
Sex-Specific Effects of Estradiol and Progesterone in Ischemic Kidney Injury
 , , , , , , and
, , , , , , and 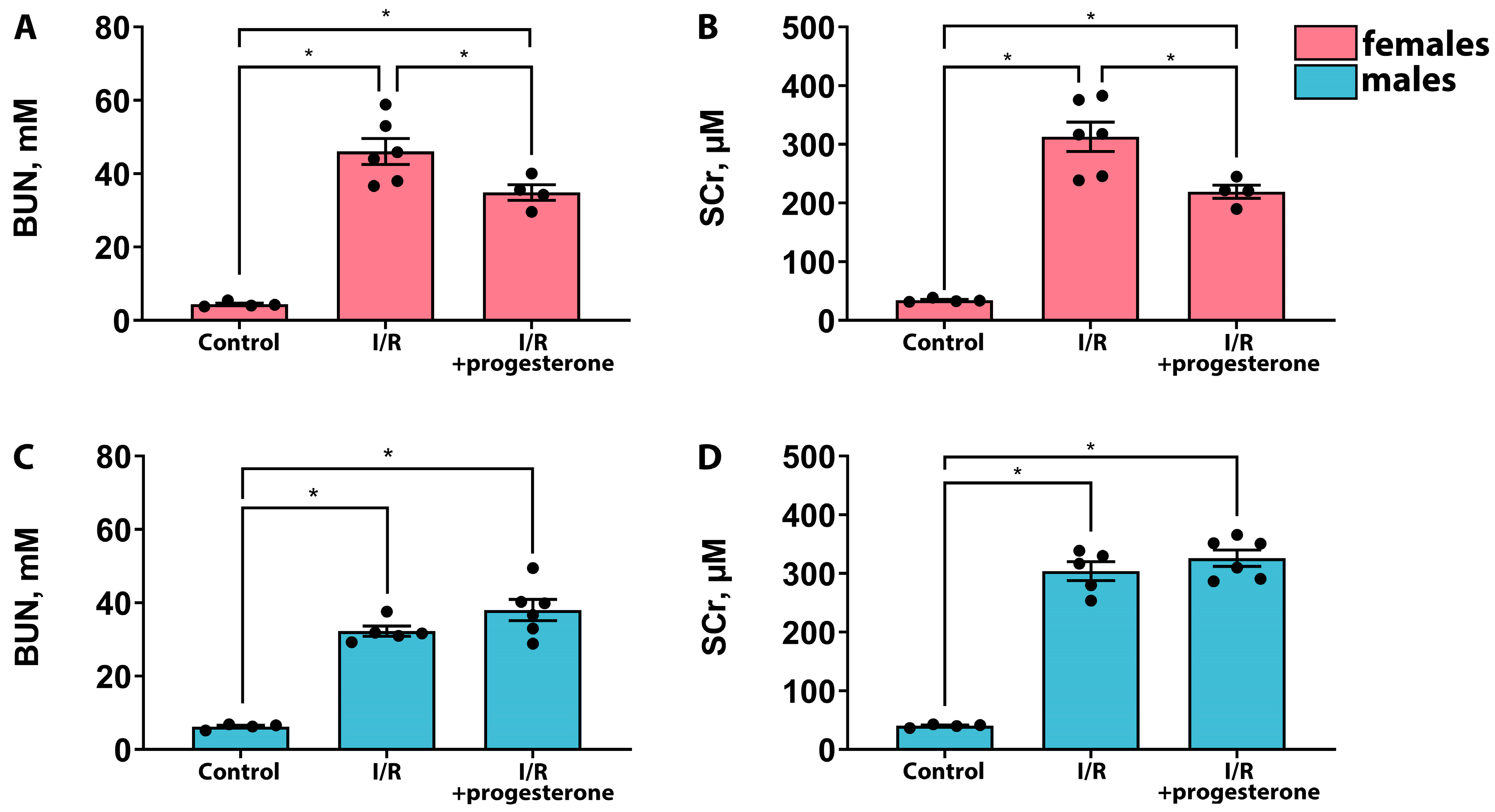{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
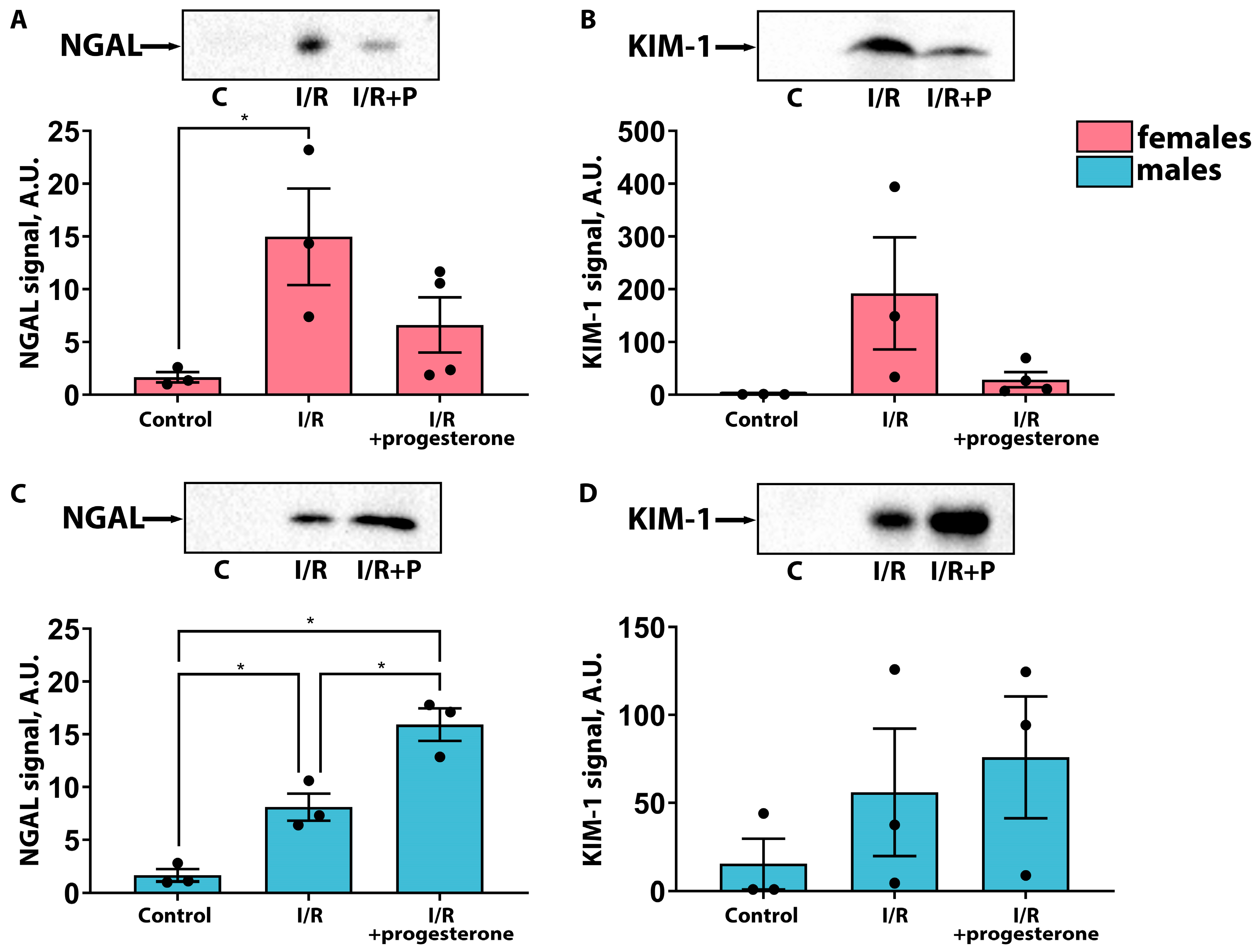
2.1. Progesterone Ameliorates the Severity of AKI in Female but Not Male Rats
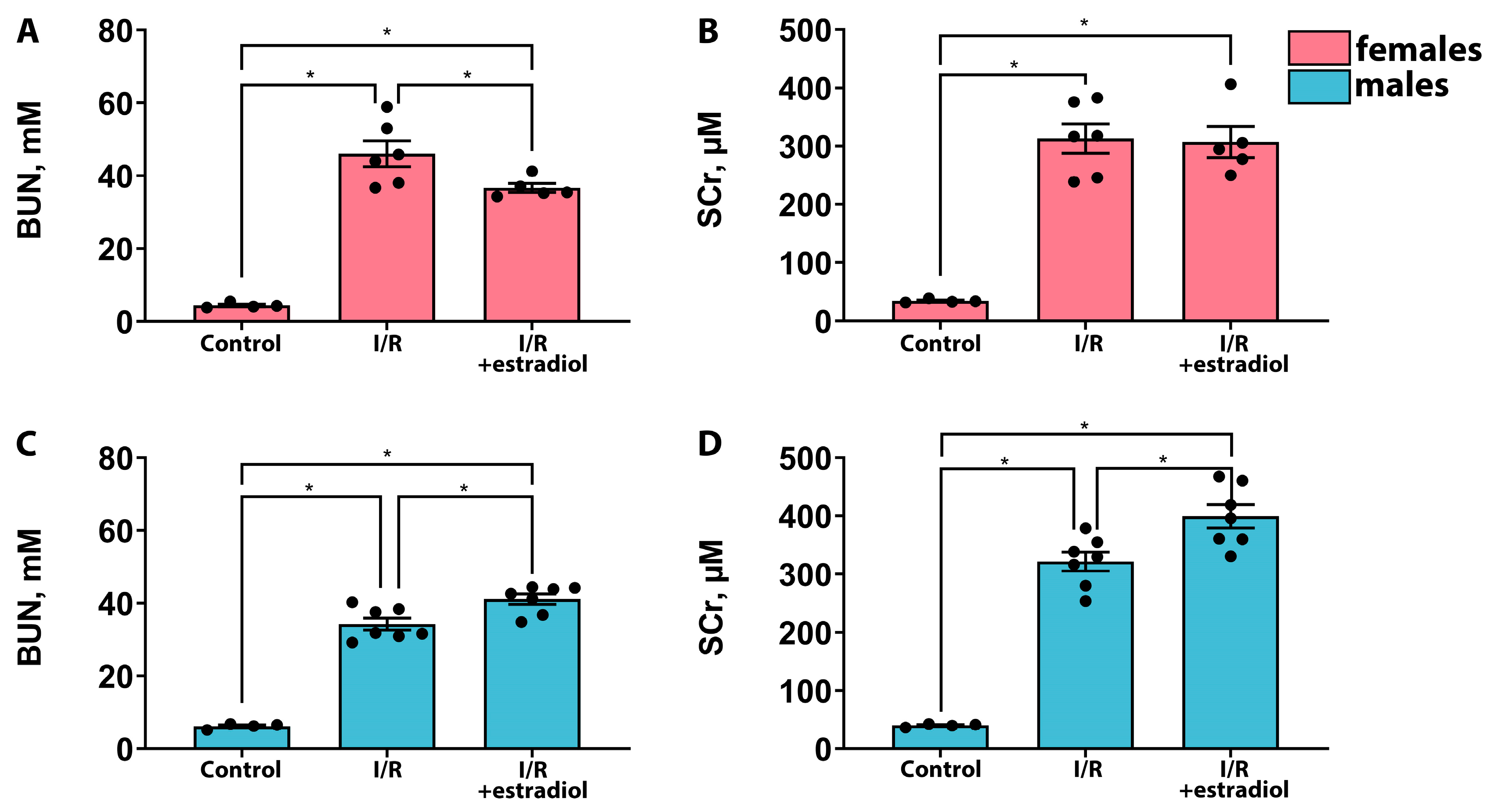
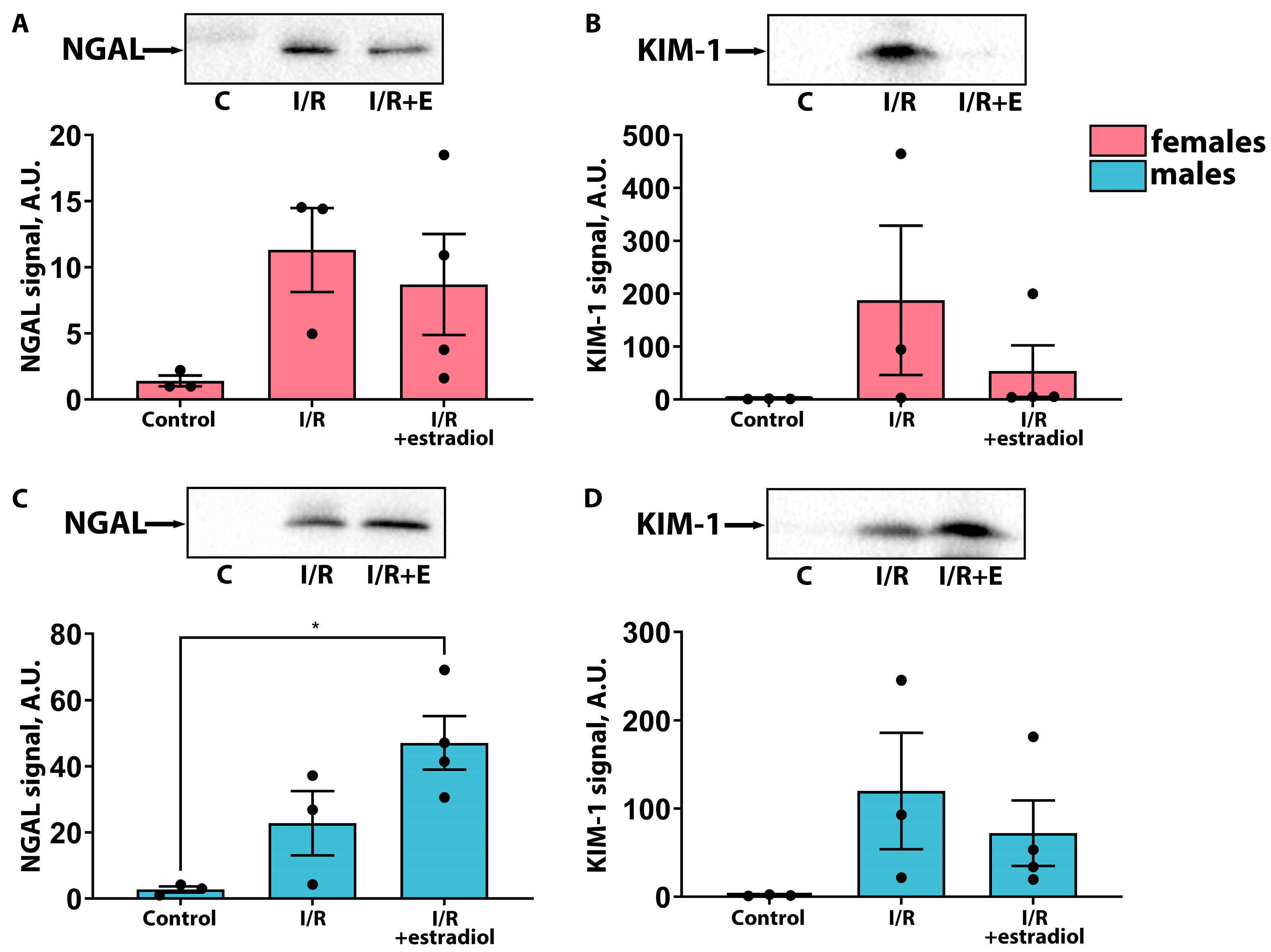
2.2. Estradiol Mitigates the Severity of AKI in Female Rats but Worsens It in Male Rats
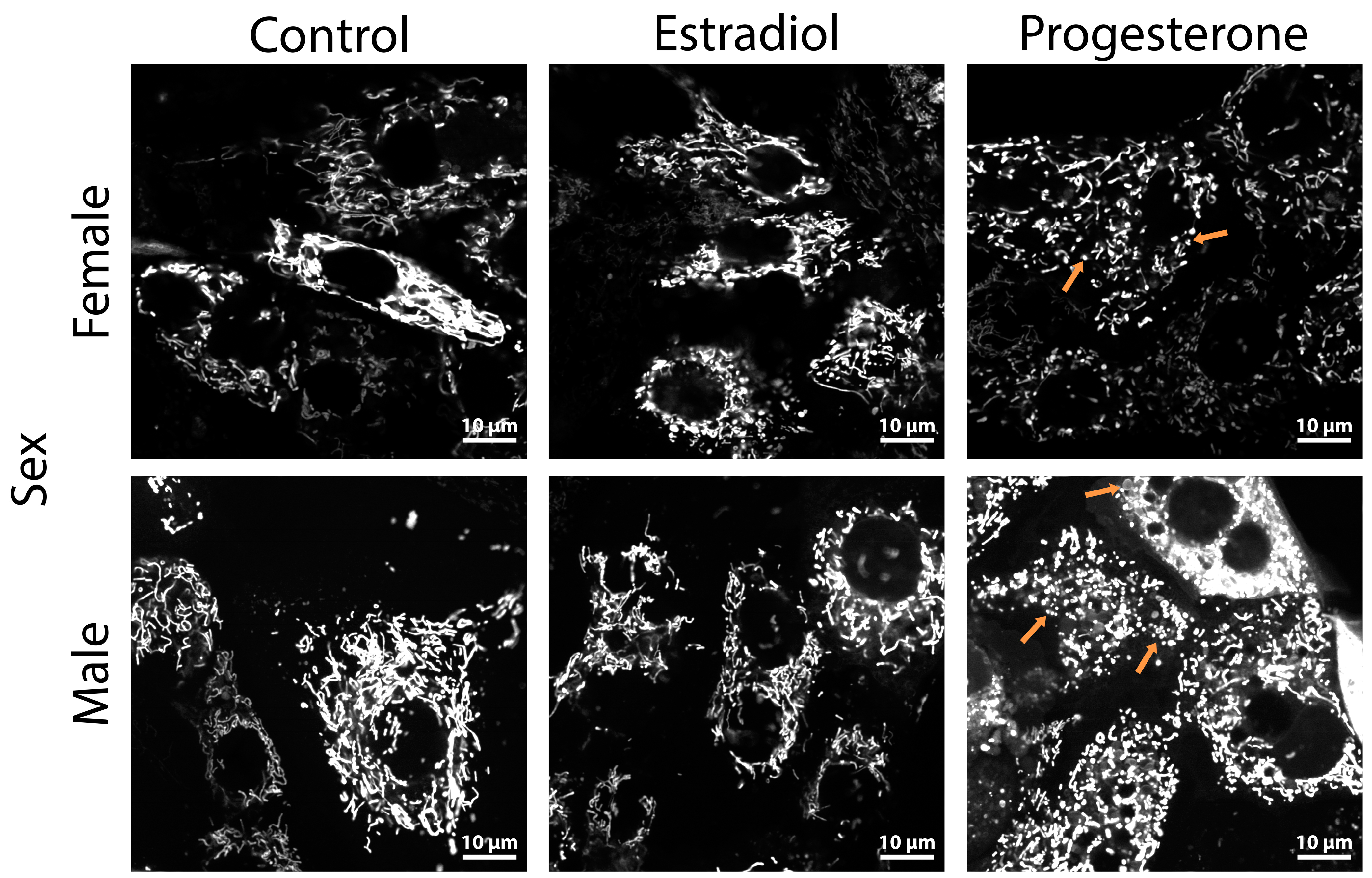
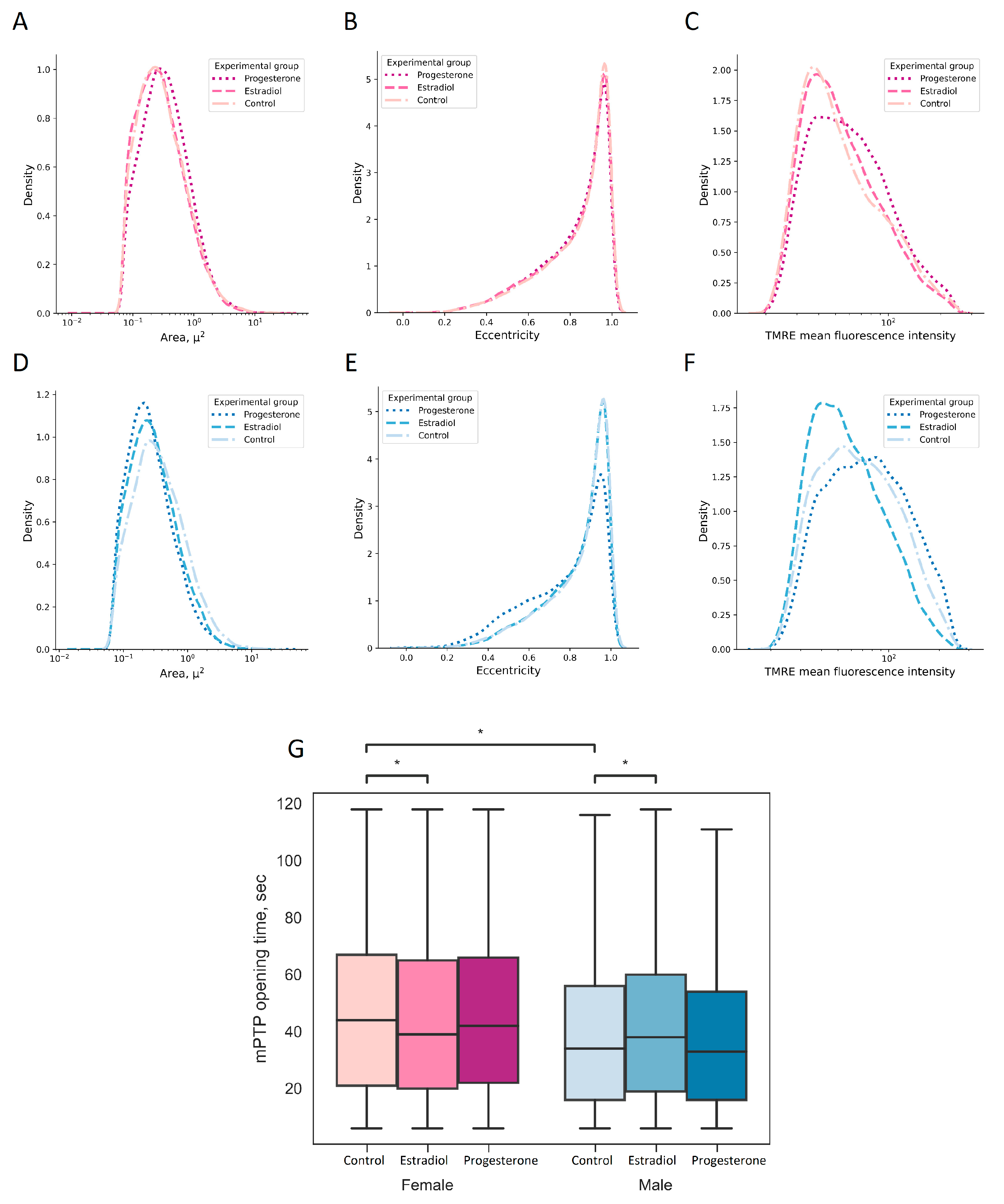
2.3. Progesterone Causes Mitochondrial Fragmentation in Kidney Cells
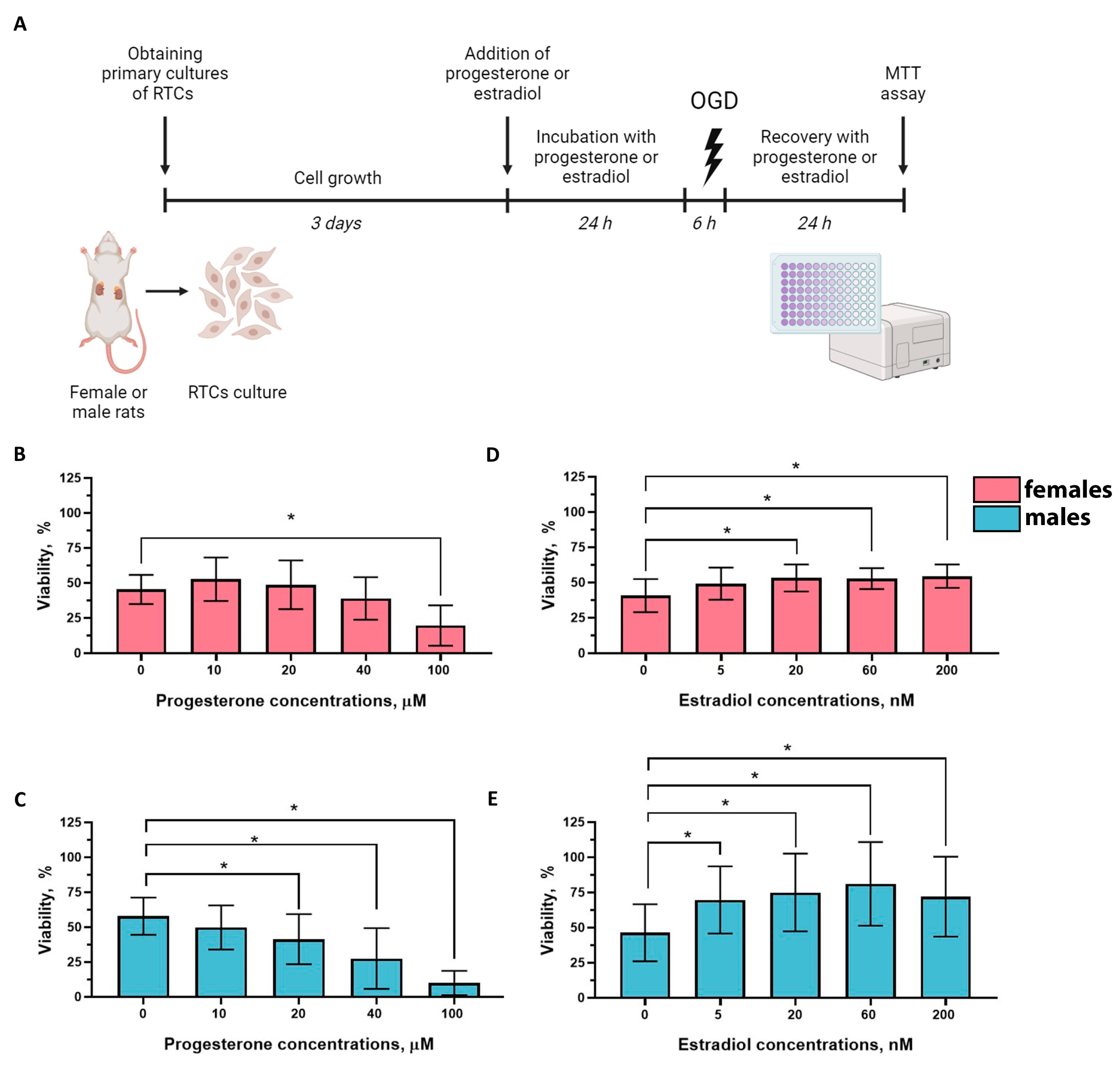
2.4. Effects of Progesterone and Estradiol on the Viability of RTCs from Female and Male Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Renal I/R Protocol
4.3. Serum Analysis
4.4. Western Blotting
4.5. Primary RTCs Cultures
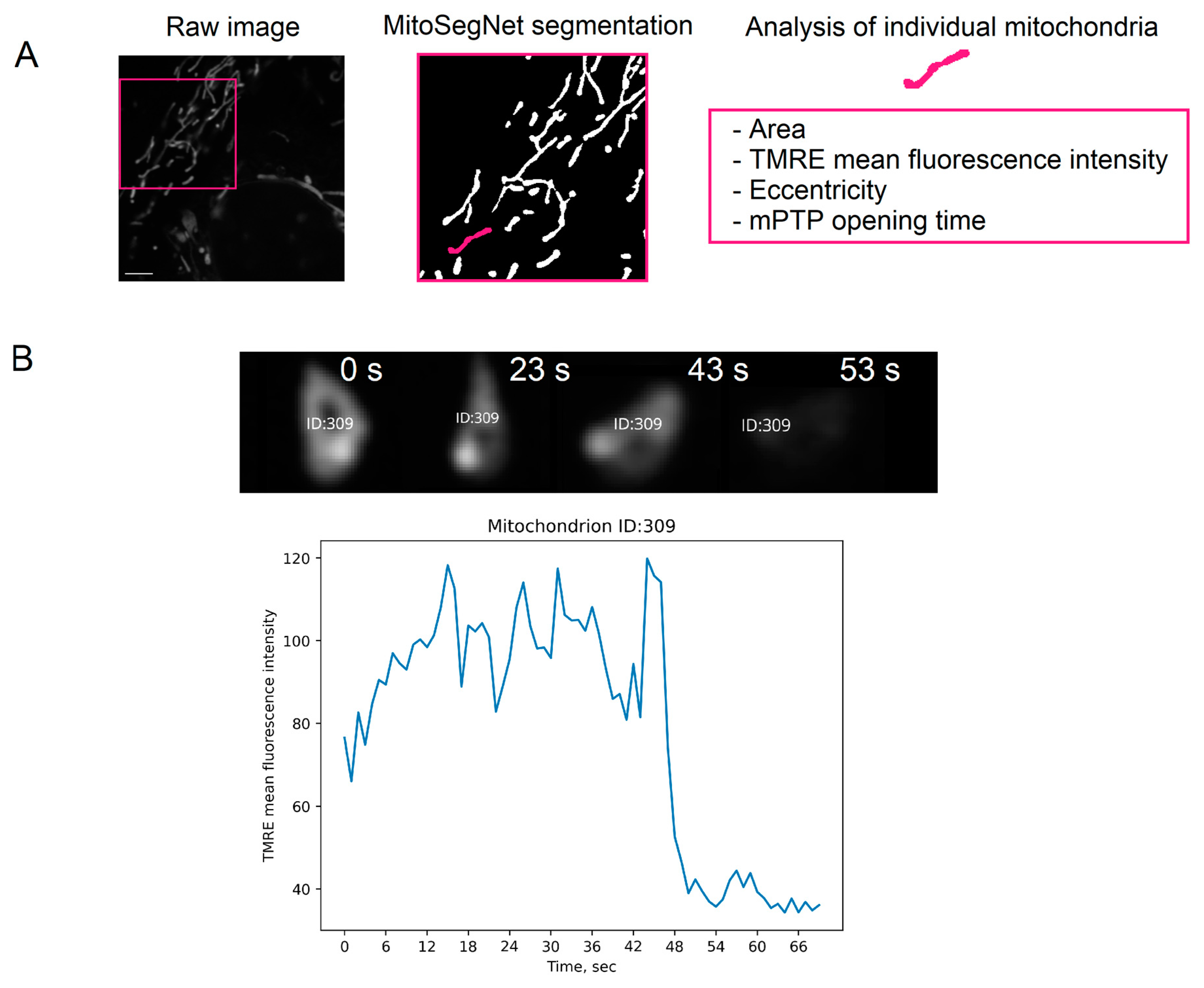
4.6. Analysis of Characteristics of Individual Mitochondria
4.7. Cell Viability
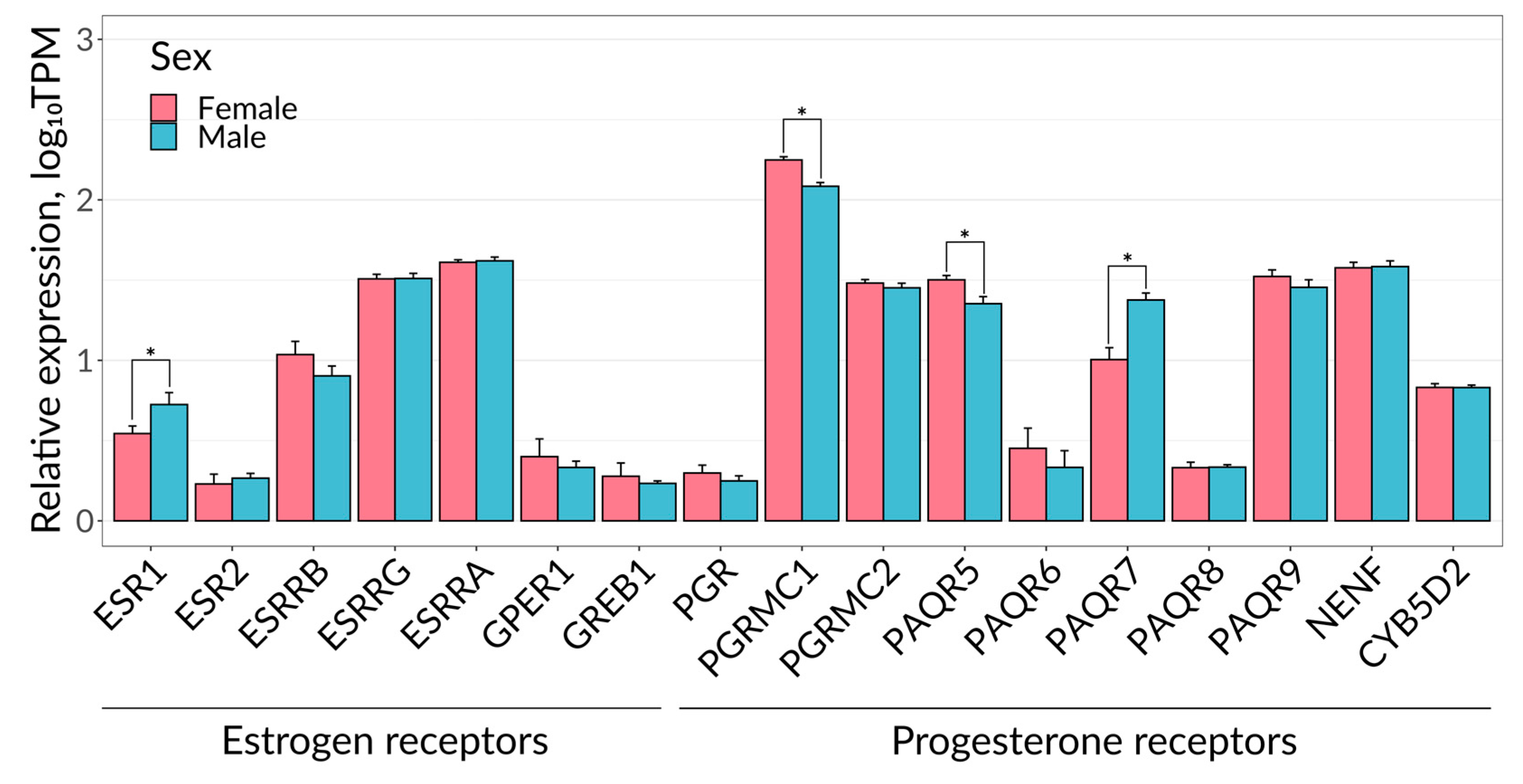
4.8. Bioinformatics Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anand, S.S.; Islam, S.; Rosengren, A.; Franzosi, M.G.; Steyn, K.; Yusufali, A.H.; Keltai, M.; Diaz, R.; Rangarajan, S.; Yusuf, S.; et al. Risk Factors for Myocardial Infarction in Women and Men: Insights from the INTERHEART Study. Eur. Heart J. 2008, 29, 932–940. [Google Scholar] [CrossRef]
- Hochman, J.S.; McCabe, C.H.; Stone, P.H.; Becker, R.C.; Cannon, C.P.; DeFeo-Fraulini, T.; Thompson, B.R.; Steingart, R.; Knatterud, G.; Braunwald, E. Outcome and Profile of Women and Men Presenting With Acute Coronary Syndromes: A Report From TIMI IIIB fn1. J. Am. Coll. Cardiol. 1997, 30, 141–148. [Google Scholar] [CrossRef]
- Heer, T.; Gitt, A.K.; Juenger, C.; Schiele, R.; Wienbergen, H.; Towae, F.; Gottwitz, M.; Zahn, R.; Zeymer, U.; Senges, J. Gender Differences in Acute Non–ST-Segment Elevation Myocardial Infarction. Am. J. Cardiol. 2006, 98, 160–166. [Google Scholar] [CrossRef]
- Costenbader, K.H.; Feskanich, D.; Stampfer, M.J.; Karlson, E.W. Reproductive and Menopausal Factors and Risk of Systemic Lupus Erythematosus in Women. Arthritis Rheum. 2007, 56, 1251–1262. [Google Scholar] [CrossRef]
- Sandberg, K. Mechanisms Underlying Sex Differences in Progressive Renal Disease. Gend. Med. 2008, 5, 10–23. [Google Scholar] [CrossRef]
- Grodstein, F.; Stampfer, M.J.; Manson, J.E.; Colditz, G.A.; Willett, W.C.; Rosner, B.; Speizer, F.E.; Hennekens, C.H. Postmenopausal Estrogen and Progestin Use and the Risk of Cardiovascular Disease. N. Engl. J. Med. 1996, 335, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Zhang, L. Gender Differences in Cardioprotection against Ischemia/reperfusion Injury in Adult Rat Hearts: Focus on Akt and Protein Kinase C Signaling. J. Pharmacol. Exp. Ther. 2005, 315, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.-C.; Yu, H.-P.; Suzuki, T.; Choudhry, M.A.; Schwacha, M.G.; Bland, K.I.; Chaudry, I.H. Upregulation of Mitochondrial Respiratory Complex IV by Estrogen Receptor-Beta Is Critical for Inhibiting Mitochondrial Apoptotic Signaling and Restoring Cardiac Functions Following Trauma-Hemorrhage. J. Mol. Cell. Cardiol. 2006, 41, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.L.; Cerdá, J.; Burdmann, E.A.; Tonelli, M.; García-García, G.; Jha, V.; Susantitaphong, P.; Rocco, M.; Vanholder, R.; Sever, M.S.; et al. International Society of Nephrology’s 0by25 Initiative for Acute Kidney Injury (zero Preventable Deaths by 2025): A Human Rights Case for Nephrology. Lancet 2015, 385, 2616–2643. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Bai, M.; Lei, J.; Xie, Y.; Xu, S.; Jia, Z.; Zhang, A. Mitochondrial Dysfunction and the AKI-to-CKD Transition. Am. J. Physiol. Renal Physiol. 2020, 319, F1105–F1116. [Google Scholar] [CrossRef] [PubMed]
- Scholz, H.; Boivin, F.J.; Schmidt-Ott, K.M.; Bachmann, S.; Eckardt, K.-U.; Scholl, U.I.; Persson, P.B. Kidney Physiology and Susceptibility to Acute Kidney Injury: Implications for Renoprotection. Nat. Rev. Nephrol. 2021, 17, 335–349. [Google Scholar] [CrossRef]
- Rossaint, J.; Zarbock, A. Acute Kidney Injury: Definition, Diagnosis and Epidemiology. Minerva Urol. Nefrol. 2016, 68, 49–57. [Google Scholar]
- Hosszu, A.; Fekete, A.; Szabo, A.J. Sex Differences in Renal Ischemia-Reperfusion Injury. Am. J. Physiol. Renal Physiol. 2020, 319, F149–F154. [Google Scholar] [CrossRef]
- Fekete, A.; Vannay, A.; Vér, A.; Rusai, K.; Müller, V.; Reusz, G.; Tulassay, T.; Szabó, A.J. Sex Differences in Heat Shock Protein 72 Expression and Localization in Rats Following Renal Ischemia-Reperfusion Injury. Am. J. Physiol. Renal Physiol. 2006, 291, F806–F811. [Google Scholar] [CrossRef] [PubMed]
- Crockett, E.T.; Spielman, W.; Dowlatshahi, S.; He, J. Sex Differences in Inflammatory Cytokine Production in Hepatic Ischemia-Reperfusion. J. Inflamm. 2006, 3, 16. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Wang, M.-H.; Dong, Z. Differential Gender Differences in Ischemic and Nephrotoxic Acute Renal Failure. Am. J. Nephrol. 2005, 25, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Li, F.; Di, Z.; Xiong, Y.; Zhang, S.; Ma, Q.; Bian, X.; Lang, Z.; Ye, Q.; Wang, Y. Estradiol Ameliorates Acute Kidney Ischemia-Reperfusion Injury by Inhibiting the TGF-βRI-SMAD Pathway. Front. Immunol. 2022, 13, 822604. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Kim, J.I.; Ahn, Y.; Bonventre, A.J.; Bonventre, J.V. Testosterone Is Responsible for Enhanced Susceptibility of Males to Ischemic Renal Injury. J. Biol. Chem. 2004, 279, 52282–52292. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Martínez, N.; Halhali, A.; Barrera, D. Steroid Hormones and Pregnancy. Gynecol. Endocrinol. 2019, 35, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Popkov, V.A.; Andrianova, N.V.; Manskikh, V.N.; Silachev, D.N.; Pevzner, I.B.; Zorova, L.D.; Sukhikh, G.T.; Plotnikov, E.Y.; Zorov, D.B. Pregnancy Protects the Kidney from Acute Ischemic Injury. Sci. Rep. 2018, 8, 14534. [Google Scholar] [CrossRef]
- Ralto, K.M.; Parikh, S.M. Mitochondria in Acute Kidney Injury. Semin. Nephrol. 2016, 36, 8–16. [Google Scholar] [CrossRef]
- Li, Z.; Lu, S.; Qian, B.; Meng, Z.; Zhou, Y.; Chen, D.; Chen, B.; Yang, G.; Ma, Y. Sex Differences in Hepatic Ischemia—Reperfusion Injury: A Cross-Sectional Study. Sci. Rep. 2023, 13, 5724. [Google Scholar] [CrossRef]
- Ostadal, B.; Ostadal, P. Sex-Based Differences in Cardiac Ischaemic Injury and Protection: Therapeutic Implications. Br. J. Pharmacol. 2014, 171, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Neugarten, J.; Golestaneh, L.; Kolhe, N.V. Sex Differences in Acute Kidney Injury Requiring Dialysis. BMC Nephrol. 2018, 19, 131. [Google Scholar] [CrossRef] [PubMed]
- Robert, R.; Ghazali, D.A.; Favreau, F.; Mauco, G.; Hauet, T.; Goujon, J.-M. Gender Difference and Sex Hormone Production in Rodent Renal Ischemia Reperfusion Injury and Repair. J. Inflamm. 2011, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Buléon, M.; Cuny, M.; Grellier, J.; Charles, P.-Y.; Belliere, J.; Casemayou, A.; Arnal, J.-F.; Schanstra, J.-P.; Tack, I. A Single Dose of Estrogen during Hemorrhagic Shock Protects against Kidney Injury Whereas Estrogen Restoration in Ovariectomized Mice Is Ineffective. Sci. Rep. 2020, 10, 17240. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Lateef, S.M.; El-Sayed, E.-S.M.; Mansour, A.M.; Salama, S.A. The Protective Role of Estrogen and Its Receptors in Gentamicin-Induced Acute Kidney Injury in Rats. Life Sci. 2019, 239, 117082. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.; Allam, S.; Mansour, A.M.; Shaheen, A.; Salama, S.A. The Potential Protective Effects of Estradiol and 2-Methoxyestradiol in Ischemia Reperfusion-Induced Kidney Injury in Ovariectomized Female Rats. Life Sci. 2022, 296, 120441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chang, Y.; Han, Z.; Ma, K.; Zeng, X.; Li, L. Estrogen Protects against Renal Ischemia-Reperfusion Injury by Regulating Th17/Treg Cell Immune Balance. Dis. Markers 2022, 2022, 7812099. [Google Scholar] [CrossRef]
- Al-Trad, B.; Ashankyty, I.M.; Alaraj, M. Progesterone Ameliorates Diabetic Nephropathy in Streptozotocin-Induced Diabetic Rats. Diabetol. Metab. Syndr. 2015, 7, 97. [Google Scholar] [CrossRef]
- Satake, A.; Takaoka, M.; Nishikawa, M.; Yuba, M.; Shibata, Y.; Okumura, K.; Kitano, K.; Tsutsui, H.; Fujii, K.; Kobuchi, S.; et al. Protective Effect of 17beta-Estradiol on Ischemic Acute Renal Failure through the PI3K/Akt/eNOS Pathway. Kidney Int. 2008, 73, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Iran-Nejad, A.; Nematbakhsh, M.; Eshraghi-Jazi, F.; Talebi, A. Preventive Role of Estradiol on Kidney Injury Induced by Renal Ischemia-Reperfusion in Male and Female Rats. Int. J. Prev. Med. 2015, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.; de Araújo, G.W.; Beyer, C. Gender-Specific Regulation of Mitochondrial Fusion and Fission Gene Transcription and Viability of Cortical Astrocytes by Steroid Hormones. J. Mol. Endocrinol. 2008, 41, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Li, L.Z.; Kastury, N.L.; Thomas, C.T.; Lam, M.P.Y.; Lau, E. Transcriptome Features of Striated Muscle Aging and Predictability of Protein Level Changes. Mol Omics 2021, 17, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Grimont, A.; Bloch-Faure, M.; El Abida, B.; Crambert, G. Mapping of Sex Hormone Receptors and Their Modulators along the Nephron of Male and Female Mice. FEBS Lett. 2009, 583, 1644–1648. [Google Scholar] [CrossRef] [PubMed]
- DeMayo, F.J.; Zhao, B.; Takamoto, N.; Tsai, S.Y. Mechanisms of Action of Estrogen and Progesterone. Ann. N. Y. Acad. Sci. 2002, 955, 48–59, discussion 86–88, 396–406. [Google Scholar] [CrossRef]
- Abramicheva, P.A.; Semenovich, D.S.; Zorova, L.D.; Pevzner, I.B.; Sokolov, I.A.; Popkov, V.A.; Kazakov, E.P.; Zorov, D.B.; Plotnikov, E.Y. Decreased Renal Expression of PAQR5 Is Associated with the Absence of a Nephroprotective Effect of Progesterone in a Rat UUO Model. Sci. Rep. 2023, 13, 12871. [Google Scholar] [CrossRef]
- Silachev, D.N.; Plotnikov, E.Y.; Pevzner, I.B.; Zorova, L.D.; Babenko, V.A.; Zorov, S.D.; Popkov, V.A.; Jankauskas, S.S.; Zinchenko, V.P.; Sukhikh, G.T.; et al. The Mitochondrion as a Key Regulator of Ischaemic Tolerance and Injury. Heart Lung Circ. 2014, 23, 897–904. [Google Scholar] [CrossRef]
- Tower, J. Sex-Specific Regulation of Aging and Apoptosis. Mech. Ageing Dev. 2006, 127, 705–718. [Google Scholar] [CrossRef]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A Central Target for Sex Differences in Pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Viña, J.; Borrás, C.; Gambini, J.; Sastre, J.; Pallardó, F.V. Why Females Live Longer than Males? Importance of the Upregulation of Longevity-Associated Genes by Oestrogenic Compounds. FEBS Lett. 2005, 579, 2541–2545. [Google Scholar] [CrossRef] [PubMed]
- Mattingly, K.A.; Ivanova, M.M.; Riggs, K.A.; Wickramasinghe, N.S.; Barch, M.J.; Klinge, C.M. Estradiol Stimulates Transcription of Nuclear Respiratory Factor-1 and Increases Mitochondrial Biogenesis. Mol. Endocrinol. 2008, 22, 609–622. [Google Scholar] [CrossRef]
- Chen, L.; Qin, Y.; Liu, B.; Gao, M.; Li, A.; Li, X.; Gong, G. PGC-1α-Mediated Mitochondrial Quality Control: Molecular Mechanisms and Implications for Heart Failure. Front. Cell Dev. Biol. 2022, 10, 871357. [Google Scholar] [CrossRef]
- Kiyama, T.; Chen, C.-K.; Wang, S.W.; Pan, P.; Ju, Z.; Wang, J.; Takada, S.; Klein, W.H.; Mao, C.-A. Essential Roles of Mitochondrial Biogenesis Regulator Nrf1 in Retinal Development and Homeostasis. Mol. Neurodegener. 2018, 13, 56. [Google Scholar] [CrossRef] [PubMed]
- Irwin, R.W.; Yao, J.; Hamilton, R.T.; Cadenas, E.; Brinton, R.D.; Nilsen, J. Progesterone and Estrogen Regulate Oxidative Metabolism in Brain Mitochondria. Endocrinology 2008, 149, 3167–3175. [Google Scholar] [CrossRef]
- Suen, D.-F.; Norris, K.L.; Youle, R.J. Mitochondrial Dynamics and Apoptosis. Genes Dev. 2008, 22, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, M.; Singh, S.; Singh, A.P.; Dasgupta, S. Mitochondrial Fusion and Fission: The Fine-Tune Balance for Cellular Homeostasis. FASEB J. 2021, 35, e21620. [Google Scholar] [CrossRef]
- Ortega-Domínguez, B.; Aparicio-Trejo, O.E.; García-Arroyo, F.E.; León-Contreras, J.C.; Tapia, E.; Molina-Jijón, E.; Hernández-Pando, R.; Sánchez-Lozada, L.G.; Barrera-Oviedo, D.; Pedraza-Chaverri, J. Curcumin Prevents Cisplatin-Induced Renal Alterations in Mitochondrial Bioenergetics and Dynamic. Food Chem. Toxicol. 2017, 107, 373–385. [Google Scholar] [CrossRef]
- Qin, L.; Xi, S. The Role of Mitochondrial Fission Proteins in Mitochondrial Dynamics in Kidney Disease. Int. J. Mol. Sci. 2022, 23, 14725. [Google Scholar] [CrossRef]
- Haworth, R.A.; Hunter, D.R. The Ca2+-Induced Membrane Transition in Mitochondria. II. Nature of the Ca2+ Trigger Site. Arch. Biochem. Biophys. 1979, 195, 460–467. [Google Scholar] [CrossRef]
- Elrod, J.W.; Molkentin, J.D. Physiologic Functions of Cyclophilin D and the Mitochondrial Permeability Transition Pore. Circ. J. 2013, 77, 1111–1122. [Google Scholar] [CrossRef]
- Zorov, D.B.; Filburn, C.R.; Klotz, L.O.; Zweier, J.L.; Sollott, S.J. Reactive Oxygen Species (ROS)-Induced ROS Release: A New Phenomenon Accompanying Induction of the Mitochondrial Permeability Transition in Cardiac Myocytes. J. Exp. Med. 2000, 192, 1001–1014. [Google Scholar] [CrossRef]
- Crompton, M. The Mitochondrial Permeability Transition Pore and Its Role in Cell Death. Biochem. J 1999, 341, 233–249. [Google Scholar] [CrossRef]
- Griffiths, E.J.; Halestrap, A.P. Protection by Cyclosporin A of Ischemia/reperfusion-Induced Damage in Isolated Rat Hearts. J. Mol. Cell. Cardiol. 1993, 25, 1461–1469. [Google Scholar] [CrossRef]
- Halestrap, A.P. A Pore Way to Die: The Role of Mitochondria in Reperfusion Injury and Cardioprotection. Biochem. Soc. Trans. 2010, 38, 841–860. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, P.; Di Lisa, F. The Mitochondrial Permeability Transition Pore: Molecular Nature and Role as a Target in Cardioprotection. J. Mol. Cell. Cardiol. 2015, 78, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.F., Jr.; Ronconi, K.S.; Morra, E.A.; Do Val Lima, P.R.; Porto, M.L.; Vassallo, D.V.; Figueiredo, S.G.; Stefanon, I. Sex Differences in the Regulation of Spatially Distinct Cardiac Mitochondrial Subpopulations. Mol. Cell. Biochem. 2016, 419, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Sandhi, J.; Singh, J.P.; Kaur, T.; Ghuman, S.S.; Singh, A.P. Involvement of Progesterone Receptors in Ascorbic Acid-Mediated Protection against Ischemia-Reperfusion-Induced Acute Kidney Injury. J. Surg. Res. 2014, 187, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-C.; Chang, C.-Y.; Chang, S.-T.; Chen, S.-H. 17β-Estradiol Accelerated Renal Tubule Regeneration in Male Rats After Ischemia/Reperfusion-Induced Acute Kidney Injury. Shock 2016, 46, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, E.Y.; Kazachenko, A.V.; Vyssokikh, M.Y.; Vasileva, A.K.; Tcvirkun, D.V.; Isaev, N.K.; Kirpatovsky, V.I.; Zorov, D.B. The Role of Mitochondria in Oxidative and Nitrosative Stress during Ischemia/reperfusion in the Rat Kidney. Kidney Int. 2007, 72, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Morosanova, M.A.; Plotnikov, E.Y.; Zorova, L.D.; Pevzner, I.B.; Popkov, V.A.; Silachev, D.N.; Jankauskas, S.S.; Babenko, V.A.; Zorov, D.B. Mechanisms of Inflammatory Injury of Renal Tubular Cells in a Cellular Model of Pyelonephritis. Biochemistry 2016, 81, 1240–1250. [Google Scholar] [CrossRef]
- Crowley, L.C.; Christensen, M.E.; Waterhouse, N.J. Measuring Mitochondrial Transmembrane Potential by TMRE Staining. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot087361. [Google Scholar] [CrossRef]
- Fischer, C.A.; Besora-Casals, L.; Rolland, S.G.; Haeussler, S.; Singh, K.; Duchen, M.; Conradt, B.; Marr, C. MitoSegNet: Easy-to-Use Deep Learning Segmentation for Analyzing Mitochondrial Morphology. iScience 2020, 23, 101601. [Google Scholar] [CrossRef]
- Prigent, S.; Valades-Cruz, C.A.; Leconte, L.; Salamero, J.; Kervrann, C. STracking: A Free and Open-Source Python Library for Particle Tracking and Analysis. Bioinformatics 2022, 38, 3671–3673. [Google Scholar] [CrossRef]
- Atif, F.; Yousuf, S.; Sayeed, I.; Ishrat, T.; Hua, F.; Stein, D.G. Combination Treatment with Progesterone and Vitamin D Hormone Is More Effective than Monotherapy in Ischemic Stroke: The Role of BDNF/TrkB/Erk1/2 Signaling in Neuroprotection. Neuropharmacology 2013, 67, 78–87. [Google Scholar] [CrossRef]
- Espinosa-Garcia, C.; Atif, F.; Yousuf, S.; Sayeed, I.; Neigh, G.N.; Stein, D.G. Progesterone Attenuates Stress-Induced NLRP3 Inflammasome Activation and Enhances Autophagy Following Ischemic Brain Injury. Int. J. Mol. Sci. 2020, 21, 3740. [Google Scholar] [CrossRef]
- Remus, E.W.; Sayeed, I.; Won, S.; Lyle, A.N.; Stein, D.G. Progesterone Protects Endothelial Cells after Cerebrovascular Occlusion by Decreasing MCP-1- and CXCL1-Mediated Macrophage Infiltration. Exp. Neurol. 2015, 271, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Hutchens, M.P.; Fujiyoshi, T.; Komers, R.; Herson, P.S.; Anderson, S. Estrogen Protects Renal Endothelial Barrier Function from Ischemia-Reperfusion in Vitro and in Vivo. Am. J. Physiol. Renal Physiol. 2012, 303, F377–F385. [Google Scholar] [CrossRef] [PubMed]
- Han, H.J.; Lim, M.J.; Lee, Y.J.; Kim, E.J.; Jeon, Y.J.; Lee, J.H. Effects of TCDD and Estradiol-17beta on the Proliferation and Na+/glucose Cotransporter in Renal Proximal Tubule Cells. Toxicol. In Vitro 2005, 19, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Frankish, A.; Diekhans, M.; Ferreira, A.-M.; Johnson, R.; Jungreis, I.; Loveland, J.; Mudge, J.M.; Sisu, C.; Wright, J.; Armstrong, J.; et al. GENCODE Reference Annotation for the Human and Mouse Genomes. Nucleic Acids Res. 2019, 47, D766–D773. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.; Huber, W.; Pagès, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.T.; Carey, V.J. Software for Computing and Annotating Genomic Ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrianova, N.V.; Brezgunova, A.A.; Buyan, M.I.; Makievskaya, C.I.; Buyan, A.I.; Cherkesova, K.S.; Pevzner, I.B.; Zorova, L.D.; Zorov, D.B.; Plotnikov, E.Y.; et al. Sex-Specific Effects of Estradiol and Progesterone in Ischemic Kidney Injury. Int. J. Mol. Sci. 2024, 25, 3155. https://doi.org/10.3390/ijms25063155
Andrianova NV, Brezgunova AA, Buyan MI, Makievskaya CI, Buyan AI, Cherkesova KS, Pevzner IB, Zorova LD, Zorov DB, Plotnikov EY, et al. Sex-Specific Effects of Estradiol and Progesterone in Ischemic Kidney Injury. International Journal of Molecular Sciences. 2024; 25(6):3155. https://doi.org/10.3390/ijms25063155
Chicago/Turabian StyleAndrianova, Nadezda V., Anna A. Brezgunova, Marina I. Buyan, Ciara I. Makievskaya, Andrey I. Buyan, Kseniia S. Cherkesova, Irina B. Pevzner, Ljubava D. Zorova, Dmitry B. Zorov, Egor Y. Plotnikov, and et al. 2024. "Sex-Specific Effects of Estradiol and Progesterone in Ischemic Kidney Injury" International Journal of Molecular Sciences 25, no. 6: 3155. https://doi.org/10.3390/ijms25063155