
Flying over Eurasia: Geographic Variation of Photoperiodic Control of Nymphal Development and Adult Diapause Induction in Native and Invasive Populations of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae)

 ,
,  , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
- (1)
- Andong, South Korea (ca. 36°41′ N, 128°44′ E; 140 m a.s.l.)—30 individuals were collected in July 2019;
- (2)
- Sochi, Krasnodar Krai, Russia (ca. 43°36′ N, 39°35′ E; 50 m a.s.l.)—more than 100 individuals were collected in July through to August 2019;
- (3)
- Basel, Switzerland (ca. 47°33′ N, 07°36′ E; 260 m a.s.l.)—more than 60 individuals were collected in July through to August 2019;
- (4)
- Torino, Italy (ca. 45°02′ N, 07°35′ E; 240 m a.s.l.)—50 individuals were collected in August 2019.
2.2. Nymphal Development
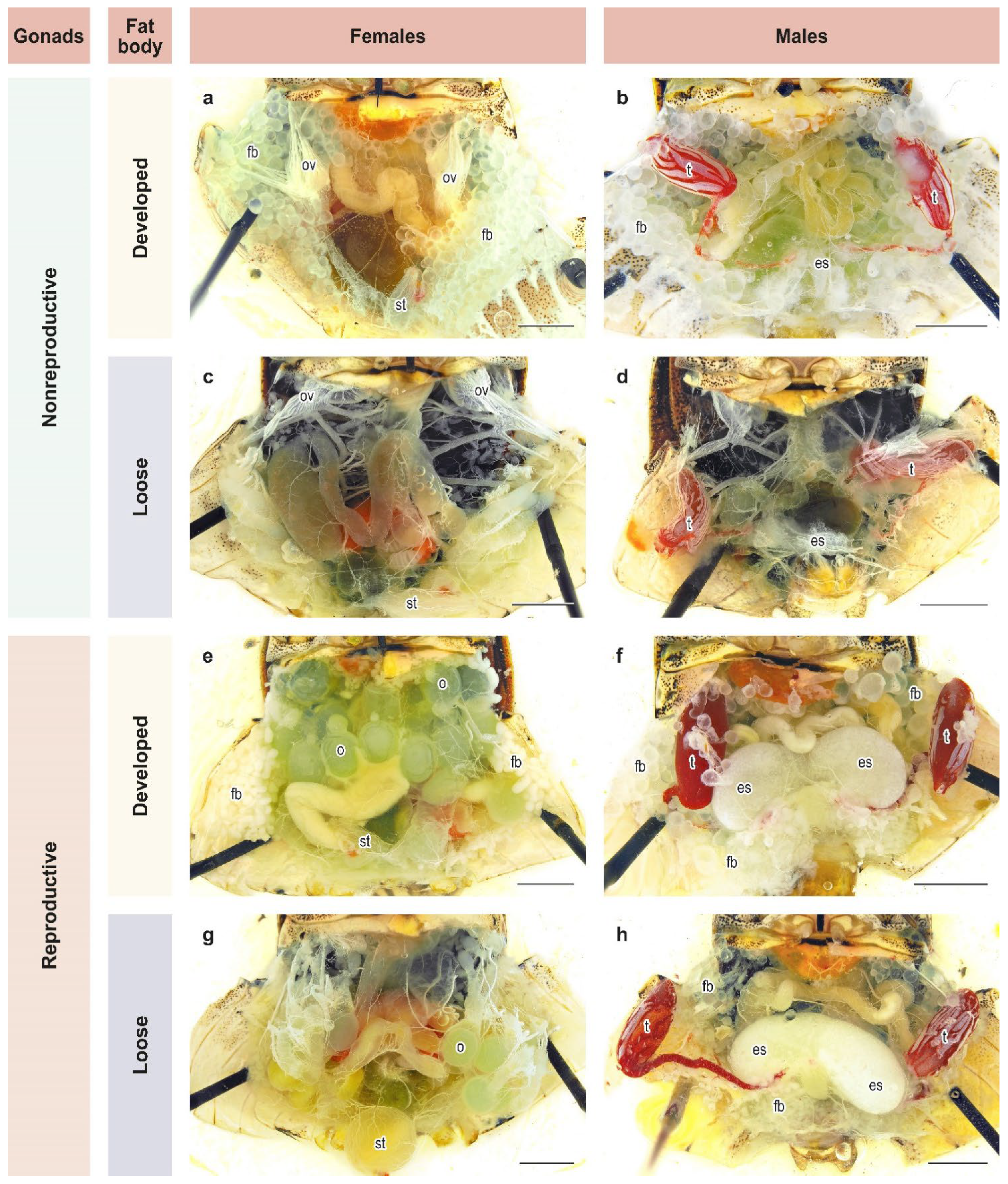
2.3. Development of Reproductive Organs and Fat Body
2.4. Statistical Analysis
3. Results
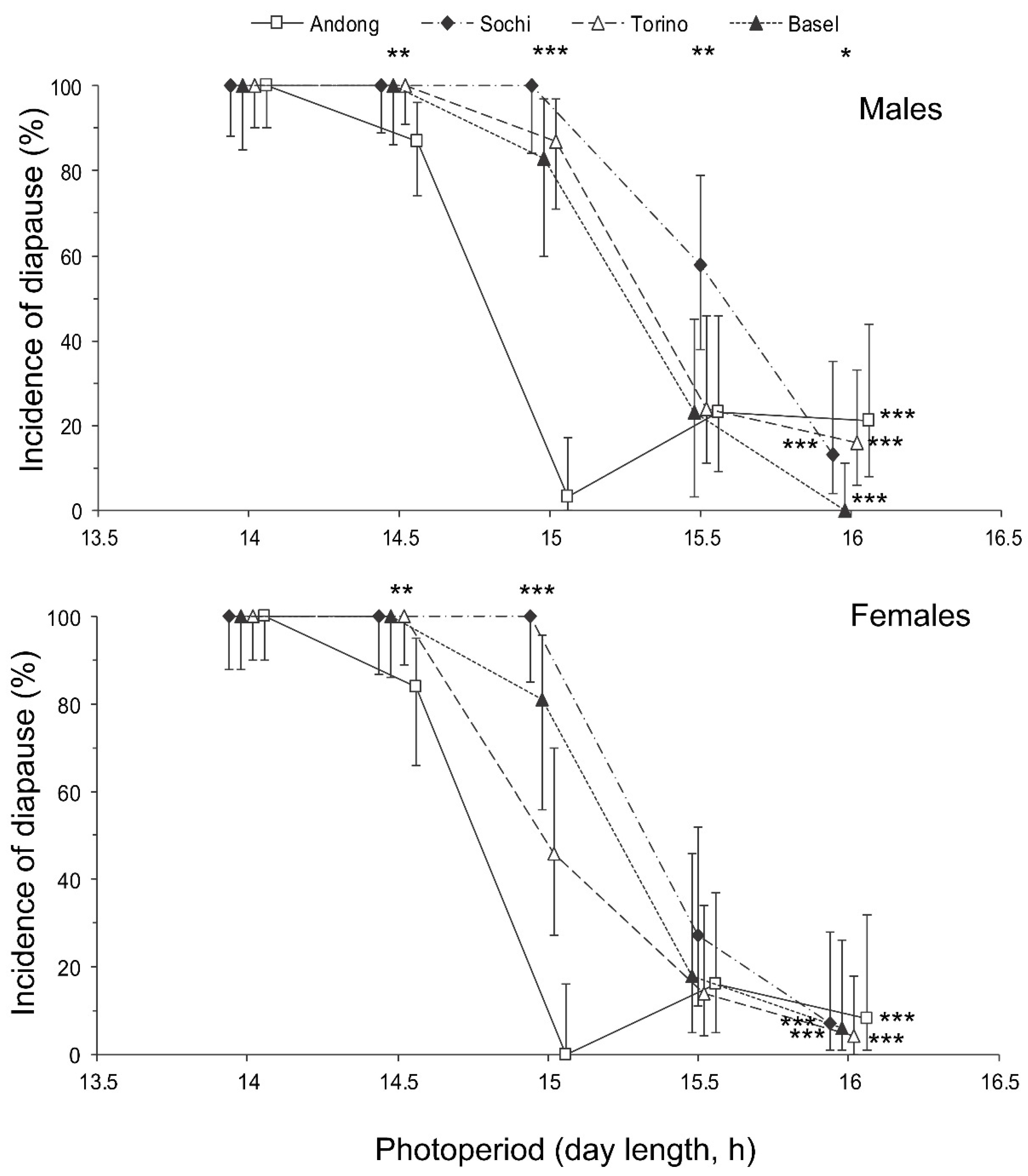
3.1. Nymphal Development
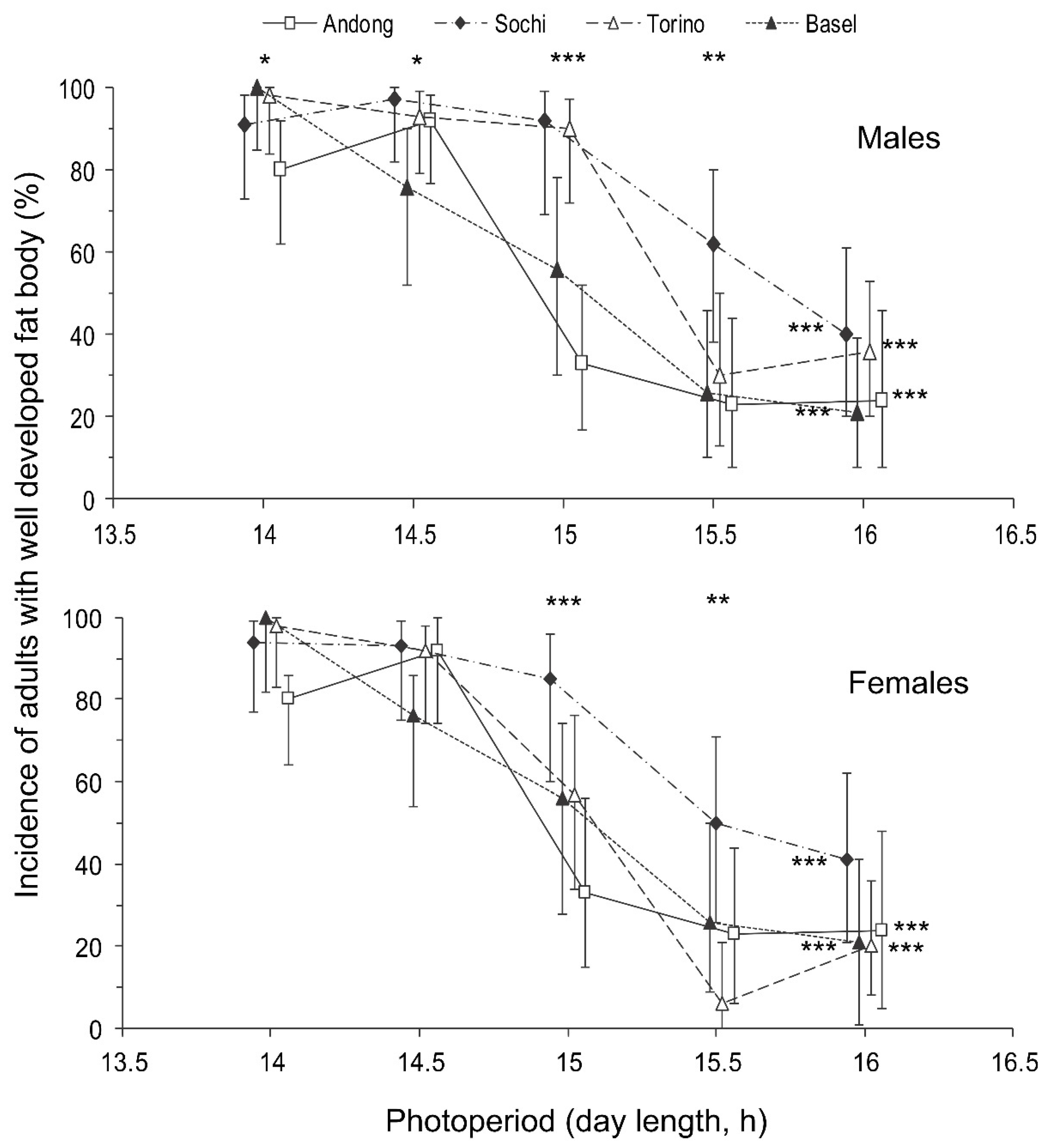
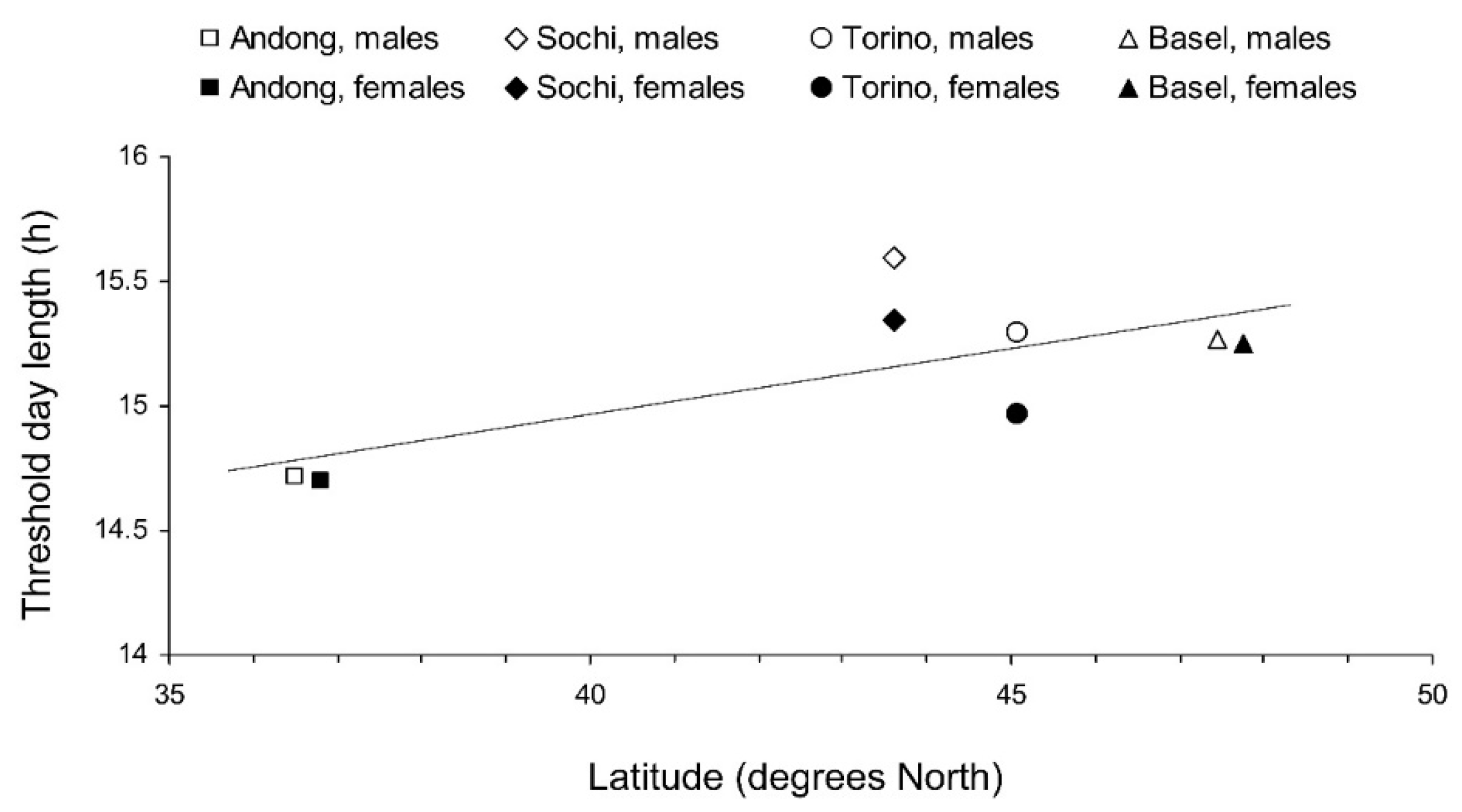
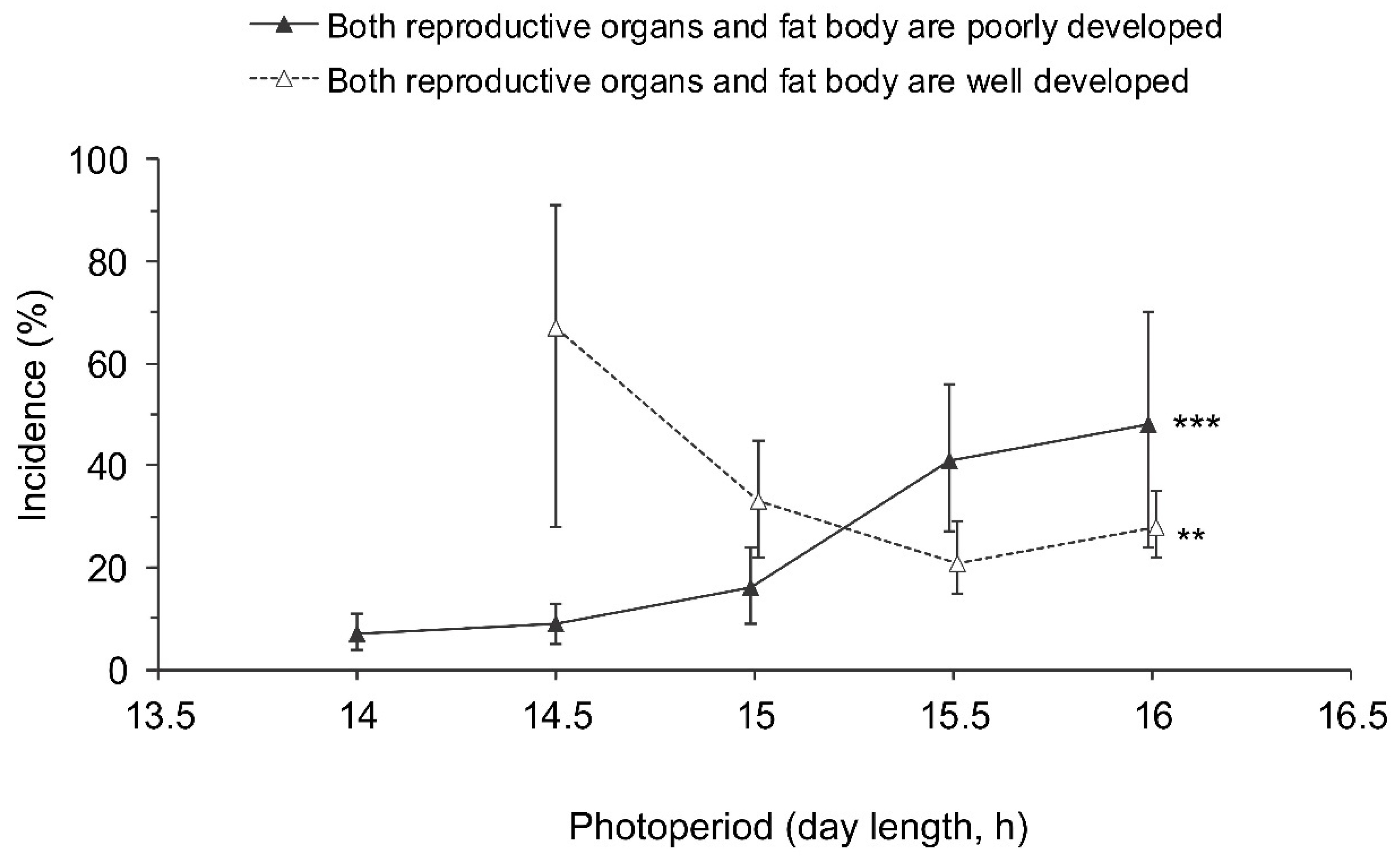
3.2. Development of Reproductive Organs and Fat Body
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simberloff, D. Biological invasions: What’s worth fighting and what can be won? Ecol. Eng. 2014, 65, 112–121. [Google Scholar] [CrossRef]
- Pimentel, D.; McNair, S.; Janecka, J.; Wightman, J.; Simmonds, C.; O’Connell, C.; Won, E.; Russel, L.; Zern, J.; Aquino, T.; et al. Economic and environmental threats of alien plant, animal, and microbe invasions. Agric. Ecosyst. Environ. 2001, 84, 1–20. [Google Scholar] [CrossRef]
- Fei, S.; Phillips, J.; Shouse, M. Biogeomorphic impacts of invasive species. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; van Kleunen, M.; Winter, M.; et al. The global rise in emerging alien species results from increased accessibility of new source pools. Proc. Natl. Acad. Sci. USA 2018, 115, E2264–E2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Handley, L.J.L.; Estoup, A.; Evans, D.M.; Thomas, C.E.; Lombaert, E.; Facon, B.; Aebi, A.; Roy, H.E. Ecological genetics of invasive alien species. BioControl 2011, 56, 409–428. [Google Scholar] [CrossRef] [Green Version]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: New York, NY, USA, 1986. [Google Scholar]
- Lee, C.E. Evolutionary genetics of invasive species. Trends Ecol. Evol. 2002, 17, 386–391. [Google Scholar] [CrossRef]
- Maslyakov, V.Y.; Izhevsky, S.S. Alien Phytophagous Insects Invasions in the European Part of Russia; Institute of Geography of Russian Academy of Sciences: Moscow, Russia, 2011. (In Russian) [Google Scholar]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef]
- Denlinger, D.L. Insect Diapause; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Saunders, D.S.; Steel, C.G.H.; Vafopoulou, X.; Lewis, R.D. Insect Clocks; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Danks, H.V. Insect Dormancy: An Ecological Perspective; The Biological Survey of Canada: Ottawa, ON, Canada, 1987. [Google Scholar]
- Danks, H.V. The elements of seasonal adaptations in insects. Can. Entomol. 2007, 139, 1–44. [Google Scholar] [CrossRef]
- Tougeron, K. Diapause research in insects: Historical review and recent work perspectives. Entomol. Exp. Appl. 2019, 167, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Urbanski, J.; Mogi, M.; O’Donnell, D.; DeCotiis, M.; Toma, T.; Armbruster, P. Rapid adaptive evolution of photoperiodic response during invasion and range expansion across a climatic gradient. Am. Nat. 2012, 179, 490–500. Available online: https://www.journals.uchicago.edu/doi/abs/10.1086/664709 (accessed on 1 March 2022). [CrossRef] [PubMed]
- Reznik, S.Y.; Dolgovskaya, M.Y.; Ovchinnikov, A.N.; Belyakova, N.A. Weak photoperiodic response facilitates the biological invasion of the harlequin ladybird Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). J. Appl. Entomol. 2015, 139, 241–249. [Google Scholar] [CrossRef]
- Reznik, S.Y.; Ovchinnikov, A.N.; Dolgovskaya, M.Y.; Belyakova, N.A. Interspecific variation in photoperiodic effect on the rate of preimaginal development in Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae). Entomol. Rev. 2015, 95, 15–27. [Google Scholar] [CrossRef]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Haye, T.; Gariepy, T.; Hoelmer, K.; Rossi, J.P.; Streito, J.C.; Tassus, X.; Desneux, N. Range expansion of the invasive brown marmorated stinkbug, Halyomorpha halys: An increasing threat to field, fruit and vegetable crops worldwide. J. Pest Sci. 2015, 88, 665–673. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Musolin, D.L.; Konjević, A.; Karpun, N.N.; Zakharchenko, V.Y.; Zhuravleva, E.N.; Tavella, L.; Bruin, A.; Haye, T. Diversity and distribution of cytochrome oxidase I (COI) haplotypes of the brown marmorated stink bug, Halyomorpha halys Stål (Hemiptera, Pentatomidae), along the eastern front of its invasive range in Eurasia. NeoBiota 2021, 68, 53–77. [Google Scholar] [CrossRef]
- Hamilton, G.C.; Ahn, J.J.; Bu, W.; Leskey, T.C.; Nielsen, A.L.; Park, Y.L.; Rabitsch, W.; Hoelmer, K.A. Halyomorpha halys (Stål). In Invasive Stink Bugs and Related Species (Pentatomoidea); McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 243–292. [Google Scholar] [CrossRef]
- Musolin, D.L.; Kirichenko, N.I.; Karpun, N.N.; Aksenenko, E.V.; Golub, V.B.; Kerchev, I.A.; Mandelshtam, M.Y.; Vasaitis, R.; Volkovitsh, M.G.; Zhuravleva, E.N.; et al. Invasive insect pests of forests and urban trees in Russia: Origin, pathways, damage, and management. Forests 2022, 13, 521. [Google Scholar] [CrossRef]
- Musolin, D.L.; Konjević, A.; Karpun, N.N.; Protsenko, V.Y.; Ayba, L.Y.; Saulich, A.K. Invasive brown marmorated stink bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Serbia: History of invasion, range expansion, early stages of establishment, and first records of damage to local crops. Arthropod-Plant Interact. 2018, 12, 517–529. [Google Scholar] [CrossRef]
- Musolin, D.L.; Dolgovskaya, M.Y.; Protsenko, V.Y.; Karpun, N.N.; Reznik, S.Y.; Saulich, A.K. Photoperiodic and temperature control of nymphal growth and adult diapause induction in the invasive Caucasian population of the brown marmorated stink bug, Halyomorpha halys. J. Pest Sci. 2019, 92, 621–631. [Google Scholar] [CrossRef]
- Watanabe, M. Ecology and extermination of Halyomorpha halys. 4. The relationship between day length and ovarian development. Ann. Rep. Toyama Inst. Health 1979, 3, 33–37. (In Japanese) [Google Scholar]
- Yanagi, T.; Hagihara, Y. Ecology of the brown marmorated stink bug. Plant Prot. 1980, 34, 315–321. (In Japanese) [Google Scholar]
- Niva, C.C.; Takeda, M. Effects of photoperiod, temperature and melatonin on nymphal development, polyphenism and reproduction in Halyomorpha halys (Heteroptera: Pentatomidae). Zool. Sci. 2003, 20, 963–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, A.L.; Hamilton, G.C. Life history of the invasive species Halyomorpha halys (Hemiptera: Pentatomidae) in northeastern United States. Ann. Entomol. Soc. Am. 2009, 102, 608–616. [Google Scholar] [CrossRef] [Green Version]
- Haye, T.; Abdallah, S.; Gariepy, T.; Wyniger, D. Phenology, life table analysis and temperature requirements of the invasive brown marmorated stink bug, Halyomorpha halys, in Europe. J. Pest Sci. 2014, 87, 407–418. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Chen, S.; Fleischer, S.J. Coupling developmental physiology, photoperiod, and temperature to model phenology and dynamics of an invasive heteropteran, Halyomorpha halys. Front. Physiol. 2016, 7, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, A.L.; Fleischer, S.; Hamilton, G.C.; Hancock, T.; Krawczyk, G.; Lee, J.C.; Ogburn, E.; Pote, J.M.; Raudenbush, A.; Rucker, A.; et al. Phenology of brown marmorated stink bug described using female reproductive development. Ecol. Evol. 2017, 7, 6680–6690. [Google Scholar] [CrossRef] [Green Version]
- Basnet, S.; Kuhar, T.P.; Laub, C.A.; Pfeiffer, D.G. Seasonality and distribution pattern of brown marmorated stink bug (Hemiptera: Pentatomidae) in Virginia vineyards. J. Econ. Entomol. 2015, 108, 1902–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, S.; Hwang, A.; Kim, H.; Lee, H.; Lee, J.H. Temperature-dependent development and oviposition models of Halyomorpha halys (Hemiptera: Pentatomidae). J. Asia-Pac. Entomol. 2017, 20, 367–375. [Google Scholar] [CrossRef]
- Kamiyama, M.T.; Matsuura, K.; Yoshimura, T.; Yang, C.C.S. Improving invasive species management using predictive phenology models: An example from brown marmorated stink bug (Halyomorpha halys) in Japan. Pest Manag. Sci. 2021, 77, 5489–5497. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Bu, W.; Gao, Y.; Liu, G. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef] [PubMed]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Kistner, E.J. Climate change impacts on the potential distribution and abundance of the brown marmorated stink bug (Hemiptera: Pentatomidae) with special reference to North America and Europe. Environ. Entomol. 2017, 46, 1212–1224. [Google Scholar] [CrossRef]
- Stoeckli, S.; Felber, R.; Haye, T. Current distribution and voltinism of the brown marmorated stink bug, Halyomorpha halys, in Switzerland and its response to climate change using a high-resolution CLIMEX model. Int. J. Biometeorol. 2020, 64, 2019–2032. [Google Scholar] [CrossRef]
- Tytar, V.M.; Kozynenko, I.I. Bioclimatic modeling of the distribution of brown marmorated stink bug Halyomorpha halys (Stål, 1855), with special reference to Ukraine. Dopov. Nac. Akad. Nauk Ukr. 2020, 2, 82–86. [Google Scholar] [CrossRef]
- Streito, J.C.; Chartois, M.; Pierre, É.; Dusoulier, F.; Armand, J.M.; Gaudin, J.; Rossi, J.P. Citizen science and niche modeling to track and forecast the expansion of the brown marmorated stinkbug Halyomorpha halys (Stål, 1855). Sci. Rep. 2021, 11, 11421. [Google Scholar] [CrossRef]
- Dolgovskaya, M.Y.; Reznik, S.Y.; Zakharchenko, V.Y.; Karpun, N.N.; Musolin, D.L. Changes in the photoperiodic response of the winter adult diapause induction in the brown marmorated stink bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) during its invasion into Europe. In Proceedings of the All-Russia Conference with International Participation Dendrobiotic Invertebrates and Fungi and Their Role in Forest Ecosystems (the Kataev Memorial Readings XI), St. Petersburg, Russia, 24–27 November 2020; Musolin, D.L., Kirichenko, N.I., Selikhovkin, A.V., Eds.; St. Petersburg State Forest Technical University: St. Petersburg, Russia, 2020; pp. 145–146. (In Russian). [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M. Study of the life cycle of the brown marmorated stink bug, Halyomorpha mista. Insectarium 1980, 17, 168–173. (In Japanese) [Google Scholar]
- Nakamura, K.; Numata, H. Seasonal life cycle of Aelia fieberi (Hemiptera: Pentatomidae) in relation to the phenology of its host plants. Ann. Entomol. Soc. Am. 1997, 90, 625–630. [Google Scholar] [CrossRef]
- Nakamura, K.; Numata, H. Environmental regulation of adult diapause of Graphosoma rubrolineatum (Westwood) (Heteroptera: Pentatomidae) in southern and northern populations of Japan. Appl. Entomol. Zool. 1999, 34, 323–326. [Google Scholar] [CrossRef]
- Nakamura, K.; Numata, H. Effects of photoperiod and temperature on the induction of adult diapause in Dolycoris baccarum (L.) (Heteroptera: Pentatomidae) from Osaka and Hokkaido, Japan. Appl. Entomol. Zool. 2006, 41, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Musolin, D.L.; Numata, H. Photoperiodic and temperature control of diapause induction and colour change in the southern green stink bug Nezara viridula. Physiol. Entomol. 2003, 28, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Esquivel, J.F. Stages of gonadal development of the southern green stink bug (Hemiptera: Pentatomidae): Improved visualization. Ann. Entomol. Soc. Am. 2009, 102, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Esquivel, J.F. Improved visualization of fat body cell conditions and abundance in the southern green stink bug (Hemiptera: Pentatomidae). J. Entomol. Sci. 2011, 46, 52–61. [Google Scholar] [CrossRef]
- Esquivel, J.F.; Musolin, D.L.; Jones, W.A.; Rabitsch, W.; Greene, J.K.; Toews, M.D.; Schwertner, C.F.; Grazia, J.; McPherson, R.M. Nezara viridula (L.). In Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management; McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 351–423. [Google Scholar] [CrossRef]
- Musolin, D.L.; Saulich, A.K. Diapause in Pentatomoidea. In Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management; McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 497–564. [Google Scholar] [CrossRef]
- SYSTAT 10.2. 2021. Available online: https://systat.informer.com/10.2/ (accessed on 1 April 2022).
- Ruberson, J.R.; Yeargan, K.V.; Newton, B.L. Variation in diapause responses between geographic populations of the predator Geocoris punctipes (Heteroptera: Geocoridae). Ann. Entomol. Soc. Am. 2001, 94, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Kawasaki, K. Geographic variability in diapause response of Japanese Orius species. Entomol. Exp. Appl. 2001, 98, 303–316. [Google Scholar] [CrossRef]
- Musolin, D.L.; Ito, K. Photoperiodic and temperature control of nymphal development and induction of reproductive diapause in two predatory Orius bugs: Interspecific and geographic differences. Physiol. Entomol. 2008, 33, 291–301. [Google Scholar] [CrossRef]
- Saulich, A.K.; Musolin, D.L. Seasonal development and ecology of anthocorids (Heteroptera, Anthocoridae). Entomol. Rev. 2009, 89, 501–528. [Google Scholar] [CrossRef]
- Danilevskii, A.S. Photoperiodism and Seasonal Development of Insects; Johnson, J., Waloff, N., Translators; Oliver & Boyd: Edinburgh, UK, 1965. [Google Scholar]
- Nielsen, A.L.; Hamilton, G.C.; Matadha, D. Developmental rate estimation and life table analysis for Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2008, 37, 348–355. [Google Scholar] [CrossRef]
- Saunders, D.S. A diapause induction-termination asymmetry in the photoperiodic responses of the Linden bug, Pyrrhocoris apterus and an effect of near-critical photoperiods on development. J. Insect Physiol. 1983, 29, 399–405. [Google Scholar] [CrossRef]
- Numata, H.; Saulich, A.H.; Volkovich, T.A. Photoperiodic responses of the linden bug, Pyrrhocoris apterus, under conditions of constant temperature and under thermoperiodic conditions. Zool. Sci. 1993, 10, 521–527. Available online: https://ci.nii.ac.jp/naid/110003323090/ (accessed on 1 March 2022).
- Musolin, D.L.; Saulich, A.K. Photoperiodic control of nymphal growth in true bugs (Heteroptera). Entomol. Rev. 1997, 77, 768–780. [Google Scholar]
- Musolin, D.L.; Saulich, A.H. Diversity of seasonal adaptations in terrestrial true bugs (Heteroptera) from the temperate zone. Entomol. Sci. 1999, 2, 623–639. [Google Scholar]
- Musolin, D.L.; Tsytsulina, K.; Ito, K. Photoperiodic and temperature control of reproductive diapause induction in the predatory bug Orius strigicollis (Heteroptera: Anthocoridae) and its implications for biological control. Biol. Control 2004, 31, 91–98. [Google Scholar] [CrossRef]
- Lopatina, E.B.; Balashov, S.V.; Kipyatkov, V.E. First demonstration of the influence of photoperiod on the thermal requirements for development in insects and in particular the linden-bug, Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae). Eur. J. Entomol. 2007, 104, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Saulich, A.K.; Musolin, D.L. Diapause in the seasonal cycle of stink bugs (Heteroptera, Pentatomidae) from the temperate zone. Entomol. Rev. 2012, 92, 1–26. [Google Scholar] [CrossRef]
- Kutcherov, D.; Lopatina, E.B.; Balashov, S. Convergent photoperiodic plasticity in developmental rate in two species of insects with widely different thermal phenotypes. Eur. J. Entomol. 2018, 115, 624–631. [Google Scholar] [CrossRef]
- Lopatina, E.B.; Gusev, I.A. A novel form of phenotypic plasticity of the thermal reaction norms for development in the bug Graphosoma lineatum (L.) (Heteroptera, Pentatomidae). Entomol. Rev. 2019, 99, 417–436. [Google Scholar] [CrossRef]
- Kidokoro, T.; Masaki, S. Photoperiodic response in relation to variable voltinism in the ground cricket, Pteronemobius fascipes Walker (Orthoptera: Gryllidae). Jpn. J. Ecol. 1978, 28, 291–298. [Google Scholar]
- Bradshaw, W.E.; Lounibos, L.P. Photoperiodic control of development in the pitcher-plant mosquito, Wyeomyia smithii. Can. J. Zool. 1972, 50, 713–719. [Google Scholar] [CrossRef]
- Tyshchenko, V.P.; Gasanov, O.G. Comparative studies on photoperiodic regulation of diapause and weight of Lepidoptera pupae. Zool. Zhurnal 1983, 62, 63–68, (In Russian, English summary). [Google Scholar]
- Tyshchenko, V.P.; Lanevich, V.P.; Gasanov, O.G. On the relationships of quantitative and qualitative manifestations of photoperiodism in the cabbage cutworm (Barathra brassicae). J. Gen. Biol. 1977, 38, 264–276, (In Russian, English summary). [Google Scholar]
- Tyshchenko, V.P.; Tyshchenko, G.F. Seasonal adaptations of the Setaceous Hebrew character Graphiphora c-nigrum L. (Lepidoptera, Noctuidae). Entomol. Rev. 1988, 67, 11–19, (In Russian, English summary). [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fat Body | Reproductive Organs | |||
|---|---|---|---|---|
| Poorly Developed | Well Developed | |||
| Males | Females | Males | Females | |
| Poorly developed | 9.0 | 6.3 | 26.6 | 31.1 |
| Well developed | 53.3 | 51.6 | 11.1 | 11.0 |
| Factor or Combination of Factors and Coding | Influence on the Proportion of Individuals with Well Developed Reproductive Organs 1 | Influence on the Proportion of Individuals with Well Developed Fat Body 1 | Influence on the Proportion of Individuals with Both Well Developed Reproductive Organs and Fat Body 2 | Influence on the Proportion of Individuals with Both Poorly Developed Reproductive Organs and Fat Body 3 |
|---|---|---|---|---|
| Photoperiod (day length) | 1.0 ± 0.2, p < 0.001 | –1.3 ± 0.2, p < 0.001 | –0.9 ± 0.3, p = 0.010 | 0.9 ± 0.3, p < 0.001 |
| Population (1—Andong, 2—Torino, 3—Basel, 4—Sochi) | –9.3 ± 1.7, p < 0.001 | –0.5 ± 0.8, p = 0.588 | –3.7 ± 2.4, p = 0.128 | 0.6 ± 1.3, p = 0.630 |
| Sex (0—males, 1—females) | –4.2 ± 3.2, p = 0.192 | 0.8 ± 1.8, p = 0.676 | –4.3 ± 4.7, p = 0.369 | –0.3 ± 3.1, p = 0.928 |
| Population × photoperiod | 0.6 ± 0.1, p < 0.001 | 0.0 ± 0.1, p = 0.425 | 0.2 ± 0.2, p = 0.118 | –0.1 ± 0.1, p = 0.561 |
| Population × sex | 0.0 ± 0.1, p = 0.958 | 0.0 ± 0.1, p = 0.955 | 0.1 ± 0.1, p = 0.658 | 0.1 ± 0.1, p = 0.254 |
| Sex × photoperiod | 0.3 ± 0.2, p = 0.171 | –0.1 ± 0.1, p = 0.649 | 0.3 ± 0.3, p = 0.404 | 0.0 ± 0.2, p = 0.971 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musolin, D.L.; Dolgovskaya, M.Y.; Zakharchenko, V.Y.; Karpun, N.N.; Haye, T.; Saulich, A.K.; Reznik, S.Y. Flying over Eurasia: Geographic Variation of Photoperiodic Control of Nymphal Development and Adult Diapause Induction in Native and Invasive Populations of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae). Insects 2022, 13, 522. https://doi.org/10.3390/insects13060522
Musolin DL, Dolgovskaya MY, Zakharchenko VY, Karpun NN, Haye T, Saulich AK, Reznik SY. Flying over Eurasia: Geographic Variation of Photoperiodic Control of Nymphal Development and Adult Diapause Induction in Native and Invasive Populations of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae). Insects. 2022; 13(6):522. https://doi.org/10.3390/insects13060522
Chicago/Turabian StyleMusolin, Dmitry L., Margarita Yu. Dolgovskaya, Vilena Ye. Zakharchenko, Natalia N. Karpun, Tim Haye, Aida Kh. Saulich, and Sergey Ya. Reznik. 2022. "Flying over Eurasia: Geographic Variation of Photoperiodic Control of Nymphal Development and Adult Diapause Induction in Native and Invasive Populations of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae)" Insects 13, no. 6: 522. https://doi.org/10.3390/insects13060522