
Phenotypic Plasticity of Common Wasps in an Industrially Polluted Environment in Southwestern Finland

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
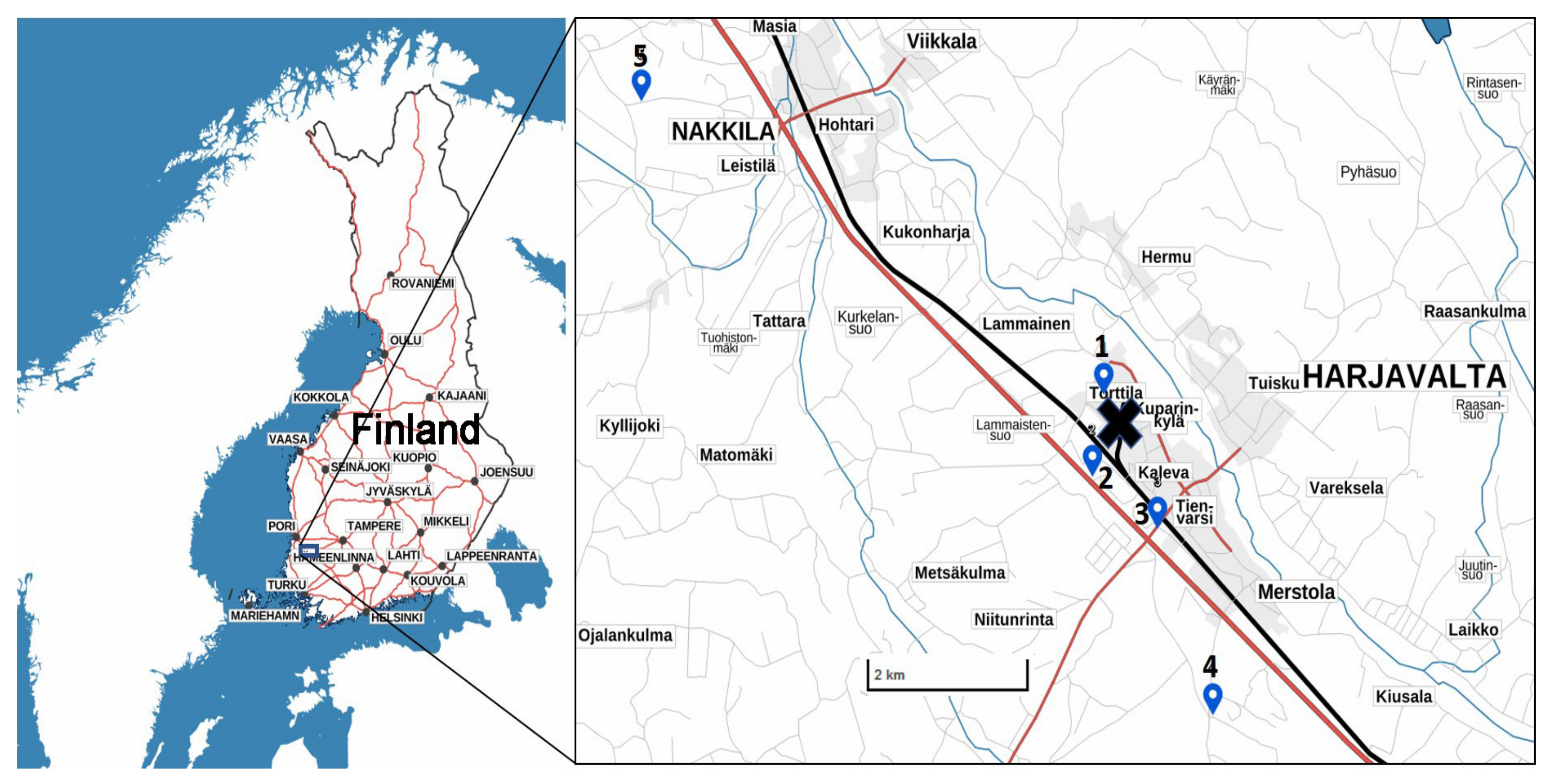
2.1. Sample Collection
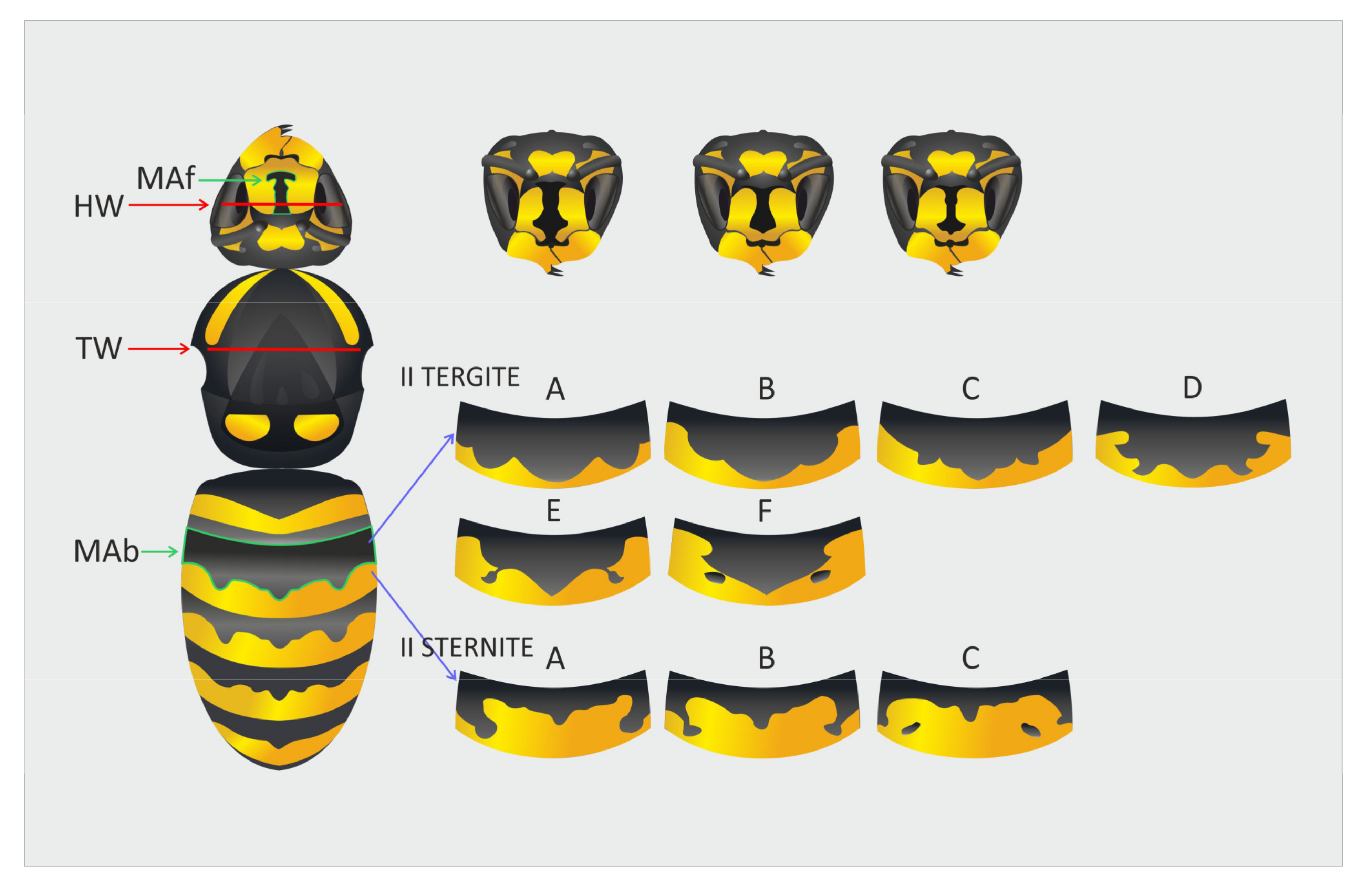
2.2. Morphological Measurements
2.3. Analyses of Color Variation
2.4. Metal Analyses
2.5. Transmission Electron Microscopy (TEM) and Energy-Dispersive X-ray Analyses (EDX)
2.6. Statistical Methods
3. Results
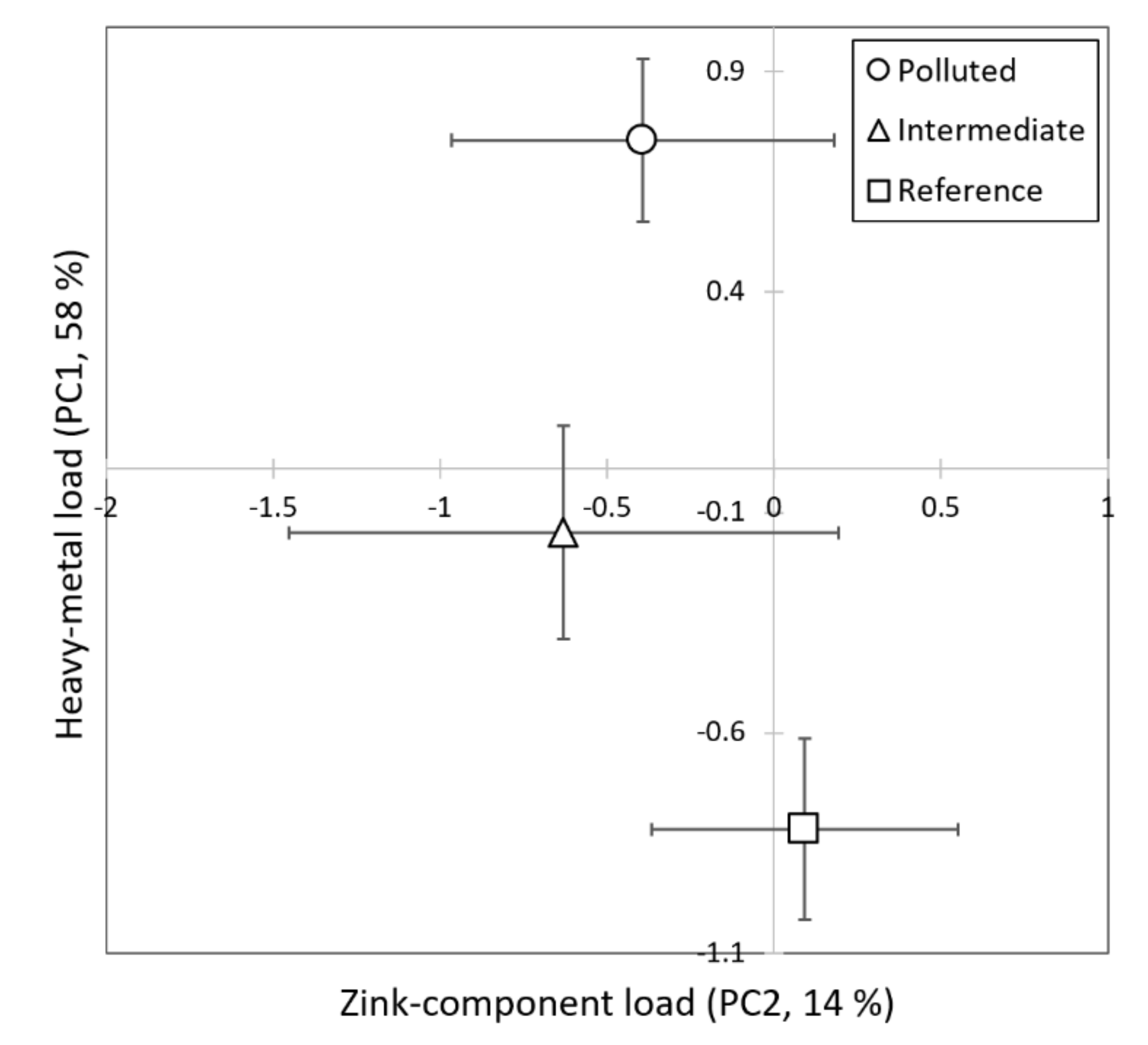
3.1. Metal Concentrations in Wasps
3.2. Body Size & Weight
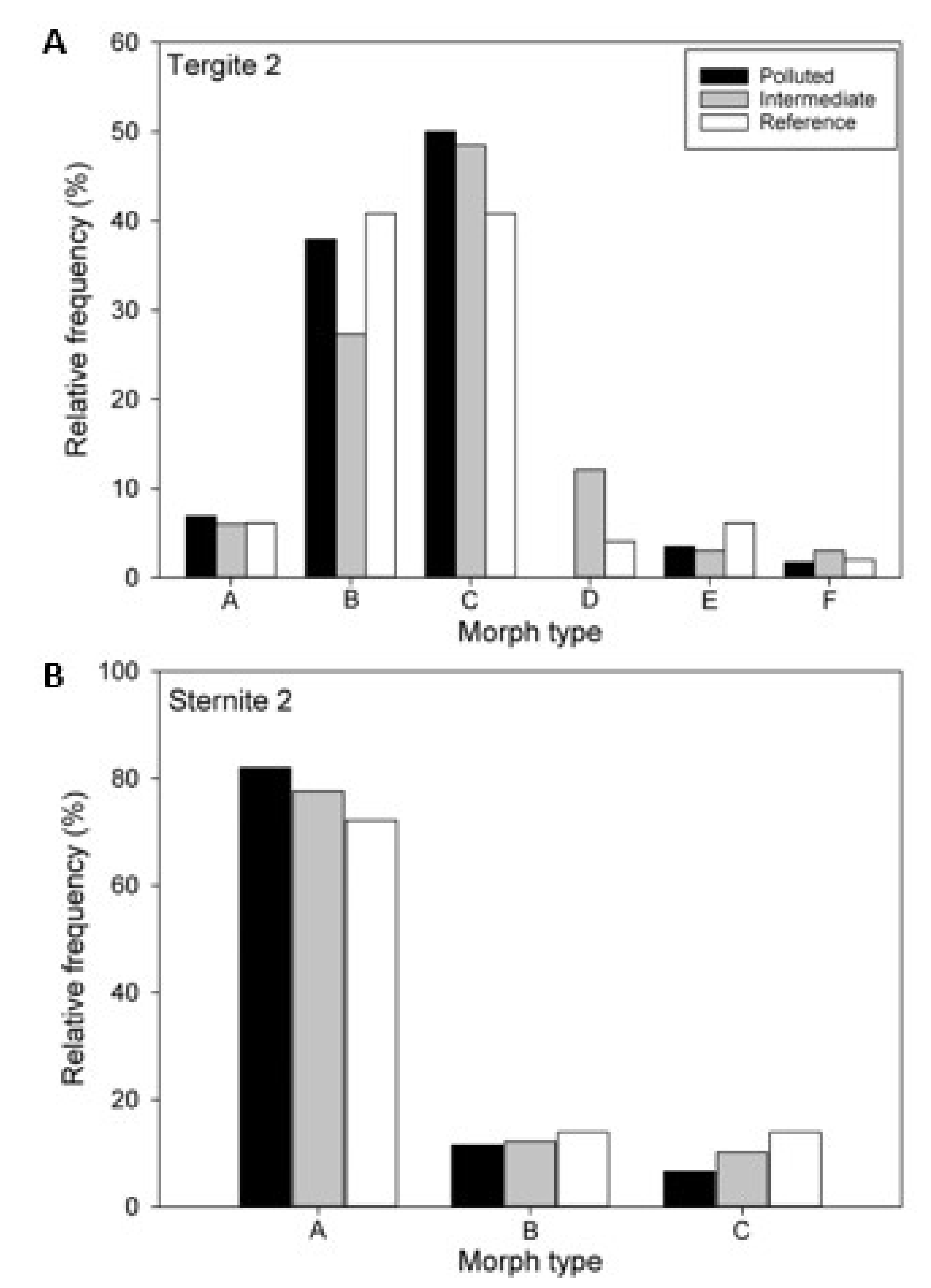
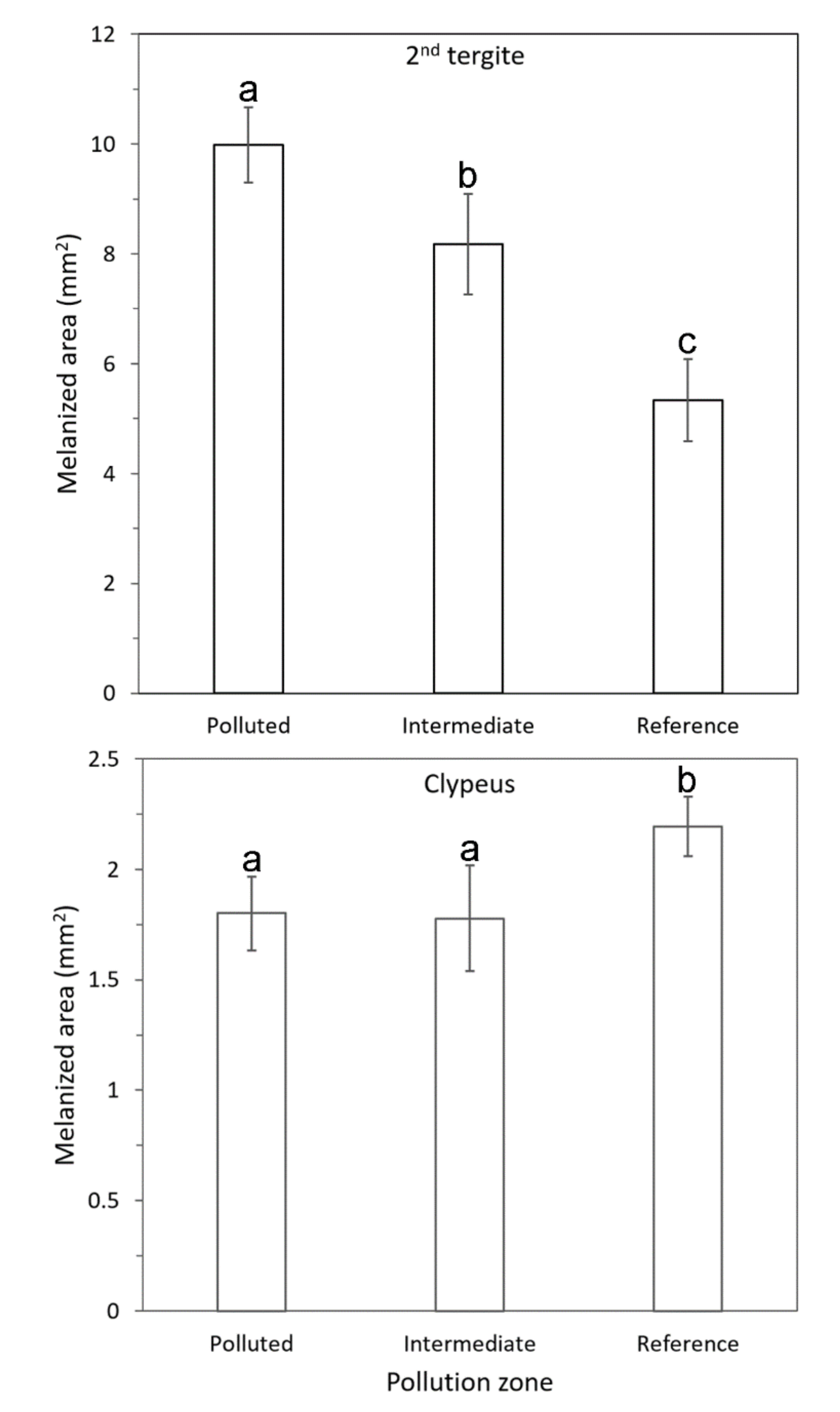
3.3. Color
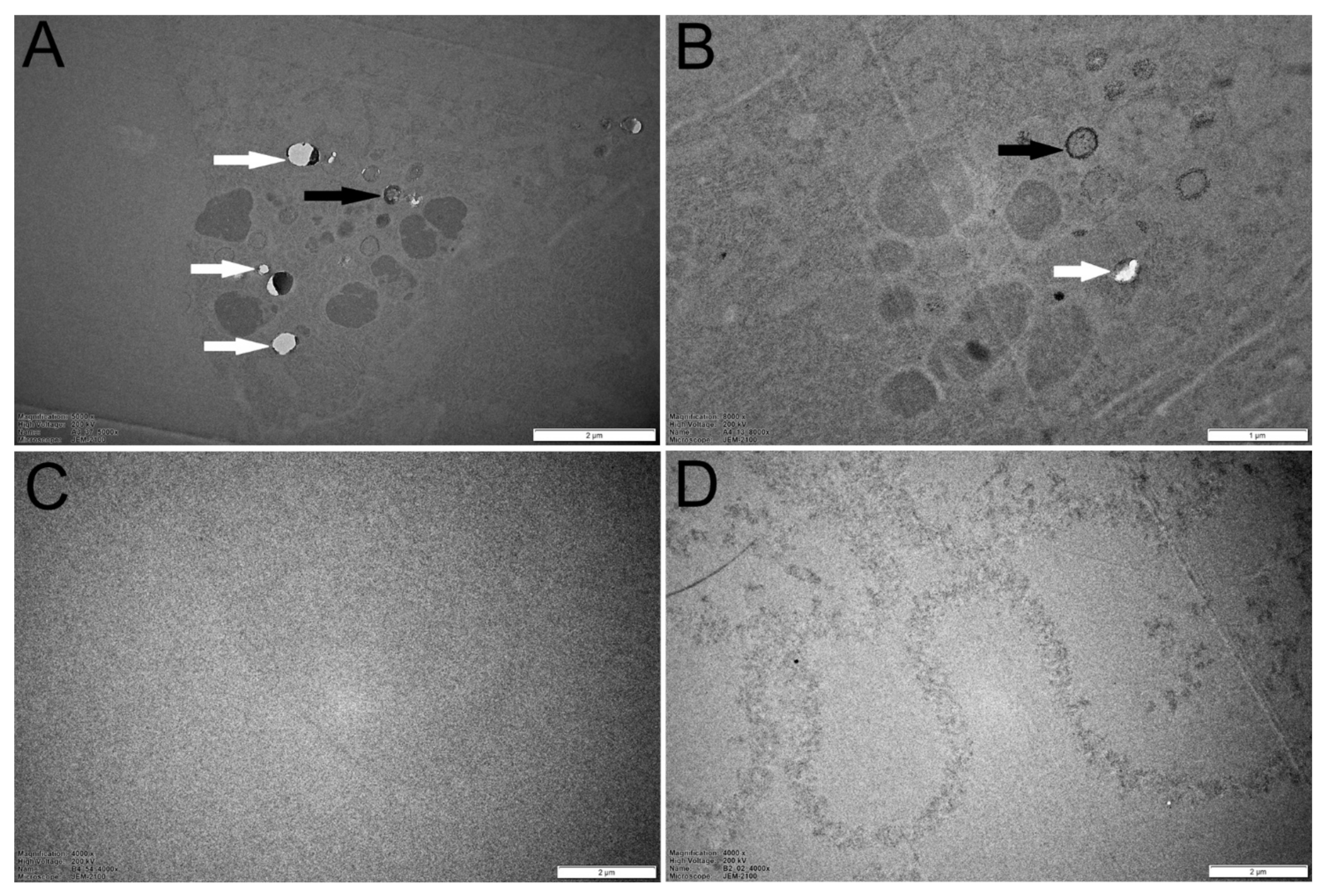
3.4. Transmission Electron Microscopy (TEM) and Energy-Dispersive X-ray Analyses (EDX) Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butler, C.D.; Beckage, N.E.; Trumble, J.T. Effects of terrestrial pollutants on insect parasitoids. Environ. Toxicol. Chem. 2009, 28, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Hillman, R.C.; Benton, A.W. Biological effects of air pollution on insects, emphasizing the reactions of the honeybee (Apis mellifera L.) to sulfur dioxide. J. Elisha Mitchel Sci. Soc. 1972, 88, 195. [Google Scholar]
- Selikhovin, A.V. Factors regulating population density of Lepidoptera in industrially polluted area. In Ecology and Protection of Forests: Interaction of Forest Ecosystem Components; Soloviev, V.A., Ed.; State Forest Technical Academy: Leningrad, Russia, 1985; pp. 123–127. [Google Scholar]
- Du, S.; Liu, Y.; Liu, J.; Zhao, J.; Champagne, C.; Tong, L.; Zhang, R.; Zhang, F.; Qin, C.-F.; Ma, P.; et al. Aedes mosquitoes acquire and transmit Zika virus by breeding in contaminated aquatic environments. Nat. Commun. 2019, 10, 1324. [Google Scholar] [CrossRef]
- Brodmann, J.; Twele, R.; Francke, W.; Hölzler, G.; Zhang, Q.; Ayasse, M. Orchids mimic green-leaf volatiles to attract prey-hunting wasps for pollination. Curr. Biol. 2008, 18, 740–744. [Google Scholar]
- Jules, E.S. Yellow jackets (Vespula vulgaris) as a second seed disperser for the myrmecochorous plant, Trillium ovatum. Am. Midl. Nat. 1996, 135, 367–369. [Google Scholar]
- Richter, M.R. Social wasp (Hymenoptera: Vespidea) foraging behaviour. Annu. Rev. Entomol. 2000, 45, 121–150. [Google Scholar] [PubMed]
- Feldhaar, H.; Otti, O. Pollutants and their interaction with diseases of social hymenoptera. Insects 2020, 11, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancrini, G.; Magi, M.; Gabrielli, S.; Arispici, M.; Tolari, F.; Dell’Omodarme, M.; Prati, M.C. Natural vectors of dirofilariasis in rural and urban areas of the Tuscan region, central Italy. J. Med. Entomol. 2006, 43, 574–579. [Google Scholar]
- Nchoutpouen, E.; Talipouo, A.; Djiappi-Tchamen, B.; Djamouko-Djonkam, L.; Kopya, E.; Ngadjeu, C.S.; Doumbe-Belisse, P.; Awono-Ambene, P.; Kekeunou, S.; Wondji, C.S.; et al. Culex species diversity, susceptibility to insecticides and role as potential vector of Lymphatic filariasis in the city of Yaounde, Cameroon. PLoS Negl. Trop. Dis. 2019, 13, e0007229. [Google Scholar]
- Rono, M.K.; Muturi, C.N.; Ochieng, R.; Mwakubabanya, R.; Wachira, F.N.; Mwangangi, J.; Kinyanjui, S.; Njunge, J.; Mireji, P.O. Cadmium tolerance pathway in Anopheles gambiae senso stricto. Acta Trop. 2019, 198, 105033. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, M.P.; Vimal, D.; Kumar, S.; Jha, R.R.; Chowdhuri, D.K. Benzene induced resistance in exposed Drosophila melanogaster: Outcome of improved detoxification and gene modulation. Chemosphere 2018, 201, 144–158. [Google Scholar] [PubMed]
- Daly, A.K.; Fairbrother, K.S.; Smart, J. Recent advances in understanding the molecular basis of polymorphisms in genes encoding cytochrome P450 enzymes. Toxicol. Lett. 1998, 102–103, 143–147. [Google Scholar]
- Steele, L.D.; Muir, W.M.; Seong, K.M.; Valero, M.C.; Rangesa, M.; Sun, W.; Clark, J.M.; Coates, B.; Pittendrigh, B.R. Genome-wide sequencing and an open reading frame analysis of dichlorodiphenyltrichloroethane (DDT) susceptible (91-C) and resistant (91-R) Drosophila melanogaster laboratory populations. PLoS ONE 2014, 9, e98584. [Google Scholar] [CrossRef] [Green Version]
- Mukhtorova, D.; Hlava, J.; Száková, J.; Kubík, S.; Vrabec, V.; Tlustoš, P. Risk element accumulation in Coleoptera and Hymenoptera (Formicidae) living in an extremely contaminated area—A preliminary study. Environ. Monit. Assess. 2019, 191, 32. [Google Scholar] [CrossRef] [PubMed]
- Talarico, F.; Brandmayr, P.; Giulianini, P.G.; Ietto, F.; Naccarato, A.; Perrotta, E.; Tagarelli, A.; Giglio, A. Effects of metal pollution on survival and physiological responses in Carabus (Chaetocarabus) lefebvrei (Coleoptera, Carabidae). Eur. J. Soil Biol. 2014, 61, 80–89. [Google Scholar] [CrossRef]
- Ballan-Dufrançais, C. Localization of metals in cells of pterygote insects. Microsc. Res. Tech. 2002, 56, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Fedorka, K.M.; Copeland, E.K.; Winterhalter, W.E. Seasonality influences cuticle melanization and immune defense in a cricket: Support for a temperature-dependent immune investment hypothesis in insects. J. Exp. Biol. 2013, 216, 4005–4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eeva, T.; Sorvari, J.; Koivunen, V. Effects of heavy metal pollution on red wood ant (Formica s. str.) populations. Environ. Pollut. 2004, 132, 533–539. [Google Scholar]
- Sorvari, J.; Rantala, L.M.; Rantala, M.J.; Hakkarainen, H.; Eeva, T. Heavy metal pollution disturbs immunity: Toxic effects in wild ant populations. Environ. Pollut. 2007, 145, 324–328. [Google Scholar] [PubMed]
- Belskaya, E.; Gilev, A.; Belskii, E. Ant (Hymenoptera, Formicidae) diversity along a pollution gradient near the Middle Ural Copper Smelter, Russia. Environ. Sci. Pollut. Res. 2017, 24, 10768–10777. [Google Scholar] [CrossRef]
- Skaldina, O.; Ciszek, R.; Peräniemi, S.; Kolehmainen, M.; Sorvari, J. Facing the threat: Common yellowjacket wasps as indicators of heavy metal pollution. Environ. Sci. Pollut. Res. 2020, 27, 29031–29042. [Google Scholar] [CrossRef] [PubMed]
- Skaldina, O.; Sorvari, J. Not simply red: Colouration of red wood ant Formica rufa (Hymenoptera: Formicidae) is polymorphic, modular and size-dependent. Eur. J. Entomol. 2017, 114, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Görür, G. Effects of heavy metal accumulation in host plants to cabbage aphid (Brevicoryne brassicae)–morphology. Ekol. Bratisl. 2006, 25, 314–321. [Google Scholar]
- Osman, W.; El-Samad, L.M.; Mokhamer, E.-H.; El-Touhamy, A.; Shonouda, M. Ecological, morphological and histological studies on Blaps polycresta (Coleoptera: Tenebrionidae) as biomonitors of cadmium soil pollution. Environ. Sci. Pollut. Res. 2015, 22, 14104–14115. [Google Scholar] [CrossRef]
- Kraus, J.M.; Schmidt, T.S.; Walters, D.M.; Wanty, R.B.; Zuellig, R.E.; Wolf, R.E. Cross-ecosystem impacts of stream pollution reduce resource and contaminant flux to riparian food webs. Ecol. Appl. 2014, 24, 235–243. [Google Scholar] [CrossRef]
- Stanković, J.; Milošević, D.; Savić-Zdraković, D.; Yalçın, G.; Yildiz, D.; Beklioğlu, M.; Jovanović, B. Exposure to a microplastic mixture is altering the life traits and is causing deformities in the non-biting midge Chironomus riparius Meigen (1804). Environ. Pollut. 2020, 262, 114248. [Google Scholar] [CrossRef]
- Kiikkilä, O. Heavy-metal pollution and remediation of forest soil around the Harjavalta Cu-Ni smelter, in SW Finland. Silva Fenn. 2003, 37, 497. [Google Scholar] [CrossRef] [Green Version]
- Sorvari, J. Social wasp (Hymenoptera: Vespidae) beer trapping in Finland 2008-2012: A German surprise. Entomol. Fenn. 2013, 24, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Kaisai, I.; Hunt, J.H. Food quantity affect traits of offspring in the paper wasp Polistes metricus (Hymenoptera: Vespidae). Environ. Entomol. 2002, 31, 99–106. [Google Scholar]
- Badejo, O.; Leskinen, J.T.T.; Koistinen, A.; Sorvari, J. Urban environment and climate condition-related phenotypic plasticity of the common wasp Vespula vulgaris. Bull Insectol 2020, 73, 285–294. [Google Scholar]
- Clapperton, B.K.; Lo, P.L.; Moller, H.; Sandlant, G.R. Variation in colour markings of German wasps Vespula germanica (F.) and common wasps Vespula vulgaris (L.) (Hymenoptera: Vespidae) in New Zealand. N. Zeal. J. Zool. 1989, 16, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Rako, L.; Anderson, A.R.; Sgrò, C.M.; Stocker, A.J.; Hoffmann, A.A. The association between inversion In(3R)Payne and clinally varying traits in Drosophila melanogaster. Genetica 2006, 128, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Schielzeth, H.; Dieker, P. The green-brown polymorphism of the club-legged grasshopper Gomphocerus sibiricus is heritable and appears genetically simple. BMC Evol. Biol. 2020, 20, 63. [Google Scholar] [CrossRef]
- Noh, P.; Oh, S.; Park, S.; Kwon, T.; Kim, Y.; Choe, J.C.; Jeong, G. Association between host wing morphology polymorphism and Wolbachia infection in Vollenhovia emeryi (Hymenoptera: Myrmicinae). Ecol. Evol. 2020, 10, 8827–8837. [Google Scholar] [CrossRef]
- Roff, D.A. The evolution of wing dimorphism in insects. Evolution 1986, 40, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- Zera, A.J.; Sall, J.W.; Grudzinski, K. Flight-muscle polymorphism in the cricket Gryllus firmus: Muscle characteristics and their influence on the evolution of flightlessness. Physiol. Zool. 1997, 70, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Busso, J.P.; Blankckenhorn, W.U.; Gonzáles-Tokman, D. Healthier or bigger? Trade-off mediating male dimorphism in the black scavenger fly Sepsis thoracica (Diptera: Sepsidae). Ecol. Entomol. 2017, 42, 517–525. [Google Scholar] [CrossRef]
- Butovsky, R.O. Heavy metals in carabids (Coleoptera, Carabidae). ZooKeys 2011, 100, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Mielczarek, A.; Wojciechowicz-Żytko, E. Bioaccumulation of heavy metals (Zn, Pb, Cd) in Polistes nimphus (Christ, 1791) (Hymenoptera, Vespidae) living on contaminated sites. Pol. J. Environ. Stud. 2020, 29, 4249–4256. [Google Scholar] [CrossRef]
- Urbini, A.; Sparvoli, E.; Turillazzi, S. Social paper wasps as bioindicators: A preliminary research with Polistes dominulus (Hymenoptera Vespidae) as a trace metal accumulator. Chemosphere 2006, 64, 697. [Google Scholar] [CrossRef] [PubMed]
- Andreani, G.; Ferlizza, E.; Cabbri, R.; Bellei, E.; Isani, G. Essential (Mg, Fe, Zn and Cu) and non-essential (Cd and Pb) elements in predatory insects (Vespa crabro and Vespa velutina): A molecular perspective. Int. J. Mol. Sci. 2021, 22, 228. [Google Scholar] [CrossRef]
- Kheirallah, D.A.M.; El-Samad, L.M. Histological and ultrastructure alterations in the midgut of Blaps polycresta and Trachyderma hispida (coleoptera: Tenebrionidae) induced by heavy metals pollution. Asian J. Biol. Sci. 2019, 12, 637–647. [Google Scholar] [CrossRef] [Green Version]
- Polidori, C.; Pastor, A.; Jorge, A.; Pertusa, J. Ultrastructural alterations of midgut epithelium, but not greater wing fluctuating asymmetry, in paper wasps (Polistes dominula) from urban environments. Microsc. Microanal. 2018, 24, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.; Reynoldson, T.B. Bioaccumulation and trophic transfer. In The pollution biology of aquatic Oligochaetes; Rodriguez, P., Reynoldson, T.B., Eds.; Springer: Dordrecht, The Netherlands, 2011. [Google Scholar] [CrossRef]
- Naslund, L.C.; Gerson, J.R.; Brooks, A.C.; Walters, D.M.; Bernhardt, E.S. Contaminant subsidies to riparian food webs in appalachian streams impacted by mountaintop removal coal mining. Environ. Sci. Technol. 2020, 54, 3951–3959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badejo, O.; Skaldina, O.; Gilev, A.; Sorvari, J. Benefits of insect colours: A review from social insect studies. Oecologia 2020, 194, 27–40. [Google Scholar] [CrossRef]
- Badejo, O.; Skaldina, O.; Sorvari, J. Spatial and temporal variation in thermal melanism in the aposematic common wasp (Vespula vulgaris) in northern Europe. Ann. Zool. Fenn. 2018, 55, 67–78. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polluted Zone (N = 62) | Intermediate Zone (N = 36) | Reference Zone (N = 52) | Test Result | |
|---|---|---|---|---|
| Fe | 233.18 ± 1.20 | 202.63 ± 1.19 | 186.77 ± 1.10 | F2,6.75 = 2.85, p = 0.13 |
| Co | 1.41 ± 1.25 a | 0.51 ± 1.29 b | 0.39 ± 1.26 b | F2,88.9 = 35.39, p < 0.0001 |
| Ni | 11.25 ± 1.78 a | 9.21 ± 3.67 a | 3.43 ± 1.54 b | F2,3.72 = 9.61, p = 0.034 |
| Cu | 84.24 ± 1.23 a | 56.34 ± 1.25 b | 46.05 ± 1.13 b | F2,6.49 = 16.82, p = 0.0027 |
| Zn | 544.63 ± 1.61 | 352.45 ± 1.83 | 531.74 ± 1.35 | F2,6.54 = 1.52, p = 0.29 |
| As | 6.98 ± 1.45 a | 3.97 ± 1.73 a | 1.63 ± 1.33 b | F2,6.53 = 25.60, p = 0.0008 |
| Cd | 1.60 ± 1.29 a | 0.41 ± 1.28 b | 0.34 ± 1.22 b | F2,89.4 = 50.72, p < 0.0001 |
| Pb | 1.26 ± 1.81 a | 0.82 ± 2.42 a | 0.20 ± 1.60 b | F2,8.87 = 16.05, p = 0.0011 |
| Factor Pattern | ||
|---|---|---|
| PC1 | PC2 | |
| Fe | 0.615 | −0.221 |
| Co | 0.840 | 0.317 |
| Ni | 0.722 | −0.380 |
| Cu | 0.916 | −0.087 |
| Zn | 0.303 | 0.907 |
| As | 0.845 | −0.153 |
| Cd | 0.859 | 0.076 |
| Pb | 0.831 | 0.016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badejo, O.; Skaldina, O.; Peräniemi, S.; Carrasco-Navarro, V.; Sorvari, J. Phenotypic Plasticity of Common Wasps in an Industrially Polluted Environment in Southwestern Finland. Insects 2021, 12, 888. https://doi.org/10.3390/insects12100888
Badejo O, Skaldina O, Peräniemi S, Carrasco-Navarro V, Sorvari J. Phenotypic Plasticity of Common Wasps in an Industrially Polluted Environment in Southwestern Finland. Insects. 2021; 12(10):888. https://doi.org/10.3390/insects12100888
Chicago/Turabian StyleBadejo, Oluwatobi, Oksana Skaldina, Sirpa Peräniemi, Victor Carrasco-Navarro, and Jouni Sorvari. 2021. "Phenotypic Plasticity of Common Wasps in an Industrially Polluted Environment in Southwestern Finland" Insects 12, no. 10: 888. https://doi.org/10.3390/insects12100888