
Conservation of Non-Pest Whiteflies and Natural Enemies of the Cabbage Whitefly Aleyrodes proletella on Perennial Plants for Use in Non-Crop Habitats


Section Phytomedicine, Institute of Horticultural Production Systems, Leibniz Universität Hannover, Herrenhäuser Straße 2, 30419 Hannover, Germany
*
Author to whom correspondence should be addressed.
Insects 2021, 12(9), 774; https://doi.org/10.3390/insects12090774
Submission received: 30 June 2021
/
Revised: 16 August 2021
/
Accepted: 21 August 2021
/
Published: 29 August 2021
(This article belongs to the Special Issue Conservation Biological Control in IPM and Organic Systems)
Abstract
:Simple Summary
The cabbage whitefly Aleyrodes proletella is a major insect pest of many cabbage crops. Natural enemies, in particular Encarsia tricolor as well as different hoverfly larvae and spiders, do not decrease pest populations sufficiently. The objective of this study is to promote local natural enemy populations by permanently establishing a non-pest whitefly species, which is an alternative host and additional food source when A. proletella is scarce or even absent. Therefore, the perennial abundance of the non-pest honeysuckle whitefly Aleyrodes lonicerae and natural enemies on different plants were evaluated in the open field. Wood avens Geum urbanum was the best host plant for A. lonicerae in terms of reproduction and overwintering. Most E. tricolor and spiders were also found on this plant species. In the future, G. urbanum might be used in non-crop habitats to increase natural enemy abundances in the agricultural landscape and decrease damage caused by A. proletella on adjacent cabbage plants.
Abstract
Aleyrodes proletella causes severe economic damage to several Brassica crops. Its naturally occurring enemies often immigrate late in the season or appear in low numbers on cabbage. This field study aims to permanently increase the local abundance of A. proletella’s natural enemies by providing the non-pest whitefly Aleyrodes lonicerae as an alternative and overwintering host/prey. Therefore, the population dynamics of natural enemies on different perennial herbaceous plants pre-infested with A. lonicerae were determined at two field locations over two winter periods. Most A. lonicerae colonized (on average 166.22 puparia per m²) and overwintered (342.19 adults per m²) on wood avens Geum urbanum. Furthermore, the abundance of A. proletella main parasitoid Encarsia tricolor (28.50 parasitized puparia per m²) and spiders (12.13 per m²) was 3–74 times and 3–14 times higher, respectively, on G. urbanum compared to the other experimental plants. Conclusively, G. urbanum pre-infested with A. lonicerae permanently promoted natural enemies of A. proletella by serving as shelter, reproduction, and overwintering habitat. A potential implementation of G. urbanum in conservation biological control strategies (e.g., tailored flower strips, hedgerows) against A. proletella are discussed and suggestions for future research are given.
1. Introduction
Aleyrodes proletella Linnaeus (Hemiptera: Aleyrodidae) has become a major pest on Brassica crops in different parts of the world during the past decades [1,2,3,4,5]. Its parasitoid Encarsia tricolor Förster (Hymenoptera: Aphelinidae) is distributed over large parts of Europe up to Russia and Northern Africa and is regarded as the most important natural enemy of A. proletella in Central Europe next to hoverfly larvae, coccinellids, and spiders among others [5,6,7,8,9,10,11,12,13,14,15,16,17].
Nevertheless, the migration of E. tricolor from its overwintering sites into cabbage crops often occurs too late in the season and insufficiently in numbers to regulate A. proletella populations substantially [18,19]. In close distance to the crop, E. tricolor and also other natural enemies often lack appropriate shelter, overwintering, and reproduction habitats that permanently provide natural enemies with alternative hosts and prey even if A. proletella is absent. In Europe, at least 44 naturally occurring whitefly species may potentially serve as alternative hosts for E. tricolor next to A. proletella [11,16]. However, most of these whitefly species are usually present only in relatively low numbers in the field [17].
The abundance of alternative whitefly hosts and their host plants in the agricultural landscape might be enhanced temporarily for one growing season. An annual banker plant system comprising Trialeurodes vaporariorum Westwood (Hemiptera: Aleyrodidae) as alternative host and E. tricolor on Hokkaido squash was already successfully tested in the field [20]. The enhanced abundance of alternative whitefly hosts increased hoverfly larvae abundance by 62% and A. proletella parasitism rates by 50% on adjacent cabbage crops, which led to a decrease of A. proletella populations by up to 26%. However, T. vaporariorum does not survive Central European winters in the open field and therefore does not serve as an overwintering host for E. tricolor [18].
A more long-term option to increase the local abundance of natural enemies in the agricultural landscape is to enhance the number of overwintering hosts as part of a conservation biological control strategy. E. tricolor overwinters as immature developmental stages inside whitefly nymphs [17,18]. A. proletella (e.g., on winter crops like oilseed rape) or naturally occurring non-pest whitefly species may play a role [21,22,23]. An increase of the latter could be achieved by a targeted composition of non-crop areas like field margins, hedgerows, or flowering strips with evergreen perennial host plants of non-pest whiteflies [24]. Potential plant-whitefly combinations facilitating natural enemies of A. proletella in Central Europe might for instance be Lonicera spp. (Caprifoliaceae) and several herbaceous host plants for A. lonicerae, Fraxinus spp. (Oleaceae) for Siphoninus phillyreae Haliday (Hemiptera: Aleyrodidae) or Viburnum spp. (Adoxaceae) for Aleurotuba jelinekii Frauenfeld (Hemiptera: Aleyrodidae) [5,14,25,26].
In particular, the polyphagous and widespread in Europe A. lonicerae has been evaluated as a promising host for E. tricolor on different host plants under controlled conditions. Thus, the present study aimed to identify perennial herbaceous host plants of A. lonicerae for their suitability to permanently conserve and promote E. tricolor and other natural enemies of A. proletella in the field.
2. Materials and Methods
Same experiments were installed at two locations in Germany, i.e., Hannover (52°23′39.0″ N 9°42′18.2″ E) and Sarstedt (52°14′39.8″ N 9°49′13.5″ E) in August 2015. In Hannover, the mean temperature was 11.27°C, and the precipitation 562.6 mm during the experimental period. The mean temperature and precipitation in Sarstedt were 4.63°C and 476.13 mm, respectively.
2.1. Inoculation of Plants with Alternative Hosts
Experimental plants, i.e., European columbine Aquilegia vulgaris (Ranunculaceae), peach-leaved bellflower Campanula persicifolia (Campanulaceae), wild strawberry Fragaria vesca and wood avens Geum urbanum (both Rosaceae), were selected based on the survival, development, and reproduction of A. lonicerae, A. proletella and their common parasitoid E. tricolor in a previous laboratory study [20]. They were sown from untreated seeds and grown separated by species under four gauze tents (length: 3 m, width: 1.5 m, height: 2 m; 500 plants per tent) in a greenhouse. Plants were inoculated with A. lonicerae when one to three true leaves were fully expanded. Adult A. lonicerae derived from the main rearing established on Aegopodium podagraria (ground elder) at the Section Phytomedicine, Institute of Horticultural Production Systems, Leibniz Universität Hannover, Germany. For inoculation, 1200 adult A. lonicerae females (2.4 females per plant) and 300 males were evenly released under each gauze tent. Females were allowed to oviposit for 35 days before plants were transferred to the field. At planting in August 2015, 6.91 ± 1.20 puparia (mean ± SE) were present on each plant.
2.2. Experimental Set-Up
Experimental plots (1.6 m × 1.6 m) were arranged in line in a randomized block design with three blocks and five treatments, i.e., A. vulgaris, C. persicifolia, F. vesca, G. urbanum, and an equal mixture of all four plant species, per location. Plants (64 per plot) were planted with 20 cm distance to each other and plots were separated by 40 cm bare soil. Other naturally occurring vegetation in the plots was regularly removed if needed. All assessments were done on four randomly selected plants per plot from 16 December 2015 to 14 December 2016. Border plants were not assessed and one plant of each species was selected in the mixed treatment. Every four weeks, the number of whitefly puparia (last nymphal stage) and parasitized whitefly puparia as well as the number of other herbivores and predatory arthropods were counted per plant in the field (14 assessment dates in total). Additionally, three leaves per plot with unparasitized and parasitized puparia were sampled every four months (4 samplings in total), transferred to gauze bags, and incubated at room temperature in order to identify the emerging adult whiteflies and parasitoids, respectively, to species level. The number of overwintering adult whiteflies per plant was determined twice, i.e., in December of both years.
2.3. Statistics
Data were statistically analyzed with R version 4.0.5 for each location separately [27]. Plots and data for descriptive statistics were provided by the packages ‘car’ and ‘FSA’ (Fisheries Stock Analysis), respectively [28,29]. All assessed data per plant were calculated to the number of individuals per m² for each plot before statistical analysis. Differences between treatments in terms of the number of whitefly puparia and parasitized whitefly puparia, other herbivores, and predators were determined with generalized linear mixed-effects models (glmer) fit by maximum likelihood (package ‘lme4’) [30]. Count data were fitted with negative binomial models (glmer.nb) and a log link function to deal with overdispersion [31]. Linear mixed-effects models (lmer) fit by restricted maximum likelihood (REML) were applied to compare the treatments with each other regarding the number of overwintering adult A. lonicerae (package ‘lme4’) [30]. The package ‘blmeco’ determined dispersion parameters [32]. Treatment (A. vulgaris, C. persicifolia, F. vesca, G. urbanum, mix) and assessment date were set as explanatory variables. Data were repeatedly collected from the same plots. Thus, an identification number was assigned to each plot, which was taken as a random effect to account for temporal non-independence. The Akaike information criterion (AIC) was used for model evaluation, i.e., the model with the lowest AIC value (highest accuracy) was chosen for each response variable to compute an analysis of deviance table (ANOVA function). Tukey post hoc was applied for multiple comparisons of means to determine differences between treatments (package ‘multcomp’) [33].
3. Results
3.1. Herbivore Population Development
The amount of overwintering adult A. lonicerae was different on experimental plant species in Hannover (χ2 (4, N = 30) = 16.47, p = 0.003) as well as in Sarstedt (χ2 (4, N = 30) = 61.46, p < 0.001). G. urbanum served as the best overwintering host for A. lonicerae adults at both locations. In Hannover, the average number of overwintering A. lonicerae per m² over the two experimental winters was far higher on G. urbanum (380.21 ± 164.27) than on C. persicifolia (2.08 ± 1.32) (p < 0.001). There was no difference to the treatments A. vulgaris (233.33 ± 113.99), F. vesca (172.92 ± 82.74), and mix (245.83 ± 124.74) (p > 0.05). In Sarstedt, more adult A. lonicerae adults overwintered on G. urbanum (304.17 ± 65.60) than on F. vesca (184.38 ± 58.85), A. vulgaris (102.08 ± 46.85) and C. persicifolia (23.96 ± 14.29) (p = 0.02, p < 0.001 and p < 0.001, respectively). Furthermore, less A. lonicerae adults were observed on A. vulgaris and C. persicifolia compared to the mixed treatment (230.21 ± 66.78) (p = 0.01 and p < 0.001, respectively). F. vesca was a better overwintering host for A. lonicerae than C. persicifolia (p < 0.001). No other differences between plant species were detected (p > 0.05).
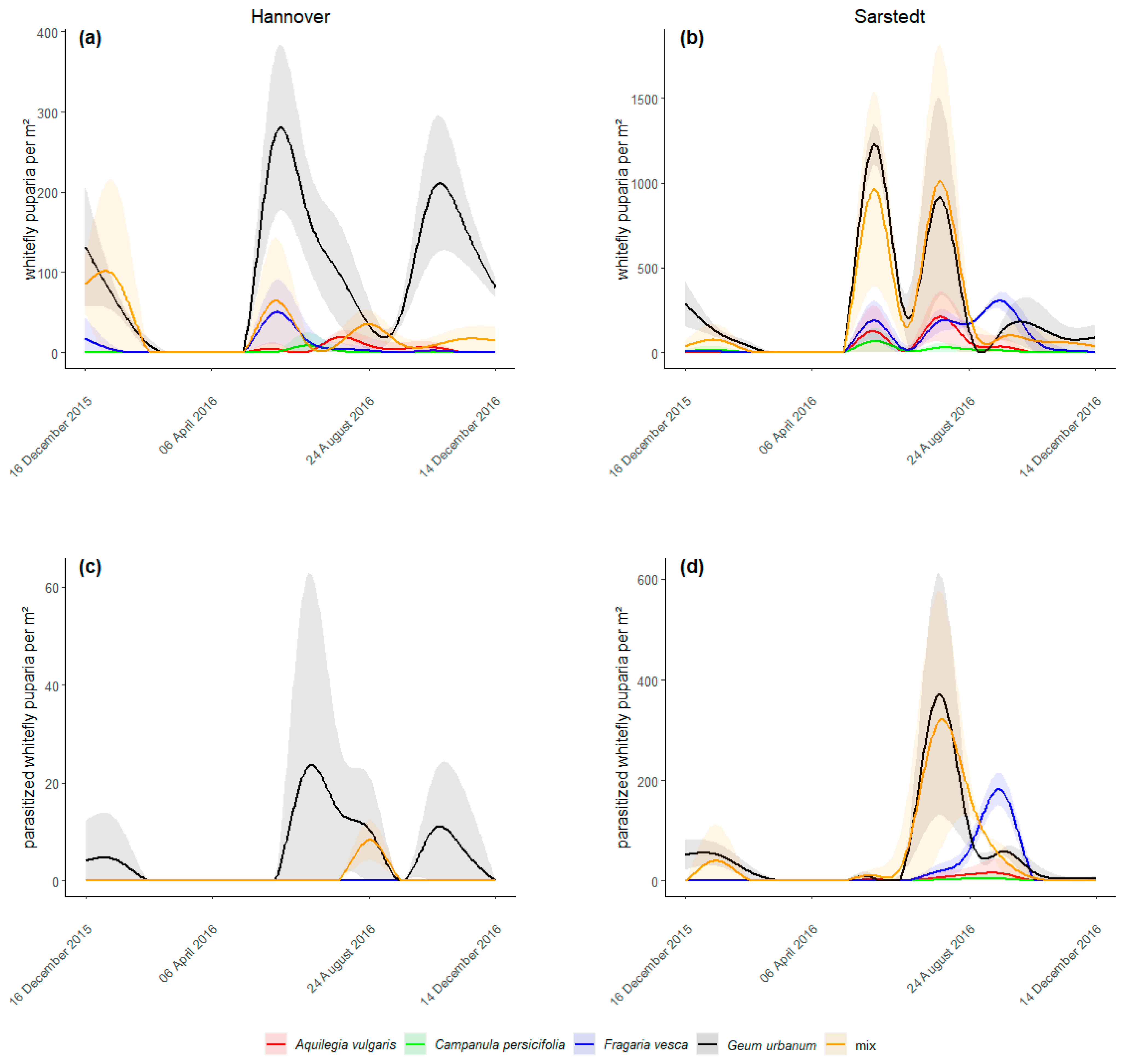
The abundance of A. lonicerae puparia differed between the treatments in Hannover (Figure 1a, χ2 (4, N = 210) = 186.08, p < 0.001) and in Sarstedt (Figure 1b, χ2 (4, N = 210) = 84.47, p < 0.001). The average number of puparia per m² over the entire experimental period ranged in Hannover from 0.74 ± 0.61 (C. persicifolia) to 91.52 ± 14.10 (G. urbanum) and in Sarstedt from 10.57 ± 3.83 (C. persicifolia) to 240.92 ± 58.57 (G. urbanum). At both locations, G. urbanum harbored more A. lonicerae puparia than any other plant species (all p < 0.001). More statistical differences are shown in Table 1.
No whitefly species other than A. lonicerae (e.g., A. proletella) was observed on any experimental plant at any assessment date. The numbers of other herbivores (i.e., aphids) were insufficient for statistical analysis.
3.2. Abundance of Naturally Occurring Enemies
All samples of parasitized whitefly puparia taken from the experimental plants were identified as E. tricolor. At both locations the abundance of parasitized puparia differed between treatments (Hannover: Figure 1c, χ2 (4, N = 210) = 37.06, p < 0.001; Sarstedt: Figure 1d, χ2 (4, N = 210) = 152.20, p < 0.001). In Hannover, parasitized puparia were only on G. urbanum (on average 5.21 ± 1.78 individuals per m²) and in the mixed treatment (0.60 ± 0.36), i.e., no whitefly parasitism on the other plant species was recorded. The number of parasitized puparia was higher on G. urbanum than in all other treatments (all p < 0.001). In Sarstedt, the average number of parasitized puparia per m² ranged from 0.74 ± 0.44 (C. persicifolia) to 51.79 ± 16.30 (G. urbanum). Statistical differences are given in Table 1.
Spider abundance was also different between the treatments in Hannover (χ2 (4, N = 210) = 53.02, p < 0.001) as well as in Sarstedt (χ2 (4, N = 210) = 23.30, p < 0.001) with G. urbanum inhabiting the most spiders at both locations. In Hannover, more spiders per m² were found on G. urbanum (on average 16.52 ± 3.81) and the mixed treatment (7.29 ± 1.40) compared to A. vulgaris (1.64 ± 0.71), F. vesca (1.34 ± 0.40) and C. persicifolia (1.19 ± 0.61) (all p < 0.01). No other differences were determined (p > 0.05). In Sarstedt, G. urbanum (7.74 ± 1.56), the mix treatment (4.32 ± 0.89) and C. persicifolia (2.98 ± 0.78) sheltered more spider per m² than F. vesca (0.89 ± 0.34; p < 0.001, p = 0.02 and p = 0.02, respectively). Additionally, G. urbanum had more spiders than A. vulgaris (1.94 ± 0.62; p = 0.04). The number of predators other than spiders (i.e., hoverfly larvae, ladybeetles, predatory bugs, lacewing larvae, predatory flies, and gall midge larvae) were insufficient and could not be analyzed statistically.
4. Discussion
This two-year field study evaluated perennial host plants of A. lonicerae for the purpose of the conservation biological control of A. proletella. Populations of alternative hosts/prey (A. lonicerae) and natural enemies (E. tricolor and spiders) of A. proletella established, overwintered, and permanently settled in the highest numbers on G. urbanum.
The parasitoid E. tricolor but also spiders are next to hoverfly larvae among the most important natural enemies of A. proletella [15]. An improvement of resilience of E. tricolor by annual banker plants has already been shown to increase parasitism rates and decrease A. proletella populations on cabbage [20]. More research is needed to investigate the impact of a permanent increase in local natural enemy abundance by G. urbanum in field margins on A. proletella populations on adjacent cabbage crops. Furthermore, it needs to be evaluated if pre-infestation with A. lonicerae (done in the current study) is necessary, or if the natural infestation can build up a suitable reservoir.
Hoverfly larvae might not have been affected significantly in this study, because hoverfly adults depend on floral resources from suitable flowers, which were hardly available [34,35]. Thus, a combination of G. urbanum with suitable flowering plants might lead to even better conservation of the natural enemies of A. proletella and should be addressed in future research.
Among all experimental plant species, G. urbanum increased A. lonicerae and natural enemy abundance at both experimental locations most, even in the second winter of the study. However, plants in Sarstedt inhabited more A. lonicerae and E. tricolor than the ones in Hannover. Soil analyses at both locations revealed that the soil in Hannover lacked nitrogen (N), potassium (K), and magnesium (Mg) resulting in generally larger and more vital plants in Sarstedt. Therefore, the plants in Sarstedt represented a qualitatively and quantitatively better nutritional source and thus may have benefited both, A. lonicerae and its parasitoid E. tricolor [36]. Next to plant nutrition, local differences in climatic parameters (e.g., on average 7°C warmer and in total 87 mm more precipitation in Hannover compared to Sarstedt during the entire experimental period), vegetation or additional food sources are further potential factors that could have influenced the abundances and colonization with herbivores and natural enemies at the two locations.
None of the investigated plant species served as a host plant for A. proletella. This whitefly pest might be able to survive and successfully reproduce to a certain extent, if caged onto these plants in the laboratory [20]. However, no adult A. proletella was observed on any plant species at any location and at any assessment date in the present field study, although cabbage plants with A. proletella populations were present in about 50 m distance to the experimental plots at both locations.
No A. lonicerae puparia and thus no parasitism was observed at either location from March to May 2016. Only whitefly adults that either overwintered or recently emerged from overwintered nymphs, first deposited eggs and already young nymphs (potential parasitism not visible in the field), and E. tricolor adults emerged from overwintered immature stages were present during this period [18]. Due to low spring temperatures, deposited whitefly eggs needed until 1st June before they developed into puparia (Figure 1a,b). First parasitized puparia appeared a few weeks later by the end of June (Figure 1c,d).
The number of E. tricolor pupae, i.e., dark whitefly puparia, during winter months was relatively low. This parameter was determined to estimate the abundance of whitefly parasitoids on the plants during the entire experimental period. However, this parameter actually underestimates the number of overwintering E. tricolor, since most E. tricolor overwinter as eggs or young larvae inside whitefly nymphs without showing morphological differences [18].
In conclusion, G. urbanum (pre-)infested with A. lonicerae is able to permanently increase the local abundance of natural enemies of A. proletella. Future research might focus on the added effect of G. urbanum in non-crop habitats (e.g., field margins, hedgerows, flowering strips) on the populations of natural enemies and A. proletella on adjacent cabbage crops and on cabbage yield. More general, similar conservation biological control strategies using alternative hosts to permanently promote natural enemy populations can be established targeting other important horticultural or agricultural pests. Finally, the conservation of non-pest herbivores in non-crop habitats might not only suppress mass outbreaks of pest populations and decrease insecticide applications but might also counteract biodiversity loss in agricultural landscapes.
Author Contributions
Conceptualization, S.L. and R.M.; Formal analysis, S.L.; Funding acquisition, R.M.; Investigation, S.L.; Methodology, S.L. and R.M.; Project administration, R.M.; Resources, R.M.; Supervision, R.M.; Validation, S.L.; Visualization, S.L.; Writing—original draft, S.L.; Writing—review & editing, S.L. and R.M. Both authors have read and agreed to the published version of the manuscript.
Funding
The project was supported by funds of the Federal Ministry of Food and Agriculture (BMEL) based on a decision of the Parliament of the Federal Republic of Germany via the Federal Office for Agriculture and Food (BLE) under the innovation support program (grant number 2812NA016).
Data Availability Statement
The data presented in this study are openly available in LUH-Projekt Seafile at https://doi.org/10.25835/0021354 (accessed on 16 August 2021) [37].
Acknowledgments
We thank Serafine Herrmann, Lisa Hildebrandt, Timo Michel, Birgit Milde, Johannes Specht and Florian Wulf for their support during data collection and plant cultivation.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Nebreda, M.; Nombela, G. Comparative host suitability of some Brassica cultivars for the whitefly, Aleyrodes proletella (Homoptera: Aleyrodidae). Environ. Entomol. 2005, 34, 205–209. [Google Scholar] [CrossRef]
- Trdan, S.; Modic, S.; Bobnar, A. The influence of cabbage whitefly (Aleyrodes proletella L., Aleyrodidae) abundance on the yield of Brussels sprouts. IOBC/WPRS Bull. 2003, 26, 265–270. [Google Scholar]
- Loomans, A.; Staneva, I.; Huang, Y.; Bukovinszkiné-Kiss, G.; van Lenteren, J.C. When native non-target species go indoors: A new challenge to biocontrol of whiteflies in European greenhouses. IOBC/WPRS Bull. 2002, 25, 139–142. [Google Scholar]
- de Barro, P.J.; CARVER, M. Cabbage whitefly, Aleyrodes proletella (L.) (Hemiptera: Aleyrodidae), newly discovered in Australia. Aust. J. Entomol. 1997, 36, 255–256. [Google Scholar] [CrossRef]
- Evans, G.A. The Whiteflies (Hemiptera: Aleyrodidae) of the World and Their Host Plants and Natural Enemies. Version: 070606. 2007. Available online: https://keys.lucidcentral.org/keys/v3/whitefly/PDF_PwP ETC/world-whitefly-catalog-Evans.pdf (accessed on 30 June 2021).
- Springate, S. The cabbage whitefly Aleyrodes proletella and its natural enemies on wild cabbage Brassica oleracea on the Kent coast. Trans. Kent Field Club 2017, 20, 42–58. [Google Scholar]
- Pütz, A.; Klausnitzer, B.; Schwartz, A.; Gebert, J. Der Bogen-Zwergmarienkäfer Clitostethus arcuatus (Rossi, 1794)—eine mediterrane Art auf Expansionskurs (Col., Coccinellidae). Entomol. Nachr. Ber. 2000, 44, 193–197. [Google Scholar]
- Springate, S.; Arnold, S.E.J. New vice-county records of Clitostethus arcuatus (Rossi) (Coleoptera: Coccinellidae) and new association with wild cabbage. Br. J. Entomol. Nat. Hist. 2011, 24, 224–225. [Google Scholar] [CrossRef] [Green Version]
- van Rijn, P.C.J.; den Belder, E.; Elderson, J.; Vlaswinkel, M.; van Alebeek, F. Perspectives for functional agro biodiversity in Brussels sprouts. IOBC/WPRS Bull. 2008, 34, 121–124. [Google Scholar]
- Stein, E. Untersuchungen über Biologie, Massenwechsel und Bekämpfung der Kohlmottenschildlaus, Aleurodes proletella L. PhD Thesis, Rheinische Friedrich Wilhelms-Universität, Bonn, Germany, 1958. [Google Scholar]
- Noyes, J.S. Universal Chalcidoidea Database. 2021. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 30 June 2021).
- Gumovsky, A. Parasitoids (Hymenoptera: Eulophidae, Aphelinidae) of the cabbage whitefly, Aleyrodes proletella (Hemiptera: Aleyrodidae), associated with the greater celindine (Chelidonium majus). In Second International Symposium on Biological Control of Arthropods; Forest Service: Davos, Switzerland, 2005; p. 108. [Google Scholar]
- Butler, C.G. The occurence of the chalcids Encarsia partenopea Masi and E. tricolor Förster in England (Hymenoptera). Proc. R. Entomol. Soc. Lond. 1936, 11, 79–80. [Google Scholar]
- Mound, L.A.; Halsey, S.H. Whitefly of the world; British Museum (Natural History): London, UK, 1978. [Google Scholar]
- Laurenz, S.; Schmidt, S.; Balkenhol, B.; Meyhöfer, R. Natural enemies associated with the cabbage whitefly Aleyrodes proletella in Germany. J. Plant Dis. Prot. 2019, 126, 47–54. [Google Scholar] [CrossRef]
- Evans, G.A. Parasitoids (Hymenoptera) Associated with Whiteflies (Aleyrodidae) of the World. Version 070202; 2007. Available online: http://www.sel.barc.usda.gov:8080/1WF/parasitoidcatalog.pdf (accessed on 30 June 2021).
- Bährmann, R. Die Mottenschildläuse: Aleyrodina, 1st ed.; Westarp Wissenschaften: Hohenwarsleben, Germany, 2002. [Google Scholar]
- Laurenz, S.; Brun, A.; Meyhöfer, R. Overwintering of Encarsia tricolor on the cabbage whitefly. IOBC-WPRS Bull. 2017, 122, 156–159. [Google Scholar]
- Springate, S. The Cabbage Whitefly, Aleyrodes Proletella: Causes of Outbreaks and Potential Solutions. Ph.D. Thesis, University of Greenwich, London, UK, 2016. [Google Scholar]
- Laurenz, S.; Meyhöfer, R. Banker plants promote functional biodiversity and decrease populations of the cabbage whitefly Aleyrodes proletella. J. Appl. Entomol. 2021, 145, 36–45. [Google Scholar] [CrossRef]
- Ludwig, M.; Ludwig, H.; Conrad, C.; Dahms, T.; Meyhöfer, R. Cabbage whiteflies colonise Brassica vegetables primarily from distant, upwind source habitats. Entomol. Exp. Appl. 2019, 167, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M.; Meyhöfer, R. Efficacy of crop cover netting against cabbage pests and their natural enemies and relevance of oilseed rape. J. Plant Dis. Prot. 2016, 123, 331–338. [Google Scholar] [CrossRef]
- Ludwig, M.; Schlinkert, H.; Meyhöfer, R. Wind-modulated landscape effects on colonization of Brussels sprouts by insect pests and their syrphid antagonists. Agric. For. Entomol. 2018, 20, 141–149. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat management to suppress pest populations: Progress and prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Pickett, C.H.; Wall, R. Biological control of ash whitefly Siphoninus phillyreae (Haliday) (Homoptera: Aleyrodidae) by Encarsia inaron (Walker) (Hymenoptera: Aphelinidae) in Northern California: 1990–2000. Pan-Pac. Entomol. 2003, 79, 156–158. [Google Scholar]
- Huldén, L. The whiteflies (Homoptera, Aleyrodidae) and their parasitoids in Finland. Not. Entomol. 1986, 66, 1–40. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Autria, 2021. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Ogle, D.H.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis. R Package Version 0.8.25. 2019. Available online: https://github.com/droglenc/FSA (accessed on 16 August 2021).
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hilbe, J.M. Negative Binomial Regression, 2nd ed.; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Korner-Nievergelt, F.; Roth, T.; von Felten, S.; Guélat, J.; Almasi, B.; Korner-Nievergelt, P. Bayesian Data Analysis in Ecology Using Linear Models with R.; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurenz, S.; Meyhöfer, R. Phenology and flower visitors of selected plant species with special respect to predators of the cabbage whitefly. IOBC-WPRS Bull. 2016, 118, 22–29. [Google Scholar]
- van Rijn, P.C.; Kooijman, J.; Wäckers, F.L. The contribution of floral resources and honeydew to the performance of predatory hoverflies (Diptera: Syrphidae). Biol. Control 2013, 67, 32–38. [Google Scholar] [CrossRef]
- Pekas, A.; Wäckers, F.L. Bottom-up effects on tri-trophic interactions: Plant fertilization enhances the fitness of a primary parasitoid mediated by its herbivore host. J. Econ. Entomol. 2020, 113, 2619–2626. [Google Scholar] [CrossRef] [PubMed]
- Laurenz, S.; Meyhöfer, R. Dataset: Conservation of Non-Pest Whiteflies and Natural Enemies of the Cabbage Whitefly Aleyrodes proletella on Perennial Plants for Use in Non-Crop Habitats. 2021. Available online: https://data.uni-hannover.de/dataset/de12a3c8-5718-4edd-b6db-f1bba771c8ff (accessed on 16 August 2021). [CrossRef]
Figure 1.
Population dynamics of whitefly puparia (a,b) and parasitized whitefly puparia (c,d) in Hannover and Sarstedt over the experimental period. The shaded areas represent the 95% confidence intervals.
Figure 1.
Population dynamics of whitefly puparia (a,b) and parasitized whitefly puparia (c,d) in Hannover and Sarstedt over the experimental period. The shaded areas represent the 95% confidence intervals.


{kind=link}
Table 1.
Mean numbers of non-parasitized and parasitized whitefly puparia per m² (mean ± SE) per plant species over the entire experimental period at the two locations (in brackets). Different small letters indicate statistical differences between plant species (α = 0.5).
Table 1.
Mean numbers of non-parasitized and parasitized whitefly puparia per m² (mean ± SE) per plant species over the entire experimental period at the two locations (in brackets). Different small letters indicate statistical differences between plant species (α = 0.5).
| Parameter (Location) | Mean Number of Individuals per m² over Experimental Period | ||||
|---|---|---|---|---|---|
| A. vulgaris | C. persicifolia | F. vesca | G. urbanum | mix | |
| whitefly puparia (Hannover) | 2.98 ± 0.98 b | 0.74 ± 0.61 a | 6.70 ± 2.62 b | 91.52 ± 14.10 d | 25.15 ± 6.56 c |
| whitefly puparia (Sarstedt) | 31.25 ± 11.76 a | 10.57 ± 3.83 a | 69.20 ± 15.95 b | 240.92 ± 58.67 c | 193.30 ± 59.45 bc |
| parasitized puparia (Hannover) | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 5.21 ± 1.78 b | 0.60 ± 0.36 a |
| parasitized puparia (Sarstedt) | 2.53 ± 1.28 ab | 0.74 ± 0.44 a | 19.64 ± 7.61 b | 51.79 ± 16.30 d | 46.58 ± 16.22 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Laurenz, S.; Meyhöfer, R. Conservation of Non-Pest Whiteflies and Natural Enemies of the Cabbage Whitefly Aleyrodes proletella on Perennial Plants for Use in Non-Crop Habitats. Insects 2021, 12, 774. https://doi.org/10.3390/insects12090774
AMA Style
Laurenz S, Meyhöfer R. Conservation of Non-Pest Whiteflies and Natural Enemies of the Cabbage Whitefly Aleyrodes proletella on Perennial Plants for Use in Non-Crop Habitats. Insects. 2021; 12(9):774. https://doi.org/10.3390/insects12090774
Chicago/Turabian StyleLaurenz, Sebastian, and Rainer Meyhöfer. 2021. "Conservation of Non-Pest Whiteflies and Natural Enemies of the Cabbage Whitefly Aleyrodes proletella on Perennial Plants for Use in Non-Crop Habitats" Insects 12, no. 9: 774. https://doi.org/10.3390/insects12090774
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.