
Analyzing Generalist Plant Species Using Topographic Characteristics of Picea jezoensis (Siebold & Zucc.) Carrière Forests in East Asia: From China (Mt. Changbai) to South Korea


Abstract
:1. Introduction
2. Materials and Methods
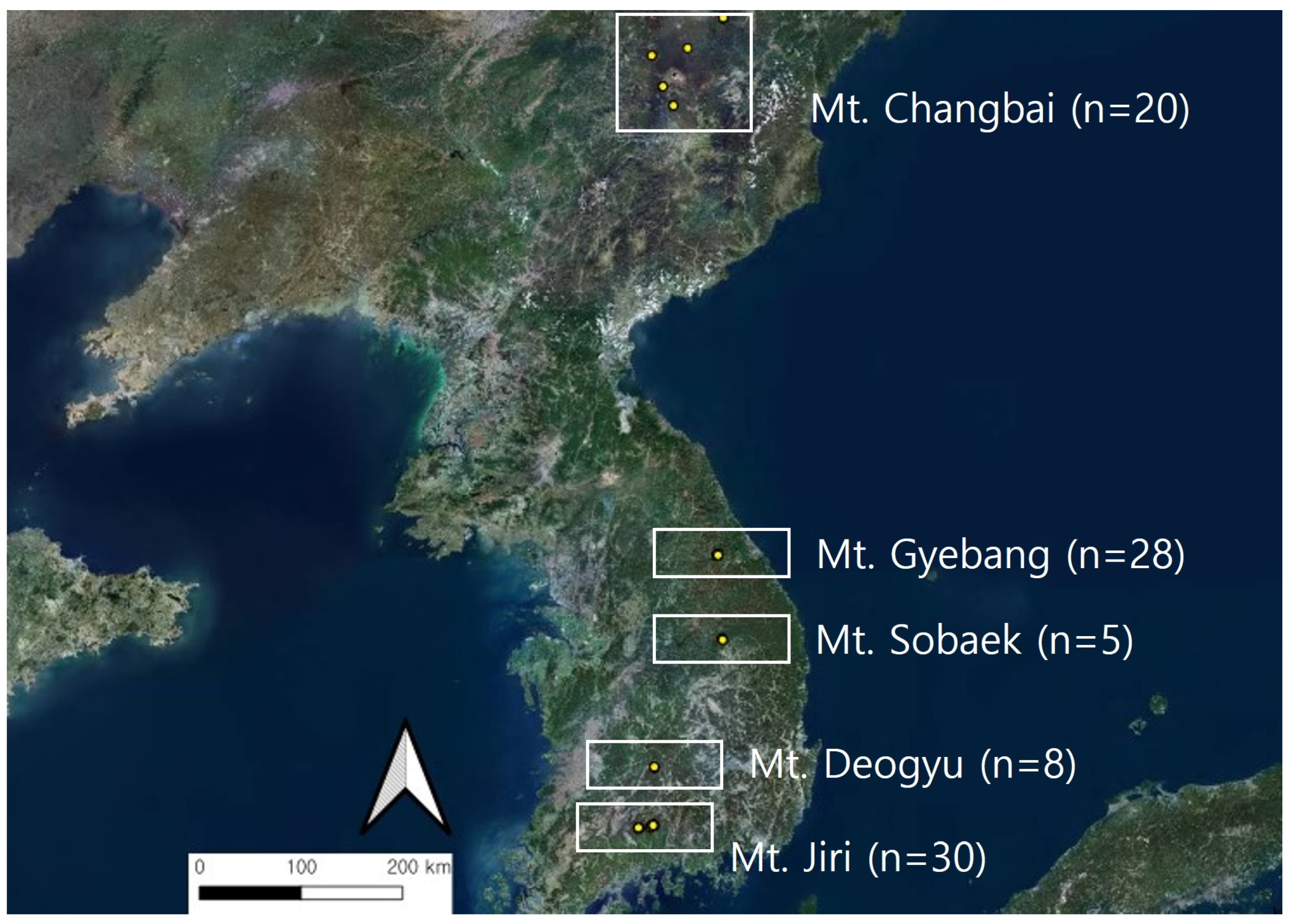
2.1. Overview of the Target Site
2.2. Field Survey Method
2.3. Statistical and Analytical Methods
3. Results
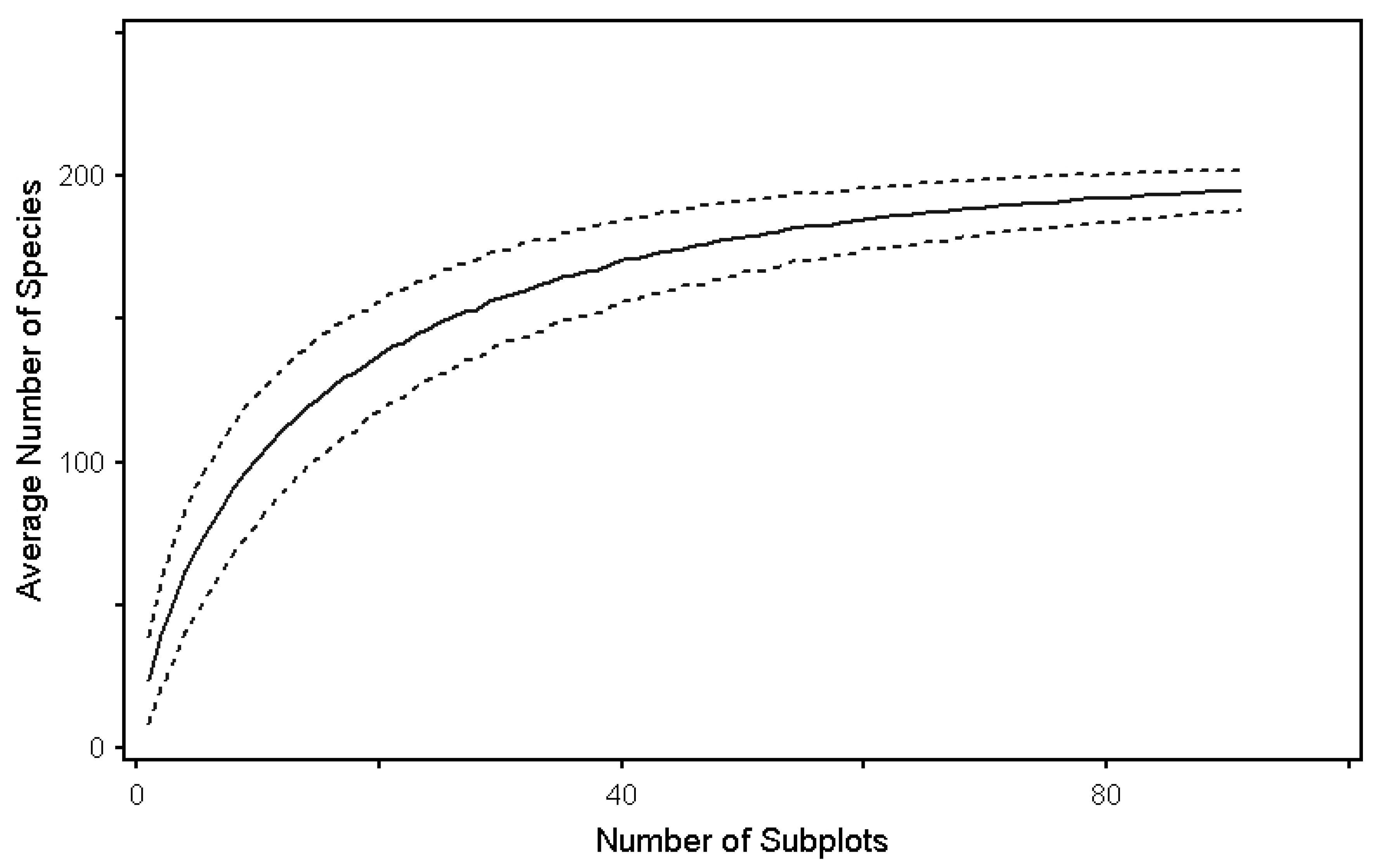
3.1. Species-Area Curves
3.2. Selection of Generalist Species
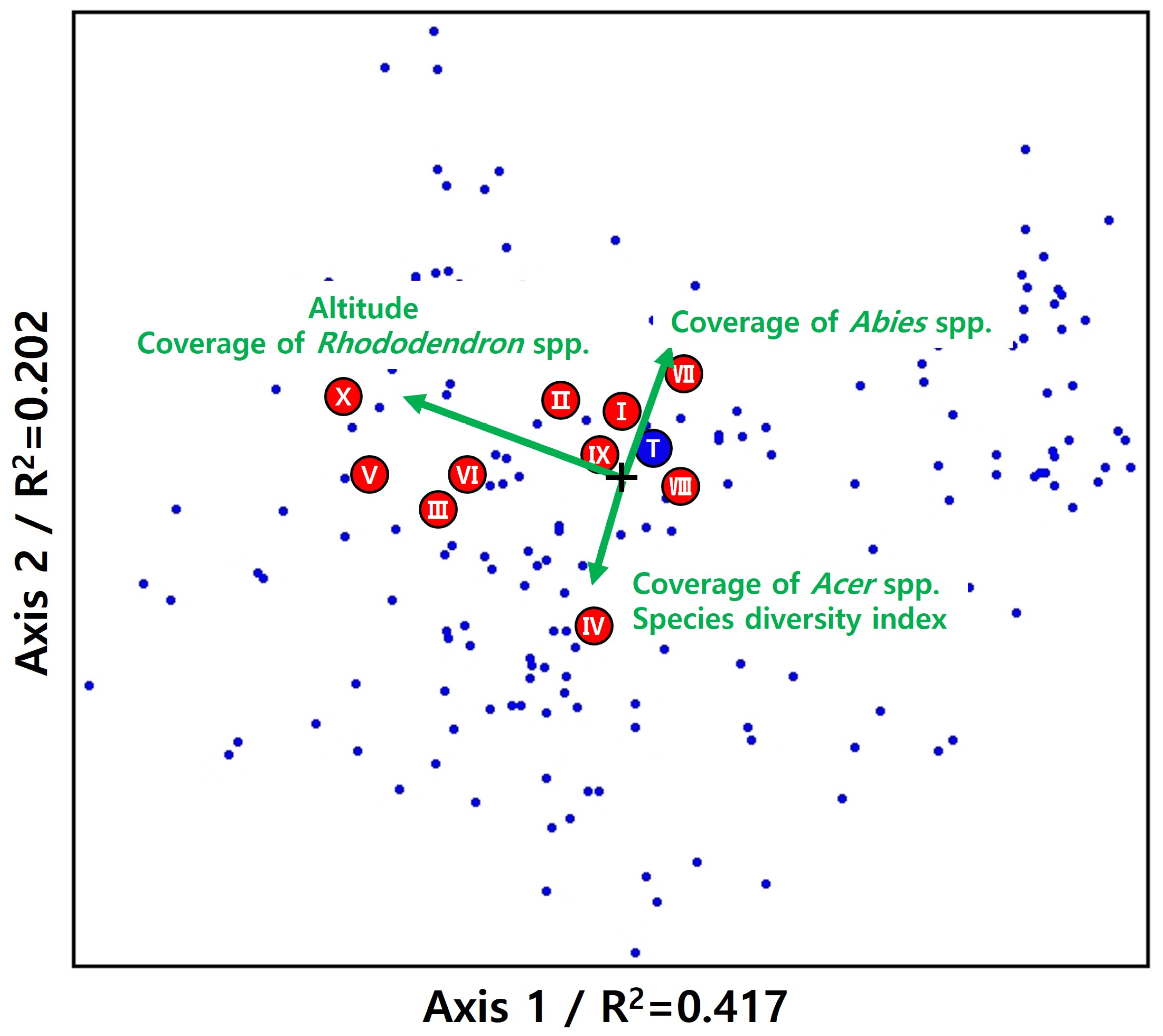
3.3. Correlations between Environmental Factors
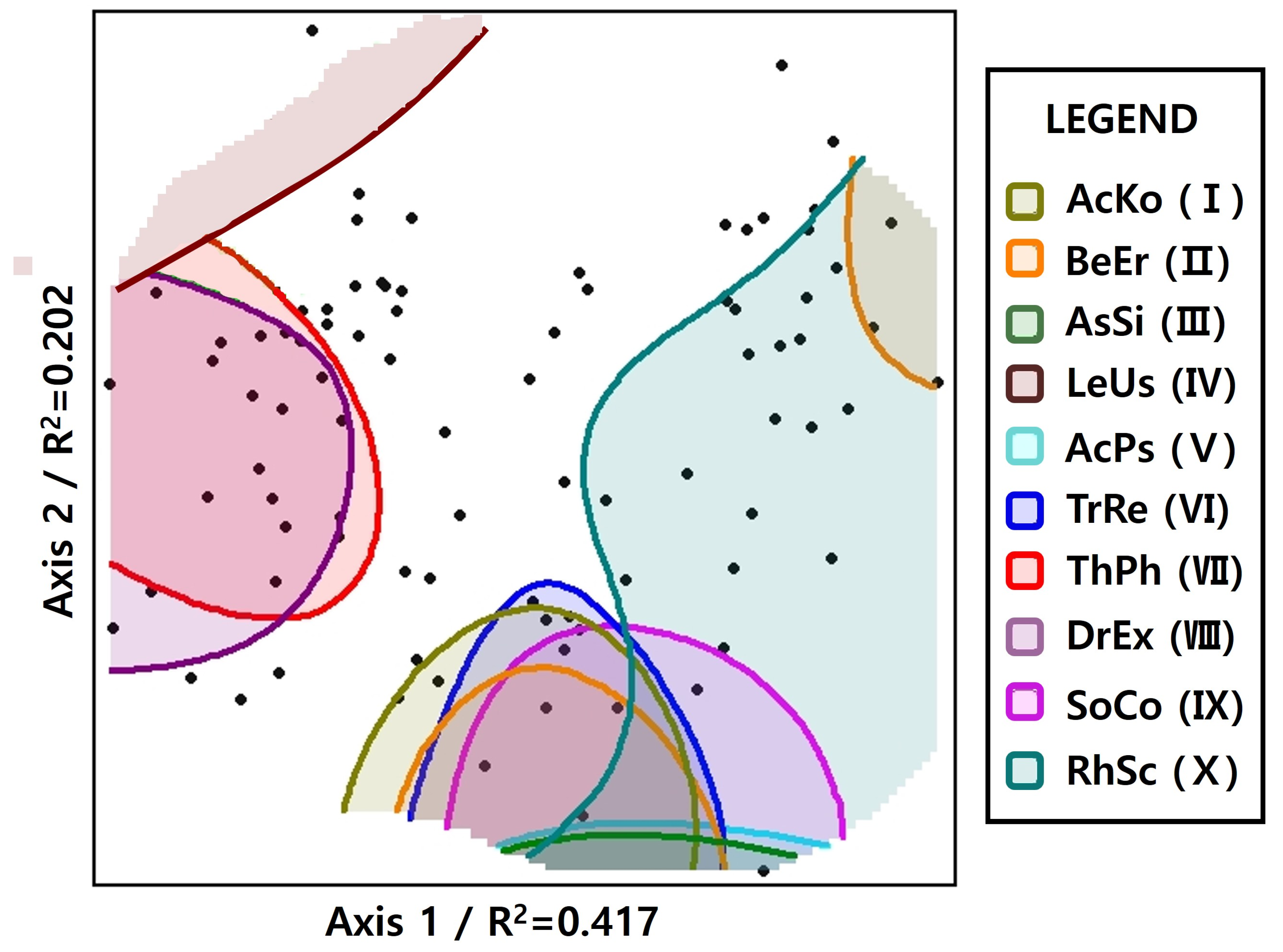
3.4. Convex Hull
4. Discussion
4.1. Ecological Characteristics of Generalist Species
4.2. Correlations between Generalist Species and Ecological Niche Distribution
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Scientific Name | ×1 | ×2 | ×3 | ×4 | ×5 | Mean |
| Abies koreana Wilson | 0.3210 | 0.2000 | 0.2467 | 0.9516 | 0.5379 | 0.4514 |
| Abies nephrolepis (Trautv.) Maxim. | 0.2409 | 0.6784 | 0.4601 | 0.7196 | 0.8919 | 0.5982 |
| Acer barbinerve Maxim. | 0.3842 | 0.7315 | 0.6249 | 0.4055 | 0.4471 | 0.5186 |
| Acer komarovii Pojark. | 0.5506 | 0.6751 | 0.6935 | 0.9049 | 0.8802 | 0.7409 |
| Acer mandshuricum Maxim. | 0.1825 | 0.2000 | 0.2000 | 0.3702 | 0.3497 | 0.2605 |
| Acer pictum subsp. mono (Maxim.) Ohashi | 0.5180 | 0.5519 | 0.5901 | 0.6604 | 0.5332 | 0.5707 |
| Acer pseudosieboldianum (Pax) Kom. | 0.6544 | 0.4904 | 0.6903 | 0.9025 | 0.7197 | 0.6915 |
| Acer tegmentosum Maxim. | 0.2722 | 0.4916 | 0.4916 | 0.7416 | 0.7063 | 0.5406 |
| Acer ukurunduense Trautv. & C.A.Mey. | 0.2805 | 0.4640 | 0.4600 | 0.5489 | 0.9036 | 0.5314 |
| Aconitum jaluense Kom. | 0.3542 | 0.4565 | 0.4565 | 0.7059 | 0.5415 | 0.5029 |
| Aconitum pseudolaeve Nakai | 0.1429 | 0.3291 | 0.2533 | 0.2953 | 0.2524 | 0.2546 |
| Actaea asiatica H. Hara | 0.1920 | 0.2000 | 0.2000 | 0.3456 | 0.4253 | 0.2726 |
| Actinidia arguta (Siebold & Zucc.) Planch. ex Miq. | 0.3094 | 0.2443 | 0.2443 | 0.6333 | 0.5115 | 0.3885 |
| Actinidia polygama (Siebold & Zucc.) Planch. ex Maxim. | 0.1429 | 0.3812 | 0.2000 | 0.3812 | 0.2383 | 0.2687 |
| Adenophora remotiflora (Siebold & Zucc.) Miq. | 0.2593 | 0.2649 | 0.2649 | 0.4667 | 0.4712 | 0.3454 |
| Agastache rugosa (Fisch. & Mey.) Kuntze | 0.2857 | 0.4000 | 0.4000 | 0.8000 | 0.5000 | 0.4771 |
| Agrimonia pilosa Ledeb. | 0.2857 | 0.4000 | 0.4000 | 0.4000 | 0.2500 | 0.3471 |
| Ainsliaea acerifolia Sch.Bip. | 0.5001 | 0.3253 | 0.3645 | 0.8053 | 0.6956 | 0.5382 |
| Angelica amurensis Schischk. | 0.4244 | 0.2477 | 0.4701 | 0.6709 | 0.4193 | 0.4465 |
| Angelica gigas Nakai | 0.3295 | 0.2535 | 0.2559 | 0.6213 | 0.7163 | 0.4353 |
| Anthriscus sylvestris (L.) Hoffm. | 0.1429 | 0.2000 | 0.2000 | 0.3171 | 0.2359 | 0.2192 |
| Aralia elata (Miq.) Seem. | 0.4947 | 0.4241 | 0.6421 | 0.6679 | 0.3174 | 0.5092 |
| Arisaema amurense Maxim. | 0.2062 | 0.2000 | 0.2000 | 0.4000 | 0.3700 | 0.2752 |
| Arisaema peninsulae Nakai | 0.2339 | 0.2000 | 0.2000 | 0.3274 | 0.3456 | 0.2614 |
| Aruncus dioicus var. kamtschaticus (Maxim.) H. Hara | 0.6377 | 0.6558 | 0.5829 | 0.5185 | 0.4685 | 0.5727 |
| Asarum sieboldii Miq. | 0.5411 | 0.5381 | 0.8826 | 0.8952 | 0.6440 | 0.7002 |
| Asplenium yokoscense (Franch. & Sav.) H.Christ | 0.4049 | 0.2115 | 0.3624 | 0.8239 | 0.6195 | 0.4844 |
| Aster scaber Thunb. | 0.5762 | 0.4822 | 0.6155 | 0.6368 | 0.3750 | 0.5371 |
| Astilbe rubra Hook.f. & Thomson | 0.5751 | 0.5410 | 0.5410 | 0.7512 | 0.7401 | 0.6297 |
| Athyrium brevifrons Kodama ex Nakai | 0.2404 | 0.8458 | 0.6476 | 0.4577 | 0.6951 | 0.5773 |
| Athyrium niponicum (Mett.) Hance | 0.3673 | 0.4000 | 0.4000 | 0.7200 | 0.3750 | 0.4525 |
| Berberis amurensis var. brevifolia Nakai | 0.3737 | 0.4000 | 0.4000 | 0.5232 | 0.2400 | 0.3874 |
| Betula costata Trautv. | 0.2822 | 0.3951 | 0.2000 | 0.3340 | 0.2088 | 0.2840 |
| Betula davurica Pall. | 0.2125 | 0.2974 | 0.2974 | 0.5324 | 0.3328 | 0.3345 |
| Betula ermanii Cham. | 0.7720 | 0.4972 | 0.7275 | 0.8812 | 0.7288 | 0.7214 |
| Betula platyphylla var. japonica (Miq.) H. Hara | 0.1429 | 0.2000 | 0.2000 | 0.3028 | 0.3012 | 0.2294 |
| Betula schmidtii Regel | 0.1429 | 0.2830 | 0.2830 | 0.2830 | 0.1769 | 0.2338 |
| Bistorta manshuriensis (Petrov ex Kom.) Kom. | 0.2407 | 0.2000 | 0.2000 | 0.3370 | 0.2106 | 0.2377 |
| Calamagrostis arundinacea (L.) Roth | 0.5561 | 0.4203 | 0.4645 | 0.8610 | 0.8218 | 0.6247 |
| Cardamine komarovii Nakai | 0.1429 | 0.5556 | 0.5556 | 0.7143 | 0.2404 | 0.4417 |
| Cardamine leucantha (Tausch) O.E.Schulz | 0.1429 | 0.2000 | 0.2000 | 0.4000 | 0.2500 | 0.2386 |
| Carex biwensis Franch. | 0.1429 | 0.2000 | 0.2000 | 0.4000 | 0.2500 | 0.2386 |
| Carex erythrobasis H.Lév. & Vaniot | 0.3136 | 0.5651 | 0.2915 | 0.5525 | 0.5679 | 0.4581 |
| Carex glabrescens Ohwi | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.1250 | 0.1736 |
| Carex hakonensis Franch. & Sav. | 0.5190 | 0.5685 | 0.5685 | 0.5310 | 0.3566 | 0.5087 |
| Carex humilis var. nana (H.Lév. & Vaniot) Ohwi | 0.5708 | 0.4575 | 0.7294 | 0.4922 | 0.5746 | 0.5649 |
| Carex lanceolata Boott | 0.5135 | 0.3155 | 0.5780 | 0.8294 | 0.2812 | 0.5035 |
| Carex okamotoi Ohwi | 0.2056 | 0.2000 | 0.2000 | 0.6939 | 0.3745 | 0.3348 |
| Carex siderosticta Hance | 0.2676 | 0.3567 | 0.3625 | 0.6276 | 0.6413 | 0.4511 |
| Caulophyllum robustum Maxim. | 0.2857 | 0.2000 | 0.2000 | 0.4000 | 0.2500 | 0.2671 |
| Chrysosplenium flagelliferum F.Schmidt | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.3621 | 0.2210 |
| Cimicifuga dahurica (Turcz. ex Fisch. & C.A.Mey.) Maxim. | 0.3155 | 0.2000 | 0.2000 | 0.6055 | 0.6106 | 0.3863 |
| Cimicifuga simplex (DC.) Turcz. | 0.1429 | 0.3340 | 0.2000 | 0.2000 | 0.1250 | 0.2004 |
| Circaea alpina L. | 0.1429 | 0.2000 | 0.2000 | 0.5155 | 0.3222 | 0.2761 |
| Clematis fusca var. violacea Maxim. | 0.3452 | 0.3951 | 0.3951 | 0.3951 | 0.2469 | 0.3555 |
| Clematis koreana Kom. | 0.1977 | 0.2000 | 0.2000 | 0.5686 | 0.4820 | 0.3296 |
| Clintonia udensis Trautv. & C.A.Mey. | 0.3632 | 0.5168 | 0.4404 | 0.7506 | 0.6750 | 0.5492 |
| Cornus controversa Hemsl. | 0.1429 | 0.3514 | 0.3514 | 0.5070 | 0.2394 | 0.3184 |
| Corylus heterophylla Fisch. ex Trautv. | 0.2163 | 0.4820 | 0.4820 | 0.4820 | 0.1892 | 0.3703 |
| Corylus sieboldiana var. mandshurica (Maxim. & Rupr.) | 0.2657 | 0.2000 | 0.2000 | 0.3797 | 0.5552 | 0.3201 |
| Cymopterus melanotilingia (H.Boissieu) C.Y.Yoon | 0.5352 | 0.3951 | 0.5686 | 0.8695 | 0.5091 | 0.5755 |
| Deutzia glabrata Kom. | 0.2740 | 0.2883 | 0.2883 | 0.5640 | 0.6025 | 0.4034 |
| Deutzia parviflora Bunge | 0.2723 | 0.2000 | 0.3812 | 0.3812 | 0.2383 | 0.2946 |
| Diarrhena fauriei (Hack.) Ohwi | 0.2723 | 0.3812 | 0.3812 | 0.3812 | 0.2383 | 0.3308 |
| Diarrhena mandshurica Maxim. | 0.1429 | 0.2000 | 0.2000 | 0.5684 | 0.2088 | 0.2640 |
| Disporum smilacinum A.Gray | 0.4861 | 0.3836 | 0.5893 | 0.4836 | 0.4253 | 0.4736 |
| Disporum viridescens (Maxim.) Nakai | 0.2857 | 0.4000 | 0.4000 | 0.4000 | 0.2500 | 0.3471 |
| Dryopteris chinensis (Baker) Koidz. | 0.4060 | 0.3951 | 0.5685 | 0.2000 | 0.2469 | 0.3633 |
| Dryopteris crassirhizoma Nakai | 0.4039 | 0.5375 | 0.5402 | 0.6442 | 0.8201 | 0.5892 |
| Dryopteris expansa (C.Presl) Fraser-Jenk. & Jermy | 0.4433 | 0.6492 | 0.6102 | 0.8299 | 0.8398 | 0.6745 |
| Eleutherococcus senticosus (Rupr. & Maxim.) Maxim. | 0.2820 | 0.2621 | 0.2000 | 0.4077 | 0.2956 | 0.2895 |
| Epilobium angustifolium L. | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.1250 | 0.1736 |
| Equisetum hyemale L. | 0.2336 | 0.2000 | 0.2000 | 0.4891 | 0.5294 | 0.3304 |
| Euonymus macropterus Rupr. | 0.6096 | 0.6037 | 0.6617 | 0.5648 | 0.7545 | 0.6389 |
| Euonymus oxyphyllus Miq. | 0.3333 | 0.3379 | 0.3379 | 0.3920 | 0.3224 | 0.3447 |
| Euonymus pauciflorus Maxim. | 0.1429 | 0.2000 | 0.2000 | 0.3774 | 0.3621 | 0.2565 |
| Euonymus sachalinensis (F.Schmidt) Maxim. | 0.2682 | 0.4977 | 0.5555 | 0.4468 | 0.3472 | 0.4231 |
| Filipendula glaberrima Nakai | 0.4622 | 0.5253 | 0.5253 | 0.5285 | 0.5873 | 0.5257 |
| Fraxinus rhynchophylla Hance | 0.3774 | 0.5440 | 0.5440 | 0.5173 | 0.4327 | 0.4831 |
| Fraxinus sieboldiana Blume | 0.4412 | 0.2000 | 0.2600 | 0.7338 | 0.6768 | 0.4624 |
| Galium kamtschaticum Steller ex (Roem. & Schult.) | 0.1429 | 0.4000 | 0.2000 | 0.2000 | 0.2500 | 0.2386 |
| Gentiana triflora var. japonica (Kusn.) H. Hara | 0.2286 | 0.3200 | 0.3200 | 0.3200 | 0.5000 | 0.3377 |
| Geranium koreanum Kom. | 0.2857 | 0.4000 | 0.4000 | 0.4000 | 0.2500 | 0.3471 |
| Hemerocallis hakuunensis Nakai | 0.1429 | 0.2000 | 0.2000 | 0.2826 | 0.2613 | 0.2173 |
| Hosta capitata (Koidz.) Nakai | 0.3507 | 0.2000 | 0.3951 | 0.4910 | 0.2470 | 0.3368 |
| Hosta plantaginea (Lam.) Asch. | 0.1429 | 0.3340 | 0.2000 | 0.3340 | 0.3553 | 0.2732 |
| Hydrangea serrata f. acuminata (Siebold & Zucc.) E.H.Wilson | 0.3280 | 0.3995 | 0.4592 | 0.2352 | 0.3512 | 0.3546 |
| Hydrocotyle sibthorpioides Lam. | 0.1429 | 0.2000 | 0.2000 | 0.3812 | 0.2382 | 0.2325 |
| Hypericum ascyron L. | 0.2857 | 0.2000 | 0.2000 | 0.4000 | 0.2500 | 0.2671 |
| Impatiens nolitangere L. | 0.2101 | 0.2000 | 0.2000 | 0.3600 | 0.4767 | 0.2894 |
| Isodon excisus (Maxim.) Kudo | 0.3321 | 0.2231 | 0.2231 | 0.7402 | 0.6529 | 0.4343 |
| Juglans mandshurica Maxim. | 0.1429 | 0.4000 | 0.4000 | 0.2000 | 0.1250 | 0.2536 |
| Kalopanax septemlobus (Thunb.) Koidz. | 0.1429 | 0.2000 | 0.2000 | 0.4000 | 0.2500 | 0.2386 |
| Larix olgensis var. koreana (Nakai) Nakai | 0.2723 | 0.3812 | 0.2000 | 0.2000 | 0.2382 | 0.2583 |
| Lepisorus ussuriensis (Regel & Maack) Ching | 0.5054 | 0.6760 | 0.8921 | 0.6043 | 0.8108 | 0.6977 |
| Ligularia fischeri (Ledeb.) Turcz. | 0.5812 | 0.5502 | 0.6168 | 0.6845 | 0.5974 | 0.6060 |
| Lilium distichum Nakai ex Kamib. | 0.4083 | 0.3802 | 0.3802 | 0.5716 | 0.3573 | 0.4195 |
| Lilium tsingtauense Gilg | 0.2339 | 0.3274 | 0.3274 | 0.5529 | 0.1250 | 0.3133 |
| Lonicera caerulea var. edulis Turcz. ex Herder | 0.1429 | 0.3456 | 0.2000 | 0.3456 | 0.2160 | 0.2500 |
| Lonicera chrysantha Turcz. | 0.3982 | 0.2999 | 0.2999 | 0.6506 | 0.7263 | 0.4750 |
| Lonicera maackii (Rupr.) Maxim. | 0.2463 | 0.2000 | 0.2000 | 0.2000 | 0.2155 | 0.2124 |
| Lonicera sachalinensis (F.Schmidt) Nakai | 0.2302 | 0.3223 | 0.3223 | 0.3486 | 0.5614 | 0.3570 |
| Lonicera tatarinowii var. leptantha (Rehder) Nakai | 0.1429 | 0.2912 | 0.2000 | 0.2722 | 0.4045 | 0.2622 |
| Lychnis cognata Maxim. | 0.4286 | 0.2000 | 0.2000 | 0.3600 | 0.3750 | 0.3127 |
| Lycopodium chinense H.Christ | 0.2368 | 0.4954 | 0.2972 | 0.5843 | 0.5630 | 0.4353 |
| Lycopodium obscurum L. | 0.2021 | 0.3618 | 0.2829 | 0.5528 | 0.4579 | 0.3715 |
| Lycopodium serratum Thunb. | 0.2923 | 0.6467 | 0.7273 | 0.6845 | 0.6056 | 0.5913 |
| Magnolia sieboldii K.Koch | 0.3708 | 0.3328 | 0.3382 | 0.5491 | 0.5104 | 0.4202 |
| Maianthemum bifolium (L.) F.W.Schmidt | 0.2482 | 0.3474 | 0.3474 | 0.3474 | 0.4765 | 0.3534 |
| Maianthemum dilatatum (Wood) A.Nelson & J.F.Macbr. | 0.1637 | 0.3969 | 0.2000 | 0.5218 | 0.5680 | 0.3701 |
| Malus baccata (L.) Borkh. | 0.2723 | 0.3812 | 0.3812 | 0.3812 | 0.2382 | 0.3308 |
| Meehania urticifolia (Miq.) Makino | 0.2740 | 0.4657 | 0.4657 | 0.7925 | 0.7707 | 0.5537 |
| Oplopanax elatus (Nakai) Nakai | 0.2405 | 0.3367 | 0.3367 | 0.7484 | 0.4458 | 0.4216 |
| Osmunda cinnamomea var. forkiensis Copel. | 0.2772 | 0.3881 | 0.2000 | 0.2000 | 0.3371 | 0.2805 |
| Ostericum grosseserratum (Maxim.) Kitag. | 0.1429 | 0.2000 | 0.2000 | 0.5104 | 0.4301 | 0.2967 |
| Oxalis acetosella L. | 0.2175 | 0.7986 | 0.4526 | 0.5912 | 0.6992 | 0.5518 |
| Oxalis corniculata L. | 0.1429 | 0.3812 | 0.2000 | 0.2000 | 0.2383 | 0.2325 |
| Paeonia japonica (Makino) Miyabe & Takeda | 0.2571 | 0.3600 | 0.3600 | 0.3600 | 0.3750 | 0.3424 |
| Parasenecio adenostyloides (Franch. & Sav. ex Maxim.) H.Koyama | 0.1798 | 0.3513 | 0.2456 | 0.5639 | 0.4906 | 0.3663 |
| Parasenecio auriculata var. kamtschatica (Maxim.) H.Koyama | 0.2608 | 0.4296 | 0.4296 | 0.4391 | 0.5727 | 0.4263 |
| Parasenecio auriculata var. matsumurana Nakai | 0.3567 | 0.3858 | 0.4486 | 0.5101 | 0.3188 | 0.4040 |
| Paris verticillata M.Bieb. | 0.4595 | 0.3701 | 0.3701 | 0.7476 | 0.5360 | 0.4966 |
| Patrinia saniculaefolia Hemsl. | 0.1429 | 0.2000 | 0.2000 | 0.4000 | 0.2500 | 0.2386 |
| Pedicularis resupinata L. | 0.6221 | 0.4593 | 0.4995 | 0.6834 | 0.7320 | 0.5993 |
| Philadelphus tenuifolius Rupr. & Maxim. | 0.3506 | 0.4194 | 0.4194 | 0.9324 | 0.4525 | 0.5149 |
| Picea jezoensis (Siebold & Zucc.) Carrière | 0.5357 | 0.7221 | 0.7366 | 0.8844 | 0.8267 | 0.7411 |
| Pilea mongolica Wedd. | 0.2857 | 0.4000 | 0.4000 | 0.4000 | 0.2500 | 0.3471 |
| Pinus densiflora Siebold & Zucc. | 0.2265 | 0.3171 | 0.3171 | 0.3171 | 0.1982 | 0.2752 |
| Pinus koraiensis Siebold & Zucc. | 0.4197 | 0.4702 | 0.5293 | 0.8479 | 0.8306 | 0.6195 |
| Polygonatum odoratum var. pluriflorum (Miq.) Ohwi | 0.2131 | 0.2983 | 0.2983 | 0.2983 | 0.4611 | 0.3138 |
| Polystichum braunii (Spenn.) Fee | 0.1429 | 0.2601 | 0.2601 | 0.3982 | 0.3388 | 0.2800 |
| Polystichum tripteron (Kunze) C.Presl | 0.3690 | 0.4297 | 0.4497 | 0.7268 | 0.7183 | 0.5387 |
| Populus maximowiczii A.Henry | 0.2857 | 0.4000 | 0.4000 | 0.4000 | 0.2500 | 0.3471 |
| Potentilla fragarioides var. major Maxim. | 0.1429 | 0.3449 | 0.3449 | 0.3449 | 0.2155 | 0.2786 |
| Primula jesoana Miq. | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.1250 | 0.1736 |
| Prunus maximowiczii Rupr. | 0.2844 | 0.2000 | 0.2000 | 0.3982 | 0.3451 | 0.2856 |
| Prunus padus L. | 0.2524 | 0.2490 | 0.2490 | 0.6954 | 0.8644 | 0.4620 |
| Prunus sargentii Rehder | 0.2266 | 0.3172 | 0.3172 | 0.3172 | 0.1982 | 0.2753 |
| Pseudostellaria heterophylla (Miq.) Pax ex Pax & Hoffm. | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.1250 | 0.1736 |
| Pseudostellaria palibiniana (Takeda) Ohwi | 0.2725 | 0.3048 | 0.3048 | 0.6185 | 0.5527 | 0.4106 |
| Pseudostellaria setulosa Ohwi | 0.3092 | 0.3821 | 0.3821 | 0.5302 | 0.4278 | 0.4063 |
| Pyrola renifolia Maxim. | 0.1429 | 0.2000 | 0.2000 | 0.4910 | 0.3069 | 0.2681 |
| Quercus mongolica Fisch. ex Ledeb. | 0.4982 | 0.5348 | 0.7782 | 0.8367 | 0.5108 | 0.6317 |
| Rhododendron mucronulatum var. ciliatum Nakai | 0.5280 | 0.2886 | 0.3532 | 0.9478 | 0.6356 | 0.5506 |
| Rhododendron schlippenbachii Maxim. | 0.7110 | 0.3748 | 0.5172 | 0.9672 | 0.7425 | 0.6625 |
| Rhododendron tschonoskii Maxim. | 0.4209 | 0.2000 | 0.3664 | 0.4504 | 0.3246 | 0.3525 |
| Ribes mandshuricum (Maxim.) Kom. | 0.4083 | 0.3802 | 0.3802 | 0.3802 | 0.3573 | 0.3812 |
| Ribes maximowiczianum Kom. | 0.5241 | 0.4558 | 0.5432 | 0.5498 | 0.5126 | 0.5171 |
| Rodgersia podophylla A.Gray | 0.3456 | 0.2000 | 0.2000 | 0.6989 | 0.3943 | 0.3678 |
| Rosa davurica Pall. | 0.2766 | 0.3990 | 0.3990 | 0.2517 | 0.2494 | 0.3151 |
| Rosa suavis Willd. | 0.3412 | 0.4040 | 0.3675 | 0.8889 | 0.7396 | 0.5482 |
| Rubia akane Nakai | 0.2920 | 0.3662 | 0.3662 | 0.5598 | 0.2555 | 0.3680 |
| Rubia chinensis Regel & Maack | 0.2206 | 0.2000 | 0.2000 | 0.2000 | 0.1930 | 0.2027 |
| Rubia cordifolia var. pratensis Maxim. | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.1250 | 0.1736 |
| Rubus crataegifolius Bunge | 0.2857 | 0.4000 | 0.4000 | 0.4000 | 0.2500 | 0.3471 |
| Rubus idaeus var. microphyllus Turcz. | 0.1429 | 0.3028 | 0.2000 | 0.4820 | 0.4747 | 0.3205 |
| Salix caprea L. | 0.5149 | 0.3581 | 0.5453 | 0.5453 | 0.2531 | 0.4433 |
| Sambucus sieboldiana var. miquelii (Nakai) Hara | 0.1429 | 0.2000 | 0.2000 | 0.3812 | 0.2383 | 0.2325 |
| Sambucus williamsii var. coreana (Nakai) Nakai | 0.6465 | 0.4043 | 0.6774 | 0.6108 | 0.4971 | 0.5672 |
| Sasa borealis (Hack.) Makino | 0.2206 | 0.2000 | 0.2363 | 0.3989 | 0.2432 | 0.2598 |
| Saussurea gracilis Maxim. | 0.4791 | 0.2324 | 0.2324 | 0.7460 | 0.4339 | 0.4248 |
| Saussurea grandifolia Maxim. | 0.4591 | 0.6427 | 0.5334 | 0.5334 | 0.3334 | 0.5004 |
| Saxifraga fortunei var. incisolobata (Engl. & Irmsch.) Nakai | 0.2986 | 0.5322 | 0.5322 | 0.2826 | 0.2612 | 0.3814 |
| Saxifraga oblongifolia Nakai | 0.1429 | 0.2000 | 0.2000 | 0.4000 | 0.1250 | 0.2136 |
| Saxifraga octopetala Nakai | 0.1429 | 0.2000 | 0.2000 | 0.4000 | 0.2500 | 0.2386 |
| Schisandra chinensis (Turcz.) Baill. | 0.3810 | 0.3200 | 0.3200 | 0.4000 | 0.3333 | 0.3509 |
| Sedum polytrichoides Hemsl. | 0.2857 | 0.4000 | 0.4000 | 0.2000 | 0.2500 | 0.3071 |
| Smilacina japonica A.Gray | 0.2774 | 0.5764 | 0.5764 | 0.5007 | 0.3966 | 0.4655 |
| Solidago virgaurea subsp. asiatica Kitam. ex Hara | 0.7979 | 0.3458 | 0.4939 | 0.8644 | 0.7344 | 0.6473 |
| Sorbus alnifolia (Siebold & Zucc.) K.Koch | 0.1429 | 0.2000 | 0.2000 | 0.3600 | 0.1250 | 0.2056 |
| Sorbus commixta Hedl. | 0.4706 | 0.5892 | 0.6250 | 0.8114 | 0.8250 | 0.6642 |
| Spiraea chamaedryfolia L. | 0.2286 | 0.3200 | 0.3200 | 0.3200 | 0.5000 | 0.3377 |
| Spiraea fritschiana C.K.Schneid. | 0.2131 | 0.2983 | 0.2983 | 0.3891 | 0.2355 | 0.2868 |
| Spodipogon cotulifer (Thunb.) Hack. | 0.2286 | 0.2000 | 0.3200 | 0.3200 | 0.5000 | 0.3137 |
| Streptopus amplexifolius var. papillatus Ohwi | 0.1429 | 0.3340 | 0.2000 | 0.2000 | 0.2088 | 0.2171 |
| Streptopus koreanus (Kom.) Ohwi | 0.1429 | 0.3787 | 0.2000 | 0.2000 | 0.4479 | 0.2739 |
| Streptopus ovalis (Ohwi) F.T.Wang & Y.C.Tang | 0.2571 | 0.2000 | 0.2000 | 0.6000 | 0.3750 | 0.3264 |
| Symplocos chinensis f. pilosa (Nakai) Ohwi | 0.3902 | 0.3999 | 0.4704 | 0.7879 | 0.4843 | 0.5065 |
| Synurus deltoides (Aiton) Nakai | 0.5387 | 0.2689 | 0.2689 | 0.6805 | 0.5690 | 0.4652 |
| Syringa patula (Palib.) Nakai | 0.3301 | 0.2638 | 0.2638 | 0.3359 | 0.2249 | 0.2837 |
| Taxus cuspidata Siebold & Zucc. | 0.4287 | 0.3665 | 0.3824 | 0.7177 | 0.7628 | 0.5316 |
| Thalictrum aquilegifolium var. sibiricum Regel & Tiling | 0.2183 | 0.3057 | 0.3057 | 0.3057 | 0.1911 | 0.2653 |
| Thalictrum filamentosum var. tenerum (Huth) Ohwi | 0.1978 | 0.3948 | 0.3255 | 0.3255 | 0.2868 | 0.3061 |
| Thelypteris japonica (Baker) Ching | 0.2857 | 0.4000 | 0.4000 | 0.4000 | 0.3333 | 0.3638 |
| Thelypteris phegopteris (L.) Sloss. | 0.6597 | 0.5766 | 0.5351 | 0.8358 | 0.7782 | 0.6771 |
| Thuja koraiensis Nakai | 0.1947 | 0.2726 | 0.2726 | 0.3554 | 0.3136 | 0.2818 |
| Tilia amurensis Rupr. | 0.2034 | 0.5254 | 0.4835 | 0.6196 | 0.5732 | 0.4810 |
| Trillium kamtschaticum Pall. ex Pursh | 0.1429 | 0.4996 | 0.4996 | 0.5331 | 0.4579 | 0.4266 |
| Trillium tschonoskii Maxim. | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.2332 | 0.1952 |
| Tripterygium regelii Sprague & Takeda | 0.7049 | 0.4501 | 0.5921 | 0.8652 | 0.8255 | 0.6876 |
| Ulmus laciniata (Trautv.) Mayr | 0.2445 | 0.2000 | 0.2000 | 0.3689 | 0.1590 | 0.2345 |
| Vaccinium hirtum var. koreanum (Nakai) Kitam. | 0.4794 | 0.5267 | 0.5267 | 0.4585 | 0.3246 | 0.4632 |
| Veratrum maackii var. japonicum (Baker) T.Schmizu | 0.1429 | 0.4000 | 0.4000 | 0.4000 | 0.1250 | 0.2936 |
| Veratrum oxysepalum Turcz. | 0.2822 | 0.3951 | 0.3951 | 0.5685 | 0.3553 | 0.3992 |
| Viburnum opulus var. calvescens (Rehder) H. Hara | 0.1997 | 0.2795 | 0.2795 | 0.2795 | 0.1747 | 0.2426 |
| Viola selkirkii Pursh ex Goldie | 0.3487 | 0.5919 | 0.5919 | 0.5222 | 0.3264 | 0.4762 |
| Viola verecunda A.Gray | 0.1429 | 0.2000 | 0.2000 | 0.2000 | 0.2500 | 0.1986 |
| Weigela florida (Bunge) A.DC. | 0.7171 | 0.4287 | 0.5220 | 0.6481 | 0.6917 | 0.6015 |
| Woodsia polystichoides D.C.Eaton | 0.3810 | 0.2000 | 0.3200 | 0.5333 | 0.3333 | 0.3535 |
References
- Rapoport, E.H. Areography: Geographical Strategies of Species; Elsevier: London, UK, 2013; Volume 1, pp. 225–230. ISBN 9780080289144. [Google Scholar]
- Whittaker, R.H.; Levin, S.A.; Root, R.B. Niche, Habitat, and Ecotope. Am. Nat. 1973, 107, 321–338. [Google Scholar] [CrossRef]
- Salazar, L.; Homeier, J.; Kessler, M.; Abrahamczyk, S.; Lehnert, M.; Krömer, T.; Kluge, J. Diversity patterns of ferns along elevational gradients in andean tropical forests. Plant Ecol. Divers. 2015, 8, 13–24. [Google Scholar] [CrossRef]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Diniz-Filho, J.A.F.; Jaramillo, C.A.; Soeller, S.A. Post-Eocene climate change, niche conservatism, and the latitudinal diversity gradient of New World birds. J. Biogeogr. 2006, 33, 770–780. [Google Scholar] [CrossRef]
- Manor, A.; Shnerb, N.M. Facilitation, competition, and vegetation patchiness: From scale free distribution to patterns. J. Theor. Biol. 2008, 253, 838–842. [Google Scholar] [CrossRef] [PubMed]
- Yeocheon Ecological Research Association. Modern Ecological Experiment; Gyomunsa: Seoul, Republic of Korea, 2005; pp. 166–179. ISBN 9788936307516. [Google Scholar]
- Germino, M.J.; Smith, W.K.; Resor, A.C. Conifer seedling distribution and survival in an alpine-treeline ecotone. Plant Ecol. 2002, 162, 157–168. [Google Scholar] [CrossRef]
- Mori, A.; Mizumachi, E.; Osono, T.; Doi, Y. Substrate-associated seedling recruitment and establishment of major conifer species in an old-growth subalpine forest in central Japan. For. Ecol. Manag. 2004, 196, 287–297. [Google Scholar] [CrossRef]
- Hunziker, U.; Brang, P. Microsite patterns of conifer seedling establishment and growth in a mixed stand in the southern Alps. For. Ecol. Manag. 2005, 210, 67–79. [Google Scholar] [CrossRef]
- Hasegawa, S.F.; Mori, A. Structural characteristics of Abies mariesii saplings in a snowy subalpine parkland in central Japan. Tree Physiol. 2007, 27, 141–148. [Google Scholar]
- Han, A.R.; Lee, S.K.; Suh, G.U.; Park, Y.; Park, P.S. Wind and topography influence the crown growth of Picea jezoensis in a subalpine forest on Mt. Deogyu, Korea. Agric. For. Meteorol. 2012, 166–167, 207–214. [Google Scholar] [CrossRef]
- Park, H.C.; Lee, H.Y.; Lee, N.Y.; Lee, H.; Song, J.Y. Survey on the distribution of Evergreen Conifers in the Major national Park—A case Study on Seoraksan, Odaesan, Taebaeksan, Sobaaeksan, Doegyusan, Jirisan National Park. J. Nat. Park. Res. 2019, 10, 224–231. [Google Scholar]
- Kong, W.S. Species composition and distribution of Korean Alpine Plants. J. Korean Geogr. Soc. 2002, 37, 357–370. [Google Scholar]
- The Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 1 August 2023).
- Nakagawa, M.; Kurahashi, A.; Hogetsu, T. The regeneration characteristics of Picea jezoensis and Abies sachalinensis on cut stumps in the sub-boreal forests of Hokkaido Tokyo University Forest. For. Ecol. Manag. 2003, 180, 353–359. [Google Scholar] [CrossRef]
- Aizawa, M.; Yoshimaru, H.; Saito, H.; Katsuki, T.; Kawahara, T.; Kitamura, K.; Shi, F.; Sabirov, R.; Kaji, M. Range-wide genetic structure in a north-east Asian spruce (Picea jezoensis) determined using nuclear microsatellite markers. J. Biogeogr. 2009, 36, 996–1007. [Google Scholar] [CrossRef]
- Park, S.J. Generality and Specificity of Landforms of the Korean Peninsula, and Its Sustainability. J. Korean Geogr. Soc. 2014, 49, 656–674. [Google Scholar]
- Park, G.E.; Kim, E.S.; Jung, S.C.; Yun, C.W.; Kim, J.S.; Kim, J.D.; Kim, J.B.; Lim, J.H. Distribution and Stand Dynamics of Subalpine Conifer Species (Abies nephrolepis, A. koreana, and Picea jezoensis) in Baekdudaegan Protected Area. J. Korean Soc. For. Sci. 2022, 111, 61–71. [Google Scholar]
- Park, B.J.; Byeon, J.G.; Heo, T.I.; Cheon, K.; Yang, J.C.; Oh, S.H. Comparison of species composition among Picea jezoensis (Siebold & Zucc.) carrière forests in Northeast Asia (from China to South Korea). J. Asia-Pac. Biodivers. 2023, 16, 272–281. [Google Scholar]
- Odion, D.C.; Sarr, D.A. Managing disturbance regimes to maintain diversity in forested ecosystems of the Pacific Northwest. For. Ecol. Manag. 2007, 246, 57–65. [Google Scholar] [CrossRef]
- Scott, T.A.; Sullivan, J.E. The Selection and Design of Multiple Species Preserves. Environ. Manag. 2000, 26, S37–S53. [Google Scholar] [CrossRef]
- Barrows, C.W.; Swartz, M.B.; Hodges, W.L.; Allen, M.F.; Rotenberry, J.T.; Li, B.L.; Scott, T.A.; Chen, X. A Framework for Monitoring Multiple-species Conservation Plans. J. Wildl. Manag. 2005, 69, 1333–1345. [Google Scholar] [CrossRef]
- The Services of Climatic Data Portal of Korea Meteorological Administration. Available online: https://data.kma.go.kr/cmmn/main.do (accessed on 1 August 2023).
- Braun-Blanquet, J. Pflanzensoziologie, Grundzfige der Vegetationskunde, 3rd ed.; Springer: New York, NY, USA, 1965; pp. 7–16. ISBN 9783540034789. [Google Scholar]
- Lee, T.B. Coloured Flora of Korea; Hyangmoonsa: Seoul, Republic of Korea, 2003; Volume 1, pp. 1–916. ISBN 9788971871959. [Google Scholar]
- Korea Fern Society. Ferns and Fern Allies of Korea; Geobook: Seoul, Republic of Korea, 2005; pp. 1–399. ISBN 9788995504925. [Google Scholar]
- Knowledge System of National Species in Korea. Available online: http://www.nature.go.kr/kpni/ (accessed on 16 August 2022).
- Newton, A.C. Forest Ecology and Conservation: A Handbook of Techniques; Oxford University Press: New York, NY, USA, 2007; pp. 85–146. ISBN 9780198567455. [Google Scholar]
- Chao, A. Nonparametric Estimation of the Number of Class in Population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Levins, R. Evolution in Changing Environments; Princeton University Press: Princeton, NJ, USA, 1968; pp. 2–120. ISBN 9780691079592. [Google Scholar]
- PC-Ord Specifications. Available online: https://www.wildblueberrymedia.net/pc-ord-specifications/ (accessed on 22 August 2022).
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.T.; Kim, H.J. Studies on the Seed Characteristics and Viabilities of Six Acer species in Relation to Natural Regeneration in Korea. Korea J. Environ. Ecol. 2011, 25, 358–364. [Google Scholar]
- Edward, E.C.C.; Richard, T.B. Secondary Succession, Gap Dynamics, and Community Structure in a Southern Appalachian Cove Forest. Ecology 1989, 70, 728–735. [Google Scholar]
- Bray, J.R. Gap phase replacement in maple-basswood forest. Ecology 1956, 37, 598–600. [Google Scholar] [CrossRef]
- Kimmins, J.P. Forest Ecology: A Foundation for Sustainable Management; Prentice-Hall: Hoboken, NJ, USA, 1997; p. 596. ISBN 9780023640711. [Google Scholar]
- Benzing, D.H. Vascular Epiphytes: General Biology and Related Biota; Cambridge University Press: Cambridge, MA, USA, 1990; pp. 210–271. ISBN 9780521048958. [Google Scholar]
- Zotz, G.; Hietz, P. The physiological ecology of vascular epiphytes: Current knowledge, open questions. J. Exp. Bot. 2001, 52, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, N.M.; Matelson, T.J. Biomass and nutrient dynamics of epiphytic litterfall in a neotropical cloud forest, Costa Rica. Biotropica 1991, 23, 225–234. [Google Scholar]
- Kramer, K.U.; Green, P.S. The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin, Germany, 1990; Volume 1, pp. 1–14. ISBN 9780947643430. [Google Scholar]
- Page, C.N. Ecological strategies in fern evolution: A neopteridological overview. Bot. Rev. 2002, 68, 345–417. [Google Scholar] [CrossRef]
- Smith, A.R.; Pryer, K.M.; Schuettpelz, E.; Korall, P.; Schneider, H.; Wolf, P.G. A classification for extant ferns. Taxon 2006, 55, 705–731. [Google Scholar] [CrossRef]
- Williams-Linera, G.; Palacios-Rios, M.; Hernández-Gómez, R. Fern richness, tree species surrogacy, and fragment complementarity in a Mexican tropical montane cloud forest. Biodivers. Conserv. 2005, 14, 119–133. [Google Scholar] [CrossRef]
- Kim, J.W.; Lee, Y.G. Classification and Assessment of Plant Communities. World Science: Seoul, Republic of Korea, 2006; pp. 1–240. ISBN 9788958810605. [Google Scholar]
- Park, B.J.; Heo, T.I.; Byeon, J.G.; Cheon, K. Study on Plant Indicator Species of Picea jezoensis (Siebold & Zucc.) Carrière Forest by Topographic Characters—From China (Baekdu-san) to South Korea. J. Environ. Impact Assess. 2022, 31, 388–408. [Google Scholar]
- Langford, A.N.; Buell, M.F. Integration, identity, and stability in the plant association. Adv. Ecol. Res. 1969, 6, 83–135. [Google Scholar]
- Li, G.; Gong, Z.; Li, W. Niches and Interspecifc Associations of Dominant Populations in Three Changed Stages of Natural Secondary Forests on Loess Plateau, P.R. China. Nature 2017, 7, 6604. [Google Scholar]
- Pickett, S.T.A. Population patterns through twenty years of old-field succession. Vegetatio 1982, 19, 45–59. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; McGraw-Hill: New York, NY, USA, 1990; pp. 9–38. ISBN 9780070478299. [Google Scholar]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.; Yang, Y.; Ohsawa, M.; Luu, H.T.; et al. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Ohsawa, M. Tertiary relic deciduous forests on a subtropical mountain, Mt. Emei, Sichuan, China. Folia Geobot. 2002, 37, 93–106. [Google Scholar] [CrossRef]
- Mulch, A.; Chamberlain, C.P. The rise and growth of Tibet. Nature 2006, 439, 670–671. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Q.; Yang, Y.; Ohsawa, M.; Momohara, A.; Hara, M.; Cheng, S.; Fan, S. Population structure of relict Metasequoia glyptostroboides and its habitat fragmentation and degradation in south-central China. Biol. Conserv. 2011, 144, 279–289. [Google Scholar] [CrossRef]
- He, L.Y.; Tang, C.Q.; Wu, Z.L.; Wang, H.C.; Ohsawa, M.; Yan, K. Forest structure and regeneration of the Tertiary relict Taiwania cryptomerioides in the Gaoligong Mountains, Yunnan, southwestern China. Phytocoenologia 2015, 45, 135–156. [Google Scholar] [CrossRef]
- Tzedakis, P.C.; Lawson, I.T.; Frogley, M.R.; Hewitt, G.M.; Preece, R.C. Buffered tree population changes in a Quaternary refugium: Evolutionary implications. Science 2002, 297, 2044–2047. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Willis, K.J. Alpines, trees, and refugia in Europe. Plant Ecol. Divers. 2008, 1, 147–160. [Google Scholar] [CrossRef]
- Keppel, G.; Van Niel, K.P.; Wardell-Johnson, G.W.; Yates, C.J.; Byrne, M.; Mucina, L.; Schut, A.G.; Hopper, S.D.; Franklin, S.E. Refugia: Identifying and understanding safe havens for biodiversity under climate change. Glob. Ecol. Biogeogr. 2012, 21, 393–404. [Google Scholar] [CrossRef]
- Morelli, T.L.; Daly, C.; Dobrowski, S.Z.; Dulen, D.M.; Ebersole, J.L.; Jackson, S.T.; Lundquist, J.D.; Millar, C.I.; Maher, S.P.; Monahan, W.B.; et al. Managing climate change refugia for climate adaptation. PLoS ONE 2016, 11, e0159909. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Noss, R. Viewpoint: Part of a special issue on endemics hotspots, endemism hotspots are linked to stable climatic refugia. Ann. Bot. 2017, 119, 207–214. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Environmental Factor (Number of Groups) | Remark |
|---|---|---|
| Vertical | Altitude (7) | ~1300 m/1300–1400 m/ 1400–1500 m/1500–1600 m/1600–1700 m/1700–1800 m/1800~ |
| Horizontal | Latitude (5) | 35°/36°/37°/41°/42° |
| Habitat (5) | JR/DG/SB/GB/BD | |
| Slope (5) | ~15°/15°–20°/20°–25° /25°–30°/30°~ | |
| Azimuth (8) | N/NE/NW/S SE/SW/W/E |
| Contents (Plots) | * Rock Exposure (%) | * Altitude (m) | * Slope (°) | Azimuth (%) | |||
|---|---|---|---|---|---|---|---|
| N | E | S | W | ||||
| Mt. Gyebang (28) | 40.8 ± 2.3 ab | 1378.1 ± 13.8 c | 28.4 ± 1.3 ab | 20.0 | 36.0 | 20.0 | 24.4 |
| Mt. Sobeak (5) | 56.3 ± 3.7 a | 1345.6 ± 10.8 c | 37.8 ± 4.4 a | 0.0 | 25.0 | 75.0 | 0.0 |
| Mt. Deogyu (8) | 35.6 ± 3.7 ab | 1552.3 ± 10.0 b | 22.4 ± 4.6 b | 12.5 | 25.0 | 0.0 | 62.5 |
| Mt. Jiri (30) | 36.3 ± 1.9 ab | 1731.7 ± 21.6 a | 21.4 ± 1.4 b | 6.7 | 10.0 | 33.3 | 50.0 |
| Mt. Changbai (20) | 24.3 ± 4.9 c | 1282.3 ± 29.4 c | 18.3 ± 2.1 c | 30.0 | 5.0 | 25.0 | 40.0 |
| Braun-Blanquet Scale | Range of Cover and Dominance (%) | Median Value |
|---|---|---|
| 5 | 75–100 | 87.5 |
| 4 | 50–75 | 62.5 |
| 3 | 25–50 | 37.5 |
| 2 | 12.5–25 | 18.75 |
| 1 | <12.5 numerous individuals | 9.375 |
| + | <5 few individuals | 4.69 |
| r | Species represented by a unique individual | 1.01 |
| Contents | Family | Genera | Species | Subspecies | Variety | Form | Total | Ratio (%) |
|---|---|---|---|---|---|---|---|---|
| Pteridophyte | 7 | 11 | 17 | 0 | 1 | 0 | 18 | 8.9 |
| Gymnosperm | 3 | 6 | 7 | 0 | 1 | 0 | 8 | 4.0 |
| Angiosperm | 49 | 110 | 147 | 2 | 24 | 3 | 176 | 87.1 |
| Dicotyledoneae | 45 | 92 | 116 | 2 | 20 | 2 | 140 | 69.3 |
| Monocotyledoneae | 4 | 18 | 31 | 0 | 4 | 1 | 36 | 17.8 |
| Total | 59 | 127 | 171 | 2 | 26 | 3 | 202 | 100.0 |
| Scientific Name | ×1 | ×2 | ×3 | ×4 | ×5 | Mean | Generalist (G) Rank |
|---|---|---|---|---|---|---|---|
| Picea jezoensis (Siebold & Zucc.) Carrière | 0.5357 | 0.7221 | 0.7366 | 0.8844 | 0.8267 | 0.7411 | Target species (T) |
| Acer komarovii Pojark. | 0.5506 | 0.6751 | 0.6935 | 0.9049 | 0.8802 | 0.7409 | I |
| Betula ermanii Cham. | 0.7720 | 0.4972 | 0.7275 | 0.8812 | 0.7288 | 0.7214 | II |
| Asarum sieboldii Miq. | 0.5411 | 0.5381 | 0.8826 | 0.8952 | 0.6440 | 0.7002 | III |
| Lepisorus ussuriensis (Regel & Maack) Ching | 0.5054 | 0.6760 | 0.8921 | 0.6043 | 0.8108 | 0.6977 | IV |
| Acer pseudosieboldianum (Pax) Kom. | 0.6544 | 0.4904 | 0.6903 | 0.9025 | 0.7197 | 0.6915 | V |
| Tripterygium regelii Sprague & Takeda | 0.7049 | 0.4501 | 0.5921 | 0.8652 | 0.8255 | 0.6876 | VI |
| Thelypteris phegopteris (L.) Sloss. | 0.6597 | 0.5766 | 0.5351 | 0.8358 | 0.7782 | 0.6771 | VII |
| Dryopteris expansa (C.Presl) Fraser-Jenk. & Jermy | 0.4433 | 0.6492 | 0.6102 | 0.8299 | 0.8398 | 0.6745 | VIII |
| Sorbus commixta Hedl. | 0.4706 | 0.5892 | 0.6250 | 0.8114 | 0.8250 | 0.6642 | IX |
| Rhododendron schlippenbachii Maxim. | 0.7110 | 0.3748 | 0.5172 | 0.9672 | 0.7425 | 0.6625 | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, B.-J.; Heo, T.-I.; Cheon, K.-I. Analyzing Generalist Plant Species Using Topographic Characteristics of Picea jezoensis (Siebold & Zucc.) Carrière Forests in East Asia: From China (Mt. Changbai) to South Korea. Int. J. Plant Biol. 2024, 15, 320-339. https://doi.org/10.3390/ijpb15020027
Park B-J, Heo T-I, Cheon K-I. Analyzing Generalist Plant Species Using Topographic Characteristics of Picea jezoensis (Siebold & Zucc.) Carrière Forests in East Asia: From China (Mt. Changbai) to South Korea. International Journal of Plant Biology. 2024; 15(2):320-339. https://doi.org/10.3390/ijpb15020027
Chicago/Turabian StylePark, Byeong-Joo, Tae-Im Heo, and Kwang-Il Cheon. 2024. "Analyzing Generalist Plant Species Using Topographic Characteristics of Picea jezoensis (Siebold & Zucc.) Carrière Forests in East Asia: From China (Mt. Changbai) to South Korea" International Journal of Plant Biology 15, no. 2: 320-339. https://doi.org/10.3390/ijpb15020027