
Mapping of a Major-Effect Quantitative Trait Locus for Seed Dormancy in Wheat
Abstract
:1. Introduction
2. Results
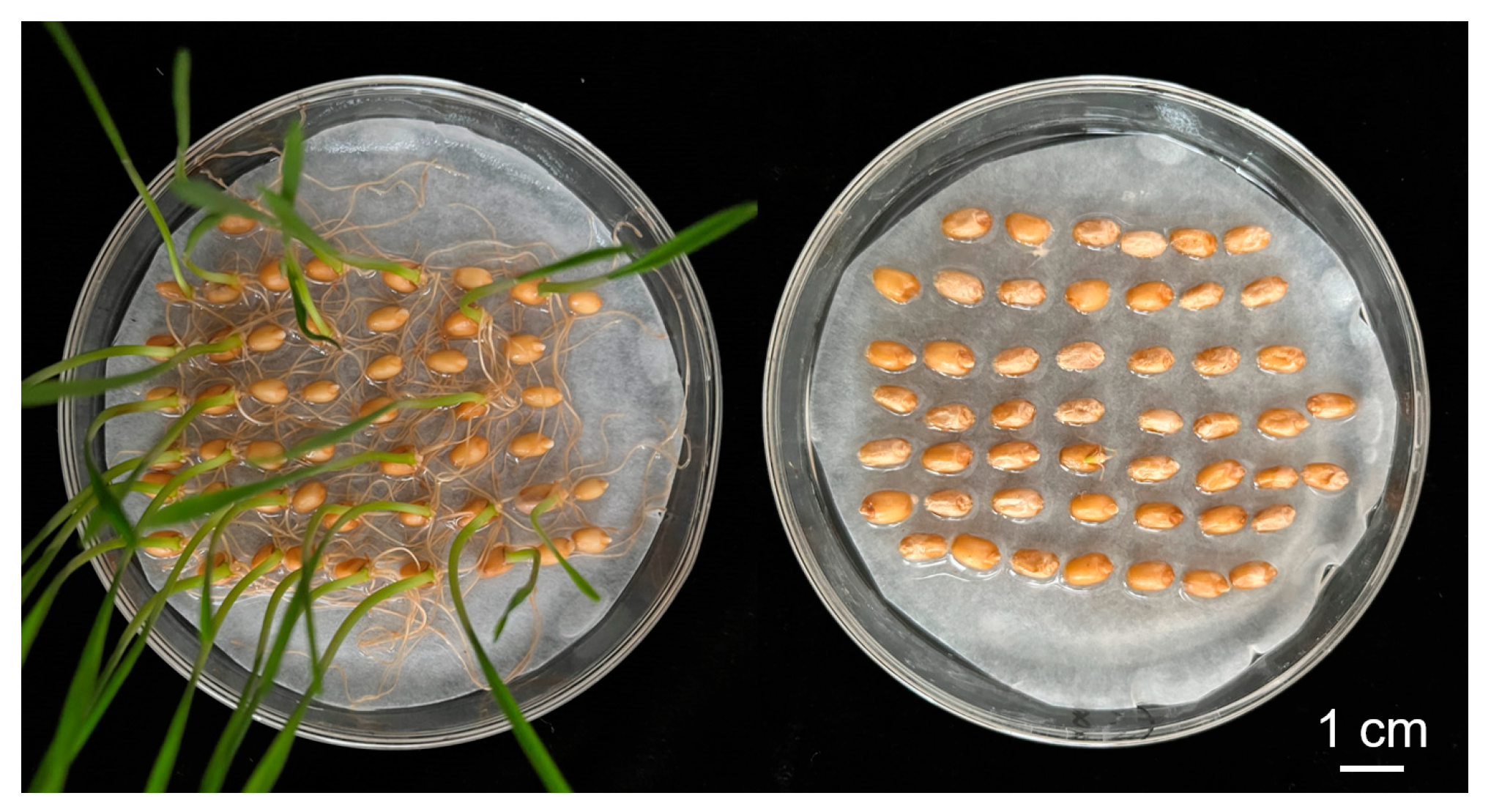
2.1. Assessment of Seed Dormancy for SY and CH
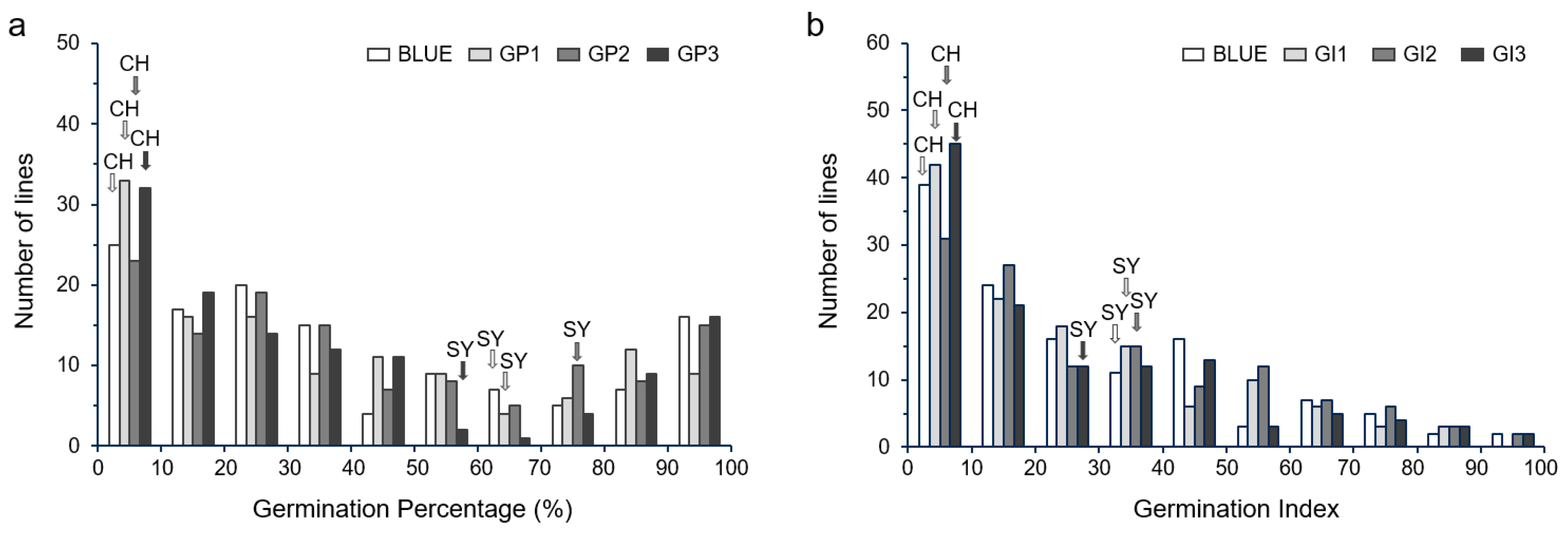
2.2. Phenotypic Variance of GP and GI in the SY × CH RILs
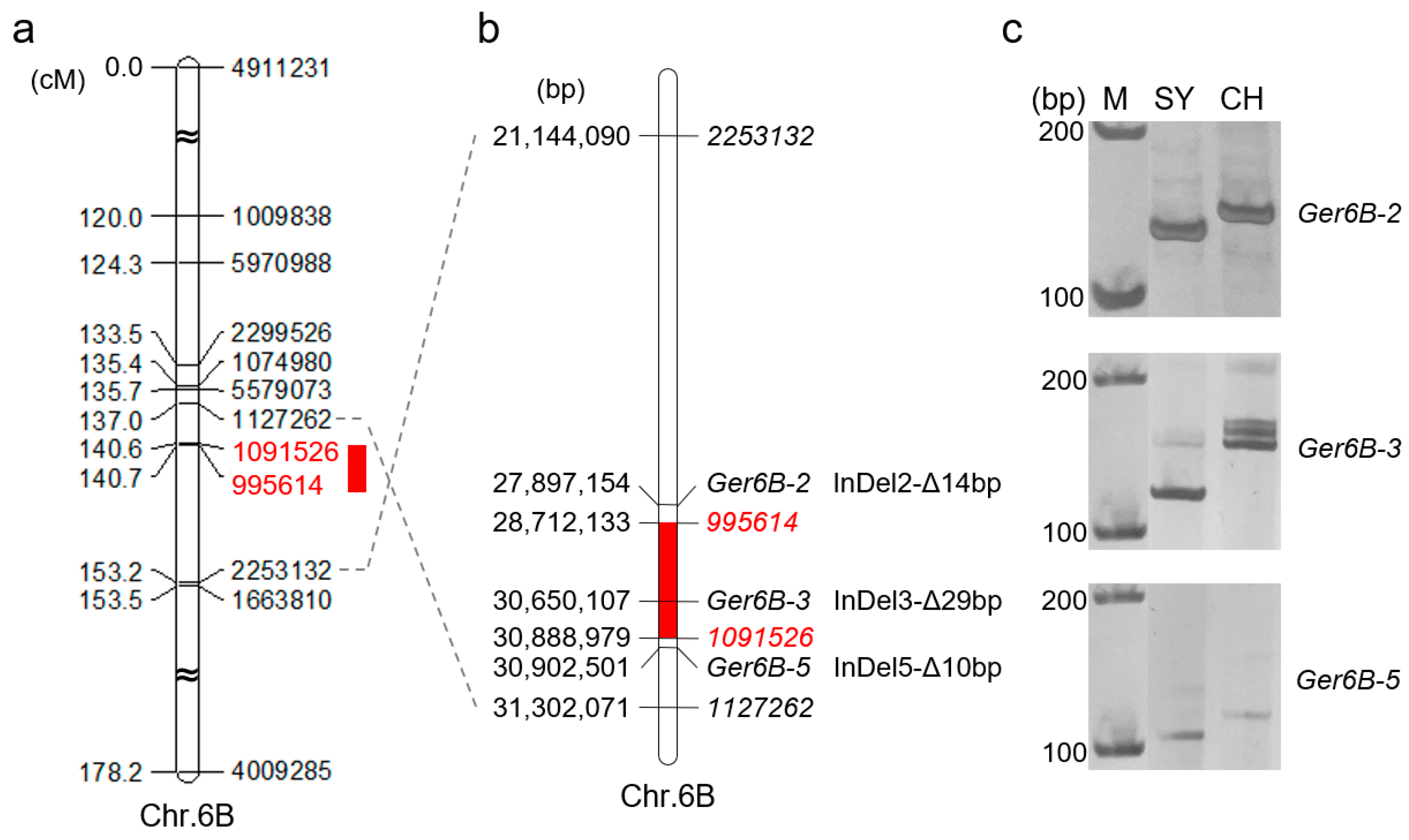
2.3. Genetic Mapping of GP and GI
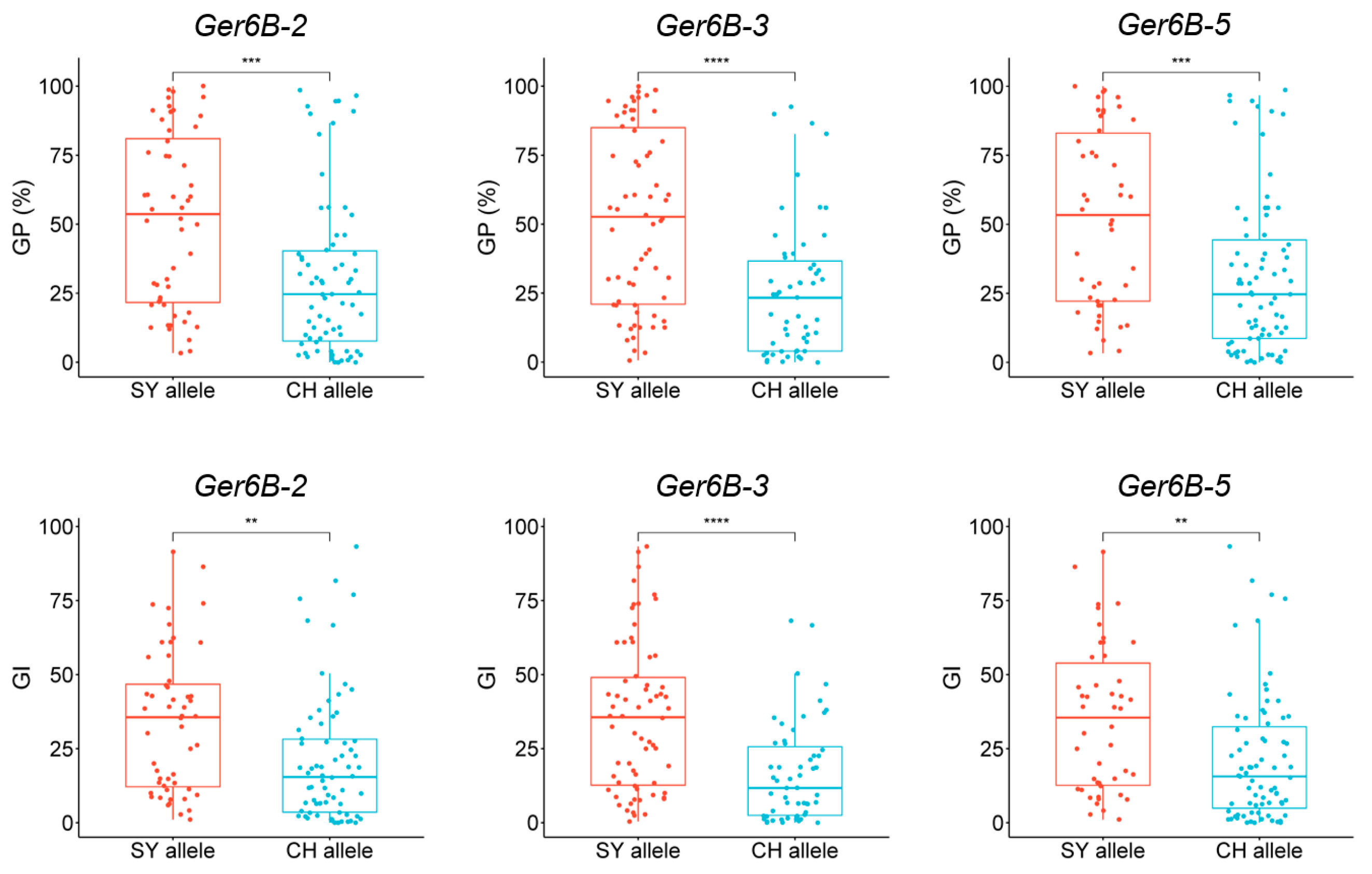
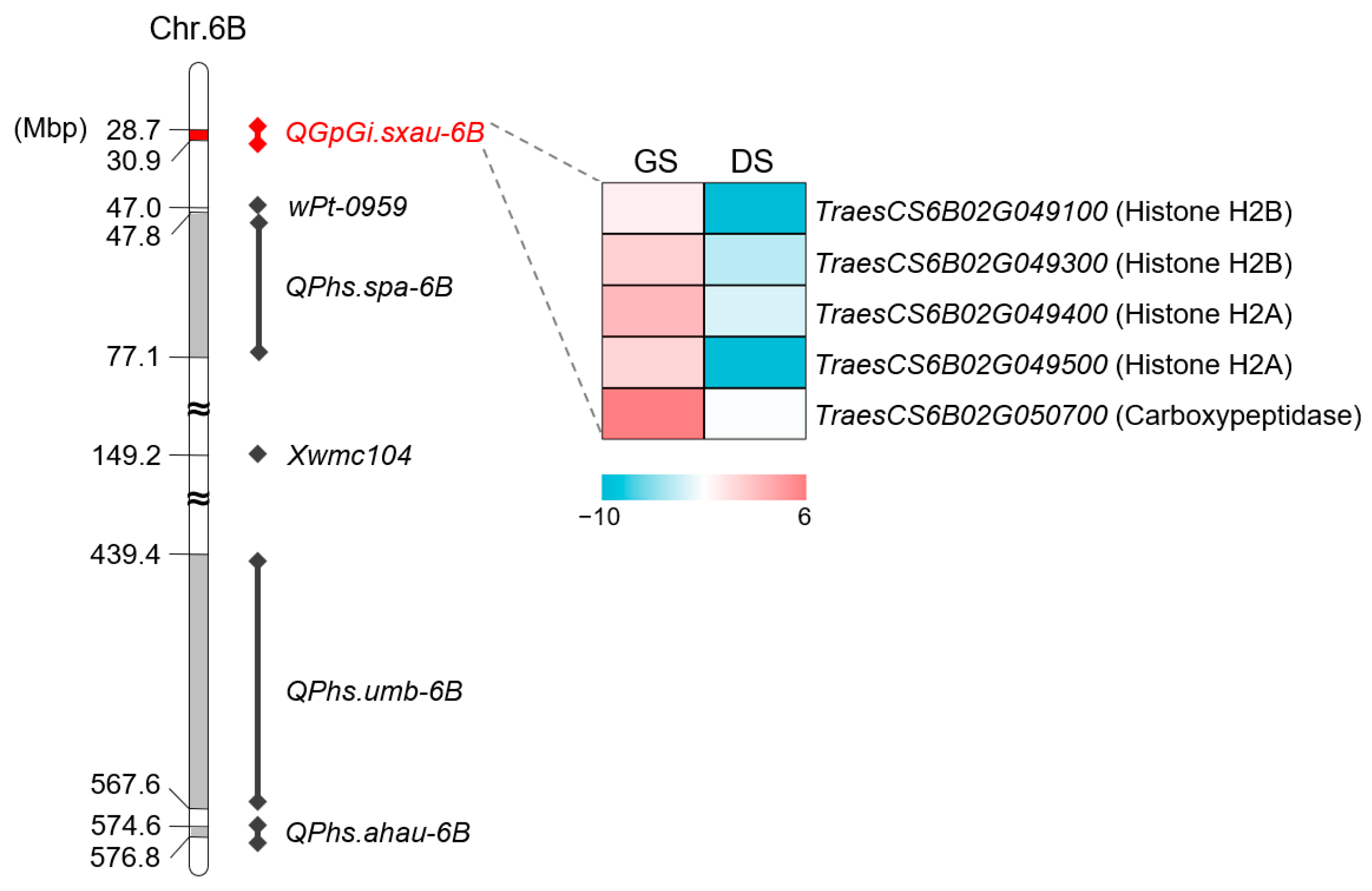
2.4. Verification of QGp/Gi.sxau-6B
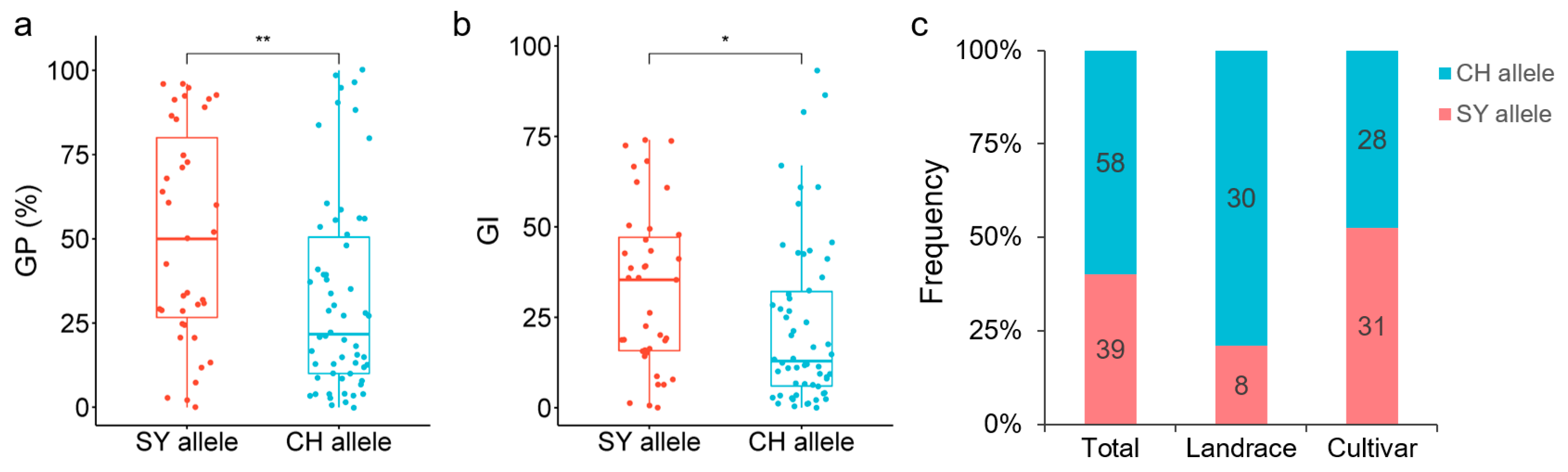
2.5. Distribution of QGp/Gi.sxau-6B Alleles in Wheat
3. Discussion
3.1. CH1539 Can Be Used to Improve the Resistance of Cultivars to PHS
3.2. QGp/Gi.sxau-6B Is a Novel Dormancy Locus
3.3. Prediction of Causing Gene for QGp/Gi.sxau-6B
4. Materials and Methods
4.1. Plant Materials
4.2. Seed Dormancy Test
4.3. Genotyping and QTL Mapping
4.4. Diagnostic Marker Developing
4.5. Analysis of QTL Alleles in Germplasms
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gao, F.; Ayele, B.T. Functional Genomics of Seed Dormancy in Wheat: Advances and Prospects. Front. Plant Sci. 2014, 5, 458. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global Food Demand and the Sustainable Intensification of Agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, R.; Spannagl, M.; Pfeifer, M.; Barker, G.L.; D’Amore, R.; Allen, A.M.; McKenzie, N.; Kramer, M.; Kerhornou, A.; Bolser, D.; et al. Analysis of the Bread Wheat Genome Using Whole-genome Shotgun Sequencing. Nature 2012, 491, 705–710. [Google Scholar] [CrossRef]
- Himi, E.; Mares, D.J.; Yanagisawa, A.; Noda, K. Effect of Grain Colour Gene (R) on Grain Dormancy and Sensitivity of the Embryo to Abscisic Acid (ABA) in Wheat. J. Exp. Bot. 2002, 53, 1569–1574. [Google Scholar] [CrossRef]
- Himi, E.; Maekawa, M.; Miura, H.; Noda, K. Development of PCR Markers for Tamyb10 Related to R-1, Red Grain Color Gene in Wheat. Theor. Appl. Genet. 2011, 122, 1561–1576. [Google Scholar] [CrossRef] [PubMed]
- Lang, J.; Fu, Y.; Zhou, Y.; Cheng, M.; Deng, M.; Li, M.; Zhu, T.; Yang, J.; Guo, X.; Gui, L.; et al. Myb10-D Confers PHS-3D Resistance to Pre-harvest Sprouting by Regulating NCED in ABA Biosynthesis Pathway of Wheat. New Phytol. 2021, 230, 1940–1952. [Google Scholar] [CrossRef]
- Bentsink, L.; Jowett, J.; Hanhart, C.J.; Koornneef, M. Cloning of DOG1, a Quantitative Trait Locus Controlling Seed Dormancy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 17042–17047. [Google Scholar] [CrossRef] [PubMed]
- Ashikawa, I.; Abe, F.; Nakamura, S. Ectopic Expression of Wheat and Barley DOG1-Like Genes Promotes Seed Dormancy in Arabidopsis. Plant Sci. 2010, 179, 536–542. [Google Scholar] [CrossRef]
- Xi, W.; Liu, C.; Hou, X.; Yu, H. MOTHER OF FT AND TFL1 Regulates Seed Germination Through a Negative Feedback Loop Modulating ABA Signaling in Arabidopsis. Plant Cell 2010, 22, 1733–1748. [Google Scholar] [CrossRef]
- Nakamura, S.; Abe, F.; Kawahigashi, H.; Nakazono, K.; Tagiri, A.; Matsumoto, T.; Utsugi, S.; Ogawa, T.; Handa, H.; Ishida, H.; et al. A Wheat Homolog of MOTHER OF FT AND TFL1 Acts in the Regulation of Germination. Plant Cell 2011, 23, 3215–3229. [Google Scholar] [CrossRef]
- Torada, A.; Koike, M.; Ogawa, T.; Takenouchi, Y.; Tadamura, K.; Wu, J.; Matsumoto, T.; Kawaura, K.; Ogihara, Y. A Causal Gene for Seed Dormancy on Wheat Chromosome 4A Encodes a MAP Kinase Kinase. Curr. Biol. 2016, 26, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Cavanagh, C.; Verbyla, K.L.; Tibbits, J.F.; Verbyla, A.P.; Huang, B.E.; Rosewarne, G.M.; Stephen, S.; Wang, P.; Whan, A.; et al. Transcriptomic Analysis of Wheat Near-isogenic Lines Identifies PM19-A1 and A2 as Candidates for a Major Dormancy QTL. Genome Biol. 2015, 16, 93. [Google Scholar] [CrossRef]
- Zhang, Y.; Miao, X.; Xia, X.; He, Z. Cloning of Seed Dormancy Genes (TaSdr) Associated with Tolerance to Pre-harvest Sprouting in Common Wheat and Development of a Functional Marker. Theor. Appl. Genet. 2014, 127, 855–866. [Google Scholar] [CrossRef]
- Penning, B.W. Gene Expression Differences Related to Pre-harvest Sprouting Uncovered in Related Wheat Varieties by RNAseq Analysis. Plant Gene 2023, 33, 100404. [Google Scholar] [CrossRef]
- Park, S.Y.; Jung, W.J.; Bang, G.; Hwang, H.; Kim, J.Y. Transcriptome and Proteome Co-Profiling Offers an Understanding of Pre-Harvest Sprouting (PHS) Molecular Mechanisms in Wheat (Triticum aestivum). Plants 2022, 11, 2807. [Google Scholar] [CrossRef] [PubMed]
- Tuan, P.A.; Yamasaki, Y.; Kanno, Y.; Seo, M.; Ayele, B.T. Transcriptomics of Cytokinin and Auxin Metabolism and Signaling Genes during Seed Maturation in Dormant and Non-dormant Wheat Genotypes. Sci. Rep. 2019, 9, 3983. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, G.; Bai, X.; Ye, Z.; Zhang, S.; Xie, K.; Sun, F.; Zhang, C.; Xi, Y. Regulatory Network of Preharvest Sprouting Resistance Revealed by Integrative Analysis of mRNA, Noncoding RNA, and DNA Methylation in Wheat. J. Agric. Food Chem. 2021, 69, 4018–4035. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.; Kamble, U.R.; Gupta, V.; Singh, G.; Sheoran, S.; Gupta, A.; Tyagi, B.S.; Kumar, P.; Mishra, C.N.; Krishannapa, G.; et al. Pre-harvest Sprouting in Wheat: Current Ctatus and Future Prospects. J. Cereal Res. 2021, 13, 1–22. [Google Scholar] [CrossRef]
- Chang, C.; Zhang, H.; Lu, J.; Si, H.; Ma, C. Genetic Improvement of Wheat with Pre-Harvest Sprouting Resistance in China. Genes 2023, 14, 837. [Google Scholar] [CrossRef]
- Roy, J.; Prasad, M.; Varshney, R.; Balyan, H.; Blake, T.; Dhaliwal, H.; Singh, H.; Edwards, K.; Gupta, P. Identification of a Microsatellite on Chromosomes 6B and a STS on 7D of Bread Wheat Showing an Association with Preharvest Sprouting Tolerance. Theor. Appl. Genet. 1999, 99, 336–340. [Google Scholar] [CrossRef]
- Rehman Arif, M.A.; Neumann, K.; Nagel, M.; Kobiljski, B.; Lohwasser, U.; Börner, A. An Association Mapping Analysis of Dormancy and Pre-harvest Sprouting in Wheat. Euphytica 2012, 188, 409–417. [Google Scholar] [CrossRef]
- Kumar, S.; Knox, R.E.; Clarke, F.R.; Pozniak, C.J.; DePauw, R.M.; Cuthbert, R.D.; Fox, S. Maximizing the Identification of QTL for Pre-harvest Sprouting Resistance Using Seed Dormancy Measures in a White-grained Hexaploid Wheat Population. Euphytica 2015, 205, 287–309. [Google Scholar] [CrossRef]
- Liton, U.A.; McCartney, C.A.; Hiebert, C.W.; Kumar, S.; Jordan, M.C.; Ayele, B.T. Identification of Loci for Pre-harvest Sprouting Resistance in the Highly Dormant Spring Wheat RL4137. Theor. Appl. Genet. 2021, 134, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, S.; Wei, W.; Xie, H.; Liu, K.; Zhang, C.; Wu, Z.; Jiang, H.; Cao, J.; Zhao, L.; et al. Genome-wide Association Study of Pre-harvest Sprouting Tolerance Using a 90K SNP Array in Common Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2019, 132, 2947–2963. [Google Scholar] [CrossRef] [PubMed]
- Sheng, D.; Qiao, L.; Zhang, X.; Li, X.; Chang, L.; Guo, H.; Zhang, S.; Chen, F.; Chang, Z. Fine Mapping of a Recessive Leaf Rust Resistance Locus on Chromosome 2BS in Wheat Accession CH1539. Mol. Breed. 2022, 42, 52. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Jia, X.; Xiang, Y.; Jiang, W. Histone Modification and Chromatin Remodeling during the Seed Life Cycle. Front. Plant Sci. 2022, 13, 865361. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, P.; Dong, Z. The Role of Histone Acetylation in Transcriptional Regulation and Seed Development. Plant Physiol. 2023. [Google Scholar] [CrossRef]
- Tognacca, R.S.; Botto, J.F. Post-transcriptional Regulation of Seed Dormancy and Germination: Current Understanding and Future Directions. Plant Commun. 2021, 2, 100169. [Google Scholar] [CrossRef]
- Yang, W.; Chen, Z.; Huang, Y.; Chang, G.; Li, P.; Wei, J.; Yuan, X.; Huang, J.; Hu, X. Powerdress as the Novel Regulator Enhances Arabidopsis Seeds Germination Tolerance to High Temperature Stress by Histone Modification of SOM Locus. Plant Sci. 2019, 284, 91–98. [Google Scholar] [CrossRef]
- Liu, Y.; Koornneef, M.; Soppe, W.J. The Absence of Histone H2B Monoubiquitination in the Arabidopsis Hub1 (rdo4) Mutant Reveals a Role for Chromatin Remodeling in Seed Dormancy. Plant Cell 2007, 19, 433–444. [Google Scholar] [CrossRef]
- Jiang, D.; Borg, M.; Lorkovi, Z.J.; Montgomery, S.A.; Osakabe, A.; Yelagandula, R.; Axelsson, E.; Berger, F. The Evolution and Functional Divergence of the Histone H2B Family in Plants. PLoS Genet. 2020, 16, e1008964. [Google Scholar] [CrossRef] [PubMed]
- Khadka, J.; Pesok, A.; Grafi, G. Plant Histone HTB (H2B) Variants in Regulating Chromatin Structure and Function. Plants 2020, 9, 1435. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tang, L.; Qiu, J.; Zhang, W.; Wang, Y.; Tong, X.; Wei, X.; Hou, Y.; Zhang, J. Serine Carboxypeptidase 46 Regulates Grain Filling and Seed Germination in Rice (Oryza sativa L.). PLoS ONE 2016, 11, e0159737. [Google Scholar] [CrossRef] [PubMed]
- Dal Degan, F.; Rocher, A.; Cameron-Mills, V.; von Wettstein, D. The Expression of Serine Carboxypeptidases during Maturation and Germination of the Barley Grain. Proc. Natl. Acad. Sci. USA 1994, 91, 8209–8213. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, F.; Cejudo, F.J. Patterns of Starchy Endosperm Acidification and Protease Gene Expression in Wheat Grains Following Germination. Plant Physiol. 1999, 119, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; González, M.C.; Cejudo, F.J. A Germination-related Gene Encoding a Serine Carboxypeptidase is Expressed During the Differentiation of the Vascular Tissue in Wheat Grains and Seedlings. Planta 2002, 215, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Mendoza, M.; Diaz, I.; Martinez, M. Insights on the Proteases Involved in Barley and Wheat Grain Germination. Int. J. Mol. Sci. 2019, 20, 2087. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, L.; Liu, Y.; Shen, X.; Guo, Y.; Ma, X.; Zhang, X.; Li, X.; Cheng, T.; Wen, H.; et al. RNA-Seq-Based WGCNA and Association Analysis Reveal the Key Regulatory Module and Genes Responding to Salt Stress in Wheat Roots. Plants 2024, 13, 274. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Zhang, X.; Li, X.; Yang, Z.; Li, R.; Jia, J.; Yan, L.; Chang, Z. Genetic Incorporation of Genes for the Optimal Plant Architecture in Common Wheat. Mol. Breed. 2022, 42, 66. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Zhang, X.; Li, X.; Zhang, L.; Zheng, J.; Chang, Z. Development of NBS-related Microsatellite (NRM) Markers in Hexaploid Wheat. Euphytica 2017, 213, 256. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Repeat | Parents | RIL Population | ||||
|---|---|---|---|---|---|---|---|
| SY | CH | Min | Max | Mean | CV | ||
| GP (%) | 1 | 66.54 | 0 | 0 | 100.00 | 38.11 | 0.82 |
| 2 | 70.21 | 2.17 | 0 | 100.00 | 43.06 | 0.75 | |
| 3 | 52.42 | 0 | 0 | 100.00 | 38.22 | 0.88 | |
| BLUE | 63.06 | 0.72 *** | 0 | 100.00 | 39.80 | 0.82 | |
| GI | 1 | 33.74 | 0 | 0 | 89.14 | 25.12 | 0.91 |
| 2 | 34.40 | 0.62 | 0 | 95.14 | 30.06 | 0.83 | |
| 3 | 28.30 | 0 | 0 | 98.00 | 25.17 | 0.97 | |
| BLUE | 32.15 | 0.21 **** | 0 | 94.10 | 26.78 | 0.90 | |
| Trait | Chromosome | Position (cM) | Left Marker | Right Marker | LOD | PVE (%) | ADD |
|---|---|---|---|---|---|---|---|
| GP | 5A | 138 | 996462 | 3064715 | 2.71 | 5.78 | 4.98 |
| 6B | 141 | 995614 | 1091526 | 6.30 | 17.66 | –13.78 | |
| GI | 5A | 140 | 3064715 | 1207181 | 3.32 | 6.41 | 2.82 |
| 6B | 141 | 995614 | 1091526 | 11.84 | 34.11 | –17.14 | |
| 6D | 87 | 1239681 | 12736348 | 2.83 | 5.06 | –2.48 | |
| 7A | 17 | 3064775 | 1230235 | 5.66 | 9.02 | 11.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Qiao, L.; Mei, C.; Nong, L.; Li, Q.; Zhang, X.; Li, R.; Gao, W.; Chen, F.; Chang, L.; et al. Mapping of a Major-Effect Quantitative Trait Locus for Seed Dormancy in Wheat. Int. J. Mol. Sci. 2024, 25, 3681. https://doi.org/10.3390/ijms25073681
Gao Y, Qiao L, Mei C, Nong L, Li Q, Zhang X, Li R, Gao W, Chen F, Chang L, et al. Mapping of a Major-Effect Quantitative Trait Locus for Seed Dormancy in Wheat. International Journal of Molecular Sciences. 2024; 25(7):3681. https://doi.org/10.3390/ijms25073681
Chicago/Turabian StyleGao, Yu, Linyi Qiao, Chao Mei, Lina Nong, Qiqi Li, Xiaojun Zhang, Rui Li, Wei Gao, Fang Chen, Lifang Chang, and et al. 2024. "Mapping of a Major-Effect Quantitative Trait Locus for Seed Dormancy in Wheat" International Journal of Molecular Sciences 25, no. 7: 3681. https://doi.org/10.3390/ijms25073681