S.; Mineo, H.; Holst, J.J. Neural regulation of glucagon-like peptide-1 secretion in pigs. Am. J. Physiol. Endocrinol. Metab. 2004, 287, e939–e947. [Google Scholar] [CrossRef] [Green Version]
- Capella, C.; Solcia, E.; Vassallo, G. Identification of six types of endocrine cells in the gastrointestinal mucosa of the rabbit. Arch. Histol. Jap. 1969, 30, 479–495. [Google Scholar] [CrossRef] [Green Version]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Prinz, C.; Kajimura, M.; Scott, D.R.; Mercier, F.; Helander, H.F.; Sachs, G. Histamine secretion from rat enterochromaffinlike cells. Gastroenterology 1993, 105, 449–461. [Google Scholar] [CrossRef]
- Stengel, A.; Goebel, M.; Yakubov, I.; Wang, L.; Witcher, D.; Coskun, T.; Tache, Y.; Sachs, G.; Lambrecht, N.W. Identification and characterization of nesfatin-1 immunoreactivity in endocrine cell types of the rat gastric oxyntic mucosa. Endocrinology 2009, 150, 232–238. [Google Scholar] [CrossRef] [Green Version]
- Stengel, A.; Hofmann, T.; Goebel-Stengel, M.; Lembke, V.; Ahnis, A.; Elbelt, U.; Lambrecht, N.W.; Ordemann, J.; Klapp, B.F.; Kobelt, P. Ghrelin and NUCB2/nesfatin-1 are expressed in the same gastric cell and differentially correlated with body mass index in obese subjects. Histochem. Cell Biol. 2013, 139, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.L.; Fujimura, M.; Hayashi, N.; Nakamura, T.; Fujimiya, M. Mechanisms in regulating the release of serotonin from the perfused rat stomach. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G1099–G1105. [Google Scholar] [CrossRef] [PubMed]
- Gonkowski, S.; Rychlik, A.; Nowicki, M.; Nieradka, R.; Bulc, M.; Całka, J. A population of nesfatin 1-like immunoreactive (LI) cells in the mucosal layer of the canine digestive tract. Res. Vet. Sci. 2012, 93, 1119–1121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.Q.; Li, X.L.; Jiang, C.Y.; Lin, L.; Shi, R.H.; Chen, J.D.; Oomura, Y. Expression of nesfatin-1/NUCB2 in rodent digestive system. World J. Gastroenterol. 2010, 16, 1735–1741. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Goebel, M.; Wang, L.; Rivier, J.; Kobelt, P.; Monnikes, H.; Lambrecht, N.W.; Taché, Y. Central nesfatin-1 reduces dark–phase food intake and gastric emptying in rats: Differential role of corticotropin–releasing factor2 receptor. Endocrinology 2009, 150, 4911–4919. [Google Scholar] [CrossRef]
- Watanabe, A.; Mochiki, E.; Kimura, A.; Kogure, N.; Tanai, M.; Ogawa, A.; Toyomasu, Y.; Ogata, K.; Ohno, T.; Suzuki, H.; et al. Nesfatin-1 suppresses gastric contractions and inhibits interdigestive migrating contractions in conscious dogs. Dig. Dis. Sci. 2015, 60, 1595–1602. [Google Scholar] [CrossRef]
- Xia, Z.F.; Fritze, D.M.; Li, J.Y.; Chai, B.; Zhang, C.; Zhang, W.; Mulholland, M.W. Nesfatin-1 inhibits gastric acid secretion via a central vagal mechanism in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, 570–577. [Google Scholar] [CrossRef] [Green Version]
- Wren, A.M.; Seal, L.J.; Cohen, M.A.; Brynes, A.E.; Frost, G.S.; Murphy, K.G.; Dhillo, W.S.; Ghatei, M.A.; Bloom, S.R. Ghrelin enhances appetite and increases food intake in humans. J. Clin. Endocrinol. Metab. 2001, 86, 5992. [Google Scholar] [CrossRef]
- Masuda, Y.; Tanaka, T.; Inomata, N.; Ohnuma, N.; Tanaka, S.; Itoh, Z.; Hosoda, H.; Kojima, M.; Kangawa, K. Ghrelin stimulates gastric acid secretion and motility in rats. Biochem. Biophys. Res. Commun. 2000, 276, 905–908. [Google Scholar] [CrossRef]
- Grönberg, M.; Tsolakis, A.V.; Magnusson, L.; Janson, E.T.; Saras, J. Distribution of obestatin and ghrelin in human tissues: Immunoreactive cells in the gastrointestinal tract, pancreas, and mammary glands. J. Histochem. Cytochem. 2008, 56, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.M.; Furnes, M.W.; Stenstrom, B.; Kulseng, B.; Chen, D. Characterization of obestatin– and ghrelin–producing cells in the gastrointestinal tract and pancreas of rats: An immunohistochemical and electron–microscopic study. Cell Tissue Res. 2008, 331, 575–587. [Google Scholar] [CrossRef]
- Cinti, S.; Matteis, R.D.; Pico, C.; Ceresi, E.; Obrador, A.; Maffeis, C.; Oliver, J.; Palou, A. Secretory granules of endocrine and chief cells of human stomach mucosa contain leptin. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 789–793. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Guyenet, S.J.; Schwartz, M.W. Regulation of food intake, energy balance, and body fat mass: Implications for the pathogenesis and treatment of obesity. J. Clin. Endocrinol. Metab. 2012, 97, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Pérez, A.; Vilariño-García, T.; Fernández-Riejos, P.; Martín-González, J.; Segura-Egea, J.J.; Sánchez-Margalet, V. Role of leptin as a link between metabolism and the immune system. Cytokine Growth Factor Rev. 2017, 35, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Morton, N.M.; Emilsson, V.; Liu, Y.L.; Cawthorne, M.A. Leptin action in intestinal cells. J. Biol. Chem. 1998, 273, 26194–26201. [Google Scholar] [CrossRef] [Green Version]
- Ducroc, R.; Guilmeau, S.; Akasbi, K.; Devaud, H.; Buyse, M.; Bado, A. Luminal leptin induces rapid inhibition of active intestinal absorption of glucose mediated by sodium-glucose cotransporter 1. Diabetes 2005, 54, 348–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhri, O.; Small, C.; Bloom, S. Gastrointestinal hormones regulating appetite. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1187–1209. [Google Scholar] [CrossRef] [Green Version]
- Mumphrey, M.B.; Patterson, L.M.; Zheng, H.; Berthoud, H. Roux-en-Y gastric bypass surgery increases number but not density of CCK-, GLP-1-, 5-HT-, and neurotensin-expressing enteroendocrine cells in rats. Neurogastroenterol. Motil. 2013, 25, e70–e79. [Google Scholar] [CrossRef] [Green Version]
- Nergård, B.J.; Lindqvist, A.; Gilason, H.G.; Groop, L.; Ekelund, M.; Wierup, N.; Hedenbro, J.L. Mucosal glucagon-like peptide-1 and glucose-dependent insulinotropic polypeptide cell numbers in the super-obese human foregut after gastric bypass. Surg. Obes. Relat. Dis. 2015, 11, 1237–1246. [Google Scholar] [CrossRef]
- Costa, M.; Furness, J.B. The origins, pathways and terminations of neurons with VIP-like immunoreactivity in the guinea-pig small intestine. Neuroscience 1983, 8, 665–676. [Google Scholar] [CrossRef]
- Varricchio, E.; Russolillo, M.G.; Russo, F.; Lomnardi, V.; Paolucci, M.; Maruccio, L. Expression and immunohistochemical detection of nesfatin-1 in the gastrointestinal tract of Casertana pig. Acta Histochem. 2014, 116, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Yarandi, S.S.; Hebbar, G.; Sauer, C.G.; Cole, C.R.; Ziegler, T.R. Diverse roles of leptin in the gastrointestinal tract: Modulation of motility, absorption, growth, and inflammation. Nutrition 2011, 27, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Edholm, T.; Levin, F.; Hellstrom, P.M.; Schmidt, P.T. Ghrelin stimulates motility in the small intestine of rats through intrinsic cholinergic neurons. Regul. Pept. 2004, 121, 25–30. [Google Scholar] [CrossRef]
- Ramesh, N.; Mortazavi, S.; Unniappan, S. Nesfatin-1 stimulates glucagon-like peptide-1 and glucose-dependent insulinotropic polypeptide secretion from STC-1 cells in vitro. Biochem. Biophys. Res. Commun. 2015, 462, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, N.; Mortazavi, S.; Unniappan, S. Nesfatin-1 stimulates cholecystokinin and suppresses peptide YY expression and secretion in mice. Biochem. Biophys. Res. Commun. 2016, 472, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Teive, M.B.; Russi, R.F.; Vieira, D.S.; Teive, A.M.; Costa, A.; d’Acampora, A.J. Quantitative immunohistochemical analysis of duodenal ghrelin cells after sleeve gastrectomy in Wistar rats. Acta Cir. Bras. 2012, 27, 595–599. [Google Scholar] [CrossRef] [Green Version]
- Banks, M.R.; Farthing, M.J.; Robberecht, P.; Burleigh, D.E. Antisecretory actions of a novel vasoactive intestinal polypeptide (VIP) antagonist in human and rat small intestine. Br. J. Pharmacol. 2005, 144, 994–1001. [Google Scholar] [CrossRef] [Green Version]
- Fung, C.; Unterweger, P.; Parry, L.J.; Bornstein, J.V.; Foong, P.P. VPAC1 receptors regulate intestinal secretion and muscle contractility by activating cholinergic neurons in guinea pig jejunum. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G748–G758. [Google Scholar] [CrossRef]
- Severi, C.; Tattoli, I.; Corleto, V.D.; Masselli, M.A.; Trisolini, P.; Delle Fave, G. Vasoactive intestinal peptide receptor subtypes and signalling pathways involved in relaxation of human stomach. Neurogastroenterol. Motil. 2006, 18, 1009–1018. [Google Scholar] [CrossRef]
- Neunlist, M.; Toumi, F.; Oreschkova, T.; Denis, M.; Leborgne, J.; Laboisse, C.L.; Galmiche, J.-P.; Jarry, A. Human ENS regulates the intestinal epithelial barrier permeability and a tight junction-associated protein ZO-1 via VIPergic pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1028–G1036. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, F.; Krueger, D.; Schemann, M. Leptin excites enteric neurons of guinea–pig submucous and myenteric plexus. Neurogastroenterol. Motil. 2011, 23, e165–e170. [Google Scholar] [CrossRef]
- Voinot, F.; Fischer, C.; Crenner, F.; Schmidt, C.; Angel, F. Leptin modulates enteric neurotransmission in the rat proximal colon: An in vitro study. Regul. Pept. 2013, 185, 73–78. [Google Scholar]
- Li, X.; Zhang, A.Q.; Lin, L.; Shi, R.; Chen, J. Expression of nesfatin–1/NUCB2 in the rodent digestive system. Gastroenterology 2010, 138 (Suppl. S1), S573. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Yeager, M.; Harris, A.L. Gap junction channel structure in the early 21st century: Facts and fantasies. Curr. Opin. Cell Biol. 2007, 19, 521–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Fanning, A.S.; Mitic, L.L.; Anderson, J.M. Transmembrane proteins in the tight junction barrier. J. Am. Soc. Nephrol. 1999, 10, 1337–1345. [Google Scholar]
- Green, K.J.; Getsios, S.; Troyanovsky, S.; Godsel, L.M. Intercellular junction assembly, dynamics, and homeostasis. CHS Perspect. Biol. 2010, 2, a000125. [Google Scholar] [CrossRef] [Green Version]
- Gumbiner, B.M. Cell adhesion: The molecular basis of tissue architecture and morphogenesis. Cell 1996, 84, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The tight junction protein ZO–1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niessen, C.M. Tight junctions/adherens junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2525–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchard, C.; Moreau, F.; Chevalier, J.; Ayer, A.; Garcon, D.; Arnaud, L.; Pais de Barros, J.-P.; Gautier, T.; Neunlist, M.; Cariou, B.; et al. Sleeve gastrectomy alters intestinal permeability in diet–induced obese mice. Obes. Surg. 2017, 27, 2590–2598. [Google Scholar] [CrossRef] [PubMed]
- Casselbrant, A.; Elias, E.; Fändriks, L.; Wallenius, V. Expression of tight–junction proteins in human proximal small intestinal mucosa before and after Roux-en-Y gastric bypass surgery. Surg. Obes. Relat. Dis. 2015, 11, 45–53. [Google Scholar] [CrossRef]
- Yang, P.-J.; Yg, W.-S.; Nien, H.-C.; Chen, C.-N.; Lee, P.-H.; Yu, L.C.-H.; Lin, M.-T. Duodenojejunal bypass leads to altered gut microbiota and strengthened epithelial barriers in rats. Obes. Surg. 2016, 26, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Rao, Z.; Laing, B.T.; Bunner, W.; Landry, T.; Prete, A.; Yuan, Y.; Zhang, Z.T.; Huang, H. Vertical sleeve gastrectomy improves liver and hypothalamic functions in obese mice. J. Endocrinol. 2019, 241, 135–147. [Google Scholar] [CrossRef]
- Bower, G.; Toma, T.; Harling, L.; Jiao, L.R.; Efthimiou, E.; Darzi, A.; Athanasiou, T.; Ashrafian, H. Bariatric surgery and non-alcoholic fatty liver disease: A systematic review of liver biochemistry and histology. Obes. Surg. 2015, 25, 2280–2989. [Google Scholar] [CrossRef]
- Keleidari, B.; Mahmoudieh, M.; Gorgi, K.; Sheikhbahaei, E.; Shahabi, S. Hepatic failure after bariatric surgery: A systematic review. Hepat. Mon. 2019, 19, e86078. [Google Scholar] [CrossRef] [Green Version]
- Marciniak, R.; Majewski, P.; Biczysko, M.; Banasiewicz, T.; Woźniak, A.; Drews, M. Effects of gastrectomy or colectomy on liver metabolism and liver morphology in an experimental rat model. Med. Sci. Monit. 2004, 10, BR34–BR40. [Google Scholar]
- Ozutemiz, C.; Obuz, F.; Taylan, A.; Atila, K.; Bora, S.; Ellidokuz, H. Volume change of segments II and III of the liver after gastrectomy in patients with gastric cancer. Diagn. Interv. Radiol. 2016, 22, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Jiang, X.; Jian, W.; Zheng, L.; Lu, L.; Zheng, C. Comparing the effectiveness of total gastrectomy and gastric bypass on glucose metabolism in diabetic rats. Obes. Surg. 2016, 26, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, M.; Dezaki, K.; Koizumi, M.; Kurashina, K.; Hosoya, Y.; Lefor, A.K.; Sata, N.; Yada, T. Total gastrectomy-induced reductions in food intake and weight are counteracted by rikkunshito by attenuating glucagon-like peptide-1 elevation in rats. Surgery 2016, 159, 1342–1350. [Google Scholar] [CrossRef] [PubMed]
- Stefater, M.A.; Pérez-Tilve, D.; Chambers, A.P.; Wilson-Pérez, H.E.; Sandoval, D.A.; Berger, J.; Toure, M.; Tschöep, M.; Woods, S.C.; Seeley, R.J. Sleeve gastrectomy induces loss of weight and fat mass in obese rats, but does not affect leptin sensitivity. Gastroenterology 2010, 138, 2426–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grayson, B.E.; Schneider, K.M.; Woods, S.C.; Seeley, R.J. Improved rodent maternal metabolism but reduced intrauterine growth after vertical sleeve gastrectomy. Sci. Transl. Med. 2013, 5, 199ra112. [Google Scholar] [CrossRef] [Green Version]
- Takiguchi, S.; Takata, A.; Murakami, K.; Miyazaki, Y.; Yanagimoto, Y.; Kurokawa, Y.; Takahashi, T.; Mori, M.; Doki, Y. Clinical application of ghrelin administration for gastric cancer patients undergoing gastrectomy. Gastric Cancer 2014, 17, 200–205. [Google Scholar] [CrossRef]
- Puzio, I.; Kapica, M.; Bieńko, M.; Radzki, R.; Pawłowska, M.; Tymicki, G. Fundectomy, antrectomy and gastrectomy influence densitometric, tomographic and mechanical bone properties as well as serum ghrelin and nesfatin-1 levels in rats. Med. Weter. 2014, 70, 604–609. [Google Scholar]
- Dogan, U.; Bulbuller, N.; Cakir, T.; Habibi, M.; Mayir, B.; Koc, U.; Aslaner, A.; Ellidag, H.Y.; Gomceli, I. Nesfatin-1 hormone levels in morbidly obese patients after laparoscopic sleeve gastrectomy. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1023–1031. [Google Scholar]
- Yang, K.; Zhang, X.; Zhou, Y.; Chen, F.; Shen, M.; Wang, Y. Changes in serum nesfatin-1 after laparoscopic sleeve gastrectomy are associated with improvements in nonalcoholic fatty liver disease. Diabetes Metab. Syndr. Obes. 2020, 13, 1459–1464. [Google Scholar] [CrossRef]
- Lee, W.-J.; Chen, C.-Y.; Ser, K.-H.; Hong, K.; Chen, S.-C.; Lee, P.-C.; Liao, Y.-D.; Lee, S.-D. Differential influences of gastric bypass and sleeve gastrectomy on plasma nesfatin-1 and obestatin levels in patients with type 2 diabetes mellitus. Curr. Pharm. Des. 2013, 19, 5830–5835. [Google Scholar] [CrossRef]
- Chambers, A.P.; Kirchner, H.; Wilson-Perez, H.E.; Willency, J.A.; Hale, J.E.; Gaylinn, B.D.; Thorner, M.O.; Pfluger, P.T.; Gutierrez, J.A.; Tschöp, M.H.; et al. The effects of vertical sleeve gastrectomy in rodents are ghrelin independent. Gastroenterology 2013, 144, 50–52.e5. [Google Scholar] [CrossRef] [Green Version]
- Bose, M.; Teixeira, J.; Olivan, B.; Bawa, B.; Arias, S.; Machineni, S.; Pi-Sunyer, F.X.; Scherer, P.E.; Laferrère, B. Weight loss and incretin responsiveness improve glucose control independently after gastric bypass surgery. J. Diabetes 2010, 2, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.P.; Jessen, L.; Ryan, K.K.; Sisley, S.; Wilson-Perez, H.E.; Stefater, M.A.; Gaitonde, S.G.; Sorrell, J.E.; Toure, M.; Berger, J.; et al. Weight-independent changes in blood glucose homeostasis after gastric bypass or vertical sleeve gastrectomy in rats. Gastroenterology 2011, 141, 950–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Duodenum | Jejunum | ||
|---|---|---|---|---|
| CONT | GAST | CONT | GAST | |
| Myenteron longitudinal lamina thickness, μm | 34.4 ± 8.85 | 29.2 ± 6.06 | 15.7 ± 4.6 | 27.2 ± 3.0 *** |
| Myenteron transversal lamina thickness, μm | 52.5 ± 12.1 | 46.0 ± 9.1 | 22.7 ± 3.7 | 40.7 ± 3.7 *** |
| Submucosa thickness, μm | 40.3 ± 8.2 | 25.4 ± 7.8 ** | 15.4 ± 7.6 | 31.7 ± 5.3 ** |
| Mucosa thickness, μm | 885 ± 73 | 825 ± 68 | 723 ± 31 | 825 ± 68 ** |
| Villus length, µm | 628 ± 34 | 622 ± 53 | 492 ± 59 | 460 ± 43 |
| Villus thickness, µm | 81.4 ± 10.3 | 81.8 ± 7.8 | 76.0 ± 9.8 | 81.3 ± 13.5 |
| Total number of villi, /mm | 9.1 ± 1.5 | 8.4 ± 0.8 | 9.9 ± 0.9 | 9.1 ± 1.0 |
| Villus epithelium thickness, µm | 30.9 ± 4.7 | 31.7 ± 4.4 | 28.9 ± 5.5 | 31.0 ± 6.7 |
| Enterocyte number, /100 µm of villus | 13.5 ± 1.4 | 14.8 ± 1.4 | 16.2 ± 1.9 | 13.5 ± 1.8* |
| Total crypts number, /mm | 13.2 ± 2.7 | 11.6 ± 1.6 | 16.9 ± 3.9 | 17.3 ± 3.3 |
| Active crypts number, /mm | 4.4 ± 1.5 | 2.6 ± 0.7 * | 5.7 ± 1.2 | 5.4 ± 1.9 |
| Inactive crypts number, /mm | 8.8 ± 2.6 | 9.0 ± 1.7 | 10.3 ± 3.8 | 11.9 ± 4.0 |
| Crypt depth, μm | 168 ± 27 | 146 ± 30 | 148 ± 19 | 136 ± 17 |
| Crypt width, µm | 52.7 ± 9.1 | 55.3 ± 8.6 | 44.3 ± 10.4 | 47.1 ± 9.0 |
| Intestine absorptive surface, µm2 | 4.2 ± 0.6 | 4.9 ± 1.0 | 3.9 ± 0.6 | 4.0 ± 0.5 |
| Immature collagen, % | 4.1 ± 2.2 | 7.6 ± 2.4 * | 3.5 ± 1.5 | 10.8 ± 2.6 *** |
| Ki index | 0.61 ± 0.06 | 0.68 ± 0.06 | 0.39 ± 0.13 | 0.56 ± 0.07 * |
| Ki number, /0.01 mm2 of the gland surface | 9.08 ± 2.01 | 8.11 ± 1.08 | 6.6 ± 1.8 | 10.1 ± 2.8 * |
| Parameter | Duodenum | Jejunum | ||
|---|---|---|---|---|
| CONT | GAST | CONT | GAST | |
| Auerbach plexus | ||||
| Area, µm2 | 828 ± 143 | 656 ± 155 | 585 ± 125 | 979 ± 149 *** |
| Perimeter, µm | 137 ± 67 | 131 ± 37 | 120 ± 55 | 179 ± 74 |
| Mean Feret diameter, µm | 41.0 ± 33.4 | 39.3 ± 40.9 | 36.2 ± 9.5 | 53.6 ± 12.1 * |
| Mean diameter, µm | 25.4 ± 10.2 | 20.2 ± 9.3 | 12.9 ± 6.5 | 13.0 ± 5.1 |
| Min diameter, µm | 15.9 ± 4.6 | 11.8 ± 3.6 | 21.9 ± 10.1 | 24.7 ± 12.3 |
| Sphericity | 0.31 ± 0.19 | 0.30 ± 0.22 | 0.21 ± 0.18 | 0.13 ± 0.11 |
| The number of the ganglia, /mm | 5.5 ± 0.9 | 5.6 ± 1.2 | 3.0 ± 1.1 | 3.1 ± 0.6 |
| Meissner plexus | ||||
| Area, µm2 | 384 ± 114 | 372 ± 142 | 355 ± 103 | 372 ± 158 |
| Perimeter, µm | 82 ± 25 | 82 ± 32 | 80 ± 22 | 79 ± 34 |
| Mean Feret diameter, µm | 24.6 ± 7.4 | 24.5 ± 9.5 | 24 ± 6.7 | 23.9 ± 10.2 |
| Mean diameter, µm | 20.8 ± 6.3 | 20.1 ± 6.3 | 19.3 ± 5.6 | 19.4 ± 7.2 |
| Min diameter, µm | 14.6 ± 4.9 | 14.2 ± 4.4 | 12.8 ± 4.5 | 13.7 ± 5.3 |
| Sphericity | 0.29 ± 0.19 | 0.31 ± 0.05 | 0.31 ± 0.18 | 0.33 ± 0.24 |
| The number of the ganglia, /mm | 1.5 ± 0.2 | 4.4 ± 0.8 *** | 3.8 ± 0.9 | 3.9 ± 0.1 |
| Parameter | CONT | GAST |
|---|---|---|
| Total cell number, /mm2 | 2084 ± 189 | 2647 ± 138 *** |
| Total hepatocyte number, /mm2 | 1565 ± 75 | 2149 ± 55 *** |
| Total hepatocyte nucleus number, /mm2 | 1651 ± 65 | 2257 ± 62 *** |
| Mononuclear hepatocytes nucleus number, /mm2 | 1479 ± 119 | 2041 ± 157 *** |
| Binuclear hepatocytes nucleus number, /mm2 | 86 ± 28 | 108 ± 29 |
| Non-hepatocyte cell number, /mm2 | 519 ± 99 | 488 ± 51 |
| Immature collagen, % | 15.8 ± 4.5 | 39.8 ± 5.3 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puzio, I.; Muszyński, S.; Dobrowolski, P.; Kapica, M.; Pawłowska-Olszewska, M.; Donaldson, J.; Tomaszewska, E. Alterations in Small Intestine and Liver Morphology, Immunolocalization of Leptin, Ghrelin and Nesfatin-1 as Well as Immunoexpression of Tight Junction Proteins in Intestinal Mucosa after Gastrectomy in Rat Model. J. Clin. Med. 2021, 10, 272. https://doi.org/10.3390/jcm10020272
Puzio I, Muszyński S, Dobrowolski P, Kapica M, Pawłowska-Olszewska M, Donaldson J, Tomaszewska E. Alterations in Small Intestine and Liver Morphology, Immunolocalization of Leptin, Ghrelin and Nesfatin-1 as Well as Immunoexpression of Tight Junction Proteins in Intestinal Mucosa after Gastrectomy in Rat Model. Journal of Clinical Medicine. 2021; 10(2):272. https://doi.org/10.3390/jcm10020272
Chicago/Turabian StylePuzio, Iwona, Siemowit Muszyński, Piotr Dobrowolski, Małgorzata Kapica, Marta Pawłowska-Olszewska, Janine Donaldson, and Ewa Tomaszewska. 2021. "Alterations in Small Intestine and Liver Morphology, Immunolocalization of Leptin, Ghrelin and Nesfatin-1 as Well as Immunoexpression of Tight Junction Proteins in Intestinal Mucosa after Gastrectomy in Rat Model" Journal of Clinical Medicine 10, no. 2: 272. https://doi.org/10.3390/jcm10020272