Bacillus thuringiensis Bioinsecticides Induce Developmental Defects in Non-Target Drosophila melanogaster Larvae

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Stocks and Genetics
2.2. Bacterial Strains and Commercial Bioinsecticides
2.3. Spore Preparation
2.4. Crystal Purification
2.5. Diet Composition
2.6. Intestinal CFU Counting and Commensal Flora Estimation
2.7. Axenic Flies
2.8. Oral Infection of Larvae
2.9. Pupation Curve
2.10. Pupal Size
2.11. Larval Feeding Behavior and Food Intake
2.12. Midgut Permeability
2.13. Metabolic Assay
2.14. Lipid Labeling
2.15. Immunohistochemistry, Image Capture and Processing
2.16. Polyploidy Estimation
2.17. Statistics
3. Results
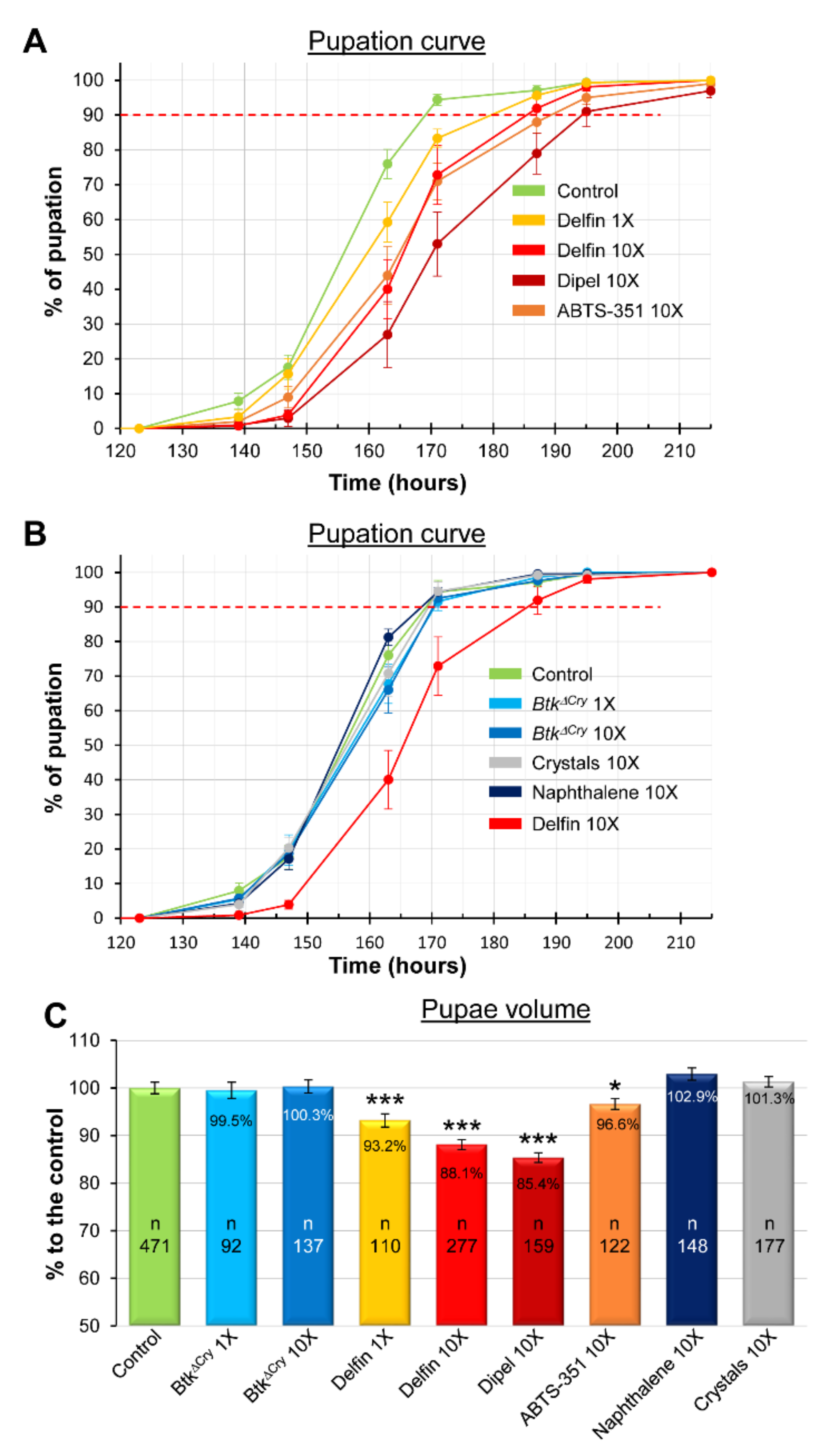
3.1. Btk Bioinsecticide Induces Developmental Delay and Growth Defects
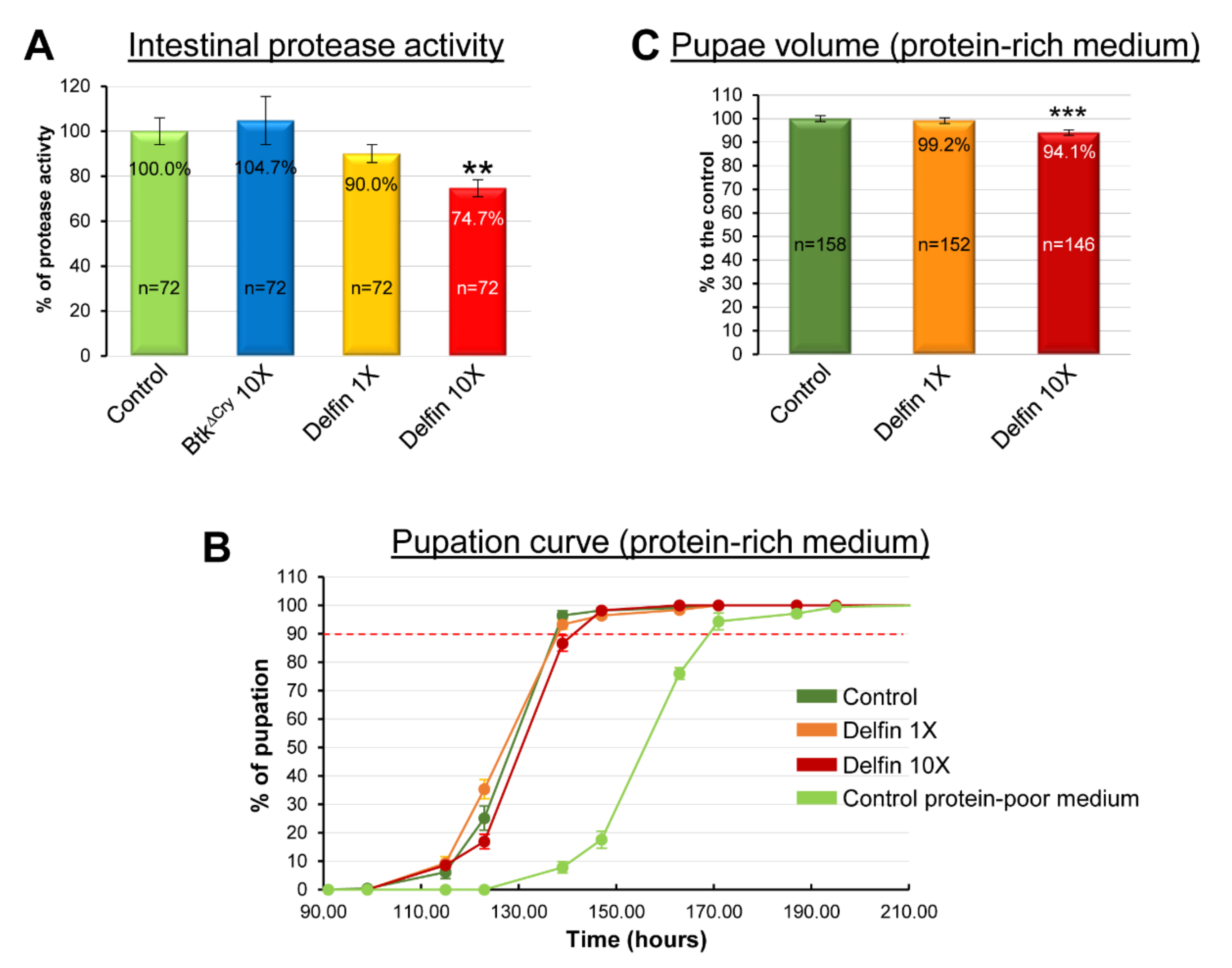
3.2. Btk Bioinsecticide Does Not Alter Feeding Behavior and Food Intake
3.3. Btk Bioinsecticide Impairs Intestinal Protein Metabolism
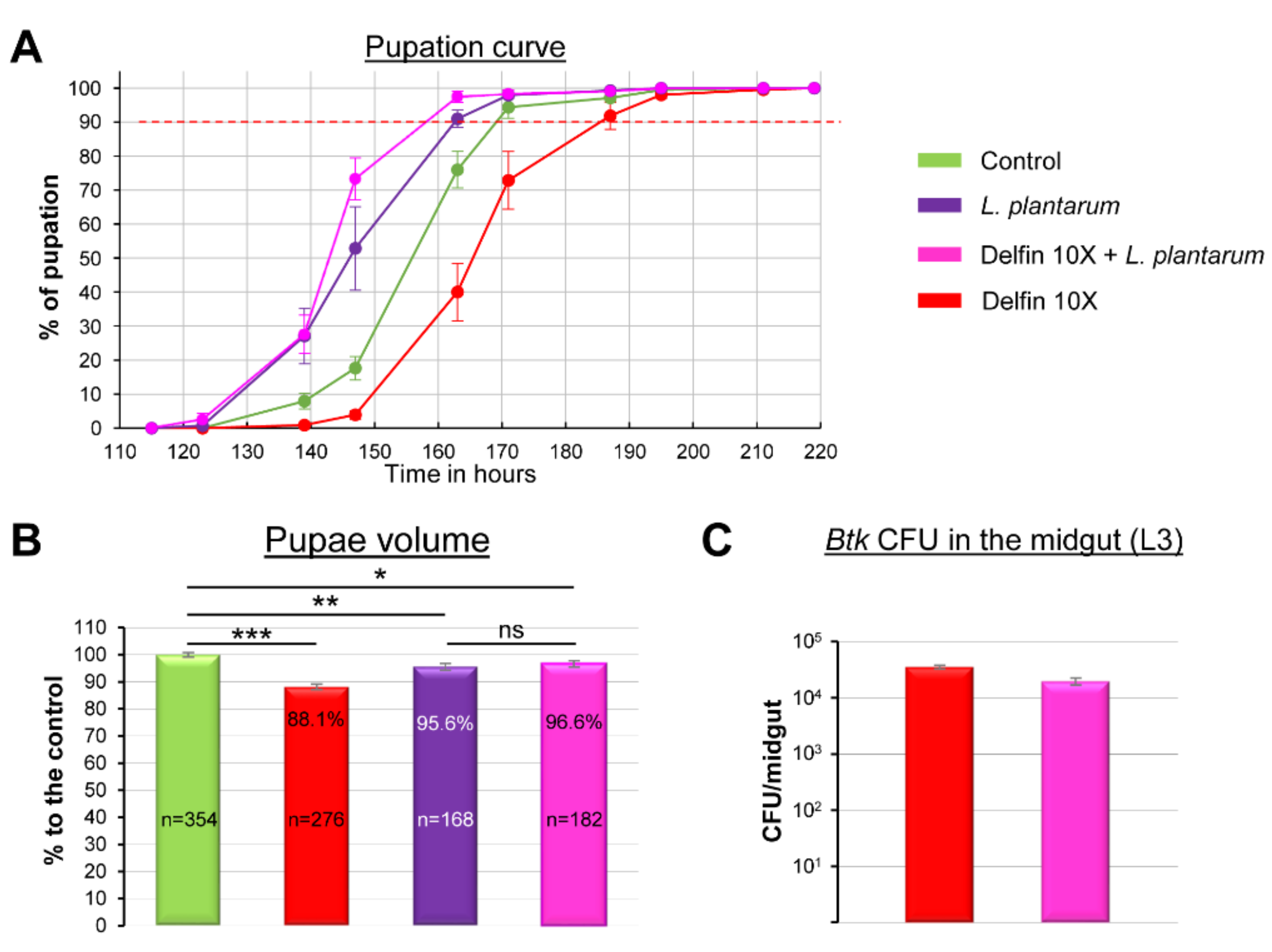
3.4. The Commensal Bacterium L. plantarum Helps to Overcome Btk Bioinsecticide Impacts
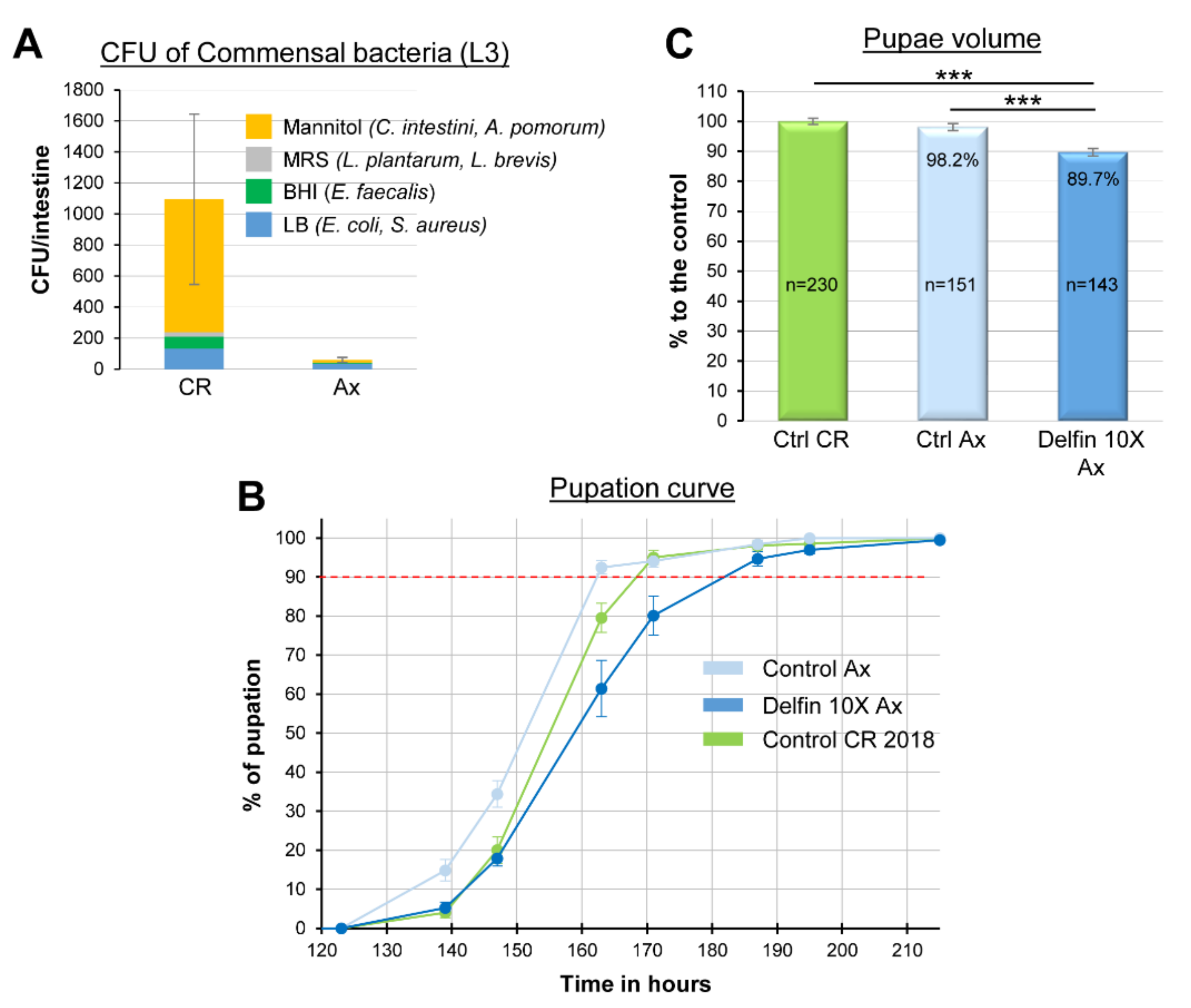
3.5. The Impacts of Btk Bioinsecticide Do Not Rely on Commensal Bacteria Disturbance
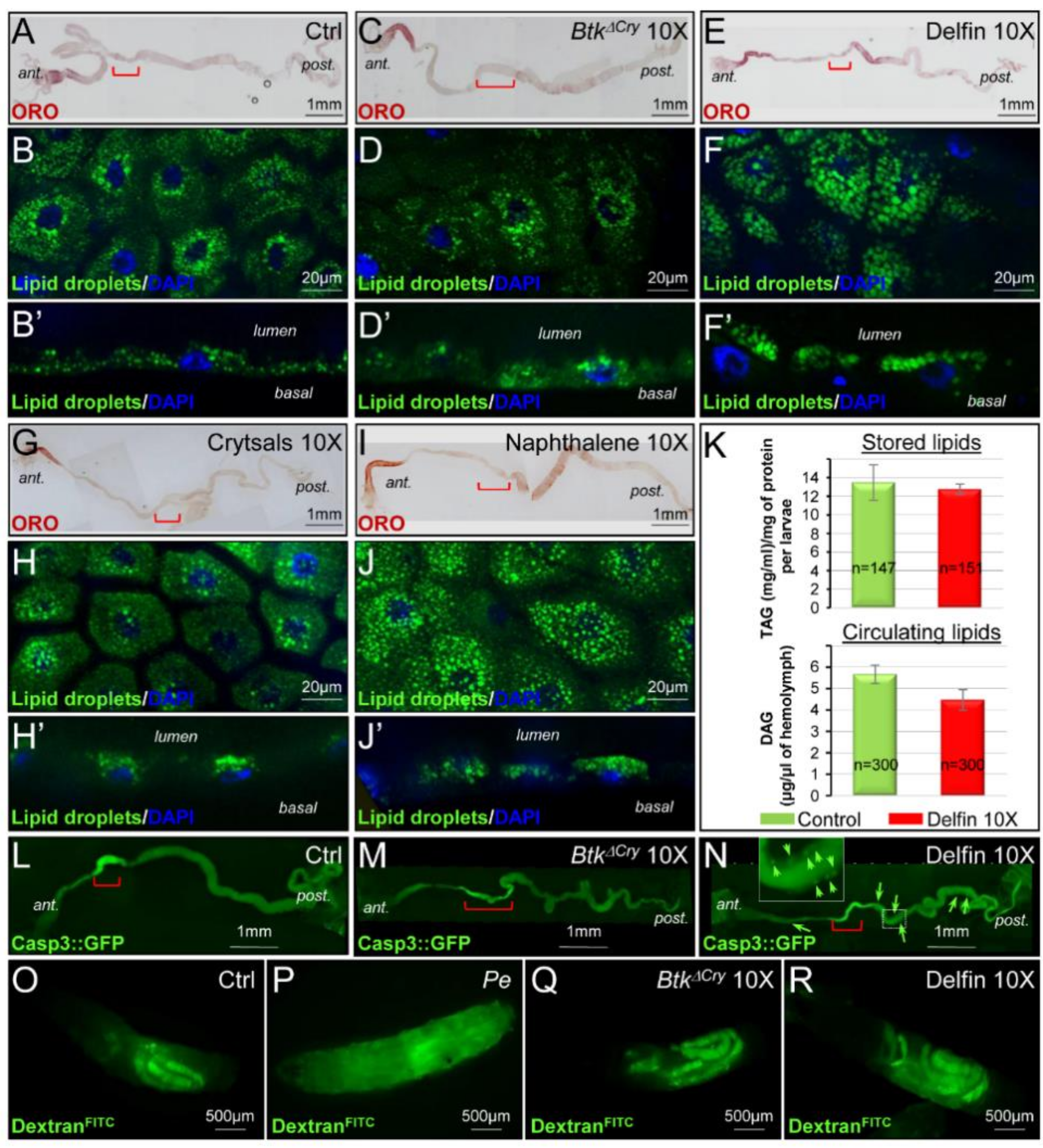
3.6. Btk Spores and Crystals Induce Midgut Perturbations
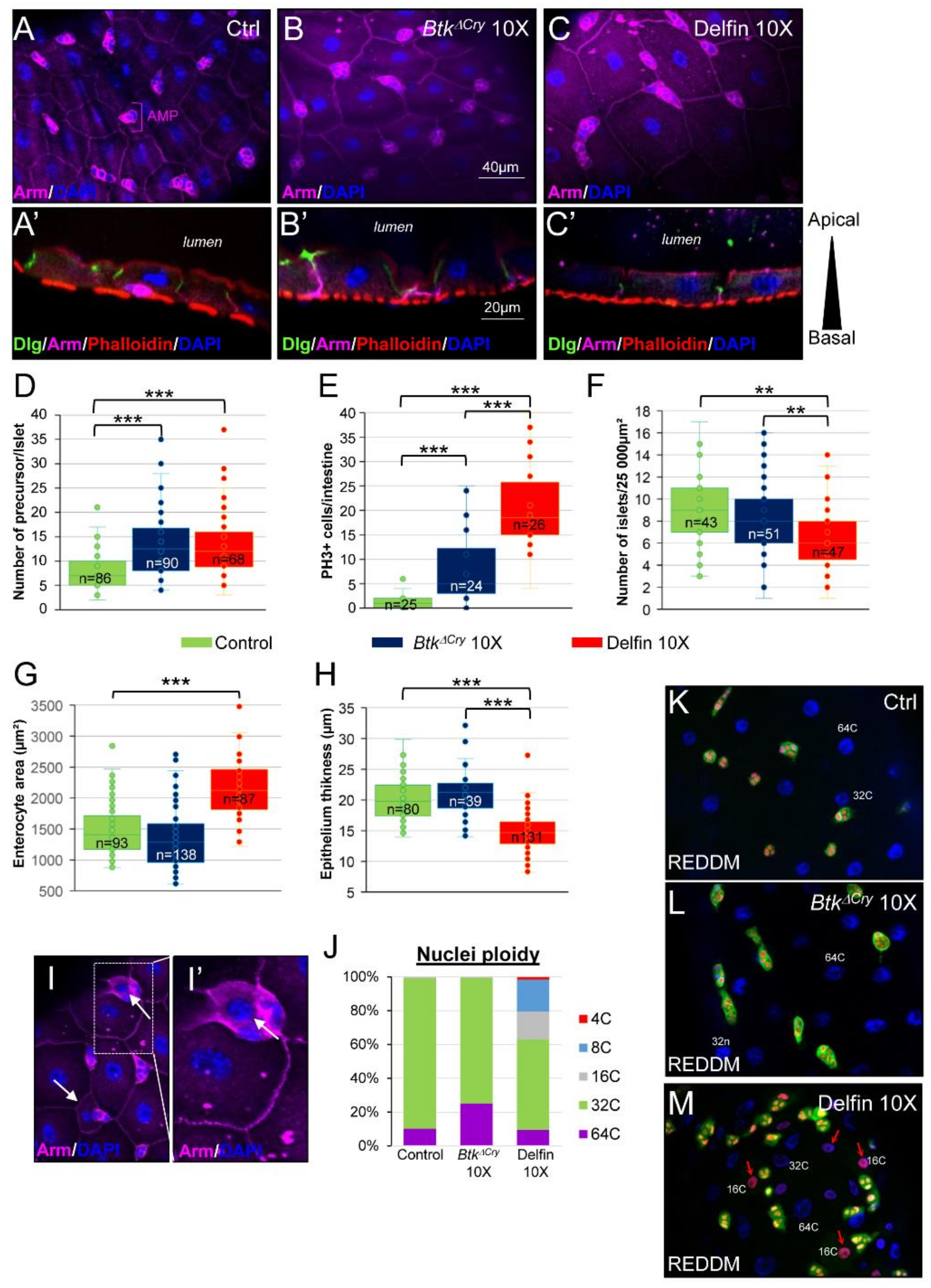
3.7. Enterocyte Flattening and Incomplete Differentiation of Adult Midgut Precursors Help to Maintain Intestinal Integrity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Frisvold, G.B. How low can you go? Estimating impacts of reduced pesticide use. Pest Manag. Sci. 2019, 75, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Casida, J.E.; Bryant, R.J. The ABCs of pesticide toxicology: Amounts, biology, and chemistry. Toxicol. Res. 2017, 6, 755–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiuza, L.M.; Polanczyk, R.A.; Crickmore, N. Bacillus thuringiensis and Lysinibacillus sphaericus; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Vilas-Bôas, G.; Peruca, A.; Arantes, O. Biology and taxonomy of Bacillus cereus, Bacillus anthracis, and Bacillus thuringiensis. Can. J. Microbiol. 2007, 53, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, L.; Vivoni, A.M.; Machado, V.; Knaak, N.; Berlitz, D.L.; Polanczyk, R.A.; Fiuza, L.M. Bacillus thuringiensis Characterization: Morphology, Physiology, Biochemistry, Pathotype, Cellular, and Molecular Aspects; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2017; pp. 1–18. [Google Scholar]
- Zeigler, D.R. The Bacillus Genetic Stock Center/Bacillus subtilis. Biol. Resour. Model Org. 2019, 556, 35–53. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2020, 107438. [Google Scholar] [CrossRef]
- Adang, M.J.; Crickmore, N.; Jurat-Fuentes, J.L. Chapter Two—Diversity of Bacillus thuringiensis Crystal Toxins and Mechanism of Action. Adv. Insect Physiol. 2014, 47, 39–87. [Google Scholar] [CrossRef]
- Van Frankenhuyzen, K. Specificity and Cross-Order Activity of Bacillus thuringiensis Pesticidal Proteins; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2017; pp. 127–172. [Google Scholar]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An international database for pesticide risk assessments and management. Hum. Ecol. Risk Assess. Int. J. 2016, 22, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis Toxins: An Overview of Their Biocidal Activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [Green Version]
- Caballero, J.; Jiménez-Moreno, N.; Orera, I.; Williams, T.; Fernández, A.B.; Villanueva, M.; Ferré, J.; Caballero, P.; Ancín-Azpilicueta, C. Unraveling the Composition of Insecticidal Crystal Proteins in Bacillus thuringiensis: A Proteomics Approach. Appl. Environ. Microbiol. 2020, 86, 00476-20. [Google Scholar] [CrossRef]
- Bizzarri, M.F.; Bishop, A.H. The Ecology of Bacillus thuringiensis on the Phylloplane: Colonization from Soil, Plasmid Transfer, and Interaction with Larvae of Pieris brassicae. Microb. Ecol. 2007, 56, 133–139. [Google Scholar] [CrossRef]
- Raymond, B.; Wyres, K.L.; Sheppard, S.K.; Ellis, R.J.; Bonsall, M.B. Environmental Factors Determining the Epidemiology and Population Genetic Structure of the Bacillus cereus Group in the Field. PLOS Pathog. 2010, 6, e1000905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettori, C.; Paffetti, D.; Saxena, D.; Stotzky, G.; Giannini, R. Persistence of toxins and cells of Bacillus thuringiensis subsp. kurstaki introduced in sprays to Sardinia soils. Soil Biol. Biochem. 2003, 35, 1635–1642. [Google Scholar] [CrossRef]
- Setlow, P. Spore Resistance Properties. Microbiol. Spectr. 2014, 2, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Infante, N.; Moreno-Fierros, L. An overview of the safety and biological effects of Bacillus thuringiensis Cry toxins in mammals. J. Appl. Toxicol. 2015, 36, 630–648. [Google Scholar] [CrossRef] [PubMed]
- Haller, S.; Romeis, J.; Meissle, M. Effects of purified or plant-produced Cry proteins on Drosophila melanogaster (Diptera: Drosophilidae) larvae. Sci. Rep. 2017, 7, 11172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosi, E.; Tank, J.L.; Royer, T.V.; Whiles, M.R.; Evans-White, M.; Chambers, C.; Griffiths, N.A.; Pokelsek, J.; Stephen, M.L. Toxins in transgenic crop byproducts may affect headwater stream ecosystems. Proc. Natl. Acad. Sci. USA 2007, 104, 16204–16208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tank, J.L.; Rosi, E.; Royer, T.V.; Whiles, M.R.; Griffiths, N.A.; Frauendorf, T.C.; Treering, D.J. Occurrence of maize detritus and a transgenic insecticidal protein (Cry1Ab) within the stream network of an agricultural landscape. Proc. Natl. Acad. Sci. USA 2010, 107, 17645–17650. [Google Scholar] [CrossRef] [Green Version]
- Amichot, M.; Curty, C.; Benguettat-Magliano, O.; Gallet, A.; Wajnberg, E. Side effects of Bacillus thuringiensis var. kurstaki on the hymenopterous parasitic wasp Trichogramma chilonis. Environ. Sci. Pollut. Res. 2015, 23, 3097–3103. [Google Scholar] [CrossRef]
- Babin, A.; Nawrot-Esposito, M.-P.; Gallet, A.; Gatti, J.-L.; Poirié, M. Differential side-effects of Bacillus thuringiensis bioinsecticide on non-target Drosophila flies. Sci. Rep. 2020, 10, 16241. [Google Scholar] [CrossRef]
- Grisolia, C.K.; Oliveira, R.; Domingues, I.; Oliveira-Filho, E.C.; Monerat, R.G.; Soares, A.M. Genotoxic evaluation of different delta-endotoxins from Bacillus thuringiensis on zebrafish adults and development in early life stages. Mutat. Res. 2009, 672, 119–123. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Houtz, P.L.; Buchon, N. Methods to Assess Intestinal Stem Cell Activity in Response to Microbes in Drosophila melanogaster. Adv. Struct. Saf. Stud. 2014, 1213, 171–182. [Google Scholar] [CrossRef]
- Troha, K.; Buchon, N. Methods for the study of innate immunity in Drosophila melanogaster. Wiley Interdiscip. Rev. Dev. Biol. 2019, 8, e344. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tennessen, J.M. Methods for studying the metabolic basis of Drosophila development. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e280. [Google Scholar] [CrossRef] [Green Version]
- Erkosar, B.; Storelli, G.; Mitchell, M.; Bozonnet, L.; Bozonnet, N.; Leulier, F. Pathogen Virulence Impedes Mutualist-Mediated Enhancement of Host Juvenile Growth via Inhibition of Protein Digestion. Cell Host Microbe 2015, 18, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Schott, S.; Ambrosini, A.; Barbaste, A.; Benassayag, C.; Gracia, M.; Proag, A.; Rayer, M.; Monier, B.; Suzanne, M. A fluorescent toolkit for spatiotemporal tracking of apoptotic cells in living Drosophila tissues. Development 2017, 144, 3840–3846. [Google Scholar] [CrossRef] [Green Version]
- A Antonello, Z.; Reiff, T.; Ballesta-Illan, E.; Dominguez, M. Robust intestinal homeostasis relies on cellular plasticity in enteroblasts mediated by miR-8–Escargot switch. EMBO J. 2015, 34, 2025–2041. [Google Scholar] [CrossRef]
- Ryu, J.-H.; Kim, S.-H.; Lee, H.-Y.; Bai, J.Y.; Nam, Y.-D.; Bae, J.-W.; Lee, D.G.; Shin, S.C.; Ha, E.-M.; Lee, W.J. Innate Immune Homeostasis by the Homeobox Gene Caudal and Commensal-Gut Mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Martino, M.E.; Bayjanov, J.R.; Joncour, P.; Hughes, S.; Gillet, B.; Kleerebezem, M.; Siezen, R.; Van Hijum, S.A.F.T.; Leulier, F. Resequencing of the Lactobacillus plantarum Strain WJL Genome. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Vodovar, N.; Vinals, M.; Liehl, P.; Basset, A.; Degrouard, J.; Spellman, P.; Boccard, F.; Lemaitre, B. Drosophila host defense after oral infection by an entomopathogenic Pseudomonas species. Proc. Natl. Acad. Sci. USA 2005, 102, 11414–11419. [Google Scholar] [CrossRef] [Green Version]
- Loudhaief, R.; Brun-Barale, A.; Benguettat, O.; Nawrot-Esposito, M.-P.; Pauron, D.; Amichot, M.; Gallet, A. Apoptosis restores cellular density by eliminating a physiologically or genetically induced excess of enterocytes in the Drosophila midgut. Development 2017, 144, 808–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, J.M.; Brown, B.J.; Carlton, B.C. Transfer of Bacillus thuringiensis plasmids coding for delta-endotoxin among strains of B. thuringiensis and B. cereus. Proc. Natl. Acad. Sci. USA 1982, 79, 6951–6955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delfin. Available online: https://www.certisusa.com/hubfs/4809084/Label%20SDS/pdf-labels/delfin-label.pdf (accessed on 1 September 2020).
- DiPel DF. Available online: https://s3-us-west-1.amazonaws.com/agrian-cg-fs1-production/pdfs/DiPelr_DF_Biological_Insecticide_Dry_Flowable_MSDS6.pdf (accessed on 1 September 2020).
- Dipel DF. Available online: https://www.edialux.fr/index.php?controller=attachment&id_attachment=2607 (accessed on 1 September 2020).
- Murty, M.; Srinivas, G.; Bora, R.S.; Sekar, V. A simple method for separation of the protein crystal from Bacillus thuringiensis using carboxymethyl cellulose column chromatography. J. Microbiol. Methods 1994, 19, 103–110. [Google Scholar] [CrossRef]
- Agaisse, H.; Lereclus, D. How does Bacillus thuringiensis produce so much insecticidal crystal protein? J. Bacteriol. 1995, 177, 6027–6032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monro, R.E. Protein turnover and the formation of protein inclusions during sporulation of Bacillus thuringiensis. Biochem. J. 1961, 81, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Bjordal, M.; Arquier, N.; Kniazeff, J.; Pin, J.P.; Léopold, P. Sensing of Amino Acids in a Dopaminergic Circuitry Promotes Rejection of an Incomplete Diet in Drosophila. Cell 2014, 156, 510–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrot-Esposito, M.-P.; Loudhaief, R.; Gallet, A. Protease Activity Assay in Fly Intestines. Bio-Protocol 2017, 7. [Google Scholar] [CrossRef]
- Tennessen, J.M.; Barry, W.E.; Cox, J.; Thummel, C.S. Methods for studying metabolism in Drosophila. Methods 2014, 68, 105–115. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Biological Hazards (BIOHAZ). Risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA J. 2016, 14. [Google Scholar] [CrossRef]
- AVIS. Available online: https://www.anses.fr/fr/system/files/DPR2013sa0039.pdf (accessed on 1 September 2020).
- Hung, T.P.; Van Truong, L.; Binh, N.D.; Frutos, R.; Quiquampoix, H.; Staunton, S. Fate of insecticidal Bacillus thuringiensis Cry protein in soil: Differences between purified toxin and biopesticide formulation. Pest Manag. Sci. 2016, 72, 2247–2253. [Google Scholar] [CrossRef]
- Hietakangas, V.; Cohen, S.M. Regulation of Tissue Growth through Nutrient Sensing. Annu. Rev. Genet. 2009, 43, 389–410. [Google Scholar] [CrossRef] [PubMed]
- Delanoue, R.; Slaidina, M.; Léopold, P. The Steroid Hormone Ecdysone Controls Systemic Growth by Repressing dMyc Function in Drosophila Fat Cells. Dev. Cell 2010, 18, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Stensmyr, M.C.; Dweck, H.K.; Farhan, A.; Ibba, I.; Strutz, A.; Mukunda, L.; Linz, J.; Grabe, V.; Steck, K.; Lavista-Llanos, S.; et al. A Conserved Dedicated Olfactory Circuit for Detecting Harmful Microbes in Drosophila. Cell 2012, 151, 1345–1357. [Google Scholar] [CrossRef] [Green Version]
- Surendran, S.; Hückesfeld, S.; Wäschle, B.; Pankratz, M.J. Pathogen-induced food evasion behavior in Drosophila larvae. J. Exp. Biol. 2017, 220, 1774–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keita, S.; Masuzzo, A.; Royet, J.; Kurz, C.L. Drosophila larvae food intake cessation following exposure to Erwinia contaminated media requires odor perception, Trpa1 channel and evf virulence factor. J. Insect Physiol. 2017, 99, 25–32. [Google Scholar] [CrossRef]
- Babin, A.; Kolly, S.; Schneider, F.; Dolivo, V.; Zini, M.; Kawecki, T.J. Fruit flies learn to avoid odours associated with virulent infection. Biol. Lett. 2014, 10. [Google Scholar] [CrossRef] [Green Version]
- Layalle, S.; Arquier, N.; Léopold, P. The TOR Pathway Couples Nutrition and Developmental Timing in Drosophila. Dev. Cell 2008, 15, 568–577. [Google Scholar] [CrossRef]
- Colombani, J.; Raisin, S.; Pantalacci, S.; Radimerski, T.; Montagne, J.; Léopold, P. A Nutrient Sensor Mechanism Controls Drosophila Growth. Cell 2003, 114, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Anagnostou, C.; Dorsch, M.; Rohlfs, M. Influence of dietary yeasts on Drosophila melanogaster life-history traits. Entomol. Exp. Appl. 2010, 136, 1–11. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Martins, N.E.; Balancé, L.F.; Broom, L.N.; Dias, A.J.; Fernandes, A.S.D.; Rodrigues, F.; Sucena, É.; Mirth, C.K. Drosophila melanogaster larvae make nutritional choices that minimize developmental time. J. Insect Physiol. 2015, 81, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Reis, T. Effects of Synthetic Diets Enriched in Specific Nutrients on Drosophila Development, Body Fat, and Lifespan. PLoS ONE 2016, 11, e0146758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Soultoukis, G.; Partridge, L. Dietary Protein, Metabolism, and Aging. Annu. Rev. Biochem. 2016, 85, 5–34. [Google Scholar] [CrossRef] [Green Version]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus plantarum Promotes Drosophila Systemic Growth by Modulating Hormonal Signals through TOR-Dependent Nutrient Sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Téfit, M.A.; Leulier, F. Lactobacillus plantarum favors the early emergence of fit and fertile adult Drosophila upon chronic undernutrition. J. Exp. Biol. 2017, 220, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, M.; Makki, K.; Storelli, G.; Machuca-Gayet, I.; Srutkova, D.; Hermanova, P.; Martino, M.E.; Balmand, S.; Hudcovic, T.; Heddi, A.; et al. Lactobacillus plantarum strain maintains growth of infant mice during chronic undernutrition. Science 2016, 351, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, P.; Parida, S.; Nanda, N.C.; Satpathy, R.; Pradhan, L.; Chandel, D.S.; Baccaglini, L.; Mohapatra, A.; Mohapatra, S.S.; Misra, P.R.; et al. A randomized synbiotic trial to prevent sepsis among infants in rural India. Nat. Cell Biol. 2017, 548, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial Communities of Diverse Drosophila Species: Ecological Context of a Host–Microbe Model System. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef]
- Broderick, N.A.; Lemaitre, B. Gut-associated microbes of Drosophila melanogaster. Gut Microbes 2012, 3, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Fast, D.; Duggal, A.; Foley, E. Monoassociation with Lactobacillus plantarum Disrupts Intestinal Homeostasis in Adult Drosophila melanogaster. mBio 2018, 9, e01114-18. [Google Scholar] [CrossRef] [Green Version]
- Téfit, M.A.; Gillet, B.; Joncour, P.; Hughes, S.; Leulier, F. Stable association of a Drosophila-derived microbiota with its animal partner and the nutritional environment throughout a fly population’s life cycle. J. Insect Physiol. 2018, 106, 2–12. [Google Scholar] [CrossRef]
- Palm, W.; Sampaio, J.L.; Brankatschk, M.; Carvalho, M.; Mahmoud, A.; Shevchenko, A.; Eaton, S. Lipoproteins in Drosophila melanogaster—Assembly, Function, and Influence on Tissue Lipid Composition. PLoS Genet. 2012, 8, e1002828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.-Z.; Lestradet, M.; Socha, C.; Schirmeier, S.; Schmitz, A.; Spenlé, C.; Lefebvre, O.; Keime, C.; Yamba, W.M.; Aoun, R.B.; et al. Enterocyte Purge and Rapid Recovery Is a Resilience Reaction of the Gut Epithelium to Pore-Forming Toxin Attack. Cell Host Microbe 2016, 20, 716–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apidianakis, Y.; Pitsouli, C.; Perrimon, N.; Rahme, L.G. Synergy between bacterial infection and genetic predisposition in intestinal dysplasia. Proc. Natl. Acad. Sci. USA 2009, 106, 20883–20888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Sayadian, A.-C.; Lowe, N.; Lovegrove, H.E.; Johnston, D.S. An alternative mode of epithelial polarity in the Drosophila midgut. PLoS Biol. 2018, 16, e3000041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, D.; Bost, A.; Driver, I.; Ohlstein, B. A Transient Niche Regulates the Specification of Drosophila Intestinal Stem Cells. Science 2010, 327, 210–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Edgar, B.A. EGFR signaling regulates the proliferation of Drosophila adult midgut progenitors. Development 2009, 136, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Micchelli, C.A.; Sudmeier, L.; Perrimon, N.; Tang, S.; Beehler-Evans, R. Identification of adult midgut precursors in Drosophila. Gene Expr. Patterns 2011, 11, 12–21. [Google Scholar] [CrossRef]
- Bonfini, A.; Liu, X.; Buchon, N. From pathogens to microbiota: How Drosophila intestinal stem cells react to gut microbes. Dev. Comp. Immunol. 2016, 64, 22–38. [Google Scholar] [CrossRef] [Green Version]
- Miguel-Aliaga, I.; Jasper, H.; Lemaitre, B. Anatomy and Physiology of the Digestive Tract of Drosophila melanogaster. Genetics 2018, 210, 357–396. [Google Scholar] [CrossRef] [Green Version]
- Bauce, E.; Carisey, N.; Dupont, A. Carry over effects of the entomopathogen Bacillus thuringiensis ssp. Kurstaki on Choristoneura fumiferana (Lepidoptera: Tortricidae) progeny under various stressful environmental conditions. Agric. For. Èntomol. 2006, 8, 63–76. [Google Scholar] [CrossRef]
- Sedaratian, A.; Fathipour, Y.; Talaei-Hassanloui, R.; Jurat-Fuentes, J. Fitness costs of sublethal exposure to Bacillus thuringiensis in Helicoverpa armigera: A carryover study on offspring. J. Appl. Èntomol. 2012, 137, 540–549. [Google Scholar] [CrossRef]
- Polanczyk, R.A.; Alves, S.B. Biological parameters of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) assayed with Bacillus thuringiensis berliner. Sci. Agric. 2005, 62, 464–468. [Google Scholar] [CrossRef]
- Houtz, P.; Bonfini, A.; Bing, X.; Buchon, N. Recruitment of Adult Precursor Cells Underlies Limited Repair of the Infected Larval Midgut in Drosophila. Cell Host Microbe 2019, 26, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Broderick, N.A.; Poidevin, M.; Pradervand, S.; Lemaitre, B. Drosophila Intestinal Response to Bacterial Infection: Activation of Host Defense and Stem Cell Proliferation. Cell Host Microbe 2009, 5, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S.; Liehl, P.; Buchon, N.; Lemaitre, B. Infection-Induced Host Translational Blockage Inhibits Immune Responses and Epithelial Renewal in the Drosophila Gut. Cell Host Microbe 2012, 12, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.C.N.; Vanhove, A.S.; Watnick, P.I. The interplay between intestinal bacteria and host metabolism in health and disease: Lessons from Drosophila melanogaster. Dis. Model. Mech. 2016, 9, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.P.; Cory, J.S.; Wilson, K.; Raubenheimer, D.; Simpson, S.J. Flexible diet choice offsets protein costs of pathogen resistance in a caterpillar. Proc. R. Soc. B Boil. Sci. 2005, 273, 823–829. [Google Scholar] [CrossRef] [Green Version]
- Shikano, I.; Cory, J.S. Dietary Mechanism behind the Costs Associated with Resistance to Bacillus thuringiensis in the Cabbage Looper, Trichoplusia ni. PLoS ONE 2014, 9, e105864. [Google Scholar] [CrossRef]
- Bing, X.; Gerlach, J.; Loeb, G.; Buchon, N. Nutrient-Dependent Impact of Microbes on Drosophila suzukii Development. mBio 2018, 9, e02199-17. [Google Scholar] [CrossRef] [Green Version]
- Ubeda, C.; Djukovic, A.; Isaac, S. Roles of the intestinal microbiota in pathogen protection. Clin. Transl. Immunol. 2017, 6, e128. [Google Scholar] [CrossRef]
- Van Arnam, E.B.; Currie, C.R.; Clardy, J. Defense contracts: Molecular protection in insect-microbe symbioses. Chem. Soc. Rev. 2018, 47, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.E.; Fischer, C.N.; Miles, J.; Handelsman, J. Frequent Replenishment Sustains the Beneficial Microbiome of Drosophila melanogaster. mBio 2013, 4, e00860-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, T.; Texada, M.J.; Halberg, K.A.; Rewitz, K.F. Metabolism and growth adaptation to environmental conditions in Drosophila. Cell. Mol. Life Sci. 2020, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Manière, G.; Alves, G.; Berthelot-Grosjean, M.; Grosjean, Y. Growth regulation by amino acid transporters in Drosophila larvae. Cell. Mol. Life Sci. 2020, 5–9. [Google Scholar] [CrossRef]
- Pasco, M.Y.; Loudhaief, R.; Gallet, A. The cellular homeostasis of the gut: What the Drosophila model points out. Histol. Histopathol. 2014, 30, 277–292. [Google Scholar]
- Joly, A.; Rousset, R. Tissue Adaptation to Environmental Cues by Symmetric and Asymmetric Division Modes of Intestinal Stem Cells. Int. J. Mol. Sci. 2020, 21, 6362. [Google Scholar] [CrossRef]
- Wells-Bennik, M.; Eijlander, R.T.; Besten, H.M.D.; Berendsen, E.M.; Warda, A.K.; Krawczyk, A.O.; Groot, M.N.N.; Xiao, Y.; Zwietering, M.H.; Kuipers, O.P.; et al. Bacterial Spores in Food: Survival, Emergence, and Outgrowth. Annu. Rev. Food Sci. Technol. 2016, 7, 457–482. [Google Scholar] [CrossRef]
- Edgar, B.A.; Zielke, N.; Gutierrez, C. Endocycles: A recurrent evolutionary innovation for post-mitotic cell growth. Nat. Rev. Mol. Cell Biol. 2014, 15, 197–210. [Google Scholar] [CrossRef]
- Xiang, J.; Bandura, J.; Zhang, P.; Jin, Y.; Reuter, H.; Edgar, B.A. EGFR-dependent TOR-independent endocycles support Drosophila gut epithelial regeneration. Nat. Commun. 2017, 8, 15125. [Google Scholar] [CrossRef] [Green Version]
- Castagnola, A.; Jurat-Fuentes, J.L. Intestinal regeneration as an insect resistance mechanism to entomopathogenic bacteria. Curr. Opin. Insect Sci. 2016, 15, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Dubovskiy, I.M.; Grizanova, E.V.; Whitten, M.M.A.; Mukherjee, K.; Greig, C.; Alikina, T.; Kabilov, M.; Vilcinskas, A.; Glupov, V.V.; Butt, T.M. Immuno-physiological adaptations confer wax moth Galleria mellonella resistance to Bacillus thuringiensis. Virulence 2016, 7, 860–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, B.M.D.C.E.; Martinez, L.C.; Barbosa, S.G.; Serrão, J.E.; Wilcken, C.F.; Soares, M.A.; Da Silva, A.A.; De Carvalho, A.G.; Zanuncio, J.C. Toxicity and cytopathology mediated by Bacillus thuringiensis in the midgut of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Sci. Rep. 2019, 9, 6667. [Google Scholar] [CrossRef] [PubMed]
- Cossentine, J.; Robertson, M.; Xu, D. Biological Activity of Bacillus thuringiensis in Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Èntomol. 2016, 109, 1071–1078. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % of Pupation | 10% | 50% | 90% | n |
|---|---|---|---|---|
| Control 2017 | 140.5 | 156.0 | 169.0 | 183 |
| Control 2018 | 142.0 | 155.0 | 168.5 | 211 |
| Control 2019 | 142.5 | 155.5 | 171.0 | 203 |
| Delfin 1X | 143.0 | 159.5 | 179.5 | 262 |
| Delfin 10X | 149.5 | 165.5 | 185.0 | 214 |
| Dipel 10X | 151.5 | 170.0 | 194.0 | 152 |
| ABTS-351 10X | 147.5 | 165.0 | 189.0 | 159 |
| BtkΔCry 1X | 141.5 | 157.0 | 170.5 | 263 |
| BtkΔCry 10X | 141.5 | 157.5 | 170.5 | 204 |
| Naphthalene sulfonic acid | 142.5 | 155.0 | 168.0 | 212 |
| Crystals | 142.0 | 156.5 | 169.5 | 143 |
| Control protein-rich medium | 117.0 | 128.5 | 137.5 | 212 |
| Delfin 1X protein-rich medium | 115.0 | 127.0 | 138.0 | 195 |
| Delfin 10X protein-rich medium | 117.0 | 130.5 | 141.0 | 207 |
| L. plantarum | 128.0 | 146.0 | 162.5 | 144 |
| Delfin 10X + L plantarum | 127.0 | 143.0 | 158.0 | 153 |
| Control Axenic | 134.0 | 151.5 | 162.5 | 171 |
| Delfin 10X Axenic | 142.0 | 159.0 | 182.0 | 172 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawrot-Esposito, M.-P.; Babin, A.; Pasco, M.; Poirié, M.; Gatti, J.-L.; Gallet, A. Bacillus thuringiensis Bioinsecticides Induce Developmental Defects in Non-Target Drosophila melanogaster Larvae. Insects 2020, 11, 697. https://doi.org/10.3390/insects11100697
Nawrot-Esposito M-P, Babin A, Pasco M, Poirié M, Gatti J-L, Gallet A. Bacillus thuringiensis Bioinsecticides Induce Developmental Defects in Non-Target Drosophila melanogaster Larvae. Insects. 2020; 11(10):697. https://doi.org/10.3390/insects11100697
Chicago/Turabian StyleNawrot-Esposito, Marie-Paule, Aurélie Babin, Matthieu Pasco, Marylène Poirié, Jean-Luc Gatti, and Armel Gallet. 2020. "Bacillus thuringiensis Bioinsecticides Induce Developmental Defects in Non-Target Drosophila melanogaster Larvae" Insects 11, no. 10: 697. https://doi.org/10.3390/insects11100697