
Phylogenomic Analyses of the Tenthredinoidea Support the Familial Rank of Athaliidae (Insecta, Tenthredinoidea)

 ,
,  , , , ,
, , , ,  ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Athalia and Its Relatives
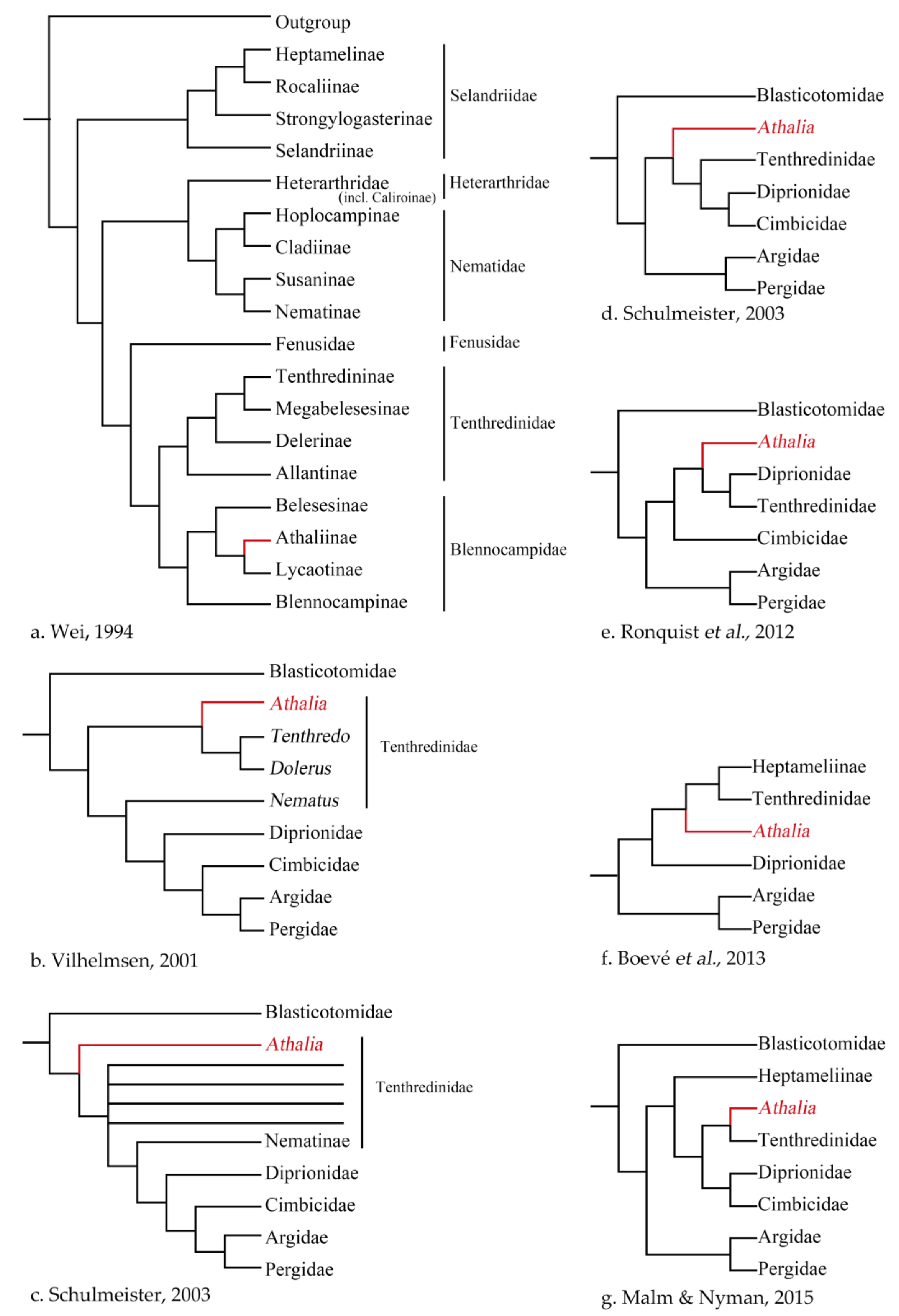
1.2. An Overview of the Systematic Position of Athalia and Its Relatives
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction and Sequencing
2.3. Genome Assembly and Single-Copy Assignment
2.4. Mitogenome Assembly, Annotation, and Structure Predictions
2.5. Phylogenomic Analyses
2.5.1. Alignment, Refinement, Supermatrix Construction, and Model Selection
2.5.2. Phylogenomic Inferences
2.5.3. Divergence Time Estimation
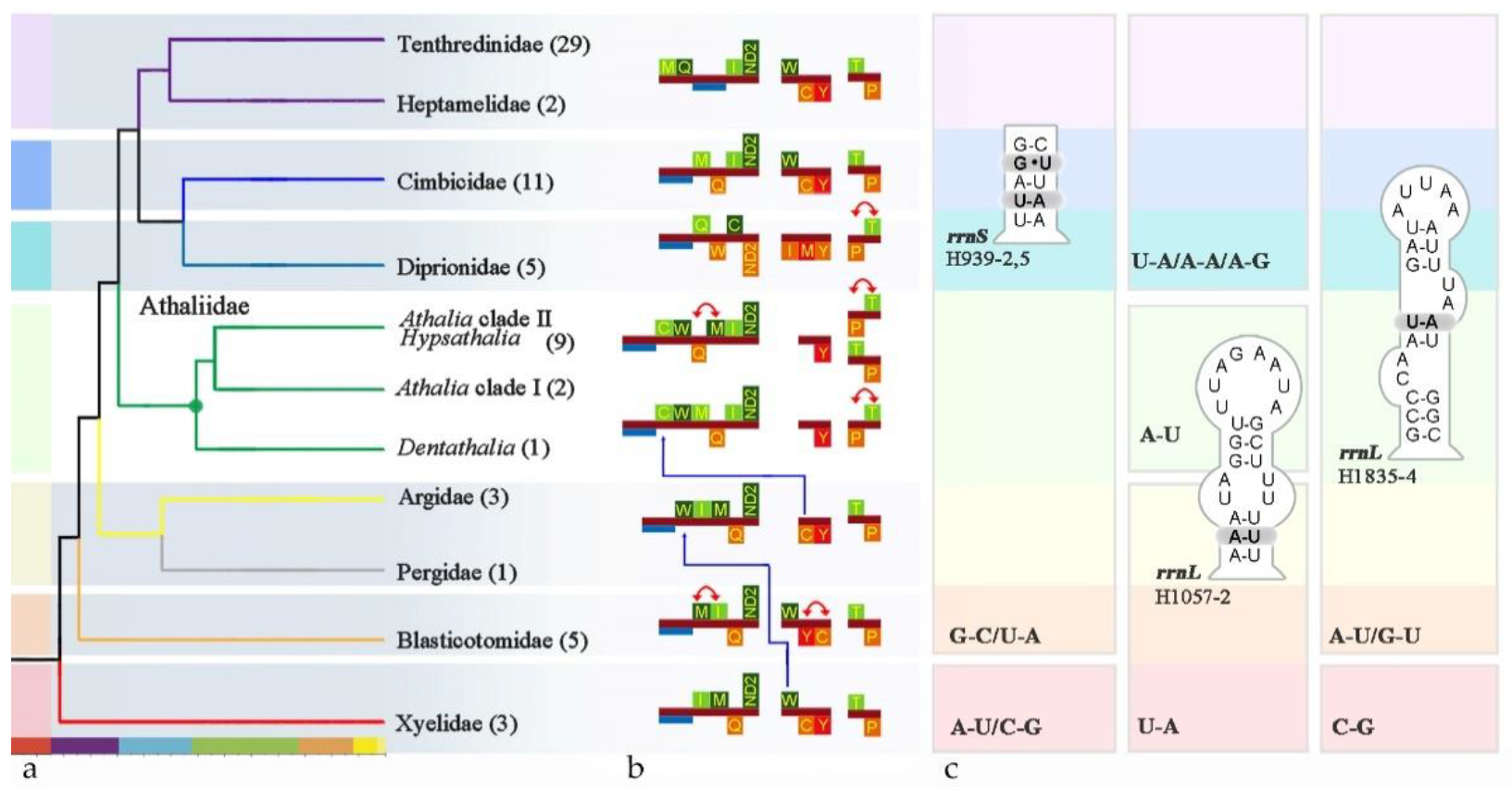
3. Results
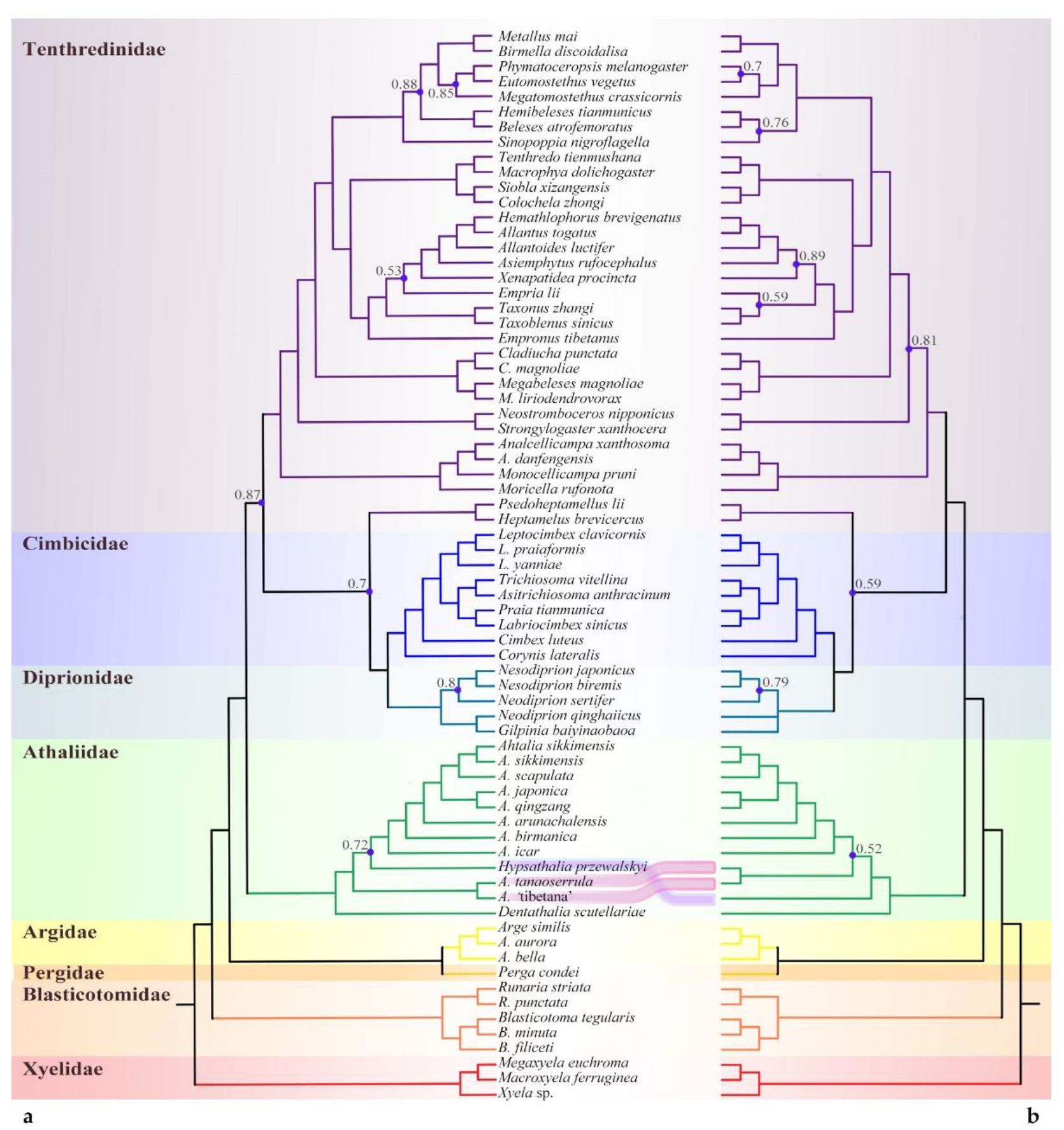
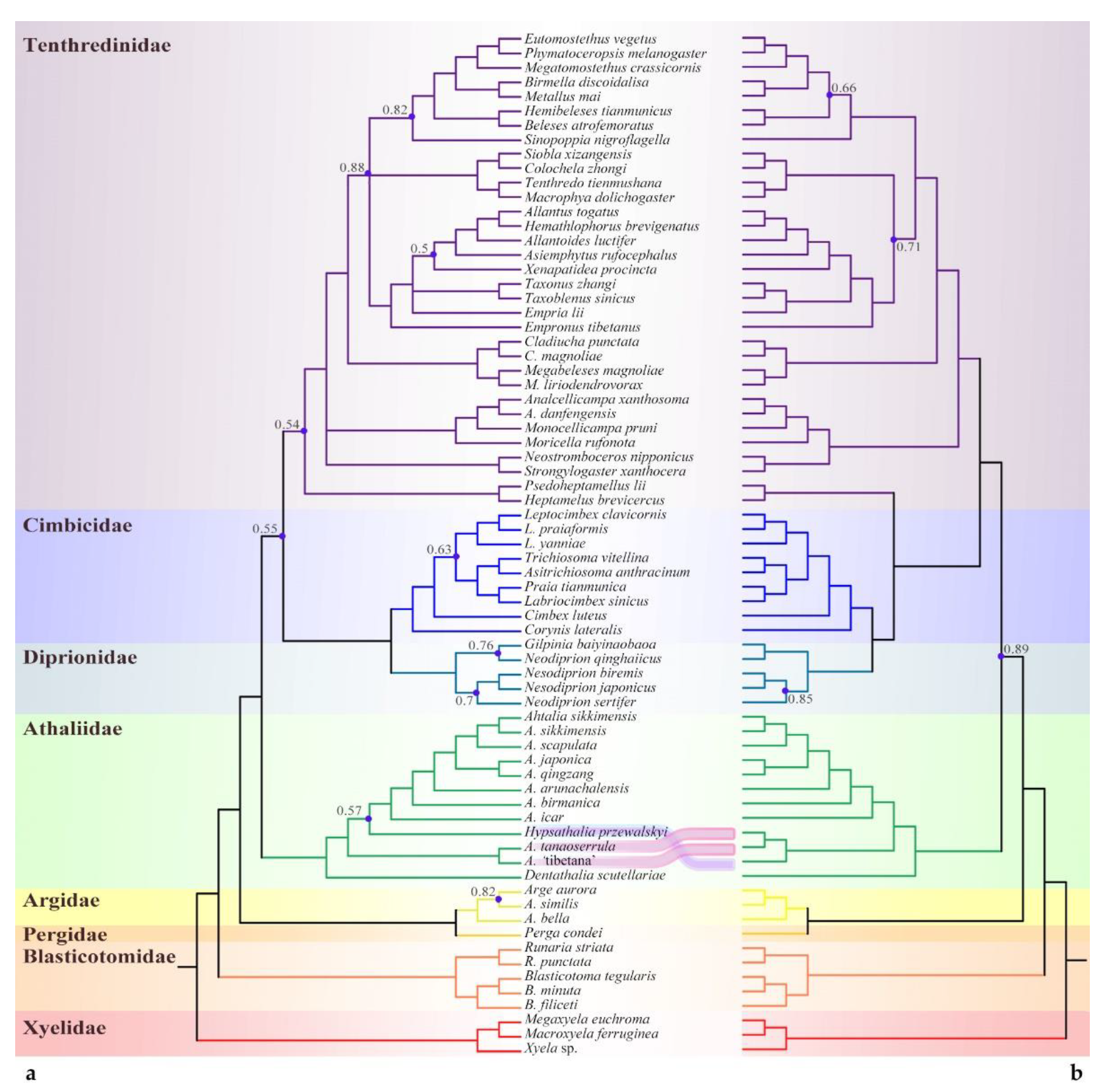
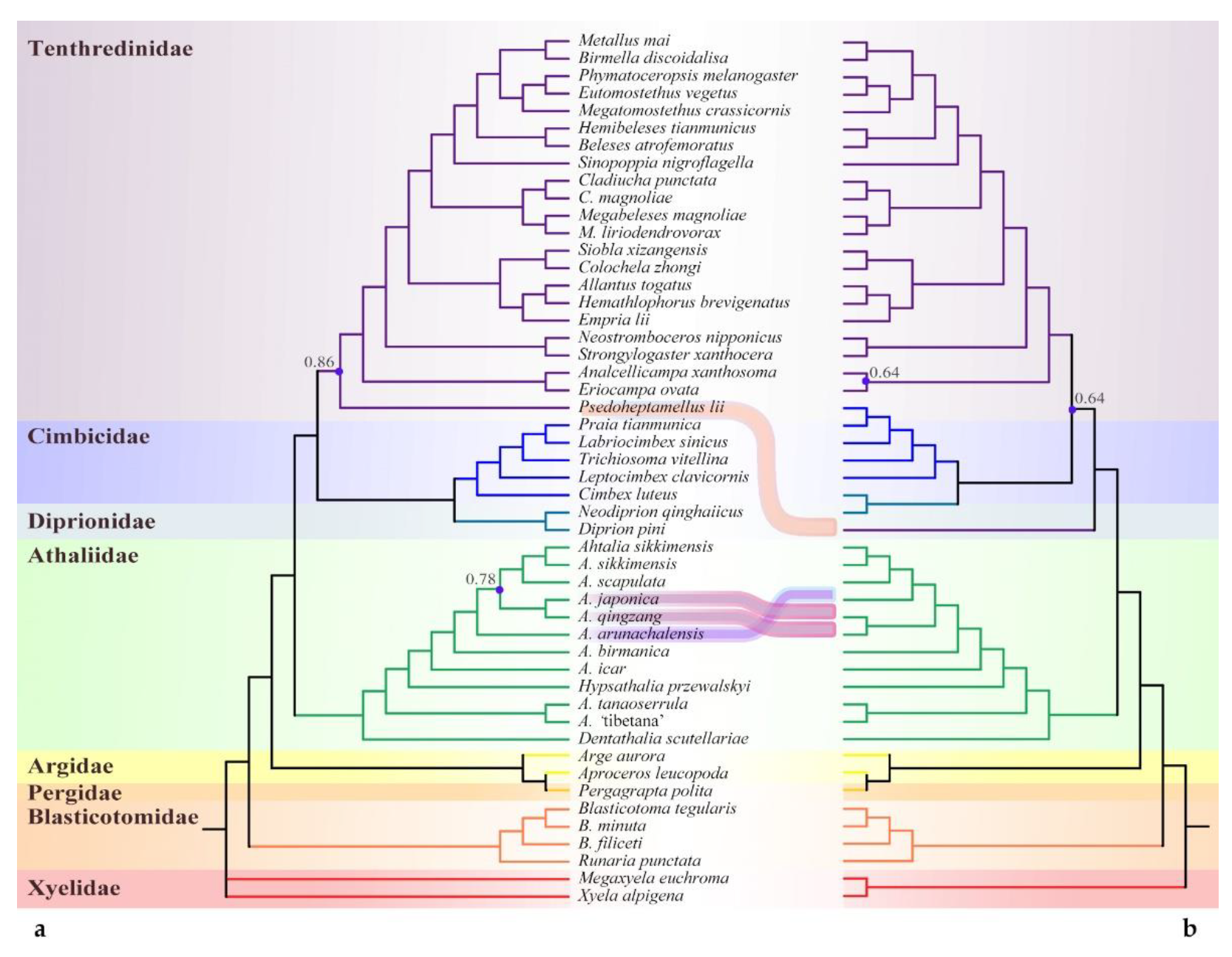
3.1. Phylogenomic Assessment of the Placement of Athalia and Its Relatives
3.1.1. Phylogenetic Analysis Based on Sequences
3.1.2. Rare Genomic Changes
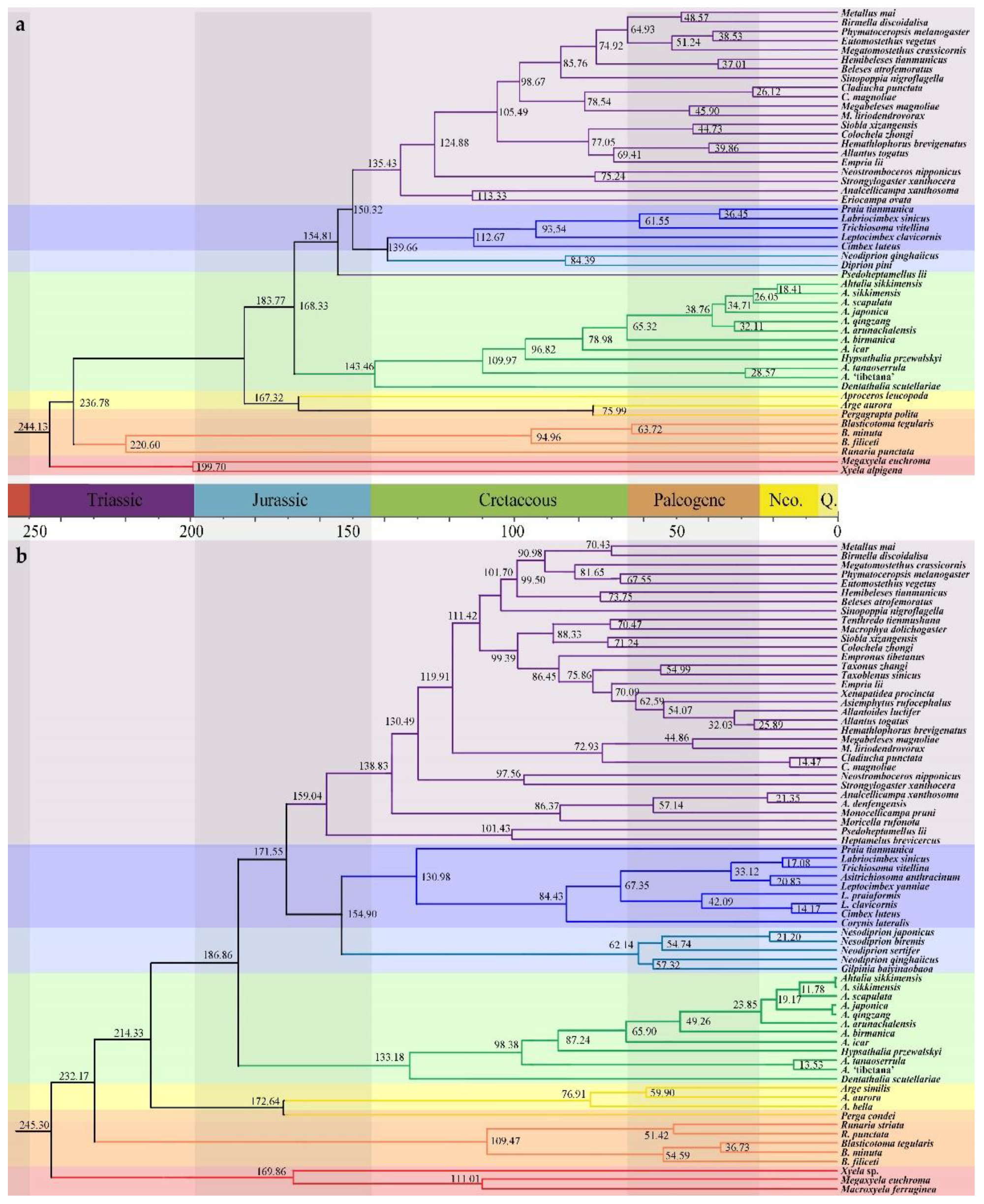
3.2. Divergence Time Estimates
4. Discussion
4.1. Sequence-Based Phylogeny, Rare Genomic Changes, and Morphology: Congruence in the Placement of Athaliidae
4.2. Spatial and Temporal Diversification of Athaliidae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics Resolves the Timing and Pattern of Insect Evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary History of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.P.; Dietrich, C.H.; Friedrich, F.; Beutel, R.G.; Wipfler, B.; Peters, R.S.; Allen, J.M.; Petersen, M.; Donath, A.; Walden, K.K.O.; et al. Phylogenomics and the Evolution of Hemipteroid Insects. Proc. Natl. Acad. Sci. USA 2018, 115, 12775–12780. [Google Scholar] [CrossRef] [PubMed]
- Wipfler, B.; Letsch, H.; Frandsen, P.B.; Kapli, P.; Mayer, C.; Bartel, D.; Buckley, T.R.; Donath, A.; Edgerly-Rooks, J.S.; Fujita, M.; et al. Evolutionary History of Polyneoptera and Its Implications for Our Understanding of Early Winged Insects. Proc. Natl. Acad. Sci. USA 2019, 116, 3024–3029. [Google Scholar] [CrossRef] [PubMed]
- Evangelista, D.A.; Wipfler, B.; Béthoux, O.; Donath, A.; Fujita, M.; Kohli, M.K.; Legendre, F.; Liu, S.; Machida, R.; Misof, B.; et al. An Integrative Phylogenomic Approach Illuminates the Evolutionary History of Cockroaches and Termites (Blattodea). Proc. R. Soc. B. 2019, 286, 20182076. [Google Scholar] [CrossRef]
- Kawahara, A.Y.; Plotkin, D.; Espeland, M.; Meusemann, K.; Toussaint, E.F.A.; Donath, A.; Gimnich, F.; Frandsen, P.B.; Zwick, A.; Dos Reis, M.; et al. Phylogenomics Reveals the Evolutionary Timing and Pattern of Butterflies and Moths. Proc. Natl. Acad. Sci. USA 2019, 116, 22657–22663. [Google Scholar] [CrossRef]
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrich, F.; Letsch, H.; et al. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef]
- Vasilikopoulos, A.; Misof, B.; Meusemann, K.; Lieberz, D.; Flouri, T.; Beutel, R.G.; Niehuis, O.; Wappler, T.; Rust, J.; Peters, R.S.; et al. An Integrative Phylogenomic Approach to Elucidate the Evolutionary History and Divergence Times of Neuropterida (Insecta: Holometabola). BMC Evol. Biol. 2020, 20, 64. [Google Scholar] [CrossRef]
- Tihelka, E.; Cai, C.; Giacomelli, M.; Lozano-Fernandez, J.; Rota-Stabelli, O.; Huang, D.; Engel, M.S.; Donoghue, P.C.J.; Pisani, D. The Evolution of Insect Biodiversity. Curr. Biol. 2021, 31, R1299–R1311. [Google Scholar] [CrossRef]
- Tihelka, E.; Engel, M.S.; Lozano-Fernandez, J.; Giacomelli, M.; Yin, Z.; Rota-Stabelli, O.; Huang, D.; Pisani, D.; Donoghue, P.C.J.; Cai, C. Compositional Phylogenomic Modelling Resolves the ‘Zoraptera Problem’: Zoraptera Are Sister to All Other Polyneopteran Insects. bioRxiv 2021. [Google Scholar]
- Whiting, M.F. Phylogeny of the Holometabolous Insect Orders: Molecular Evidence. Zool. Scr. 2002, 31, 3–15. [Google Scholar] [CrossRef]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.-W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Bayless, K.M.; Heimberg, A.M.; et al. Episodic Radiations in the Fly Tree of Life. Proc. Natl. Acad. Sci. USA 2011, 108, 5690–5695. [Google Scholar] [CrossRef] [PubMed]
- Kapli, P.; Flouri, T.; Telford, M.J. Systematic Errors in Phylogenetic Trees. Curr. Biol. 2021, 31, R59–R64. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.J. Key Innovations, Convergence, and Success: Macroevolutionary Lessons from Plant Phylogeny. Paleobiology 2005, 31, 77–93. [Google Scholar] [CrossRef]
- Rokas, A.; Holland, P.W.H. Rare Genomic Changes as a Tool for Phylogenetics. Trends Ecol. Evol. 2000, 15, 454–459. [Google Scholar] [CrossRef]
- Moret, B.M.E.; Tang, J.; Warnow, T.J. Reconstructing Phylogenies from Gene-Content and Gene-Order Data. In Proceedings of the Mathematics of Evolution and Phylogeny, Paris, France, 17–18 June 2007. [Google Scholar]
- Donath, A.; Stadler, P.F. 25 Molecular Morphology: Higher Order Characters Derivable from Sequence Information. In Deep Metazoan Phylogeny: The Backbone of the Tree of Life; De Gruyter: Berlin, Germany, 2014; pp. 549–562. ISBN 978-3-11-027752-4. [Google Scholar]
- Watterson, G.A.; Ewens, W.J.; Hall, T.E.; Morgan, A. The Chromosome Inversion Problem. J. Theor. Biol. 1982, 99, 1–7. [Google Scholar] [CrossRef]
- Bernt, M.; Merkle, D.; Middendorf, M.; Schierwater, B.; Schlegel, M.; Stadler, P.F. 22 Computational Methods for the Analysis of Mitochondrial Genome Rearrangements. In Deep Metazoan Phylogeny: The Backbone of the Tree of Life; Wägele, J.W., Bartolomaeus, T., Eds.; De Gruyter: Berlin, Germany, 2014; pp. 515–530. ISBN 978-3-11-027746-3. [Google Scholar]
- Hartmann, T.; Bernt, M.; Middendorf, M. EqualTDRL: Illustrating Equivalent Tandem Duplication Random Loss Rearrangements. BMC Bioinform. 2018, 19, 192. [Google Scholar] [CrossRef]
- Lavrov, D.V.; Boore, J.; Brown, W. Complete MtDNA Sequences of Two Millipedes Suggest a New Model for Mitochondrial Gene Rearrangements: Duplication and Non-Random Loss. Mol. Biol. Evol. 2002, 19, 163–169. [Google Scholar] [CrossRef]
- Podsiadlowski, L.; Mwinyi, A.; Lesný, P.; Bartolomaeus, T. 19 Mitochondrial Gene Order in Metazoa—Theme and Variations. In Deep Metazoan Phylogeny: The Backbone of the Tree of Life; Wägele, J.W., Bartolomaeus, T., Eds.; De Gruyter: Berlin, Germany, 2014; pp. 459–472. ISBN 978-3-11-027746-3. [Google Scholar]
- Moreno-Carmona, M.; Cameron, S.L.; Prada Quiroga, C.F. How Are the Mitochondrial Genomes Reorganized in Hexapoda? Differential Evolution and the First Report of Convergences within Hexapoda. Gene 2021, 791, 145719. [Google Scholar] [CrossRef]
- Dowton, M.; Cameron, S.L.; Dowavic, J.I.; Austin, A.D.; Whiting, M.F. Characterization of 67 Mitochondrial TRNA Gene Rearrangements in the Hymenoptera Suggests That Mitochondrial TRNA Gene Position Is Selectively Neutral. Mol. Biol. Evol. 2009, 26, 1607–1617. [Google Scholar] [CrossRef]
- Niu, G.; Jiang, S.; Doğan, Ö.; Korkmaz, E.M.; Budak, M.; Wu, D.; Wei, M. Mitochondrial Phylogenomics of Tenthredinidae (Hymenoptera: Tenthredinoidea) Supports the Monophyly of Megabelesesinae as a Subfamily. Insects 2021, 12, 495. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Cheng, Y.; Cai, C.; Meng, G.; Yan, Y.; Sun, Z.; Wan, S.; Wei, M. Phylogenomics Illuminates the Divergence Timeline and the Dynamic Mitogenome of Cimbicidae. Insects 2022, 13, 858. [Google Scholar] [CrossRef]
- Donath, A.; Findeiβ, S.; Hertel, J.; Marz, M.; Otto, W.; Schulz, C.; Stadler, P.F.; Wirth, S. Noncoding RNA. In Evolutionary Genomics and Systems Biology; Caetano-Anollés, G., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 251–293. ISBN 978-0-470-57041-8. [Google Scholar]
- Wei, M. Studies on the Phylogeny of Tenthredinoidea (s.Str.) With the Taxonomy of Chinese Tenthredinoidea Excl. Nematidae & Tenthredininae. Ph.D. Thesis, Institute of Zoology, Academia Sinica, Beijing, China, 1994. [Google Scholar]
- Vilhelmsen, L. Phylogeny and Classification of the Extant Basal Lineages of the Hymenoptera (Insecta). Zool. J. Linn. Soc. 2001, 131, 393–442. [Google Scholar] [CrossRef]
- Schulmeister, S. Review of Morphological Evidence on the Phylogeny of Basal Hymenoptera (Insecta), with a Discussion of the Ordering of Characters. Biol. J. Linn. Soc. 2003, 79, 209–243. [Google Scholar] [CrossRef]
- Schulmeister, S. Simultaneous Analysis of Basal Hymenoptera (Insecta): Introducing Robust-Choice Sensitivity Analysis. Biol. J. Linn. Soc. 2003, 79, 245–275. [Google Scholar] [CrossRef]
- Ronquist, F.; Klopfstein, S.; Vilhelmsen, L.; Schulmeister, S.; Murray, D.L.; Rasnitsyn, A.P. A Total-Evidence Approach to Dating with Fossils, Applied to the Early Radiation of the Hymenoptera. Syst. Biol. 2012, 61, 973–999. [Google Scholar] [CrossRef] [PubMed]
- Malm, T.; Nyman, T. Phylogeny of the Symphytan Grade of Hymenoptera: New Pieces into the Old Jigsaw (Fly) Puzzle. Cladistics 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Boevé, J.-L.; Blank, S.M.; Meijer, G.; Nyman, T. Invertebrate and Avian Predators as Drivers of Chemical Defensive Strategies in Tenthredinid Sawflies. BMC Evol. Biol. 2013, 13, 198. [Google Scholar] [CrossRef]
- Leach, W.E. The Zoological Miscellany: Being Descriptions of New, or Interesting Animals; R. and A. Taylor, Shoe-Lane: London, UK, 1817; Volume 3, p. 155. [Google Scholar]
- Cameron, P. Hymenopterological Notices. Mem. Proc. Manch. Lit. Philos. Soc. 1891, 4, 182–194. [Google Scholar]
- Taeger, A.; Blank, S.M.; Liston, A. European Sawflies (Hymenoptera: Symphyta)—A Species Checklist for the Countries. In Recent Sawfly Research—Synthesis and Prospects; Blank, S.M., Schmidt, S., Taeger, A., Eds.; Goecke & Evers: Keltern, Germany, 2006; pp. 399–504. ISBN 978-3-937783-19-2. [Google Scholar]
- Benson, R.B. Notes on the British Sawflies of the Genus Athalia (Hymenoptera, Tenthredinidae), with the Description of a New Species. Entomol. Mon. Mag. 1931, 67, 109–114. [Google Scholar]
- Benson, R.B. A Revision of the Athaliini (Hymenoptera: Symphyta). Bull. Br. Mus. (Nat. Hist.). Entomol. Ser. 1962, 11, 333–382. [Google Scholar]
- Forsius, R. Über Eine Neue Selandriinen-Gattung Mit Vielgliedrigen Fühlern. Not. Entomol. 1935, 15, 57–59. [Google Scholar]
- Togashi, I. Symphyta of Thailand (Insecta, Hymenoptera). Steenstrupia 1988, 14, 101–119. [Google Scholar]
- Saini, M.S.; Blank, S.M.; Smith, D.R. Checklist of the Sawflies (Hymenoptera: Symphyta) of India. In Recent Sawfly Research—Synthesis and Prospects; Blank, S.M., Schmidt, S., Taeger, A., Eds.; Goecke & Evers: Keltern, Germany, 2006; pp. 575–617. ISBN 978-3-937783-19-2. [Google Scholar]
- Hartig, T. Die Aderflügler Deutschlands Mit Besonderer Berücksichtigung Ihres Larvenzustandes Und Ihres Wirkens in Wäldern Und Gärten Für Entomologen, Wald-Und Gartenbesitzer; Haude und Spener: Berlin, Germany, 1837; p. 416. [Google Scholar]
- Kirby, W.F. List of Hymenoptera with Descriptions and Figures of the Typical Specimens in the British Museum. 1. Tenthredinidae and Siricidae Vol. 1; British Museum (Natural History): London, UK, 1882; p. 450. [Google Scholar]
- Cameron, P. A Monograph of the British Phytophagous Hymenoptera. (Tenthredo, Sirex and Cynips, Linné.); Ray Society: London, UK, 1882; p. 340. [Google Scholar]
- Ashmead, W.H. Classification of the Horntails and Sawflies, or the Sub-Order Phytophaga. Can. Entomol. 1898, 30, 249–257. [Google Scholar] [CrossRef]
- Konow, F.W. Hymenoptera. Fam. Tenthredinidae. Genera Insectorum 1905, 29, 1–176. [Google Scholar]
- Rohwer, S.A. Technical Papers on Miscellaneous Forest Insects. II. The Genotypes of Sawflies and Woodwasps, or the Superfamily Tenthredinoidea. Tech. Ser. U.S. Dep. Agric. Bur. Entomol. 1911, 20, 69–109. [Google Scholar]
- Benson, R.B. On the Classification of Sawflies (Hymenoptera Symphyta). Trans. R. Entomol. Soc. Lond. 1938, 87, 353–384. [Google Scholar] [CrossRef]
- Takeuchi, K. A Generic Classification of the Japanese Tenthredinidae (Hymenoptera: Symphyta); Shinbi Print. Co.: Kyoto, Japan, 1952; Volume 91. [Google Scholar]
- Benson, R.B. Hymenoptera 2. Handb. Identif. Br. Insects 1952, 6, 51–137. [Google Scholar]
- Lorenz, H.; Kraus, M. Die Larvalsystematik Der Blattwespen. (Tenthredinoidea Und Megalodontoidea); Akademie Verlag: Berlin, Germany, 1957; pp. 1–389. [Google Scholar]
- Zhelochovtsev, A.N. Otrjad Hymenoptera—Pereponchatokrylye Podotrjad Symphyta (Chalastogastra)—Sidjachebrjuhie. [Order Hymenoptera—Wasps Suborder Symphyta (Chalastogastra)—Sawflies and woodwasps.]; Opredelitel’ nasekomyh evropejskoj chasti SSSR. T. III; Nauka: Leningrad, Russia, 1988; Volume 27, ISBN 978-5-02-025709-2. [Google Scholar]
- Abe, M.; Smith, D.R. The Genus-Group Names of Symphyta (Hymenoptera) and Their Type Species. Esakia 1991, 31, 115. [Google Scholar] [CrossRef]
- Wei, M.; Nie, H. Generic List of Tenthredinoidea s. Str. in New Systematic Arrangement with Synonyms and Distribution Data. J. Cent.-South For. Coll. 1998, 18, 23–31. [Google Scholar]
- Lacourt, J. Répertoire Des Tenthredinidae Ouest-Paléarctiques (Hymenoptera, Symphyta). Mémoires De La Société Entomol. De Fr. 1999, 3, 1–432. [Google Scholar]
- Heraty, J.; Ronquist, F.; Carpenter, J.M.; Hawks, D.; Schulmeister, S.; Dowling, A.P.; Murray, D.; Munro, J.; Wheeler, W.C.; Schiff, N.; et al. Evolution of the Hymenopteran Megaradiation. Mol. Phylogenetics Evol. 2011, 60, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Isaka, Y.; Sato, T. Was Species Diversification in Tenthredinoidea (Hymenoptera: Symphyta) Related to the Origin and Diversification of Angiosperms? Can. Entomol. 2015, 147, 443–458. [Google Scholar] [CrossRef]
- Vilhelmsen, L.; Mikó, I.; Krogmann, L. Beyond the Wasp-Waist: Structural Diversity and Phylogenetic Significance of the Mesosoma in Apocritan Wasps (Insecta: Hymenoptera): Phylogeny of Hymenoptera. Zool. J. Linn. Soc. 2010, 159, 22–194. [Google Scholar] [CrossRef]
- Vilhelmsen, L. Morphological Phylogenetics of the Tenthredinidae (Insecta: Hymenoptera). Invert. Syst. 2015, 29, 164–190. [Google Scholar] [CrossRef]
- Taeger, A.; Blank, S.M.; Liston, A.D. World Catalog of Symphyta (Hymenoptera). Zootaxa 2010, 2580, 1. [Google Scholar] [CrossRef]
- Castro, L.R.; Dowton, M. The Position of the Hymenoptera within the Holometabola as Inferred from the Mitochondrial Genome of Perga Condei (Hymenoptera: Symphyta: Pergidae). Mol. Phylogenetics Evol. 2005, 34, 469–479. [Google Scholar] [CrossRef]
- Du, S.; Niu, G.; Nyman, T.; Wei, M. Characterization of the Mitochondrial Genome of Arge Bella Wei & Du Sp. Nov. (Hymenoptera: Argidae). PeerJ 2018, 6, e6131. [Google Scholar] [CrossRef]
- Tang, P.; Zhu, J.; Zheng, B.; Wei, S.; Sharkey, M.; Chen, X.; Vogler, A.P. Mitochondrial Phylogenomics of the Hymenoptera. Mol. Phylogenetics Evol. 2019, 131, 8–18. [Google Scholar] [CrossRef]
- He, H.; Niu, G.; Zhang, B.; Wei, M. The Complete Mitochondrial Genome of Athalia Proxima (Hymenoptera: Tenthredinidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2019, 4, 3868–3869. [Google Scholar] [CrossRef]
- Yan, Y.; Li, K.; Liu, S.; Niu, G.; Wei, M. The Complete Mitochondrial Genome of Cimbex Luteus (Hymenoptera: Cimbicidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2021, 6, 2031–2032. [Google Scholar] [CrossRef]
- Yan, Y.; Niu, G.; Zhang, Y.; Ren, Q.; Du, S.; Lan, B.; Wei, M. Complete Mitochondrial Genome Sequence of Labriocimbex Sinicus, a New Genus and New Species of Cimbicidae (Hymenoptera) from China. PeerJ 2019, 7, e7853. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Yan, Y.; Wei, M.; Niu, G. Characterization of Mitochondrial Genomes of Three New Species: Leptocimbex Praiaformis, L. Clavicornis, and L. Yanniae (Hymenoptera: Cimbicidae). Entomol. Res. 2021, 51, 287–304. [Google Scholar] [CrossRef]
- Cheng, Y.; Yan, Y.; Wei, M.; Niu, G. The Complete Mitochondrial Genome of Praia Tianmunica (Hymenoptera: Cimbicidae) with Related Phylogenetic Analysis. Mitochondrial DNA Part B 2020, 5, 3037–3038. [Google Scholar] [CrossRef] [PubMed]
- Song, S.-N.; Tang, P.; Wei, S.-J.; Chen, X.-X. Comparative and Phylogenetic Analysis of the Mitochondrial Genomes in Basal Hymenopterans. Sci. Rep. 2016, 6, 20972. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wei, M.; Yang, H.; Wang, H.; Niu, G. Nearly Complete Mitochondrial Genome of Trichiosoma Vitellina Linné, 1760 (Hymenoptera: Tenthredinidae): Sequencing and Phylogenetic Analysis. Mitochondrial DNA Part B 2020, 5, 802–803. [Google Scholar] [CrossRef]
- Doğan, Ö.; Korkmaz, E.M. Nearly Complete Mitogenome of Hairy Sawfly, Corynis Lateralis (Brullé, 1832) (Hymenoptera: Cimbicidae): Rearrangements in the IQM and ARNS1EF Gene Clusters. Genetica 2017, 145, 341–350. [Google Scholar] [CrossRef]
- Aydemir, M.N.; Korkmaz, E.M. Comparative Mitogenomics of Hymenoptera Reveals Evolutionary Differences in Structure and Composition. Int. J. Biol. Macromol. 2020, 144, 460–472. [Google Scholar] [CrossRef]
- Wei, S.-J.; Niu, F.-F.; Du, B.-Z. Rearrangement of TrnQ-TrnM in the Mitochondrial Genome of Allantus Luctifer (Smith) (Hymenoptera: Tenthredinidae). Mitochondrial DNA 2016, 27, 856–858. [Google Scholar] [CrossRef]
- Yang, J.; Sun, Z.; Wei, M.; Niu, G. The Complete Mitochondrial Genome of Allantus Togatus (Panzer, 1801), in View of Possible Cryptic Species. Mitochondrial DNA Part B 2021, 6, 1114–1115. [Google Scholar] [CrossRef]
- Liu, J.; Wei, M.; Niu, G. The Complete Mitochondrial Genome of Hemathlophorus Brevigenatus Wei, 2005 (Hymenoptera: Tenthredinidae) with Phylogenetic Analysis. Mitochondrial DNA Part B 2021, 6, 2759–2761. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wei, M.; Niu, G. The Complete Mitochondrial Genome of Taxoblenus Sinicus Wei & Nie, 1999 (Hymenoptera: Tenthredinidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2021, 6, 2061–2062. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Tan, B.; Wei, M.; Niu, G. The Complete Mitochondrial Genome Sequence of Taxonus Zhangi Wei, 1997 (Hymenoptera: Tenthredinidae) with Phylogenetic Analysis. Mitochondrial DNA Part B 2021, 6, 3188–3189. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wei, M.; Liu, J.; Niu, G. Characterization of the Mitochondrial Genome of Eutomostethus Vegetus Konow, 1898 (Hymenoptera: Tenthredinidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2020, 5, 3033–3034. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Wei, M.; Liu, M.; Niu, G. Advancement in Sequencing the Mitochondrial Genome of Birmella Discoidalisa Wei, 1994 (Hymenoptera: Tenthredinidae) and the Phylogenetic Classification of Fenusini. Mitochondrial DNA Part B 2019, 4, 4100–4101. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Wei, M.; Niu, G. The Complete Mitochondrial Genome Sequence of Sinopoppia Nigroflagella Wei, 1997 (Hymenoptera: Tenthredinidae) Reveals a New Gene Order. Mitochondrial DNA Part B 2021, 6, 999–1000. [Google Scholar] [CrossRef]
- Wu, D.; Wang, H.; Wei, M.; Niu, G. The Nearly Complete Mitochondrial Genome of Colochela Zhongi Wei, 2016 (Hymenoptera: Tenthredinidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2020, 5, 3341–3342. [Google Scholar] [CrossRef]
- Luo, X.; Wei, M.; Niu, G. Nearly Complete Mitochondrial Genome of Siobla Xizangensis Xiao, Huang & Zhou, 1988 (Hymenoptera: Tenthredinidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2019, 4, 4102–4103. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, M.; Wei, M.; Li, Z. The Complete Mitochondrial Genome of Eriocampa Ovata Linne, 1760 (Hymenoptera: Tenthredinidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2015, 29, 164–190. [Google Scholar]
- Song, S.-N.; Wang, Z.-H.; Li, Y.; Wei, S.-J.; Chen, X.-X. The Mitochondrial Genome of Tenthredo Tienmushana (Takeuchi) and a Related Phylogenetic Analysis of the Sawflies (Insecta: Hymenoptera). Mitochondrial DNA Part A 2016, 27, 2860–2861. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, M.; Niu, G. The First Mitochondrial Genome of a Fern Sawfly, Strongylogaster Xanthocera Stephens, 1835 (Hymenoptera: Tenthredinidae). Mitochondrial DNA Part B 2021, 6, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, Q.; Liu, Y.; Pu, D.; Chen, Z.; Liu, X. The Complete Mitochondrial Genome of a Sawfly Species, Analcellicampa Danfengensis (Hymenoptera: Tenthredinidae). Mitochondrial DNA Part B 2019, 4, 3124–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.-J.; Wu, Q.-L.; Liu, W. Sequencing and Characterization of the Monocellicampa Pruni (Hymenoptera: Tenthredinidae) Mitochondrial Genome. Mitochondrial DNA 2015, 26, 157–158. [Google Scholar] [CrossRef]
- Tan, B.; Wei, M.; Niu, G. The Complete Mitochondrial Genome of Moricella Rufonota Rohwer, 1916 (Hymenoptera: Tenthredinidae) and Phylogenetic Analysis. Mitochondrial DNA Part B 2021, 6, 2285–2287. [Google Scholar] [CrossRef]
- Zheng, B.-Y.; Li, Z.-K.; Li, X.-F.; Zhu, J.-C.; Sharkey, M.; Tang, P.; Chen, X.-X. The First Mitochondrial Genome of the Living-Fossil Sawfly Macroxyela Ferruginea (Hymenoptera: Xyelidae, Macroxyelinae). Mitochondrial DNA Part B 2020, 5, 23–24. [Google Scholar] [CrossRef]
- Zhang, F.; Ding, Y.; Zhu, C.; Zhou, X.; Orr, M.C.; Scheu, S.; Luan, Y. Phylogenomics from Low-coverage Whole-genome Sequencing. Methods Ecol. Evol. 2019, 10, 507–517. [Google Scholar] [CrossRef]
- Bushnell, B. BBtools. Available online: Https://Sourceforge.Net/Projects/Bbmap/ (accessed on 1 October 2021).
- Chikhi, R.; Rizk, G. Space-Efficient and Exact de Bruijn Graph Representation Based on a Bloom Filter. Algorithms Mol. Biol. 2013, 8, 22. [Google Scholar] [CrossRef]
- Pryszcz, L.P.; Gabaldón, T. Redundans: An Assembly Pipeline for Highly Heterozygous Genomes. Nucleic Acids Res. 2016, 44, e113. [Google Scholar] [CrossRef]
- Sahlin, K.; Chikhi, R.; Arvestad, L. Assembly Scaffolding with PE-Contaminated Mate-Pair Libraries. Bioinformatics 2016, 32, 1925–1932. [Google Scholar] [CrossRef]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An Empirically Improved Memory-Efficient Short-Read de Novo Assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved Annotation of Protein-Coding Genes Boundaries in Metazoan Mitochondrial Genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef]
- Niu, G.; Zhang, Y.; Li, Z.; Wei, M. Characterization of the Mitochondrial Genome of Analcellicampa Xanthosoma Gen. et Sp. Nov. (Hymenoptera: Tenthredinidae). PeerJ 2019, 7, e6866. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An Integrated and Scalable Desktop Platform for Streamlined Molecular Sequence Data Management and Evolutionary Phylogenetics Studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Lartillot, N.; Rodrigue, N.; Stubbs, D.; Richer, J. PhyloBayes MPI: Phylogenetic Reconstruction with Infinite Mixtures of Profiles in a Parallel Environment. Syst. Biol. 2013, 62, 611–615. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Aberer, A.J.; Kobert, K.; Stamatakis, A. ExaBayes: Massively Parallel Bayesian Tree Inference for the Whole-Genome Era. Mol. Biol. Evol. 2014, 31, 2553–2556. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1.4.2, A Graphical Viewer of Phylogenetic Trees. Available online: Http://Tree.Bio.Ed.Ac.Uk/Softw./Figtree/ (accessed on 1 January 2014).
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Malagón-Aldana, L.A.; Jensen, A.R.; Smith, D.R.; Shinohara, A.; Vilhelmsen, L. Sawflies out of Gondwana: Phylogenetics and Biogeography of Argidae (Hymenoptera). Syst. Entomol. 2022, 47, 231–258. [Google Scholar] [CrossRef]
- Nel, A.; Roques, P.; Nel, P.; Prokin, A.A.; Bourgoin, T.; Prokop, J.; Szwedo, J.; Azar, D.; Desutter-Grandcolas, L.; Wappler, T.; et al. The Earliest Known Holometabolous Insects. Nature 2013, 503, 257–261. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Sterelny, K. Another View of Life. Stud. Hist. Philos. Sci. Part C Stud. Hist. Philos. Biol. Biomed. Sci. 2005, 36, 585–593. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P. Proishozhdenie i Jevoljucija Nizshih Pereponchatokrylyh. [Origin and Evolution of the Lower Hymenoptera.]. Tr. Paleontol. Inst. Acad. Nauk SSSR 1969, 123, 1–196. [Google Scholar]
- Rasnitsyn, A.P. Proishozhdenie i Jevoljucija Pereponchatokrylyh Nasekomyh. [Origin and Evolution of Hymenoptera.]. Tr. Paleontol. Inst. Acad. Nauk SSSR 1980, 174, 1–192. [Google Scholar]
- Piton, L. Paléontologie Du Gisement Éocéne de Menat (Puy-de-Dôme) (Flore et Faune); Imprimeries P. Vallier: Clermont-Ferrand, France, 1940; p. 303. [Google Scholar]
- Stocsits, R.R.; Letsch, H.; Hertel, J.; Misof, B.; Stadler, P.F. Accurate and Efficient Reconstruction of Deep Phylogenies from Structured RNAs. Nucleic Acids Res. 2009, 37, 6184–6193. [Google Scholar] [CrossRef]
- Cockerell, T.D.A. Fossil Hymenoptera from Florissant, Colorado. Bull. Mus. Comp. Zoölogy Harv. Coll. 1906, 50, 3–58. [Google Scholar]
- Li, H.-T.; Yi, T.-S.; Gao, L.-M.; Ma, P.-F.; Zhang, T.; Yang, J.-B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of Angiosperms and the Puzzle of the Jurassic Gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Cardinal-McTeague, W.M.; Sytsma, K.J.; Hall, J.C. Biogeography and Diversification of Brassicales: A 103 Million Year Tale. Mol. Phylogenetics Evol. 2016, 99, 204–224. [Google Scholar] [CrossRef]
- Yu, X.-Q.; Maki, M.; Drew, B.T.; Paton, A.J.; Li, H.-W.; Zhao, J.-L.; Conran, J.G.; Li, J. Phylogeny and Historical Biogeography of Isodon (Lamiaceae): Rapid Radiation in South-West China and Miocene Overland Dispersal into Africa. Mol. Phylogenetics Evol. 2014, 77, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Opitz, S.E.W.; Boevé, J.-L.; Nagy, Z.T.; Sonet, G.; Koch, F.; Müller, C. Host Shifts from Lamiales to Brassicaceae in the Sawfly Genus Athalia. PLoS ONE 2012, 7, e33649. [Google Scholar] [CrossRef] [Green Version]
- Jablonski, D.; Bottjer, D.J. The Origin and Diversification of Major Groups: Environmental Patterns and Macroevolutionary Lags. In Major Evolutionary Radiation; Taylor, P.D., Larwood, G.P., Eds.; Clarendon Press: Oxford, UK, 1991; pp. 17–57. [Google Scholar]
- Stroud, J.T.; Losos, J.B. Ecological Opportunity and Adaptive Radiation. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 507–532. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Accession Number | References | BUSCO | Matrix | Voucher Number |
|---|---|---|---|---|---|---|
| Blasticotomidae | Runaria striata | ON964462 | This study | / | / | CSCS-Hym-MC0014 |
| Runaria punctata | ON808427 | This study | 5321 | 4877 | CSCS-Hym-MC0019 | |
| Blasticotoma minuta | ON964461 | This study | 5576 | 5115 | CSCS-Hym-MC0060 | |
| Blasticotoma tegularis | ON840089 | This study | 5484 | 5037 | CSCS-Hym-MC0083 | |
| Blasticotoma filiceti | ON840091 | This study | 5343 | 4901 | CSCS-Hym-MC0140 | |
| Pergidae | Perga condei | AY787816 | [62] | / | / | / |
| Pergagrapta polita | / | SRX642961 | 2292 | 2132 | / | |
| Argidae | Arge bella | MF287761 | [63] | / | / | CSCS-Hym-MC0008 |
| Arge similis | MG923484 | [64] | / | / | / | |
| Arge aurora | MN913350 | This study | 5298 | 4864 | CSCS-Hym-MC0179 | |
| Aproceros leucopoda | / | SRX642913 | 3615 | 3378 | / | |
| Athaliidae | Athalia icar | MN527306 | [65] | 2184 | 1934 | CSCS-Hym-MC0021 |
| Athalia birmanica | ON840085 | This study | 5789 | 5312 | CSCS-Hym-MC0077 | |
| Athalia japonica | ON964466 | This study | 5713 | 5251 | CSCS-Hym-MC0081 | |
| Athalia sikkimensis | ON840087 | This study | 5672 | 5219 | CSCS-Hym-MC0345 | |
| Athalia scapulata | ON840088 | This study | 5662 | 5193 | CSCS-Hym-MC0080 | |
| Athalia arunachalensis | ON840086 | This study | 5794 | 5320 | CSCS-Hym-MC0355 | |
| Athalia rosae | / | ERX8976896 | 4480 | 4165 | / | |
| Athalia ‘tibetana’ | ON964467 | This study | / | / | CSCS-Hym-MC0731 | |
| Athalia qingzang | ON964468 | This study | / | / | CSCS-Hym-MC0732 | |
| Athalia tanaoserrula | ON964469 | This study | / | / | CSCS-Hym-MC0733 | |
| Athalia sikkimensis | ON964470 | This study | / | / | CSCS-Hym-MC0734 | |
| Hypsathalia przewalskyi | ON840090 | This study | 5701 | 5240 | CSCS-Hym-MC0187 | |
| Dentathalia scutellariae | ON808426 | This study | 5780 | 5310 | CSCS-Hym-MC0360 | |
| Cimbicidae | Cimbex luteus | MW136447 | [66] | 607 | 502 | CSCS-Hym-MC0035 |
| Labriocimbex sinicus | MH136623 | [67] | 5338 | 4897 | CSCS-Hym-MC0009 | |
| Leptocimbex clavicornis | MT478109 | [68] | 5423 | 4973 | CSCS-Hym-MC0162 | |
| Leptocimbex praiaformis | MT478110 | [68] | / | / | CSCS-Hym-MC0167 | |
| Leptocimbex yanniae | MT478111 | [68] | / | / | CSCS-Hym-MC0133 | |
| Praia tianmunica | MT665975 | [69] | 4092 | 3710 | CSCS-Hym-MC0049 | |
| Asitrichiosoma anthracinum | KT921411 | [70] | / | / | / | |
| Trichiosoma vitellina | MN853777 | [71] | 5186 | 4743 | CSCS-Hym-MC0165 | |
| Corynis lateralis | KY063728 | [72] | / | / | / | |
| Diprionidae | Neodiprion sertifer | MK994526 | [73] | / | / | / |
| Neodiprion qinghaiicus | ON964471 | This study | 5657 | 5192 | CSCS-Hym-MC0198 | |
| Gilpinia baiyinaobaoa | ON840092 | This study | / | / | CSCS-Hym-MC0178 | |
| Nesodiprion biremis | ON964465 | This study | / | / | CSCS-Hym-MC0055 | |
| Nesodiprion japonicus | ON964464 | This study | / | / | CSCS-Hym-MC0010 | |
| Diprion pini | / | SRX642914 | 3933 | 3651 | / | |
| Heptamelinae | Psedoheptamellus lii | ON964463 | This study | 5653 | 5202 | CSCS-Hym-MC0175 |
| Heptamelus brevicercus | MW632128 | [25] | / | / | CSCS-Hym-MC0018 | |
| Tenthredinidae | Allantoides luctifer | KJ713152 | [74] | / | / | / |
| Allantus togatus | MW464859 | [75] | 5676 | 5206 | CSCS-Hym-MC0142 | |
| Asiemphytus rufocephalus | KR703582 | [70] | / | / | / | |
| Empria lii | MW632124 | [25] | 4737 | 4323 | CSCS-Hym-MC0079 | |
| Empronus tibetanus | MZ265343 | [25] | 5148 | 4703 | CSCS-Hym-MC0152 | |
| Hemathlophorus brevigenatus | MW632125 | [76] | 4789 | 4373 | CSCS-Hym-MC0177 | |
| Taxoblenus sinicus | MW632126 | [77] | / | / | CSCS-Hym-MC0193 | |
| Xenapatidea procincta | MW487928 | [25] | / | / | CSCS-Hym-MC0022 | |
| Taxonus zhangi | MZ461490 | [78] | / | / | CSCS-Hym-MC0342 | |
| Beleses atrofemoratus | MZ265347 | [25] | 1520 | 1311 | CSCS-Hym-MC0011 | |
| Eutomostethus vegetus | MT663219 | [79] | 5398 | 4939 | CSCS-Hym-MC0184 | |
| Megatomostethus crassicornis | MZ265345 | [25] | 5682 | 5222 | CSCS-Hym-MC0075 | |
| Phymatoceropsis melanogaster | MZ265346 | [25] | 947 | 792 | CSCS-Hym-MC0032 | |
| Monophadnus latus | / | SRX6352475 | 4237 | 3914 | / | |
| Birmella discoidalisa | MF197548 | [80] | 494 | 399 | CSCS-Hym-MC0029 | |
| Metallus mai | MW255941 | [25] | 5643 | 5168 | CSCS-Hym-MC0182 | |
| Sinopoppia nigroflagella | MW487927 | [81] | 5405 | 4945 | CSCS-Hym-MC0073 | |
| Colochela zhongi | MT702984 | [82] | 5714 | 5246 | CSCS-Hym-MC0061 | |
| Macrophya dolichogaster | MW544890 | [25] | / | / | / | |
| Siobla xizangensis | MN562486 | [83] | 5753 | 5280 | CSCS-Hym-MC0150 | |
| Eriocampa ovata | / | [84] | 5811 | 5337 | CSCS-Hym-MC0143 | |
| Tenthredo tienmushana | KR703581 | [85] | / | / | / | |
| Tenthredo koehleri | / | SRX314899 | 4563 | 4230 | / | |
| Cladiucha magnoliae | MT295305 | [25] | 2104 | 1844 | CSCS-Hym-MC0015 | |
| Cladiucha punctata | MT295306 | [25] | 5733 | 5266 | CSCS-Hym-MC0044 | |
| Megabeleses liriodendrovorax | MW255939 | [25] | 1808 | 1570 | CSCS-Hym-MC0030 | |
| Megabeleses magnoliae | MW255940 | [25] | 5793 | 5316 | CSCS-Hym-MC0042 | |
| Neostromboceros nipponicus | MW632127 | [25] | 817 | 679 | CSCS-Hym-MC0026 | |
| Strongylogaster xanthocera | MW324676 | [86] | 5727 | 5255 | CSCS-Hym-MC0070 | |
| Analcellicampa xanthosoma | MH992752 | [25] | 1250 | 1066 | CSCS-Hym-MC0017 | |
| Analcellicampa danfengensis | MN163004 | [87] | / | / | / | |
| Monocellicampa pruni | JX566509 | [88] | / | / | / | |
| Moricella rufonota | MW487926 | [89] | / | / | CSCS-Hym-MC0068 | |
| Nematus ribesii | / | SRX643001 | 3544 | 3329 | / | |
| Hemibeleses tianmunicus | MZ265344 | [25] | 5072 | 4631 | CSCS-Hym-MC0027 | |
| Xyelidae | Xyela sp. | MG923517 | [64] | / | / | / |
| Xyela alpigena | / | SRX642930 | 3517 | 3308 | / | |
| Megaxyela euchroma | OL794667 | This study | 5393 | 4938 | CSCS-Hym-MC0001 | |
| Macroxyela ferruginea | MK270536 | [90] | / | / | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, G.; Budak, M.; Korkmaz, E.M.; Doğan, Ö.; Nel, A.; Wan, S.; Cai, C.; Jouault, C.; Li, M.; Wei, M. Phylogenomic Analyses of the Tenthredinoidea Support the Familial Rank of Athaliidae (Insecta, Tenthredinoidea). Insects 2022, 13, 858. https://doi.org/10.3390/insects13100858
Niu G, Budak M, Korkmaz EM, Doğan Ö, Nel A, Wan S, Cai C, Jouault C, Li M, Wei M. Phylogenomic Analyses of the Tenthredinoidea Support the Familial Rank of Athaliidae (Insecta, Tenthredinoidea). Insects. 2022; 13(10):858. https://doi.org/10.3390/insects13100858
Chicago/Turabian StyleNiu, Gengyun, Mahir Budak, Ertan Mahir Korkmaz, Özgül Doğan, André Nel, Siying Wan, Chenyang Cai, Corentin Jouault, Min Li, and Meicai Wei. 2022. "Phylogenomic Analyses of the Tenthredinoidea Support the Familial Rank of Athaliidae (Insecta, Tenthredinoidea)" Insects 13, no. 10: 858. https://doi.org/10.3390/insects13100858