Bone Marrow Ts65Dn Trisomy-Induced Changes in Platelet Functionality and Lymphocytopenia Do Not Impact Atherosclerosis Susceptibility in Mice


Abstract
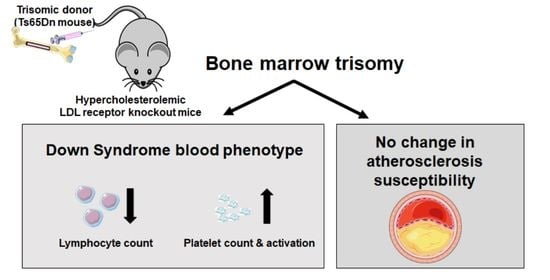
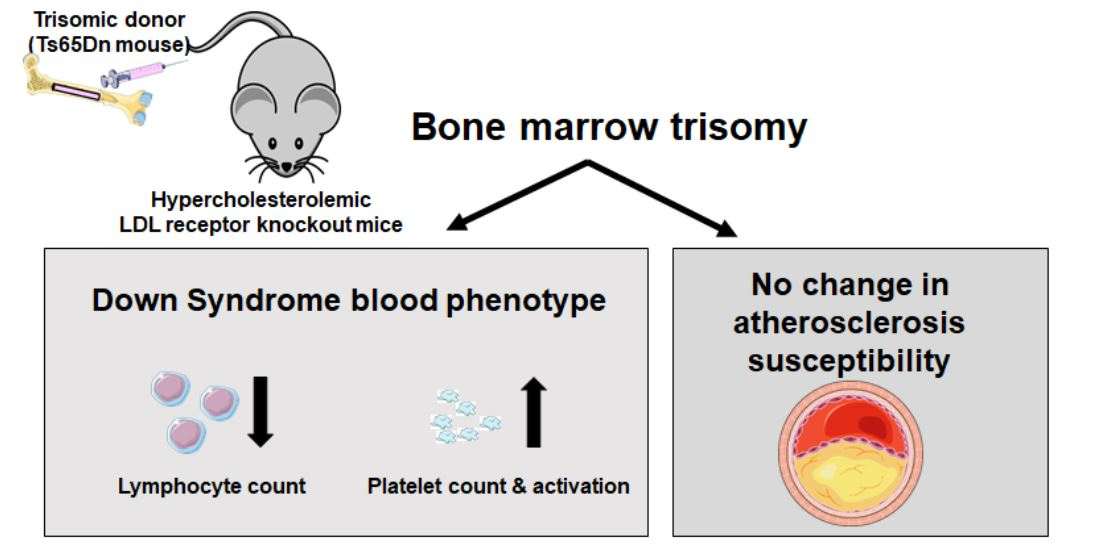
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice and Bone Marrow Transplantation
2.2. Generation of Bone Marrow-Derived Macrophages
2.3. Analysis of Gene Expression by Real-Time Quantitative PCR
2.4. Blood Cell Analysis
2.5. Histological Analysis of Atherosclerosis
2.6. Platelet-Related Studies
2.7. Data Analysis
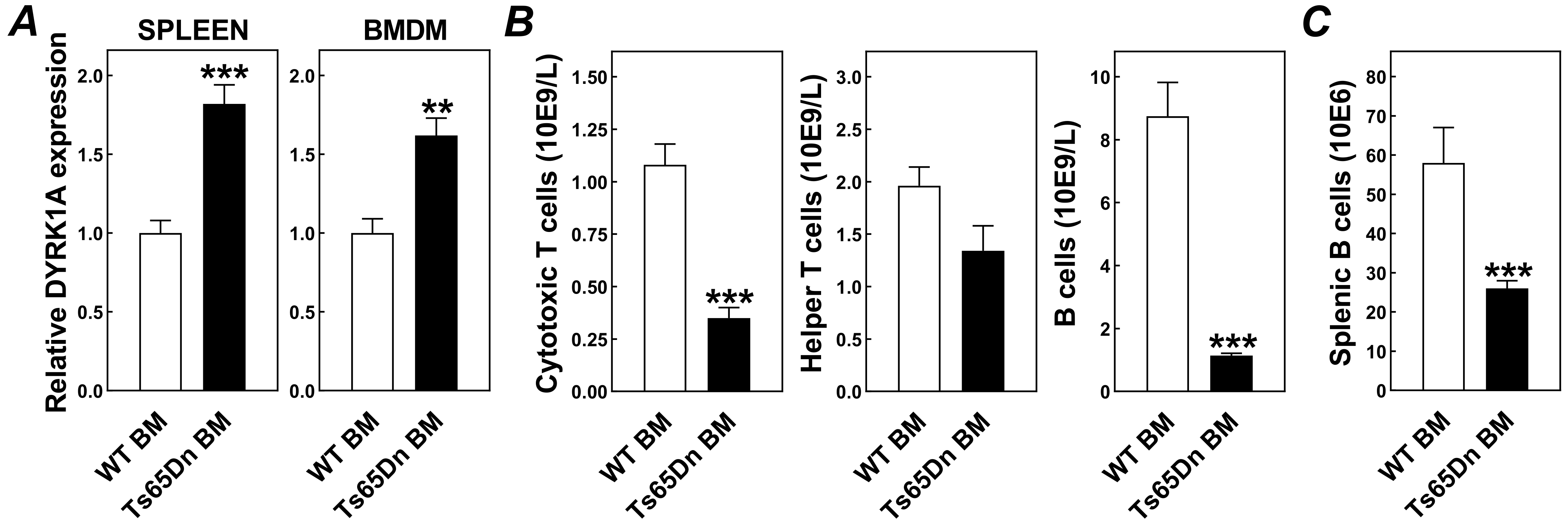
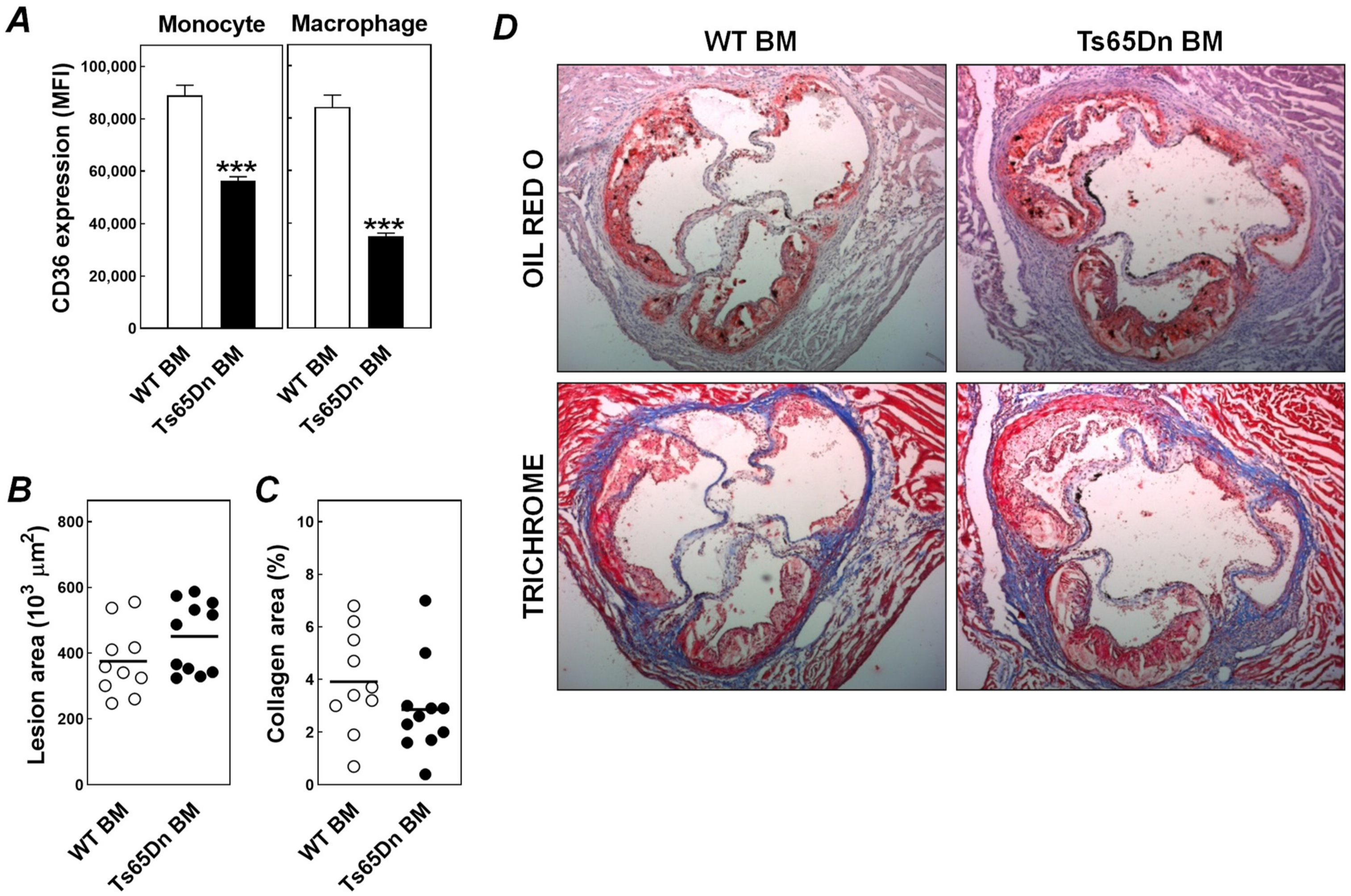
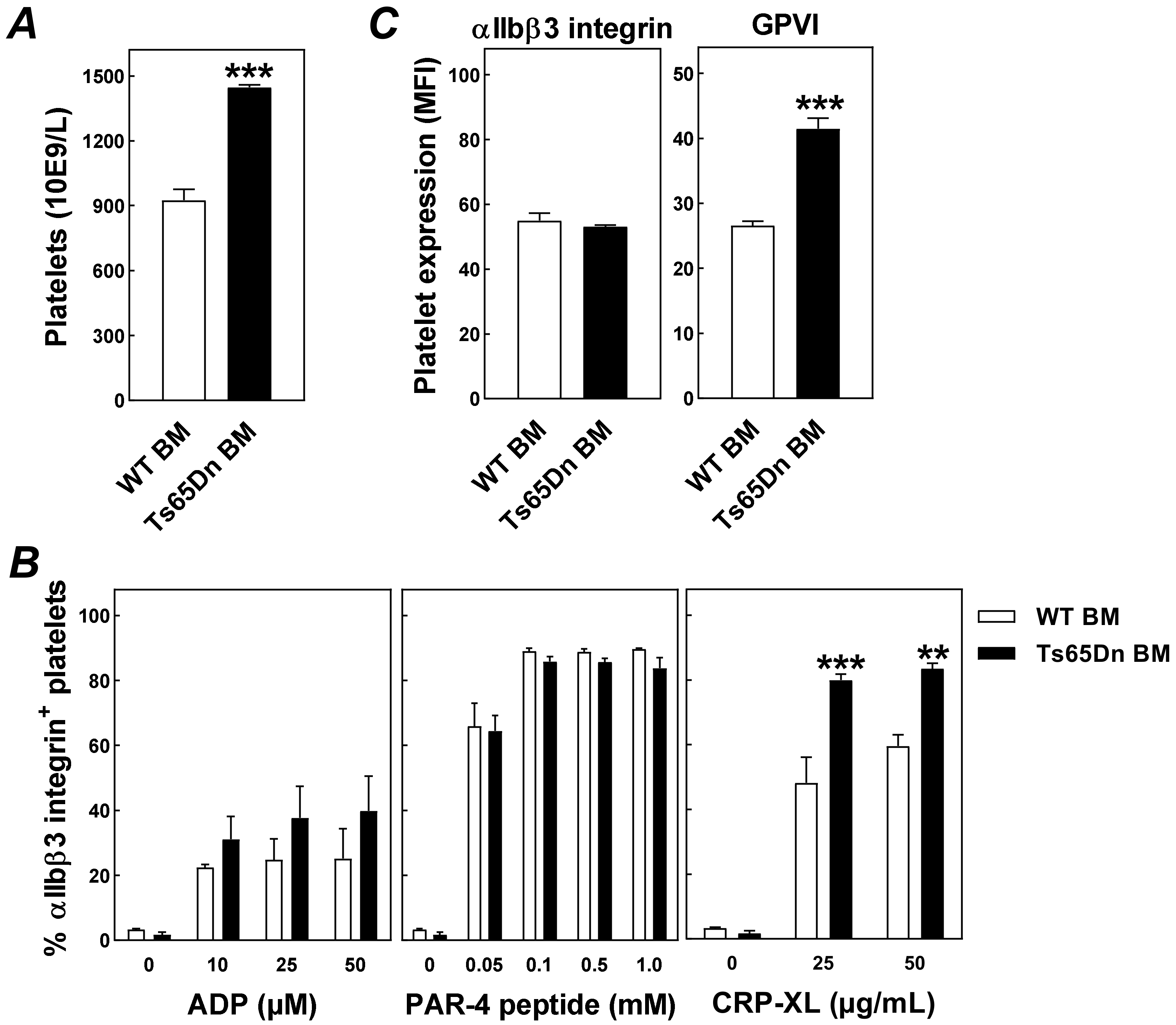
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murdoch, J.C.; Rodger, J.C.; Rao, S.S.; Fletcher, C.D.; Dunnigan, M.G. Down’s syndrome: An atheroma-free model? Br. Med. J. 1977, 2, 226–228. [Google Scholar] [CrossRef] [Green Version]
- Ylä-Herttuala, S.; Luoma, J.; Nikkari, T.; Kivimäki, T. Down’s syndrome and atherosclerosis. Atherosclerosis 1989, 76, 269–272. [Google Scholar] [CrossRef]
- Draheim, C.C.; Geijer, J.R.; Dengel, D.R. Comparison of intima-media thickness of the carotid artery and cardiovascular disease risk factors in adults with versus without the Down syndrome. Am. J. Cardiol. 2010, 106, 1512–1516. [Google Scholar] [CrossRef]
- Tansley, G.; Holmes, D.T.; Lütjohann, D.; Head, E.; Wellington, C.L. Sterol lipid metabolism in down syndrome revisited: Down syndrome is associated with a selective reduction in serum brassicasterol levels. Curr. Gerontol. Geriatr. Res. 2012, 2012, 179318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ram, G.; Chinen, J. Infections and immunodeficiency in Down syndrome. Clin. Exp. Immunol. 2011, 164, 9–16. [Google Scholar] [CrossRef]
- Satgé, D.; Seidel, M.G. The Pattern of Malignancies in Down syndrome and Its Potential Context With the Immune System. Front. Immunol. 2018, 9, 3058. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef]
- Davisson, M.T.; Schmidt, C.; Reeves, R.H.; Irving, N.G.; Akeson, E.C.; Harris, B.S.; Bronson, R.T. Segmental trisomy as a mouse model for Down syndrome. Prog. Clin. Biol. Res. 1993, 384, 117–133. [Google Scholar]
- Reeves, R.H.; Irving, N.G.; Moran, T.H.; Wohn, A.; Kitt, C.; Sisodia, S.S.; Schmidt, C.; Bronson, R.T.; Davisson, M.T. A mouse model for Down syndrome exhibits learning and behaviour deficits. Nat. Genet. 1995, 11, 177–184. [Google Scholar] [CrossRef]
- Holtzman, D.M.; Santucci, D.; Kilbridge, J.; Chua-Couzens, J.; Fontana, D.J.; Daniels, S.E.; Johnson, R.M.; Chen, K.; Sun, Y.; Carlson, E.; et al. Developmental abnormalities and age-related neurodegeneration in a mouse model of Down syndrome. Proc. Natl. Acad. Sci. USA 1996, 93, 13333–13338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahon, J.E.; Hoekstra, M.; van Hulst, S.; Manta, C.; Goerdt, S.; Geerling, J.J.; Géraud, C.; Van Eck, M. Hematopoietic Stabilin-1 deficiency does not influence atherosclerosis susceptibility in LDL receptor knockout mice. Atherosclerosis 2019, 281, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, M.; Kruijt, J.K.; Van Eck, M.; Van Berkel, T.J. Specific gene expression of ATP-binding cassette transporters and nuclear hormone receptors in rat liver parenchymal, endothelial, and Kupffer cells. J. Biol. Chem. 2003, 278, 25448–25453. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Vergara, M.; Shiri-Sverdlov, R.; de Haan, G.; Hofker, M.H. Bone marrow transplantation in mice as a tool for studying the role of hematopoietic cells in metabolic and cardiovascular diseases. Atherosclerosis 2010, 213, 335–344. [Google Scholar] [CrossRef]
- Lorenzo, L.P.; Chen, H.; Shatynski, K.E.; Clark, S.; Yuan, R.; Harrison, D.E.; Yarowsky, P.J.; Williams, M.S. Defective hematopoietic stem cell and lymphoid progenitor development in the Ts65Dn mouse model of Down syndrome: Potential role of oxidative stress. Antioxid Redox Signal. 2011, 15, 2083–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, P.; Rusconi, S.; Mavilio, D.; Fogli, M.; Murdaca, G.; Pende, D.; Mingari, M.C.; Galli, M.; Moretta, L.; De Maria, A. Differential disappearance of inhibitory natural killer cell receptors during HAART and possible impairment of HIV-1-specific CD8 cytotoxic T lymphocytes. AIDS 2001, 15, 965–974. [Google Scholar] [CrossRef]
- Ciprandi, G.; Murdaca, G.; Colombo, B.M.; De Amici, M.; Marseglia, G.L. Serum vascular endothelial growth factor in allergic rhinitis and systemic lupus erythematosus. Hum. Immunol. 2008, 69, 510–512. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, L.P.; Shatynski, K.E.; Clark, S.; Yarowsky, P.J.; Williams, M.S. Defective thymic progenitor development and mature T-cell responses in a mouse model for Down syndrome. Immunology 2013, 139, 447–458. [Google Scholar] [CrossRef]
- Wang, L.; Peters, J.M.; Fuda, F.; Li, L.; Karandikar, N.J.; Koduru, P.; Wang, H.Y.; Chen, W. Acute megakaryoblastic leukemia associated with trisomy 21 demonstrates a distinct immunophenotype. Cytometry B Clin. Cytom. 2015, 88, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, P.I.; Lappalainen, J.P.; Heinonen, S.E.; Leppänen, P.; Lähteenvuo, M.T.; Aarnio, J.V.; Heikkilä, J.; Turunen, M.P.; Ylä-Herttuala, S. Silencing of either SR-A or CD36 reduces atherosclerosis in hyperlipidaemic mice and reveals reciprocal upregulation of these receptors. Cardiovasc. Res. 2010, 88, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Febbraio, M.; Guy, E.; Silverstein, R.L. Stem cell transplantation reveals that absence of macrophage CD36 is protective against atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2333–2338. [Google Scholar] [CrossRef] [Green Version]
- Pogribna, M.; Melnyk, S.; Pogribny, I.; Chango, A.; Yi, P.; James, S.J. Homocysteine metabolism in children with Down syndrome: In vitro modulation. Am. J. Hum. Genet. 2001, 69, 88–95. [Google Scholar] [CrossRef] [Green Version]
- De la Torre, R.; De Sola, S.; Pons, M.; Duchon, A.; de Lagran, M.M.; Farré, M.; Fitó, M.; Benejam, B.; Langohr, K.; Rodriguez, J.; et al. Epigallocatechin-3-gallate, a DYRK1A inhibitor, rescues cognitive deficits in Down syndrome mouse models and in humans. Mol. Nutr. Food Res. 2014, 58, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Helm, S.; Blayney, M.; Whited, T.; Noroozi, M.; Lin, S.; Kern, S.; Green, D.; Salehi, A. Deleterious Effects of Chronic Folate Deficiency in the Ts65Dn Mouse Model of Down syndrome. Front. Cell Neurosci. 2017, 11, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noll, C.; Planque, C.; Ripoll, C.; Guedj, F.; Diez, A.; Ducros, V.; Belin, N.; Duchon, A.; Paul, J.L.; Badel, A.; et al. DYRK1A, a novel determinant of the methionine-homocysteine cycle in different mouse models overexpressing this Down-syndrome-associated kinase. PLoS ONE 2009, 4, e7540. [Google Scholar] [CrossRef]
- Latour, A.; Salameh, S.; Carbonne, C.; Daubigney, F.; Paul, J.L.; Kergoat, M.; Autier, V.; Delabar, J.M.; De Geest, B.; Janel, N. Corrective effects of hepatotoxicity by hepatic Dyrk1a gene delivery in mice with intermediate hyperhomocysteinemia. Mol. Genet. Metab. Rep. 2015, 2, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirsammer, G.; Jilani, S.; Liu, H.; Davis, E.; Gurbuxani, S.; Le Beau, M.M.; Crispino, J.D. Highly penetrant myeloproliferative disease in the Ts65Dn mouse model of Down syndrome. Blood 2008, 111, 767–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheppard, J.R.; Schumacher, W.; White, J.G.; Jakobs, K.H.; Schultz, G. The alpha adrenergic response of Down’s syndrome platelets. J. Pharmacol. Exp. Ther. 1983, 225, 584–588. [Google Scholar]
- Journeycake, J.M.; Brumley, L.E. Down syndrome as an Independent Risk Factor for Thrombosis in Children. Blood 2006, 108, 1489. [Google Scholar] [CrossRef]
- Tarlaci, S.; Sagduyu, A. Cerebral venous thrombosis in Down’s syndrome. Clin. Neurol. Neurosurg. 2001, 103, 242–244. [Google Scholar] [CrossRef]
- Kurokami, T.; Takasawa, R.; Takeda, S.; Kurobe, M.; Takasawa, K.; Nishioka, M.; Shimohira, M. Venous thromboembolism in two adolescents with Down syndrome. Turk. J. Pediatr. 2018, 60, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korporaal, S.J.A.; van der Sluis, R.J.; Van Eck, M.; Hoekstra, M. Bone Marrow Ts65Dn Trisomy-Induced Changes in Platelet Functionality and Lymphocytopenia Do Not Impact Atherosclerosis Susceptibility in Mice. J. Cardiovasc. Dev. Dis. 2021, 8, 110. https://doi.org/10.3390/jcdd8090110
Korporaal SJA, van der Sluis RJ, Van Eck M, Hoekstra M. Bone Marrow Ts65Dn Trisomy-Induced Changes in Platelet Functionality and Lymphocytopenia Do Not Impact Atherosclerosis Susceptibility in Mice. Journal of Cardiovascular Development and Disease. 2021; 8(9):110. https://doi.org/10.3390/jcdd8090110
Chicago/Turabian StyleKorporaal, Suzanne J. A., Ronald J. van der Sluis, Miranda Van Eck, and Menno Hoekstra. 2021. "Bone Marrow Ts65Dn Trisomy-Induced Changes in Platelet Functionality and Lymphocytopenia Do Not Impact Atherosclerosis Susceptibility in Mice" Journal of Cardiovascular Development and Disease 8, no. 9: 110. https://doi.org/10.3390/jcdd8090110