
Slow-Release Sachets of Neoseiulus cucumeris Predatory Mites Reduce Intraguild Predation by Dalotia coriaria in Greenhouse Biological Control Systems

Abstract
:1. Introduction
2. Experimental Section
2.1. Experiment One—Materials and Methods
2.2. Experiment Two—Materials and Methods
3. Results
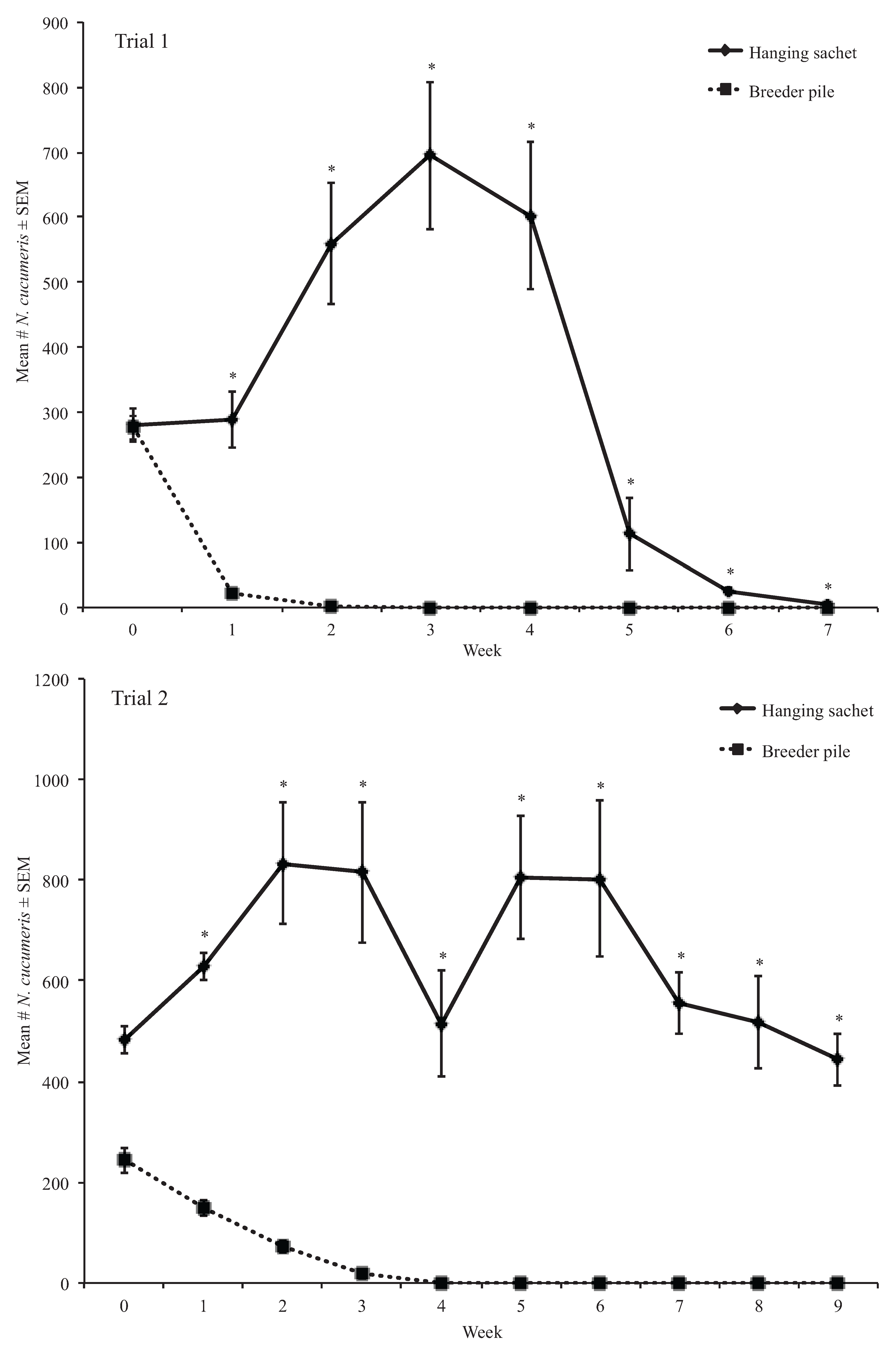
3.1. Experiment One Results
3.1.1. Dalotia coriaria Extracted from Samples

3.1.2. Weekly Proportion Change of Neoseiulus cucumeris

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week | Treatment | Percentage Change ± SEM | H-Value, df = 1 | p-Value |
|---|---|---|---|---|
| 1 | Breeder pile | −92.49 ± 01.22 | 6.8182 | 0.0090 * |
| Hanging sachet | 02.86 ± 15.21 | |||
| 2 | Breeder pile | −99.71 ± 00.18 | 7.0313 | 0.0080 * |
| Hanging sachet | 99.71 ± 33.53 | |||
| 3 | Breeder pile | −99.86 ± 00.14 | 7.2581 | 0.0071 * |
| Hanging sachet | 148.00 ± 40.29 | |||
| 4 | Breeder pile | −99.86 ± 00.09 | 7.0313 | 0.0080 * |
| Hanging sachet | 115.07 ± 40.55 | |||
| 5 | Breeder pile | −99.93 ± 00.07 | 7.2581 | 0.0071 * |
| Hanging sachet | −59.50 ± 20.04 | |||
| 6 | Breeder pile | −100.00 ± 00.00 | 7.7586 | 0.0053 * |
| Hanging sachet | −91.14 ± 01.65 | |||
| 7 | Breeder pile | −100.00 ± 00.00 | 5.5814 | 0.0182 * |
| Hanging sachet | −98.36 ± 00.88 |

| Week | Treatment | Percentage Change ± SEM | H-Value, df =1 | p-Value |
|---|---|---|---|---|
| 1 | Breeder pile | −38.90 ± 06.78 | 6.8182 | 0.0090 * |
| Hanging sachet | 30.53 ± 05.72 | |||
| 2 | Breeder pile | −70.23 ± 04.92 | 6.8182 | 0.0090 * |
| Hanging sachet | 72.87 ± 25.19 | |||
| 3 | Breeder pile | −92.52 ± 03.72 | 6.8598 | 0.0088 * |
| Hanging sachet | 69.02 ± 28.86 | |||
| 4 | Breeder pile | −99.84 ± 00.10 | 7.0313 | 0.0080 * |
| Hanging sachet | 06.89 ± 22.15 | |||
| 5 | Breeder pile | −99.92 ± 00.08 | 7.2581 | 0.0071 * |
| Hanging sachet | 66.98 ± 25.21 | |||
| 6 | Breeder pile | −100.00 ± 00.00 | 7.7586 | 0.0053 * |
| Hanging sachet | 66.49 ± 32.02 | |||
| 7 | Breeder pile | −99.92 ± 00.08 | 7.2581 | 0.0071 * |
| Hanging sachet | 15.47 ± 12.87 | |||
| 8 | Breeder pile | −99.92 ± 00.08 | 7.2581 | 0.0071 * |
| Hanging sachet | 07.55 ± 19.15 | |||
| 9 | Breeder pile | −100.00 ± 00.00 | 7.7586 | 0.0053 * |
| Hanging sachet | −08.13 ± 10.54 |
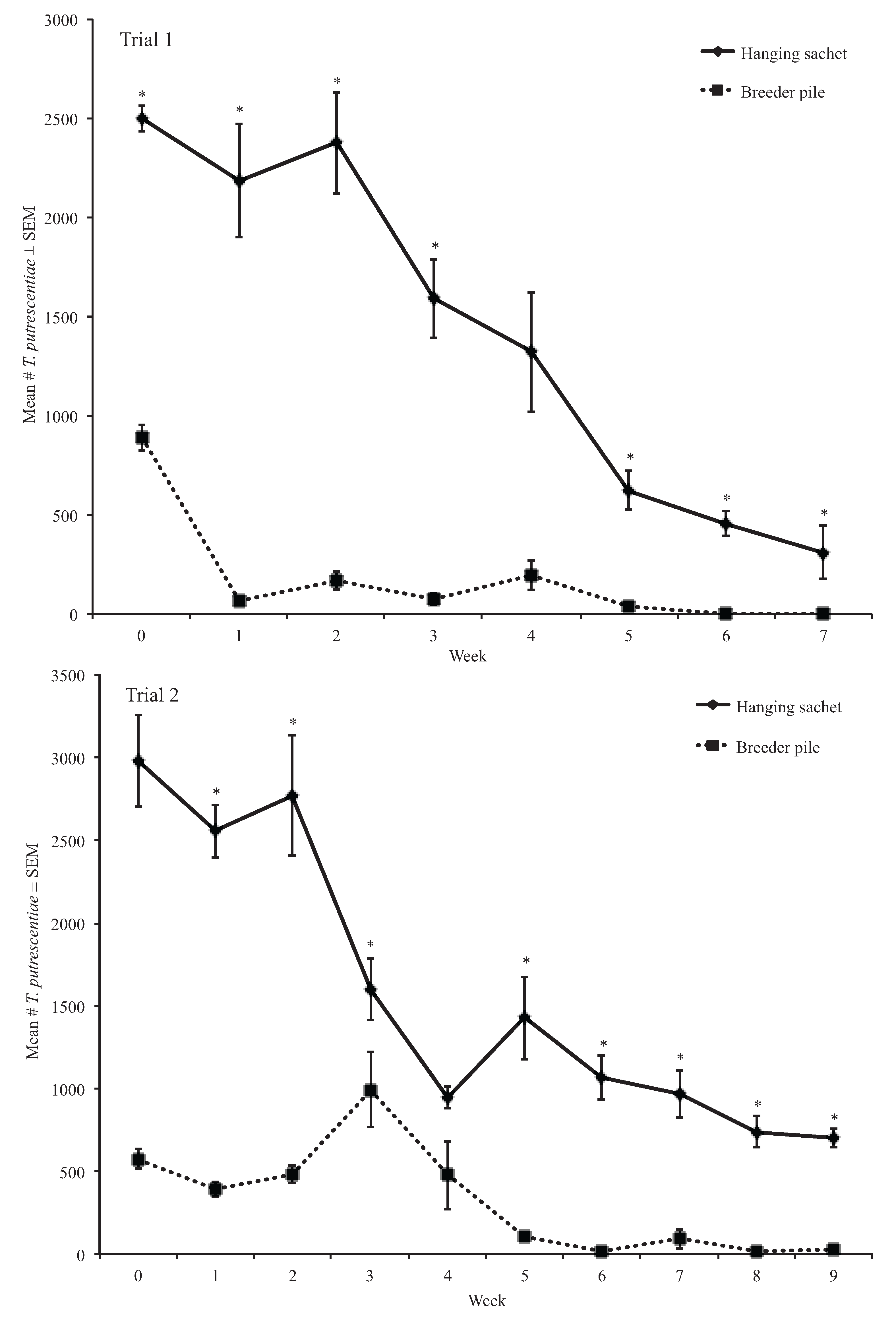
3.1.3. Weekly Proportion Change of Tyrophagus putrescentiae
| Week | Treatment | Percentage Change ± SEM | H-Value, df =1 | p-Value |
|---|---|---|---|---|
| 1 | Breeder pile | −92.97 ± 00.58 | 6.8182 | 0.0090 * |
| Hanging sachet | −12.41 ± 11.48 | |||
| 2 | Breeder pile | −81.10 ± 05.12 | 6.8182 | 0.0090 * |
| Hanging sachet | −04.89 ± 10.26 | |||
| 3 | Breeder pile | −91.61 ± 02.93 | 6.8182 | 0.0090 * |
| Hanging sachet | −36.24 ± 07.97 | |||
| 4 | Breeder pile | −78.02 ± 08.56 | 3.1527 | 0.0758 |
| Hanging sachet | −47.08 ± 12.13 | |||
| 5 | Breeder pile | −95.49 ± 01.81 | 6.8182 | 0.0090 * |
| Hanging sachet | −75.04 ± 03.97 | |||
| 6 | Breeder pile | −97.41 ± 01.90 | 6.8182 | 0.0090 * |
| Hanging sachet | −81.68 ± 02.58 | |||
| 7 | Breeder pile | −100.00 ± 00.00 | 7.7586 | 0.0053 * |
| Hanging sachet | −87.63 ± 05.44 |

| Week | Treatment | Percentage Change ± SEM | H-Value, df =1 | p-Value |
|---|---|---|---|---|
| 1 | Breeder pile | −32.38 ± 07.85 | 3.1527 | 0.0758 |
| Hanging sachet | −14.23 ± 05.39 | |||
| 2 | Breeder pile | −16.24 ± 09.11 | 0.2727 | 0.6015 |
| Hanging sachet | −06.99 ± 12.23 | |||
| 3 | Breeder pile | 72.31 ± 39.61 | 6.8182 | 0.0090 * |
| Hanging sachet | −46.25 ± 6.30 | |||
| 4 | Breeder pile | −17.15 ± 35.82 | 0.8836 | 0.3472 |
| Hanging sachet | −68.29 ± 02.11 | |||
| 5 | Breeder pile | −81.74 ± 04.17 | 6.8182 | 0.0090 * |
| Hanging sachet | −52.14 ± 08.39 | |||
| 6 | Breeder pile | −96.31 ± 01.59 | 6.8598 | 0.0088 * |
| Hanging sachet | −64.30 ± 04.44 | |||
| 7 | Breeder pile | −83.83 ± 10.36 | 2.4545 | 0.1172 |
| Hanging sachet | −67.65 ± 04.84 | |||
| 8 | Breeder pile | −97.88 ± 01.50 | 6.8182 | 0.0090 * |
| Hanging sachet | −75.28 ± 03.18 | |||
| 9 | Breeder pile | −95.44 ± 03.15 | 6.8182 | 0.0090 * |
| Hanging sachet | −76.48 ± 01.91 |
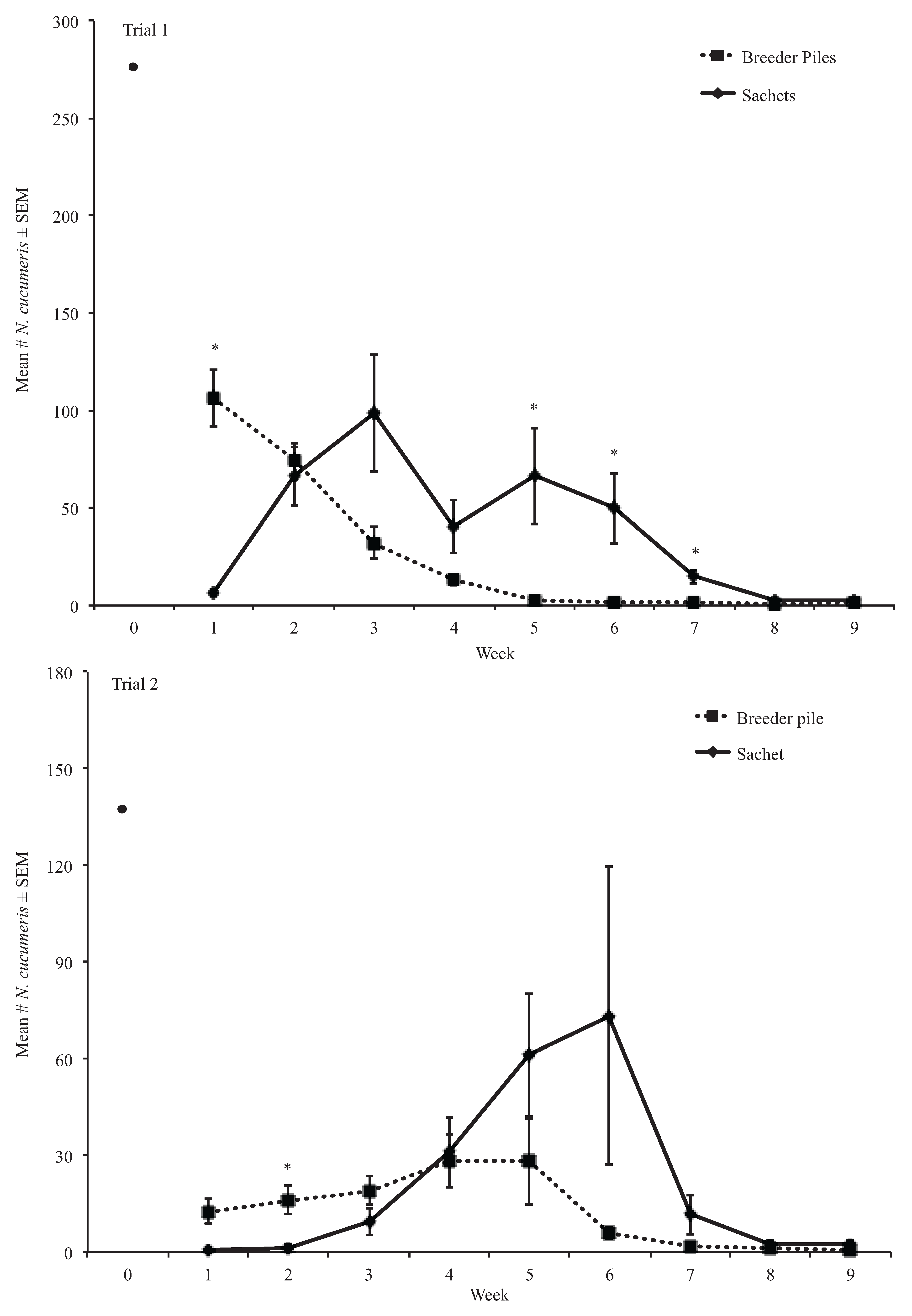
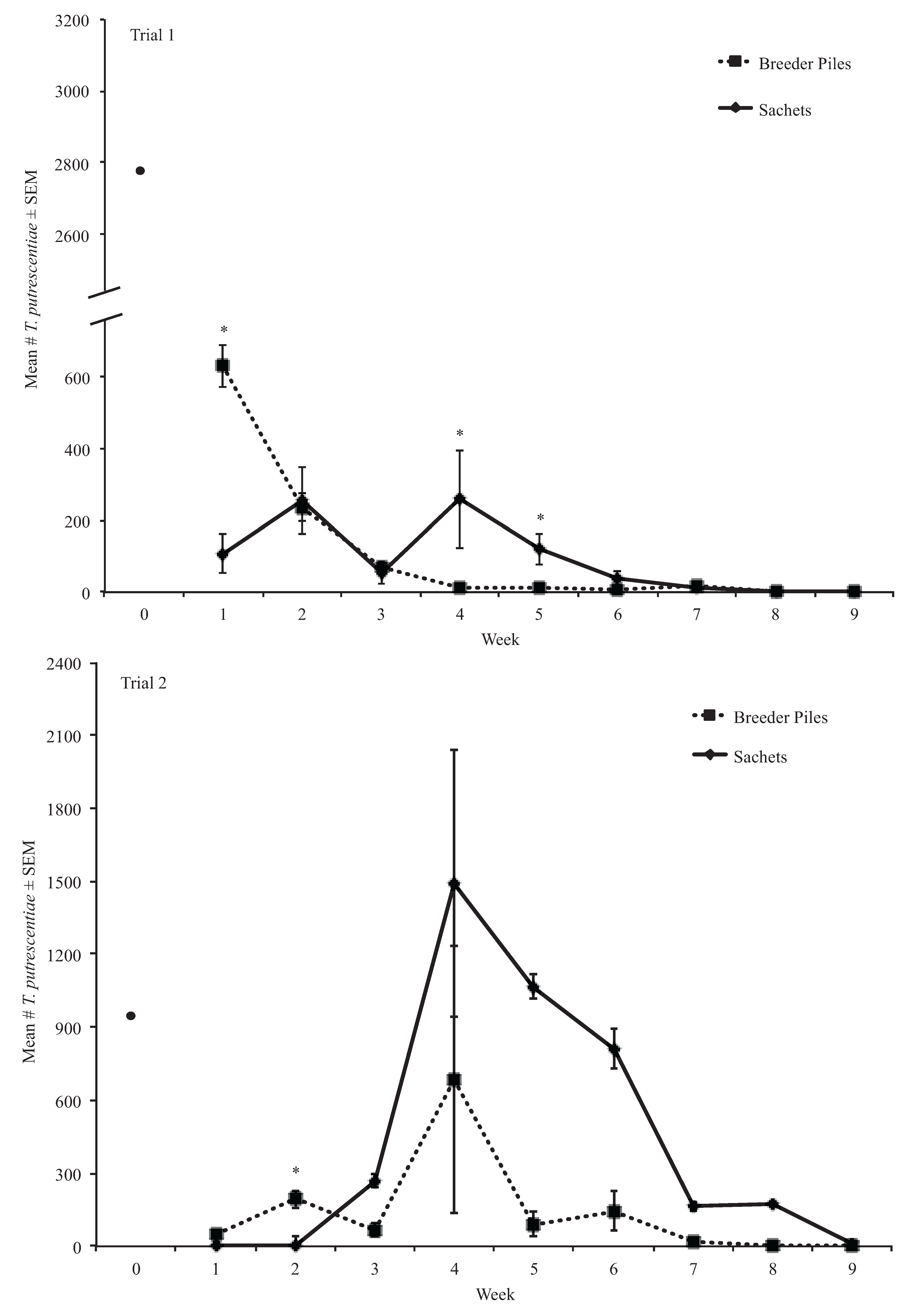
3.2. Experiment Two Results
3.2.1. Neoseiulus cucumeris Predatory Mites


3.2.2. Tyrophagus putrescentiae Mold Mites
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van Lenteren, J.C. A greenhouse without pesticides: Fact or fantasy? Crop Prot. 2000, 19, 375–384. [Google Scholar] [CrossRef]
- Stacey, D.L. Banker plant production of Encarsia-Formosa Gahan and its use in control of glasshouse whitefly on tomatoes. Plant Pathol. 1977, 26, 63–66. [Google Scholar] [CrossRef]
- Van Steenis, M.J. Biological control of the cotton aphid, Aphis gossypii Glover (Horn., Aphididae): Pre-Introduction evaluation of natural enemies. J. Appl. Entomol. 1992, 114, 362–380. [Google Scholar]
- Huang, N.X.; Enkegaard, A.; Osborne, L.S.; Ramakers, P.M.J.; Messelink, G.J.; Pijnakker, J.; Murphy, G. The Banker Plant Method in Biological Control. Crit. Rev. Plant Sci. 2011, 30, 259–278. [Google Scholar] [CrossRef]
- Janssen, A.; Pallini, A.; Venzon, M.; Sabelis, M.W. Review Behaviour and indirect interactions in food webs of plant-inhabiting arthropods. Exp. Appl. Acarol. 1998, 22, 497–521. [Google Scholar] [CrossRef]
- Janssen, A.; Sabelis, M.W.; Magalhães, S.; Montserrat, M.; van der Hammen, T. Habitat structure affects intraguild predation. Ecol. Soc. Am. 2007, 88, 2713–2719. [Google Scholar] [CrossRef]
- Messelink, G.J.; Sabelis, M.W.; Janssen, A. Generalist Predators, Food Web Complexities and Biological Pest Control in Greenhouse Crops. In Integrated Pest Management and Pest Control—Current and Future Tactics; Larramendy, M.L., Soloneski, S., Eds.; InTech: Rijeka, Croatia, 2012; pp. 191–214. [Google Scholar]
- Wittmann, E.J.; Leather, S.R. Compatibility of Orius laevigatus Fieber (Hemiptera: Anthocoridae) with Neoseiulus (Amblyseius) cucumeris Oudemans (Acari: Phytoseiidae) and Iphiseius (Amblyseius) degenerans Berlese (Acari: Phytoseiidae) in the biocontrol of Frankliniella occidentalis Pergande (Thysanoptera: Thripidae). Exp. Appl. Acarol. 1997, 21, 523–538. [Google Scholar]
- Jandricic, S.; Scott-Dupree, C.D.; Broadbent, A.B.; Harris, C.R.; Murphy, G. Compatibility of Atheta coriaria with other biological control agents and reduced-risk insecticides used in greenhouse floriculture integrated pest management programs for fungus gnats. Can. Entomol. 2006, 138, 712–722. [Google Scholar] [CrossRef]
- Buitenhuis, R.; Shipp, L.; Scott-Dupree, C. Intra-Guild vs. extra-guild prey: Effect on predator fitness and preference of Amblyseius swirskii (Athias-Henriot) and Neoseiulus cucumeris (Oudemans) (Acari: Phytoseiidae). Bull. Entomol. Res. 2009, 100, 167–173. [Google Scholar] [PubMed]
- Shipp, J.L.; Wang, K. Evaluation of Amblyseius cucumeris (Acari: Phytoseiidae) and Orius insidiosus (Hemiptera: Anthocoridae) for control of Frankliniella occidentalis (Thysanoptera: Thripidae) on greenhouse tomatoes. Biol. Control 2003, 28, 271–281. [Google Scholar] [CrossRef]
- Wright, E.M.; Chambers, R.J. The biology of the predatory mite Hypoaspis miles (Acari: Laelapidae), a potential biological control agent of Bradysia paupera (Dipt.: Sciaridae). BioControl 1994, 39, 225–235. [Google Scholar]
- Carney, V.A.; Diamond, J.C.; Murphy, G.D.; Marshall, D. The potential of Atheta coriaria Kraatz (Coleoptera: Staphylinidae), as a biological control agent for use in greenhouse crops. IOBC/WPRS Bull. 2002, 25, 37–40. [Google Scholar]
- Berndt, O.; Meyhöfer, R.; Poehling, H.M. The edaphic phase in the ontogenesis of Frankliniella occidentalis and comparison of Hypoaspis miles & H. aculeifer as predators of soil-dwelling thrips stages. Biol. Control 2004, 30, 17–24. [Google Scholar]
- Birken, E.M.; Cloyd, R.A. Food preference of the rove beetle, Atheta coriaria Kraatz (Coleoptera: Staphylinidae) under laboratory conditions. Insect Sci. 2007, 14, 53–56. [Google Scholar] [CrossRef]
- Pochubay, E.A.; Grieshop, M.J. Intraguild Predation of Neoseiulus cucumeris by Stratiolaelaps miles and Atheta coriaria in greenhouse open rearing systems. Biol. Control 2012, 63, 195–200. [Google Scholar] [CrossRef]
- Weintraub, P.G.; Kleitman, S.; Mori, R.; Shapira, N.; Palevsky, E. Control of the broad mite (Polyphagotarsonemus latus (Banks)) on organic greenhouse sweet peppers (Capsicum annuum L.) with the predatory mites, Neosiulus cucumeris (Oudemans). Biol. Control 2003, 27, 300–309. [Google Scholar]
- Van Houten, Y.M.; Østlie, M.L.; Hoogerbrugge, H.; Bolckmans, K. Biological control of western flower thrips on sweet pepper using the predatory mites Amblyseius cucumeris, Iphiseius degenerans, A. andersoni, and A. swirskii. IOBC/WPRS Bull. 2005, 28, 283–286. [Google Scholar]
- Krantz, G.W.; Walter, D.E. Collecting, Rearing, and Preparing Specimens. In A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 83–96. [Google Scholar]
- Van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Klowden, M.J. Metabolic Systems. In Physiological Systems in Insects, 2nd ed.; Elsevier Academic Press: Burlington, MA, USA, 2007; pp. 293–357. [Google Scholar]
- Fullaway, D.T. Orchid Insects. Proc. Hawaii. Entomol. Soc. 1937, 10, 45–49. [Google Scholar]
- Appel, A.G.; Rust, M.K.; Reierson, D.A. The smokybrown cockroach: Potential new pest in California. Calif. Agric. 1990, 44, 23–24. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pochubay, E.; Tourtois, J.; Himmelein, J.; Grieshop, M. Slow-Release Sachets of Neoseiulus cucumeris Predatory Mites Reduce Intraguild Predation by Dalotia coriaria in Greenhouse Biological Control Systems. Insects 2015, 6, 489-507. https://doi.org/10.3390/insects6020489
Pochubay E, Tourtois J, Himmelein J, Grieshop M. Slow-Release Sachets of Neoseiulus cucumeris Predatory Mites Reduce Intraguild Predation by Dalotia coriaria in Greenhouse Biological Control Systems. Insects. 2015; 6(2):489-507. https://doi.org/10.3390/insects6020489
Chicago/Turabian StylePochubay, Emily, Joseph Tourtois, Jeanne Himmelein, and Matthew Grieshop. 2015. "Slow-Release Sachets of Neoseiulus cucumeris Predatory Mites Reduce Intraguild Predation by Dalotia coriaria in Greenhouse Biological Control Systems" Insects 6, no. 2: 489-507. https://doi.org/10.3390/insects6020489