
Acute Impacts of Ionizing Radiation Exposure on the Gastrointestinal Tract and Gut Microbiome in Mice
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
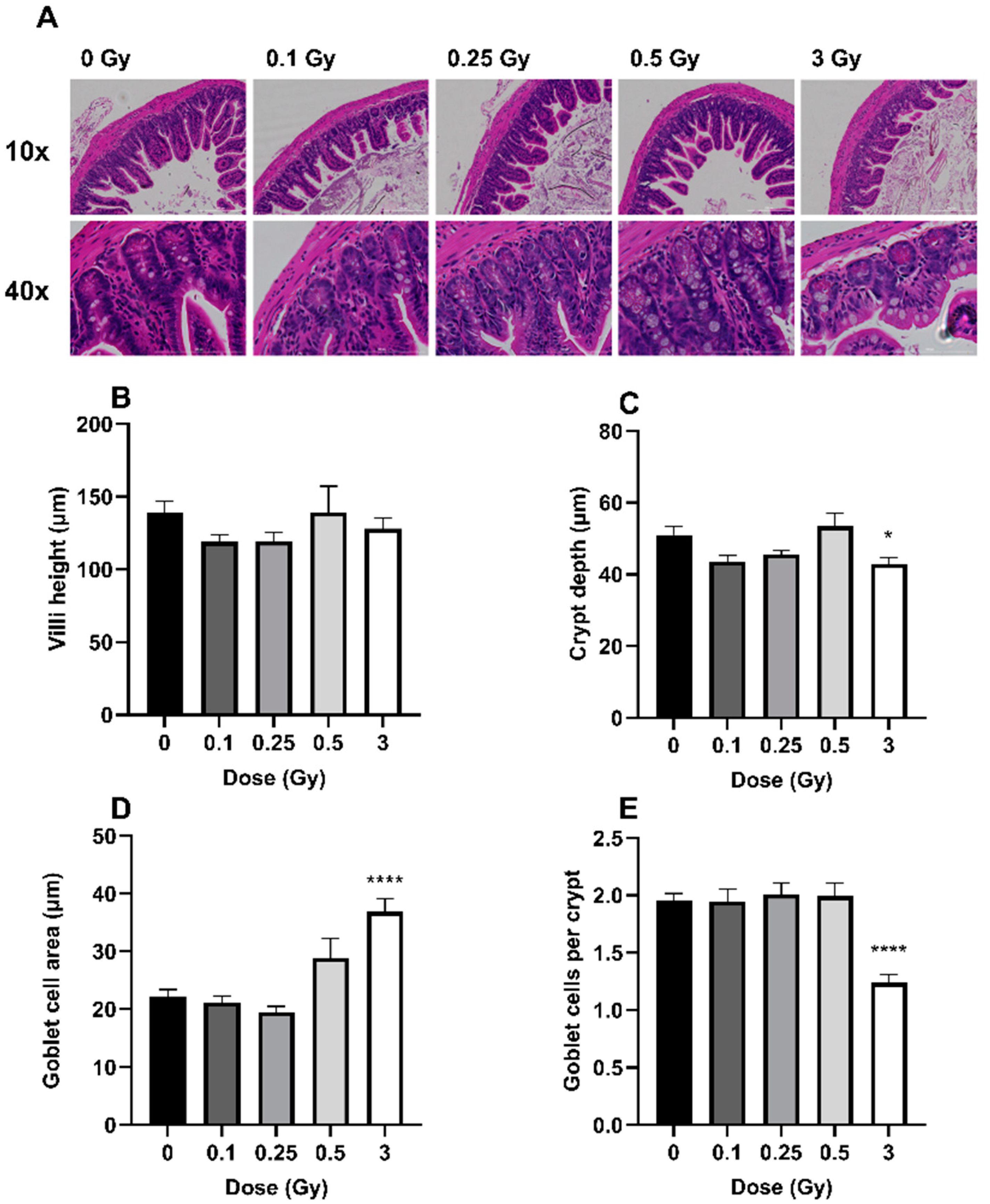
2.1. The Effects of Radiation on GIT Histology
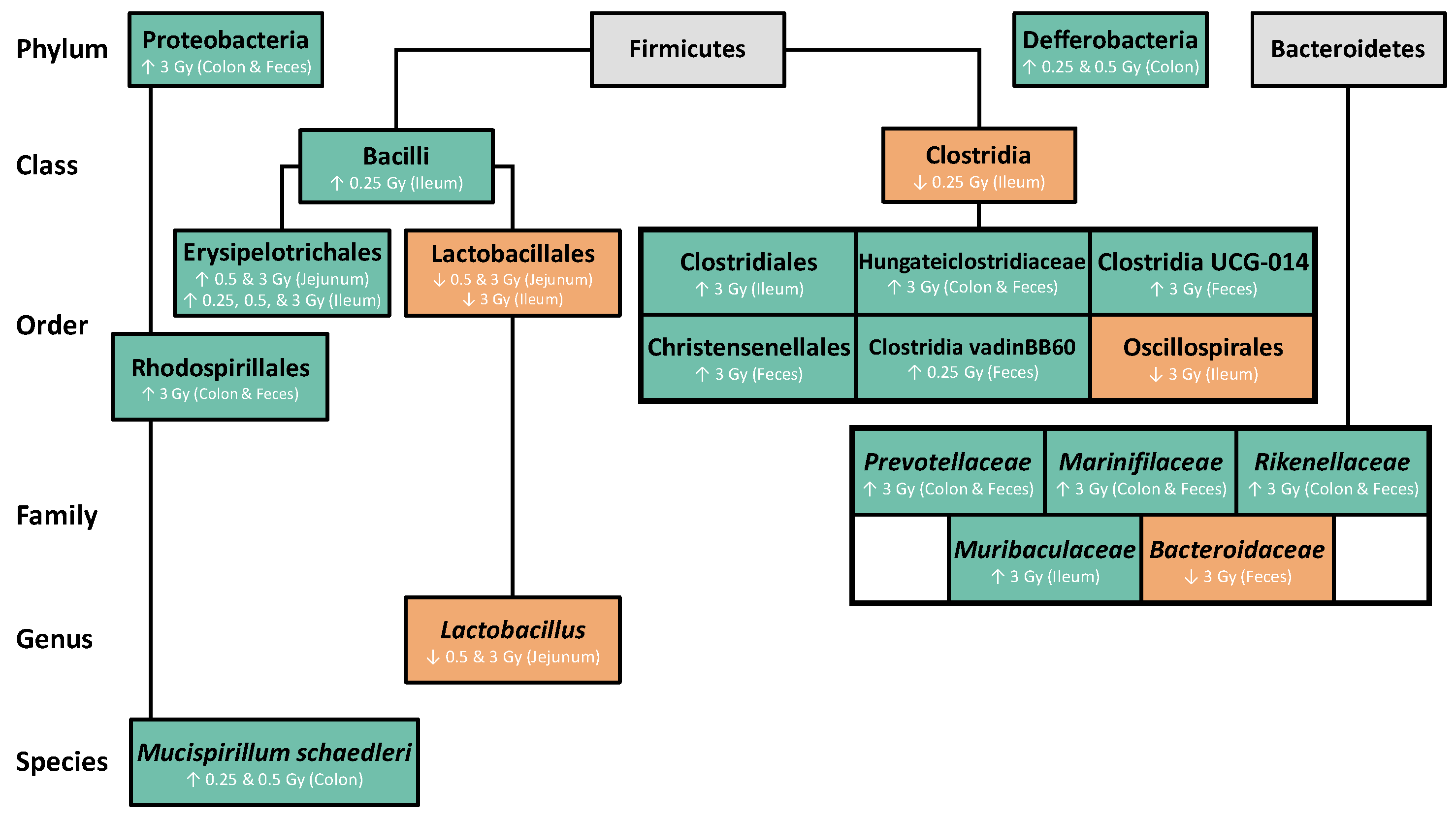
2.2. The Effects of Radiation on the GIT Microbiome
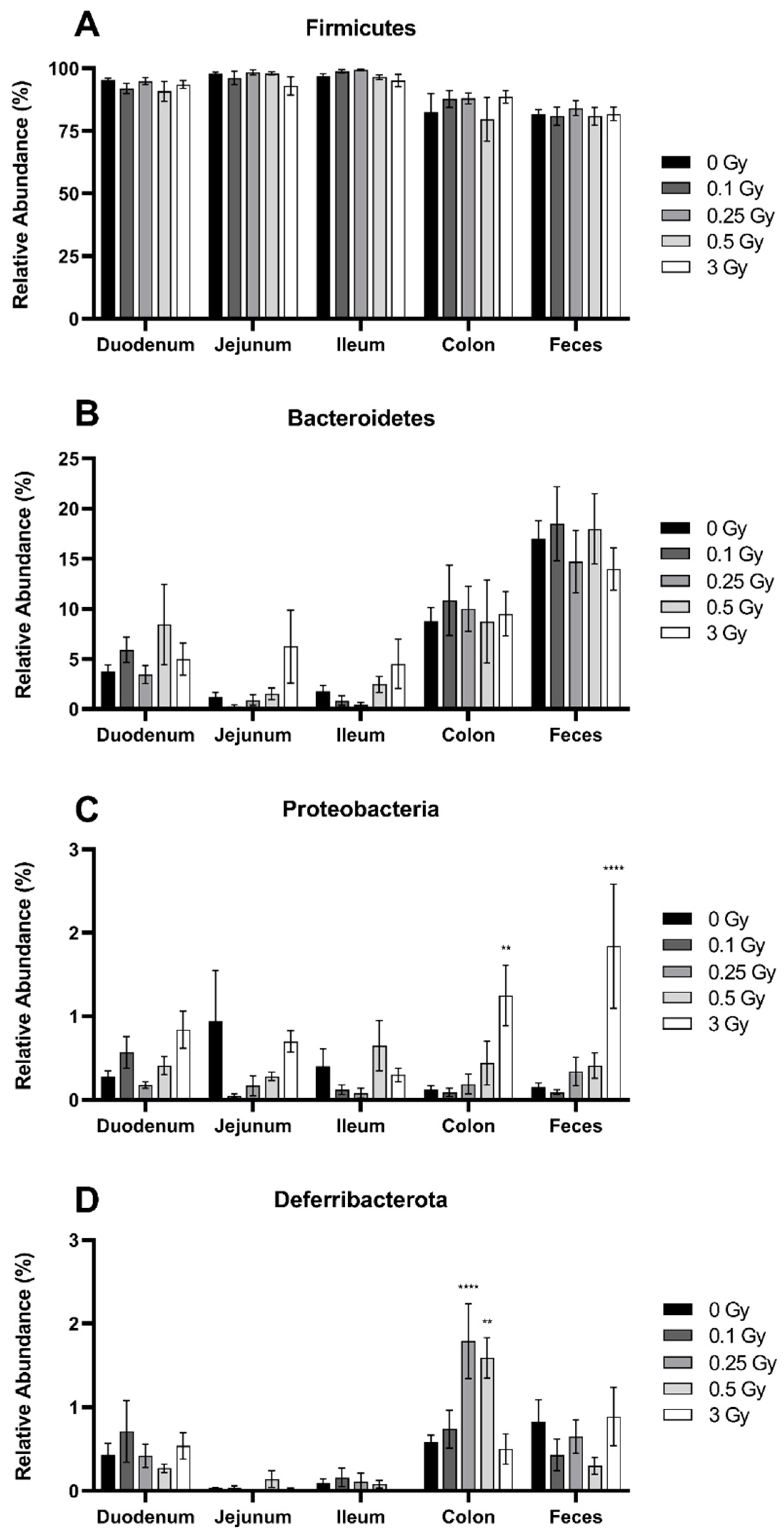
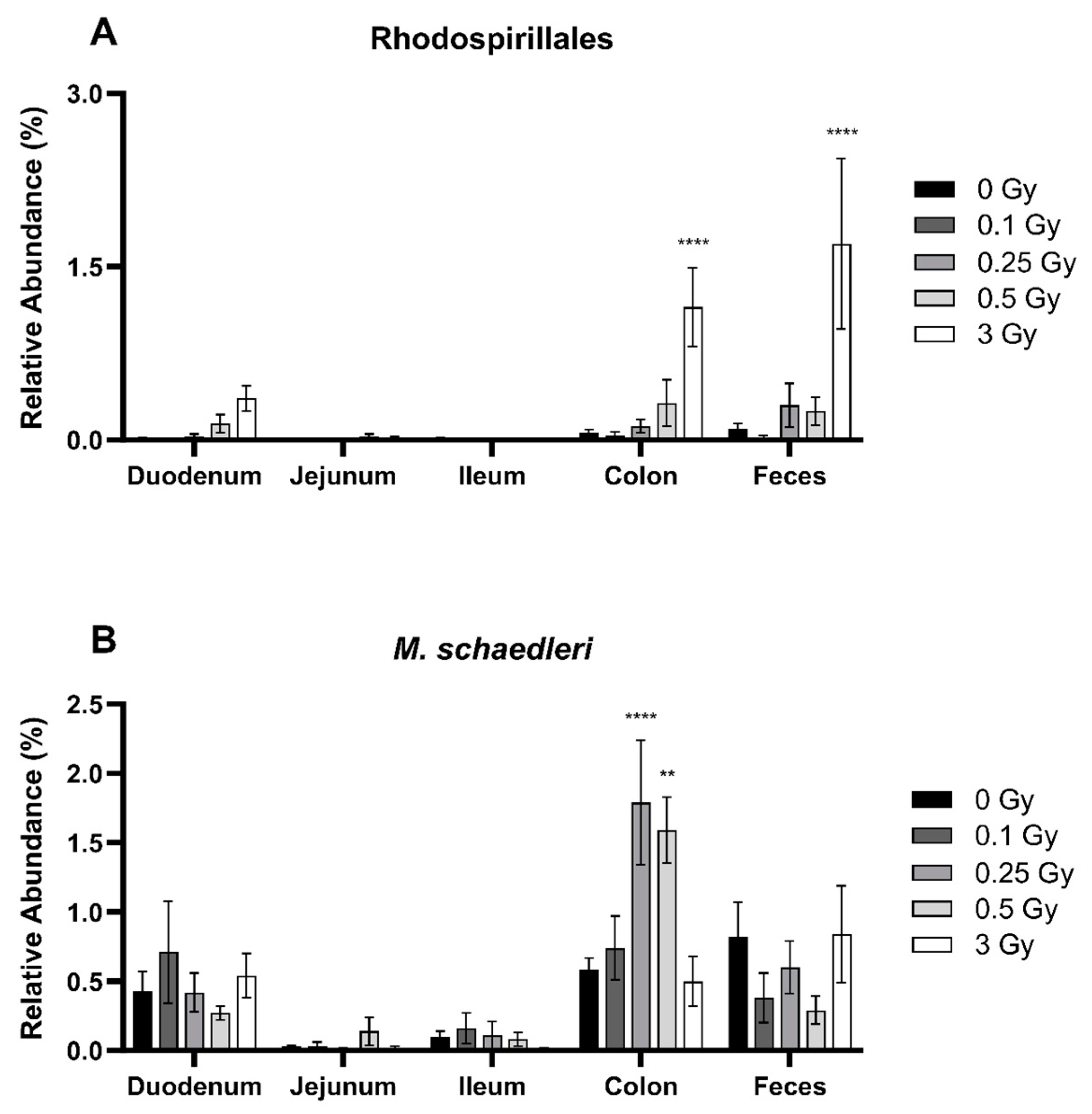
2.2.1. Radiation Increased the Abundance of Proteobacteria and Deferribacterota in the GIT
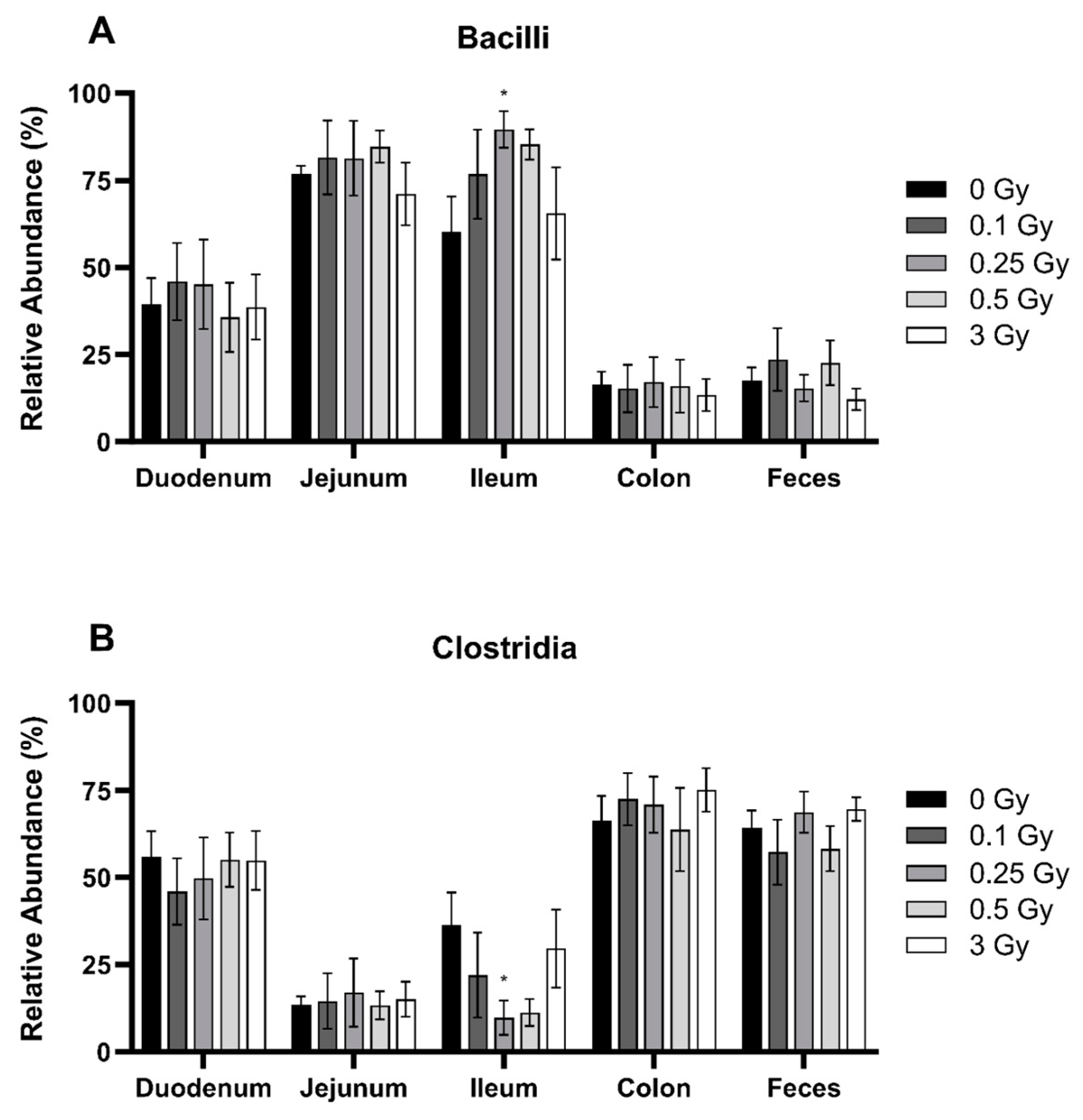
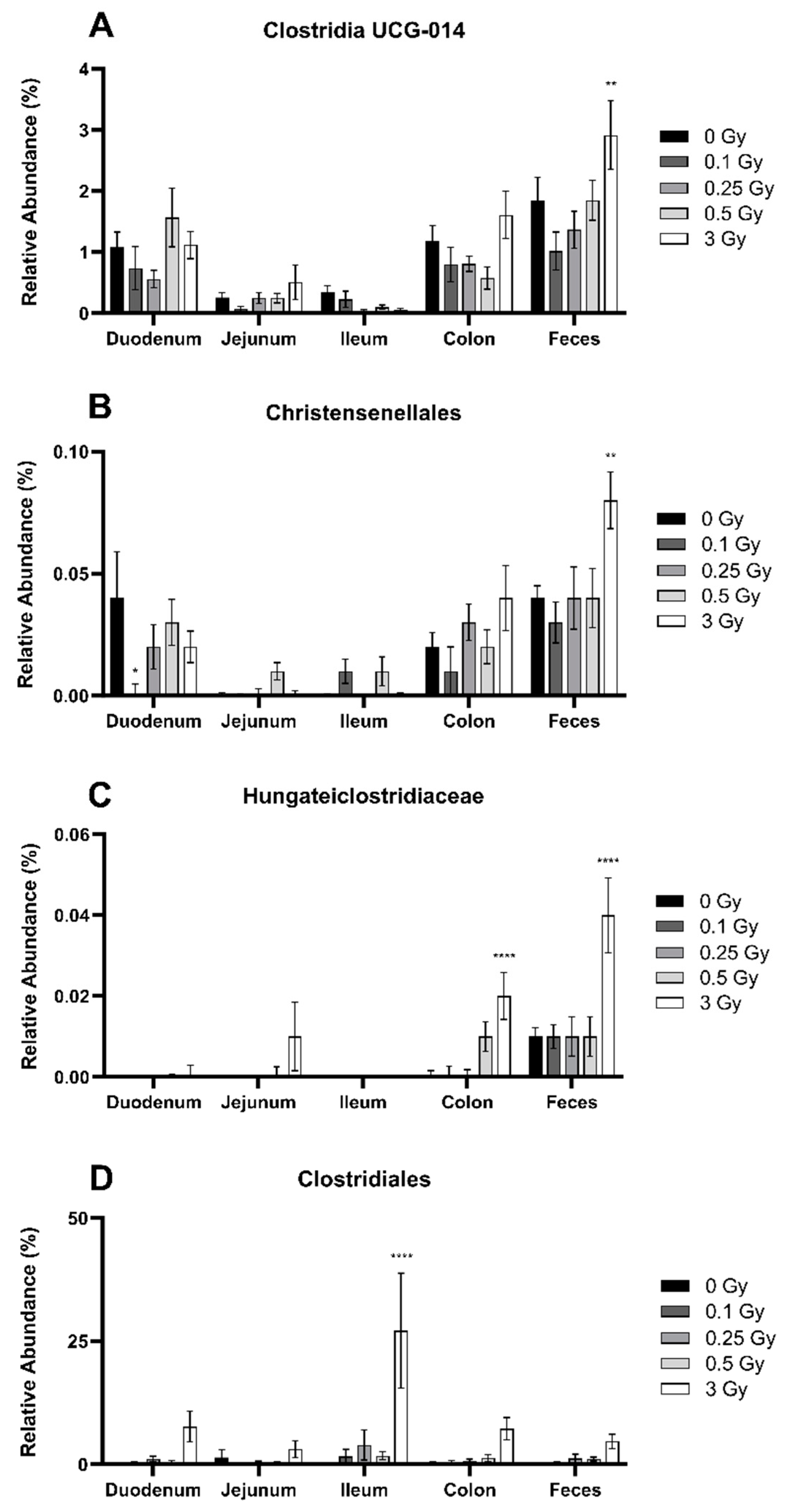
2.2.2. Radiation Altered the Distribution of Firmicutes in the GIT
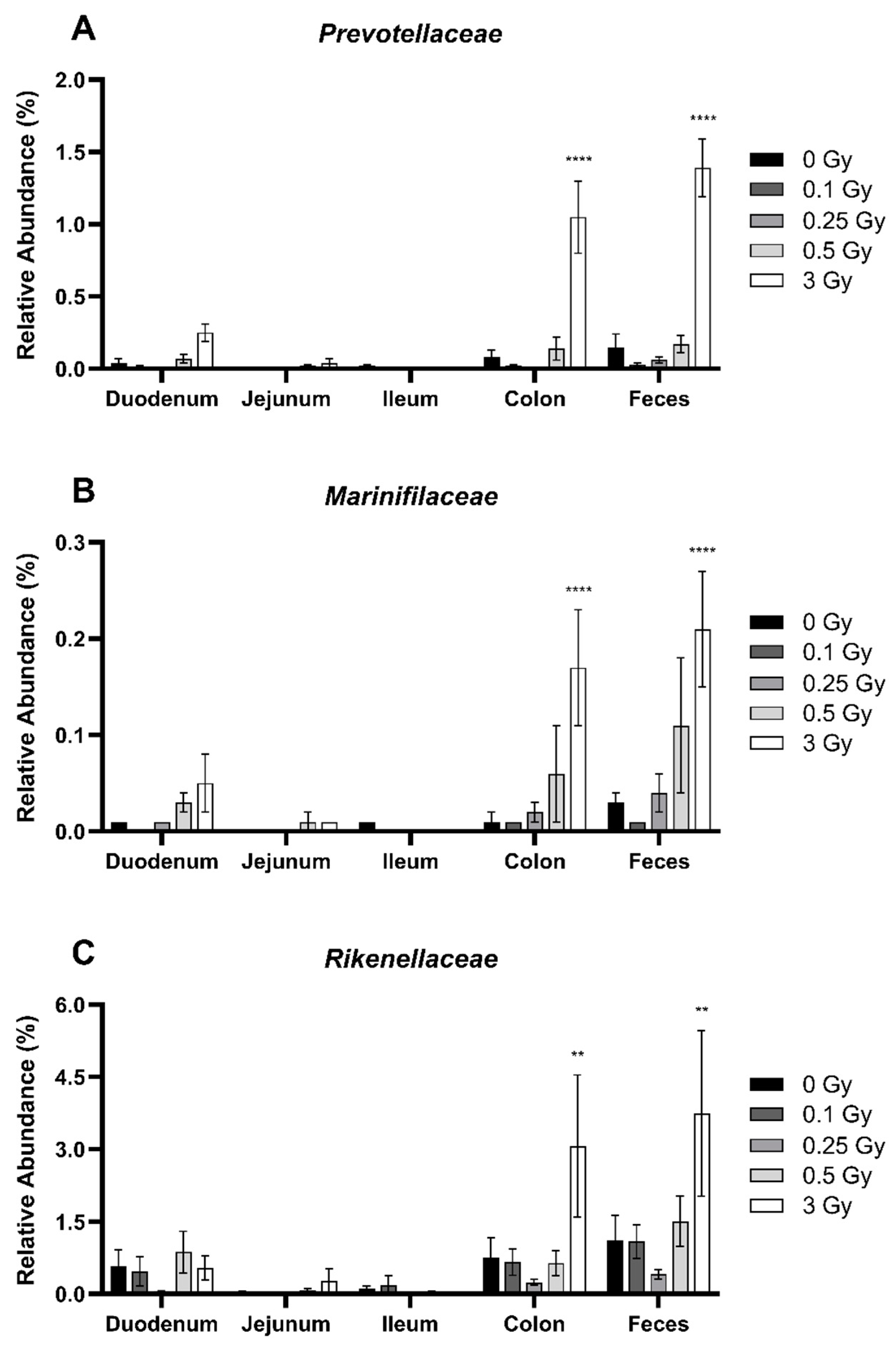
2.2.3. Radiation Altered the Distribution of Bacteroidetes in the GIT
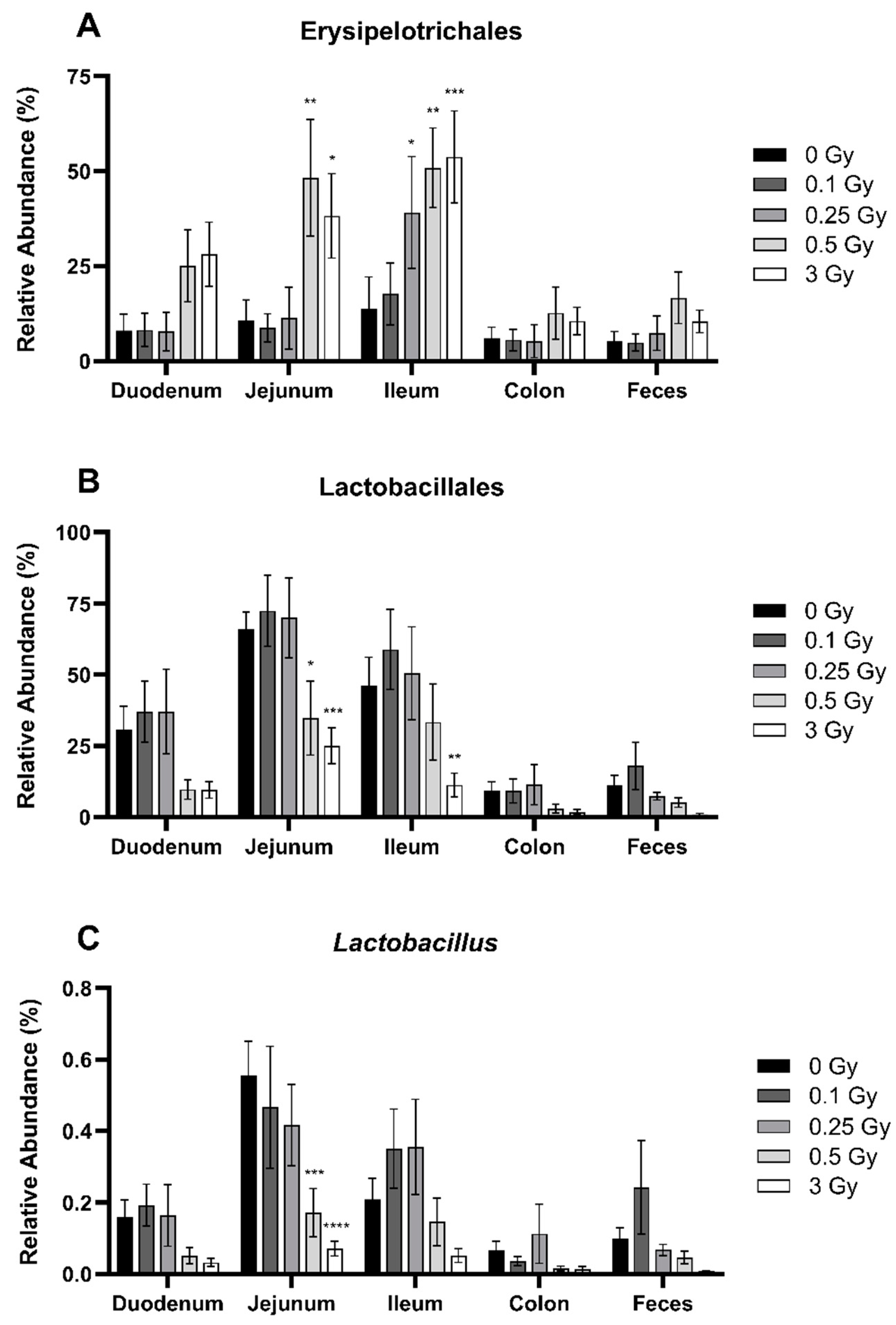
2.2.4. Radiation Altered the Distribution of Select Members of Proteobacteria and Deferribacterota Phyla in the GIT
3. Discussion
4. Materials and Methods
4.1. Animal Handling
4.2. Irradiations
4.3. Tissue Collection
4.4. Histological Analysis
4.5. 16S Metagenomics Microbiome Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaffray, D.A.; Gospodarowicz, M.K. Radiation Therapy for Cancer. In Cancer: Disease Control Priorities; Gelband, H., Jha, P., Sankaranarayanan, R., Horton, S., Eds.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2015; Volume 3, pp. 142–146. ISBN 9781464803499. [Google Scholar]
- McCaughan, H.; Boyle, S.; McGoran, J.J. Update on the Management of the Gastrointestinal Effects of Radiation. World J. Gastrointest. Oncol. 2021, 13, 400. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, T.; Rahman, F.; Smith, A.M. The Microbiome and Radiation Induced-Bowel Injury: Evidence for Potential Mechanistic Role in Disease Pathogenesis. Nutrients 2018, 10, 1405. [Google Scholar] [CrossRef]
- Hollingsworth, B.A.; Cassatt, D.R.; DiCarlo, A.L.; Rios, C.I.; Satyamitra, M.M.; Winters, T.A.; Taliaferro, L.P. Acute Radiation Syndrome and the Microbiome: Impact and Review. Front. Pharmacol. 2021, 12, 643283. [Google Scholar] [CrossRef]
- Stacey, R.; Green, J.T. Radiation-Induced Small Bowel Disease: Latest Developments and Clinical Guidance. Ther. Adv. Chronic Dis. 2014, 5, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Malipatlolla, D.K.; Patel, P.; Sjöberg, F.; Devarakonda, S.; Kalm, M.; Angenete, E.; Lindskog, E.B.; Grandér, R.; Persson, L.; Stringer, A.; et al. Long-Term Mucosal Injury and Repair in a Murine Model of Pelvic Radiotherapy. Sci. Rep. 2019, 9, 13803. [Google Scholar] [CrossRef] [PubMed]
- Appanna, V.D. Human Microbes-the Power within: Health, Healing and Beyond; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–175. [Google Scholar]
- Paquette, S.; Thomas, S.C.; Venkataraman, K.; Appanna, V.D.; Tharmalingam, S. The Effects of Oral Probiotics on Type 2 Diabetes Mellitus (T2DM): A Clinical Trial Systematic Literature Review. Nutrients 2023, 15, 4690. [Google Scholar] [CrossRef]
- Lkhagva, D.E.; Chung, H.J.; Hong, J.; Tang, W.H.W.; Lee, S.I.; Hong, S.T.; Lee, S. The Regional Diversity of Gut Microbiome along the GI Tract of Male C57BL/6 Mice. BMC Microbiol. 2021, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Song, C.; Liu, Y.; Wu, X.; Dong, W.; Zhu, H.; Xiang, Z.; Qin, C. Characteristics of Gut Microbiota in Representative Mice Strains: Implications for Biological Research. Anim. Model. Exp. Med. 2022, 5, 337–349. [Google Scholar] [CrossRef]
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef]
- Tian, T.; Zhao, Y.; Yang, Y.; Wang, T.; Jin, S.; Guo, J.; Liu, Z. The Protective Role of Short-Chain Fatty Acids Acting as Signal Molecules in Chemotherapy- or Radiation-Induced Intestinal Inflammation. Am. J. Cancer Res. 2020, 10, 3508–3531. [Google Scholar]
- Nam, Y.-D.; Kim, H.J.; Seo, J.-G.; Kang, S.W.; Bae, J.-W. Impact of Pelvic Radiotherapy on Gut Microbiota of Gynecological Cancer Patients Revealed by Massive Pyrosequencing. PLoS ONE 2013, 8, e82659. [Google Scholar] [CrossRef]
- Wang, A.; Ling, Z.; Yang, Z.; Kiela, P.R.; Wang, T.; Wang, C.; Cao, L.; Geng, F.; Shen, M.; Ran, X.; et al. Gut Microbial Dysbiosis May Predict Diarrhea and Fatigue in Patients Undergoing Pelvic Cancer Radiotherapy: A Pilot Study. PLoS ONE 2015, 10, e0126312. [Google Scholar] [CrossRef]
- Villéger, R.; Lopès, A.; Carrier, G.; Veziant, J.; Billard, E.; Barnich, N.; Gagnière, J.; Vazeille, E.; Bonnet, M. Intestinal Microbiota: A Novel Target to Improve Anti-Tumor Treatment? Int. J. Mol. Sci. 2019, 20, 4584. [Google Scholar] [CrossRef] [PubMed]
- Kalkeri, R.; Walters, K.; Van Der Pol, W.; McFarland, B.C.; Fisher, N.; Koide, F.; Morrow, C.D.; Singh, V.K. Changes in the Gut Microbiome Community of Nonhuman Primates Following Radiation Injury. BMC Microbiol. 2021, 21, 93. [Google Scholar] [CrossRef] [PubMed]
- Carbonero, F.; Mayta, A.; Bolea, M.; Yu, J.Z.; Lindeblad, M.; Lyubimov, A.; Neri, F.; Szilagyi, E.; Smith, B.; Halliday, L.; et al. Specific Members of the Gut Microbiota Are Reliable Biomarkers of Irradiation Intensity and Lethality in Large Animal Models of Human Health. Radiat. Res. 2018, 191, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, Y.; Wang, S.; Guan, H.; Hu, S.; Huang, R.; Zhou, P. Impact of Low-Dose Ionising Radiation on the Composition of the Gut Microbiota of Mice. Toxicol. Sci. 2019, 171, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.R.; Muls, A.; Dearnaley, D.P.; Andreyev, H.J.N. Microbiota and Radiation-Induced Bowel Toxicity: Lessons from Inflammatory Bowel Disease for the Radiation Oncologist. Lancet Oncol. 2014, 15, e139–e147. [Google Scholar] [CrossRef] [PubMed]
- Touchefeu, Y.; Montassier, E.; Nieman, K.; Gastinne, T.; Potel, G.; Bruley Des Varannes, S.; Le Vacon, F.; De La Cochetière, M.F. Systematic Review: The Role of the Gut Microbiota in Chemotherapy- or Radiation-Induced Gastrointestinal Mucositis—Current Evidence and Potential Clinical Applications. Aliment. Pharmacol. Ther. 2014, 40, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Segers, C.; Mastroleo, F.; Verslegers, M.; Baatout, S.; Leys, N.; Lebeer, S. Food Supplements to Mitigate Detrimental Effects of Pelvic Radiotherapy. Microorganisms 2019, 7, 97. [Google Scholar] [CrossRef]
- Macià i Garau, M.; Lucas Calduch, A.; López, E.C. Radiobiology of the Acute Radiation Syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 123–130. [Google Scholar] [CrossRef]
- Segers, C.; Mysara, M.; Claesen, J.; Baatout, S.; Leys, N.; Lebeer, S.; Verslegers, M.; Mastroleo, F. Intestinal Mucositis Precedes Dysbiosis in a Mouse Model for Pelvic Irradiation. ISME Commun. 2021, 11, 24. [Google Scholar] [CrossRef]
- Ciorba, M.A.; Riehl, T.E.; Rao, M.S.; Moon, C.; Ee, X.; Nava, G.M.; Walker, M.R.; Marinshaw, J.M.; Stappenbeck, T.S.; Stenson, W.F. Lactobacillus Probiotic Protects Intestinal Epithelium from Radiation Injury in a TLR-2/Cyclo-Oxygenase-2-Dependent Manner. Gut 2012, 61, 829–838. [Google Scholar] [CrossRef]
- Jian, Y.P.; Yang, G.; Zhang, L.H.; Liang, J.Y.; Zhou, H.L.; Wang, Y.S.; Xu, Z.X. Lactobacillus Plantarum Alleviates Irradiation-Induced Intestinal Injury by Activation of FXR-FGF15 Signaling in Intestinal Epithelia. J. Cell. Physiol. 2022, 237, 1845–1856. [Google Scholar] [CrossRef]
- Zhao, Z.; Cheng, W.; Qu, W.; Shao, G.; Liu, S. Antibiotic Alleviates Radiation-Induced Intestinal Injury by Remodeling Microbiota, Reducing Inflammation, and Inhibiting Fibrosis. ACS Omega 2020, 5, 2967–2977. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Kainthola, A.; Tiwari, M.; Agrawala, P.K. Gut Microbiota Response to Ionizing Radiation and Its Modulation by HDAC Inhibitor TSA. Int. J. Radiat. Biol. 2020, 96, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Livanova, A.A.; Fedorova, A.A.; Zavirsky, A.V.; Krivoi, I.I.; Markov, A.G. Dose- and Segment-Dependent Disturbance of Rat Gut by Ionizing Radiation: Impact of Tight Junction Proteins. Int. J. Mol. Sci. 2023, 24, 1753. [Google Scholar] [CrossRef]
- Zhang, D.; Zhong, D.; Ouyang, J.; He, J.; Qi, Y.; Chen, W.; Zhang, X.; Tao, W.; Zhou, M. Microalgae-Based Oral Microcarriers for Gut Microbiota Homeostasis and Intestinal Protection in Cancer Radiotherapy. Nat. Commun. 2022, 13, 1413. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.; Sottili, M.; Gerini, C.; Desideri, I.; Bastida, C.; Pallotta, S.; Castiglione, F.; Bonomo, P.; Meattini, I.; Greto, D.; et al. A PPAR-Gamma Agonist Protects from Radiation-Induced Intestinal Toxicity. United Eur. Gastroenterol. J. 2017, 5, 218–226. [Google Scholar] [CrossRef]
- Jang, H.; Kim, S.; Kim, H.; Oh, S.H.; Kwak, S.Y.; Joo, H.W.; Lee, S.B.; Jang, W.I.; Park, S.; Shim, S. Metformin Protects the Intestinal Barrier by Activating Goblet Cell Maturation and Epithelial Proliferation in Radiation-Induced Enteropathy. Int. J. Mol. Sci. 2022, 23, 5929. [Google Scholar] [CrossRef]
- Umar, S. Intestinal Stem Cells. Curr. Gastroenterol. Rep. 2010, 12, 340. [Google Scholar] [CrossRef]
- Etienne-Mesmin, L.; Chassaing, B.; Desvaux, M.; De Paepe, K.; Gresse, R.; Sauvaitre, T.; Forano, E.; De Wiele, T.V.; Schüller, S.; Juge, N.; et al. Experimental Models to Study Intestinal Microbes–Mucus Interactions in Health and Disease. FEMS Microbiol. Rev. 2019, 43, 457–489. [Google Scholar] [CrossRef]
- Paone, P.; Cani, P.D. Mucus Barrier, Mucins and Gut Microbiota: The Expected Slimy Partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Peredo-Lovillo, A.; Romero-Luna, H.E.; Jiménez-Fernández, M. Health Promoting Microbial Metabolites Produced by Gut Microbiota after Prebiotics Metabolism. Food Res. Int. 2020, 136, 109473. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ho, S.B. Intestinal Goblet Cells and Mucins in Health and Disease: Recent Insights and Progress. Curr. Gastroenterol. Rep. 2010, 12, 319. [Google Scholar] [CrossRef]
- Deplancke, B.; Gaskins, H.R. Microbial Modulation of Innate Defense: Goblet Cells and the Intestinal Mucus Layer. Am. J. Clin. Nutr. 2001, 73, 1131S–1141S. [Google Scholar] [CrossRef]
- Park, J.C.; Im, S.H. Of Men in Mice: The Development and Application of a Humanized Gnotobiotic Mouse Model for Microbiome Therapeutics. Exp. Mol. Med. 2020, 52, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E.W. Anaerobic Bacteria, the Colon and Colitis. Aust. N. Z. J. Surg. 1980, 50, 73–75. [Google Scholar] [CrossRef]
- Larsen, J.M. The Immune Response to Prevotella Bacteria in Chronic Inflammatory Disease. Immunology 2017, 151, 363. [Google Scholar] [CrossRef]
- Dempsey, E.; Corr, S.C. Lactobacillus Spp. for Gastrointestinal Health: Current and Future Perspectives. Front. Immunol. 2022, 13, 840245. [Google Scholar] [CrossRef]
- Shuryak, I. Review of Microbial Resistance to Chronic Ionizing Radiation Exposure under Environmental Conditions. J. Environ. Radioact. 2019, 196, 50–63. [Google Scholar] [CrossRef]
- Lai, J. Radioresistance: Implications for Astrobiological and Medical Research. Health Sci. Isc. 2019, 4, 75–79. [Google Scholar]
- Pavlopoulou, A.; Savva, G.D.; Louka, M.; Bagos, P.G.; Vorgias, C.E.; Michalopoulos, I.; Georgakilas, A.G. Unraveling the Mechanisms of Extreme Radioresistance in Prokaryotes: Lessons from Nature. Mutat. Res.-Rev. Mutat. Res. 2016, 767, 92–107. [Google Scholar] [CrossRef]
- Davis-Richardson, A.G.; Triplett, E.W. A Model for the Role of Gut Bacteria in the Development of Autoimmunity for Type 1 Diabetes. Diabetologia 2015, 58, 1386–1393. [Google Scholar] [CrossRef]
- Mitra, A.; Grossman Biegert, G.W.; Delgado, A.Y.; Karpinets, T.V.; Solley, T.N.; Mezzari, M.P.; Yoshida-Court, K.; Petrosino, J.F.; Mikkelson, M.D.; Lin, L.; et al. Microbial Diversity and Composition Is Associated with Patient-Reported Toxicity during Chemoradiation Therapy for Cervical Cancer. Int. J. Radiat. Oncol. Biol. Phys. 2020, 107, 163–171. [Google Scholar] [CrossRef]
- Cénit, M.C.; Matzaraki, V.; Tigchelaar, E.F.; Zhernakova, A. Rapidly Expanding Knowledge on the Role of the Gut Microbiome in Health and Disease. Biochim. Biophys. Acta-Mol. Basis Dis. 2014, 1842, 1981–1992. [Google Scholar] [CrossRef]
- Singh, V.; Ahlawat, S.; Mohan, H.; Gill, S.S.; Sharma, K.K. Balancing Reactive Oxygen Species Generation by Rebooting Gut Microbiota. J. Appl. Microbiol. 2022, 132, 4112–4129. [Google Scholar] [CrossRef] [PubMed]
- Mohan, H.; Vandna; Soni, S.; Syed, S. Targeting Reactive Oxygen Species (ROS) for Cancer Therapy. In Handbook of Oxidative Stress in Cancer: Therapeutic Aspects; Springer: Berlin/Heidelberg, Germany, 2022; pp. 3181–3196. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.M.; Duckworth, C.A.; Vowell, K.; Burkitt, M.D.; Pritchard, D.M. Intestinal Preparation Techniques for Histological Analysis in the Mouse. Curr. Protoc. Mouse Biol. 2016, 6, 148–168. [Google Scholar] [CrossRef] [PubMed]
- Shahi, S.K.; Zarei, K.; Guseva, N.V.; Mangalam, A.K. Microbiota Analysis Using Two-Step PCR and Next-Generation 16S RRNA Gene Sequencing. J. Vis. Exp. 2019, 2019, e59980. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Technol. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jameus, A.; Dougherty, J.; Narendrula, R.; Levert, D.; Valiquette, M.; Pirkkanen, J.; Lalonde, C.; Bonin, P.; Gagnon, J.D.; Appanna, V.D.; et al. Acute Impacts of Ionizing Radiation Exposure on the Gastrointestinal Tract and Gut Microbiome in Mice. Int. J. Mol. Sci. 2024, 25, 3339. https://doi.org/10.3390/ijms25063339
Jameus A, Dougherty J, Narendrula R, Levert D, Valiquette M, Pirkkanen J, Lalonde C, Bonin P, Gagnon JD, Appanna VD, et al. Acute Impacts of Ionizing Radiation Exposure on the Gastrointestinal Tract and Gut Microbiome in Mice. International Journal of Molecular Sciences. 2024; 25(6):3339. https://doi.org/10.3390/ijms25063339
Chicago/Turabian StyleJameus, Alexandra, Jessica Dougherty, Ramya Narendrula, Daniela Levert, Manon Valiquette, Jake Pirkkanen, Christine Lalonde, Patrice Bonin, Jeffrey D. Gagnon, Vasu D. Appanna, and et al. 2024. "Acute Impacts of Ionizing Radiation Exposure on the Gastrointestinal Tract and Gut Microbiome in Mice" International Journal of Molecular Sciences 25, no. 6: 3339. https://doi.org/10.3390/ijms25063339