
The Role of Metabolomics and Microbiology in Urinary Tract Infection
 ,
,
Abstract
:1. Introduction
2. Human Microbiota and UTI
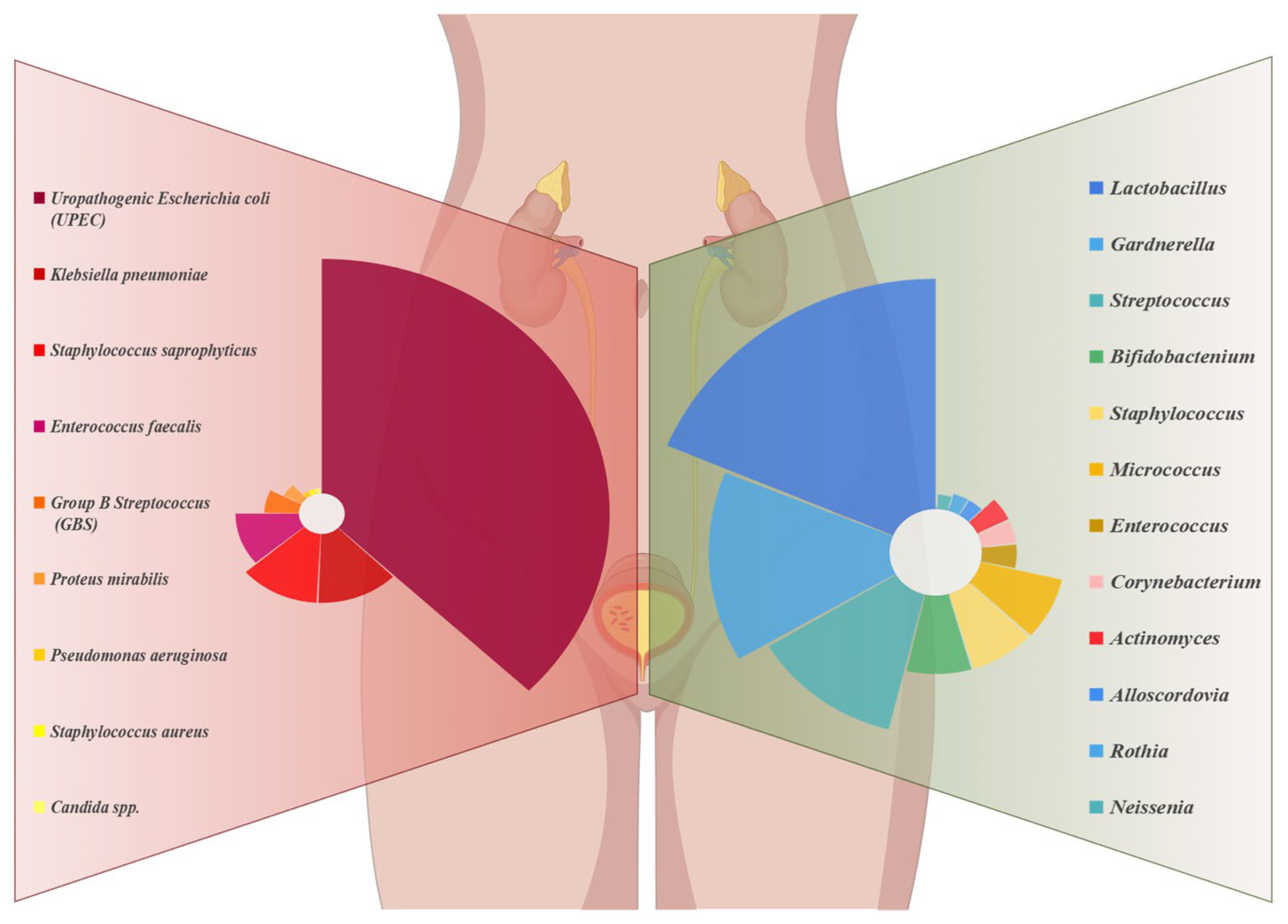
2.1. Normal Microbiological Composition of Urine
2.2. Microbiota Structure of UTI-Infected Patients
2.2.1. Changes in the Urinary Microbiome and Pathogenesis in UTIs
2.2.2. Gut Microbiota and UTI
2.2.3. Vaginal Microbiota and UTI
2.2.4. Environmentally Altered Microbiota and UTI
2.3. Relationship between Microbiome and UTI
3. Urinary Metabolome and UTI
3.1. Common Methods and Techniques for Studying Metabolites in Urine
3.2. Effect of UTIs on the Metabolome
4. Interactions and Regulatory Relationships between the Microbiome and Metabolome during UTI
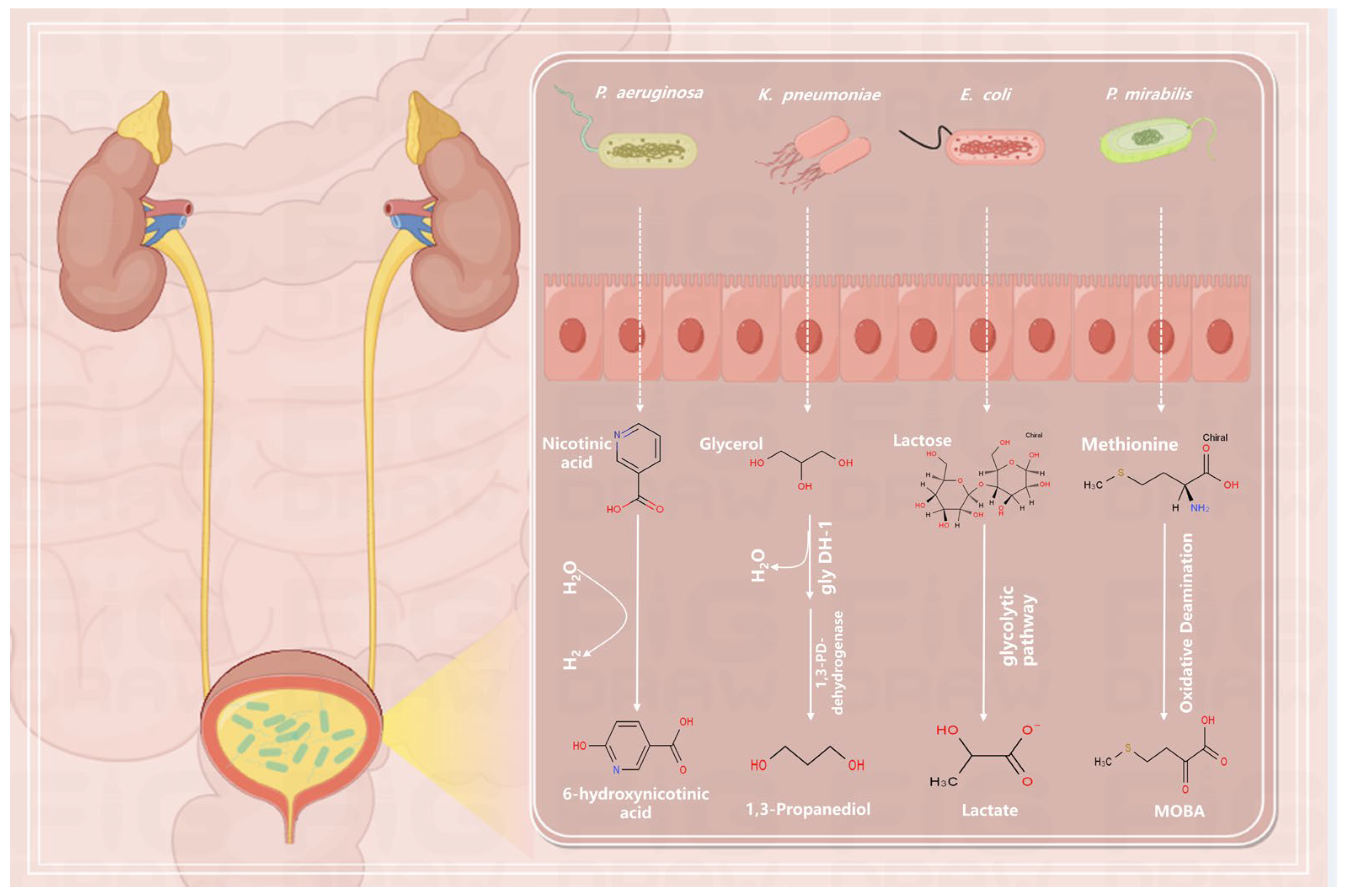
4.1. Effects of Metabolites on Microorganisms
4.2. Regulation of Metabolites by Microorganisms
4.3. The Significance of Microbiome and Metabolome Synergy in the Development and Recovery of UTIs
5. Summary and Outlook
5.1. Potential Applications of the Metabolome in the Diagnosis and Prediction of UTI
5.2. Potential Applications of the Microbiome in the Diagnosis and Prediction of UTI
5.3. Application and Advantages of Emerging Therapies in UTI Research
5.3.1. Mannoside FimH Antagonist
5.3.2. Probiotic Therapy
5.3.3. Transfer of Fecal Microbiota
5.4. Future Research Needs and Possible Challenges
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tamadonfar, K.O.; Omattage, N.S.; Spaulding, C.N.; Hultgren, S.J. Reaching the End of the Line: Urinary Tract Infections. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Asadi Karam, M.R.; Habibi, M.; Bouzari, S. Urinary Tract Infection: Pathogenicity, Antibiotic Resistance and Development of Effective Vaccines against Uropathogenic Escherichia Coli. Mol. Immunol. 2019, 108, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) Infections: Virulence Factors, Bladder Responses, Antibiotic, and Non-Antibiotic Antimicrobial Strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef]
- Gupta, K.; Bhadelia, N. Management of Urinary Tract Infections from Multidrug-Resistant Organisms. Infect. Dis. Clin. N. Am. 2014, 28, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Nielubowicz, G.R.; Mobley, H.L.T. Host–Pathogen Interactions in Urinary Tract Infection. Nat. Rev. Urol. 2010, 7, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary Tract Infections: Epidemiology, Mechanisms of Infection and Treatment Options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The Healthy Human Microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-Sensing Receptors GPR43 and GPR109A Facilitate Dietary Fibre-Induced Gut Homeostasis through Regulation of the Inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef]
- Jonsson, A.L.; Bäckhed, F. Role of Gut Microbiota in Atherosclerosis. Nat. Rev. Cardiol. 2017, 14, 79–87. [Google Scholar] [CrossRef]
- Jung, H.D.; Cho, S.; Lee, J.Y. Update on the Effect of the Urinary Microbiome on Urolithiasis. Diagnostics 2023, 13, 951. [Google Scholar] [CrossRef]
- Friedrich, V.; Choi, H.W. The Urinary Microbiome: Role in Bladder Cancer and Treatment. Diagnostics 2022, 12, 2068. [Google Scholar] [CrossRef]
- Simon, J.-C.; Marchesi, J.R.; Mougel, C.; Selosse, M.-A. Host-Microbiota Interactions: From Holobiont Theory to Analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Mann, R.; Mediati, D.G.; Duggin, I.G.; Harry, E.J.; Bottomley, A.L. Metabolic Adaptations of Uropathogenic E. coli in the Urinary Tract. Front. Cell. Infect. Microbiol. 2017, 7, 241. [Google Scholar] [CrossRef]
- Alteri, C.J.; Himpsl, S.D.; Mobley, H.L.T. Preferential Use of Central Metabolism In Vivo Reveals a Nutritional Basis for Polymicrobial Infection. PLoS Pathog. 2015, 11, e1004601. [Google Scholar] [CrossRef]
- Alteri, C.J.; Mobley, H.L. Escherichia coli Physiology and Metabolism Dictates Adaptation to Diverse Host Microenvironments. Curr. Opin. Microbiol. 2012, 15, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Hilt, E.E.; McKinley, K.; Pearce, M.M.; Rosenfeld, A.B.; Zilliox, M.J.; Mueller, E.R.; Brubaker, L.; Gai, X.; Wolfe, A.J.; Schreckenberger, P.C. Urine Is Not Sterile: Use of Enhanced Urine Culture Techniques to Detect Resident Bacterial Flora in the Adult Female Bladder. J. Clin. Microbiol. 2014, 52, 871–876. [Google Scholar] [CrossRef]
- Wolfe, A.J.; Toh, E.; Shibata, N.; Rong, R.; Kenton, K.; FitzGerald, M.; Mueller, E.R.; Schreckenberger, P.; Dong, Q.; Nelson, D.E.; et al. Evidence of Uncultivated Bacteria in the Adult Female Bladder. J. Clin. Microbiol. 2012, 50, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Fouts, D.E.; Pieper, R.; Szpakowski, S.; Pohl, H.; Knoblach, S.; Suh, M.-J.; Huang, S.-T.; Ljungberg, I.; Sprague, B.M.; Lucas, S.K.; et al. Integrated Next-Generation Sequencing of 16S rDNA and Metaproteomics Differentiate the Healthy Urine Microbiome from Asymptomatic Bacteriuria in Neuropathic Bladder Associated with Spinal Cord Injury. J. Transl. Med. 2012, 10, 174. [Google Scholar] [CrossRef]
- Pace, N.R. A Molecular View of Microbial Diversity and the Biosphere. Science 1997, 276, 734–740. [Google Scholar] [CrossRef]
- Wolfe, A.J.; Brubaker, L. “Sterile Urine” and the Presence of Bacteria. Eur. Urol. 2015, 68, 173–174. [Google Scholar] [CrossRef]
- Price, T.K.; Dune, T.; Hilt, E.E.; Thomas-White, K.J.; Kliethermes, S.; Brincat, C.; Brubaker, L.; Wolfe, A.J.; Mueller, E.R.; Schreckenberger, P.C. The Clinical Urine Culture: Enhanced Techniques Improve Detection of Clinically Relevant Microorganisms. J. Clin. Microbiol. 2016, 54, 1216–1222. [Google Scholar] [CrossRef]
- Neugent, M.L.; Hulyalkar, N.V.; Nguyen, V.H.; Zimmern, P.E.; De Nisco, N.J. Advances in Understanding the Human Urinary Microbiome and Its Potential Role in Urinary Tract Infection. mBio 2020, 11, e00218-20. [Google Scholar] [CrossRef]
- Price, T.; Hilt, E.; Thomas-White, K.; Mueller, E.; Wolfe, A.; Brubaker, L. The Urobiome of Continent Adult Women: A Cross-Sectional Study. BJOG Int. J. Obstet. Gynaecol. 2020, 127, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Komesu, Y.M.; Dinwiddie, D.L.; Richter, H.E.; Lukacz, E.S.; Sung, V.W.; Siddiqui, N.Y.; Zyczynski, H.M.; Ridgeway, B.; Rogers, R.G.; Arya, L.A.; et al. Defining the Relationship between Vaginal and Urinary Microbiomes. Am. J. Obstet. Gynecol. 2020, 222, 154.e1–154.e10. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.M.; Hilt, E.E.; Rosenfeld, A.B.; Zilliox, M.J.; Thomas-White, K.; Fok, C.; Kliethermes, S.; Schreckenberger, P.C.; Brubaker, L.; Gai, X.; et al. The Female Urinary Microbiome: A Comparison of Women with and without Urgency Urinary Incontinence. mBio 2014, 5, e01283-14. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ling, Z.; Xiao, Y.; Yang, Q.; Zheng, L.; Jiang, P.; Li, L.; Wang, W. Characterization of the Urinary Microbiota of Elderly Women and the Effects of Type 2 Diabetes and Urinary Tract Infections on the Microbiota. Oncotarget 2017, 8, 100678–100690. [Google Scholar] [CrossRef] [PubMed]
- Curtiss, N.; Balachandran, A.; Krska, L.; Peppiatt-Wildman, C.; Wildman, S.; Duckett, J. Age, Menopausal Status and the Bladder Microbiome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 228, 126–129. [Google Scholar] [CrossRef]
- Elsayed, N.S.; Wolfe, A.J.; Burk, R.D. Urine Microbiome in Individuals with an Impaired Immune System. Front. Cell. Infect. Microbiol. 2024, 13, 1308665. [Google Scholar] [CrossRef]
- Gilbert, N.M.; O’Brien, V.P.; Waller, C.; Batourina, E.; Mendelsohn, C.L.; Lewis, A.L. Gardnerella Exposures Alter Bladder Gene Expression and Augment Uropathogenic Escherichia coli Urinary Tract Infection in Mice. Front. Cell. Infect. Microbiol. 2022, 12, 909799. [Google Scholar] [CrossRef]
- Moustafa, A.; Li, W.; Singh, H.; Moncera, K.J.; Torralba, M.G.; Yu, Y.; Manuel, O.; Biggs, W.; Venter, J.C.; Nelson, K.E.; et al. Microbial Metagenome of Urinary Tract Infection. Sci. Rep. 2018, 8, 4333. [Google Scholar] [CrossRef]
- Klein, R.D.; Hultgren, S.J. Urinary Tract Infections: Microbial Pathogenesis, Host-Pathogen Interactions and New Treatment Strategies. Nat. Rev. Microbiol. 2020, 18, 211–226. [Google Scholar] [CrossRef]
- Al Mohajer, M.; Musher, D.M.; Minard, C.G.; Darouiche, R.O. Clinical significance of Staphylococcus aureus bacteriuria at a tertiary care hospital. Scand. J. Infect. Dis. 2013, 45, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Li, S.; Gan, R.-Y.; Zhou, T.; Xu, D.-P.; Li, H.-B. Impacts of Gut Bacteria on Human Health and Diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Schembri, M.A.; Nhu, N.T.K.; Phan, M.-D. Gut-bladder axis in recurrent UTI. Nat. Microbiol. 2022, 7, 601–602. [Google Scholar] [CrossRef]
- Yamamoto, S.; Tsukamoto, T.; Terai, A.; Kurazono, H.; Takeda, Y.; Yoshida, O. Genetic Evidence Supporting the Fecal-Perineal-Urethral Hypothesis in Cystitis Caused by Escherichia coli. J. Urol. 1997, 157, 1127–1129. [Google Scholar] [CrossRef] [PubMed]
- De Nisco, N.J.; Neugent, M.; Mull, J.; Chen, L.; Kuprasertkul, A.; De Souza Santos, M.; Palmer, K.L.; Zimmern, P.; Orth, K. Direct Detection of Tissue-Resident Bacteria and Chronic Inflammation in the Bladder Wall of Postmenopausal Women with Recurrent Urinary Tract Infection. J. Mol. Biol. 2019, 431, 4368–4379. [Google Scholar] [CrossRef] [PubMed]
- Magruder, M.; Sholi, A.N.; Gong, C.; Zhang, L.; Edusei, E.; Huang, J.; Albakry, S.; Satlin, M.J.; Westblade, L.F.; Crawford, C.; et al. Gut Uropathogen Abundance Is a Risk Factor for Development of Bacteriuria and Urinary Tract Infection. Nat. Commun. 2019, 10, 5521. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.R.; Muthukumar, T.; Dadhania, D.; Toussaint, N.C.; Ling, L.; Pamer, E.; Suthanthiran, M. Gut Microbial Community Structure and Complications Following Kidney Transplantation: A Pilot Study. Transplantation 2014, 98, 697–705. [Google Scholar] [CrossRef]
- Paalanne, N.; Husso, A.; Salo, J.; Pieviläinen, O.; Tejesvi, M.V.; Koivusaari, P.; Pirttilä, A.M.; Pokka, T.; Mattila, S.; Jyrkäs, J.; et al. Intestinal Microbiome as a Risk Factor for Urinary Tract Infections in Children. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1881–1891. [Google Scholar] [CrossRef]
- Worby, C.J.; Schreiber, H.L.; Straub, T.J.; Van Dijk, L.R.; Bronson, R.A.; Olson, B.S.; Pinkner, J.S.; Obernuefemann, C.L.P.; Muñoz, V.L.; Paharik, A.E.; et al. Longitudinal Multi-Omics Analyses Link Gut Microbiome Dysbiosis with Recurrent Urinary Tract Infections in Women. Nat. Microbiol. 2022, 7, 630–639. [Google Scholar] [CrossRef]
- Perez-Carrasco, V.; Soriano-Lerma, A.; Soriano, M.; Gutiérrez-Fernández, J.; Garcia-Salcedo, J.A. Urinary Microbiome: Yin and Yang of the Urinary Tract. Front. Cell. Infect. Microbiol. 2021, 11, 617002. [Google Scholar] [CrossRef]
- Stapleton, A.E.; Au-Yeung, M.; Hooton, T.M.; Fredricks, D.N.; Roberts, P.L.; Czaja, C.A.; Yarova-Yarovaya, Y.; Fiedler, T.; Cox, M.; Stamm, W.E. Randomized, Placebo-Controlled Phase 2 Trial of a Lactobacillus Crispatus Probiotic given Intravaginally for Prevention of Recurrent Urinary Tract Infection. Clin. Infect. Dis. 2011, 52, 1212–1217. [Google Scholar] [CrossRef]
- Sumati, A.; Saritha, N. Association of Urinary Tract Infection in Women with Bacterial Vaginosis. J. Glob. Infect. Dis. 2009, 1, 151. [Google Scholar] [CrossRef]
- Gilbert, N.M.; O’Brien, V.P.; Lewis, A.L. Transient Microbiota Exposures Activate Dormant Escherichia coli Infection in the Bladder and Drive Severe Outcomes of Recurrent Disease. PLoS Pathog. 2017, 13, e1006238. [Google Scholar] [CrossRef]
- Guiton, P.S.; Hannan, T.J.; Ford, B.; Caparon, M.G.; Hultgren, S.J. Enterococcus faecalis Overcomes Foreign Body-Mediated Inflammation to Establish Urinary Tract Infections. Infect. Immun. 2013, 81, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Pinkner, J.S.; Caparon, M.G.; Hultgren, S.J. EbpA Vaccine Antibodies Block Binding of Enterococcus faecalis to Fibrinogen to Prevent Catheter-Associated Bladder Infection in Mice. Sci. Transl. Med. 2014, 6, 254ra127. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Flores-Mireles, A.L.; Cusumano, Z.T.; Takagi, E.; Hultgren, S.J.; Caparon, M.G. Host and Bacterial Proteases Influence Biofilm Formation and Virulence in a Murine Model of Enterococcal Catheter-Associated Urinary Tract Infection. Npj Biofilms Microbiomes 2017, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Weiner-Lastinger, L.M.; Abner, S.; Edwards, J.R.; Kallen, A.J.; Karlsson, M.; Magill, S.S.; Pollock, D.; See, I.; Soe, M.M.; Walters, M.S.; et al. Antimicrobial-Resistant Pathogens Associated with Adult Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network, 2015–2017. Infect. Control Hosp. Epidemiol. 2020, 41, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.N.; Flores-Mireles, A.L.; Pinkner, C.L.; Schreiber, H.L.; Joens, M.S.; Park, A.M.; Potretzke, A.M.; Bauman, T.M.; Pinkner, J.S.; Fitzpatrick, J.A.J.; et al. Catheterization Alters Bladder Ecology to Potentiate Staphylococcus aureus Infection of the Urinary Tract. Proc. Natl. Acad. Sci. USA 2017, 114, E8721–E8730. [Google Scholar] [CrossRef] [PubMed]
- Bichler, K.-H.; Eipper, E.; Naber, K.; Braun, V.; Zimmermann, R.; Lahme, S. Urinary Infection Stones. Int. J. Antimicrob. Agents 2002, 19, 488–498. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Prenovost, K.; Mobley, H.L.T.; Mody, L. How Often Do Clinically Diagnosed Catheter-Associated Urinary Tract Infections in Nursing Homes Meet Standardized Criteria? J. Am. Geriatr. Soc. 2017, 65, 395–401. [Google Scholar] [CrossRef]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and Urinary Tract Infections. Microbiol. Spectr. 2015, 3, 383–433. [Google Scholar] [CrossRef] [PubMed]
- Stickler, D.J. Clinical Complications of Urinary Catheters Caused by Crystalline Biofilms: Something Needs to Be Done. J. Intern. Med. 2014, 276, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Giovannetti, O.; Tomalty, D.; Velikonja, L.; Gray, G.; Boev, N.; Gilmore, S.; Oladipo, J.; Sjaarda, C.; Sheth, P.M.; Adams, M.A. Pre- and Post-LEEP: Analysis of the Female Urogenital Tract Microenvironment and Its Association with Sexual Dysfunction. Sex. Med. 2023, 11, qfad039. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.E.; Van Der Pol, B.; Dong, Q.; Revanna, K.V.; Fan, B.; Easwaran, S.; Sodergren, E.; Weinstock, G.M.; Diao, L.; Fortenberry, J.D. Characteristic Male Urine Microbiomes Associate with Asymptomatic Sexually Transmitted Infection. PLoS ONE 2010, 5, e14116. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, A.E. The Vaginal Microbiota and Urinary Tract Infection. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-J.; Shin, H.; Song, J.; Kim, M.; Yun, J.; Kim, Z.; Lee, Y.; Lee, S.; Lee, K.; Kim, W.; et al. Urinary Microbiome Characteristics in Female Patients with Acute Uncomplicated Cystitis and Recurrent Cystitis. J. Clin. Med. 2021, 10, 1097. [Google Scholar] [CrossRef] [PubMed]
- Jeney, S.E.S.; Lane, F.; Oliver, A.; Whiteson, K.; Dutta, S. Fecal Microbiota Transplantation for the Treatment of Refractory Recurrent Urinary Tract Infection. Obstet. Gynecol. 2020, 136, 771–773. [Google Scholar] [CrossRef]
- Nickel, J.C.; Stephens, A.; Landis, J.R.; Mullins, C.; Van Bokhoven, A.; Anger, J.T.; Ackerman, A.L.; Kim, J.; Sutcliffe, S.; Krol, J.E.; et al. Urinary Fungi Associated with Urinary Symptom Severity among Women with Interstitial Cystitis/Bladder Pain Syndrome (IC/BPS). World J. Urol. 2020, 38, 433–446. [Google Scholar] [CrossRef]
- Santiago-Rodriguez, T.M.; Ly, M.; Bonilla, N.; Pride, D.T. The Human Urine Virome in Association with Urinary Tract Infections. Front. Microbiol. 2015, 6, 14. [Google Scholar] [CrossRef]
- Josephs-Spaulding, J.; Krogh, T.J.; Rettig, H.C.; Lyng, M.; Chkonia, M.; Waschina, S.; Graspeuntner, S.; Rupp, J.; Møller-Jensen, J.; Kaleta, C. Recurrent Urinary Tract Infections: Unraveling the Complicated Environment of Uncomplicated rUTIs. Front. Cell. Infect. Microbiol. 2021, 11, 562525. [Google Scholar] [CrossRef]
- Thomas-White, K.J.; Gao, X.; Lin, H.; Fok, C.S.; Ghanayem, K.; Mueller, E.R.; Dong, Q.; Brubaker, L.; Wolfe, A.J. Urinary Microbes and Postoperative Urinary Tract Infection Risk in Urogynecologic Surgical Patients. Int. Urogynecol. J. 2018, 29, 1797–1805. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Anger, J.; Lee, U.; Ackerman, A.L.; Chou, R.; Chughtai, B.; Clemens, J.Q.; Hickling, D.; Kapoor, A.; Kenton, K.S.; Kaufman, M.R.; et al. Recurrent Uncomplicated Urinary Tract Infections in Women: AUA/CUA/SUFU Guideline. J. Urol. 2019, 202, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.; Traynor, P.; Hopson, D. Evaluation of Microbiological Processing of Urine Specimens: Comparison of Overnight versus Two-Day Incubation. J. Clin. Microbiol. 1992, 30, 1600–1601. [Google Scholar] [CrossRef]
- Nanda, N.; Juthani-Mehta, M. Novel Biomarkers for the Diagnosis of Urinary Tract Infection—A Systematic Review. Biomark. Insights 2009, 4, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Lenz, E.M.; Bright, J.; Wilson, I.D.; Morgan, S.R.; Nash, A.F.P. A 1H NMR-Based Metabonomic Study of Urine and Plasma Samples Obtained from Healthy Human Subjects. J. Pharm. Biomed. Anal. 2003, 33, 1103–1115. [Google Scholar] [CrossRef]
- Wang, T.; Liu, J.; Luo, X.; Hu, L.; Lu, H. Functional Metabolomics Innovates Therapeutic Discovery of Traditional Chinese Medicine Derived Functional Compounds. Pharmacol. Ther. 2021, 224, 107824. [Google Scholar] [CrossRef]
- Lam, C.-W.; Law, C.-Y.; Sze, K.-H.; To, K.K.-W. Quantitative Metabolomics of Urine for Rapid Etiological Diagnosis of Urinary Tract Infection: Evaluation of a Microbial–Mammalian Co-Metabolite as a Diagnostic Biomarker. Clin. Chim. Acta 2015, 438, 24–28. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wu, X.; Wang, X. Urine metabolomics. Clin. Chim. Acta 2012, 414, 65–69. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X. Recent and Potential Developments of Biofluid Analyses in Metabolomics. J. Proteom. 2012, 75, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zheng, S.; Li, M.; Liu, X.; Sun, H.; Guo, Z.; Wei, J.; Jia, L.; Sun, W. A Comprehensive 2D-LC/MS/MS Profile of the Normal Human Urinary Metabolome. Diagnostics 2022, 12, 2184. [Google Scholar] [CrossRef]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The Human Urine Metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef] [PubMed]
- Fadil, F.; Samol, C.; Berger, R.S.; Kellermeier, F.; Gronwald, W.; Oefner, P.J.; Dettmer, K. Isotope Ratio Outlier Analysis (IROA) for HPLC-TOFMS-Based Metabolomics of Human Urine. Metabolites 2022, 12, 741. [Google Scholar] [CrossRef]
- Qin, S.; Xin, G.; Wei, J.; He, G.; Yuan, Z.; Liu, H.; Zhang, X.; Wang, Y.; Zhang, W.; Lu, J. Metabolic Profiles of 5F-MDMB-PICA in Human Urine, Serum and Hair Samples Using LC-Q Exactive HF-MS. J. Anal. Toxicol. 2022, 46, 408–420. [Google Scholar] [CrossRef]
- Klepacki, J.; Klawitter, J.; Klawitter, J.; Thurman, J.M.; Christians, U. A High-Performance Liquid Chromatography-Tandem Mass Spectrometry-Based Targeted Metabolomics Kidney Dysfunction Marker Panel in Human Urine. Clin. Chim. Acta 2015, 446, 43–53. [Google Scholar] [CrossRef]
- Jiménez Girón, A.; Deventer, K.; Roels, K.; Van Eenoo, P. Development and Validation of an Open Screening Method for Diuretics, Stimulants and Selected Compounds in Human Urine by UHPLC-HRMS for Doping Control. Anal. Chim. Acta 2012, 721, 137–146. [Google Scholar] [CrossRef]
- Byeon, S.K.; Kim, J.Y.; Lee, J.-S.; Moon, M.H. Variations in Plasma and Urinary Lipids in Response to Enzyme Replacement Therapy for Fabry Disease Patients by Nanoflow UPLC-ESI-MS/MS. Anal. Bioanal. Chem. 2016, 408, 2265–2274. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque Cavalcanti, G.; Carius Garrido, B.; Dias Leal, F.; Costa Padilha, M.; Mazzarino, M.; De La Torre, X.; Botre, F.; Radler De Aquino Neto, F. Detection of New Exemestane Metabolites by Liquid Chromatography Interfaced to Electrospray-Tandem Mass Spectrometry. J. Steroid Biochem. Mol. Biol. 2011, 127, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, M.J.; Powers, R. Multiplatform untargeted metabolomics. Magn. Reson. Chem. 2023, 61, 628–653. [Google Scholar] [CrossRef]
- Reitzer, L.; Zimmern, P. Rapid Growth and Metabolism of Uropathogenic Escherichia coli in Relation to Urine Composition. Clin. Microbiol. Rev. 2019, 33, e00101-19. [Google Scholar] [CrossRef]
- Ronald, A. The Etiology of Urinary Tract Infection: Traditional and Emerging Pathogens. Am. J. Med. 2002, 113, 14–19. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Han, Y.; Yuan, Y.; Wang, P.; Song, G.; Yuan, X.; Zhang, M.; Xie, N.; Wang, X. Exploratory Urinary Metabolic Biomarkers and Pathways Using UPLC-Q-TOF-HDMS Coupled with Pattern Recognition Approach. Analyst 2012, 137, 4200. [Google Scholar] [CrossRef] [PubMed]
- Puebla-Barragan, S.; Renaud, J.; Sumarah, M.; Reid, G. Malodorous Biogenic Amines in Escherichia coli-Caused Urinary Tract Infections in Women-a Metabolomics Approach. Sci. Rep. 2020, 10, 9703. [Google Scholar] [CrossRef] [PubMed]
- Nevedomskaya, E.; Pacchiarotta, T.; Artemov, A.; Meissner, A.; Van Nieuwkoop, C.; Van Dissel, J.T.; Mayboroda, O.A.; Deelder, A.M. (1)H NMR-Based Metabolic Profiling of Urinary Tract Infection: Combining Multiple Statistical Models and Clinical Data. Metabolomics 2012, 8, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.-W.; Law, C.-Y.; To, K.K.-W.; Cheung, S.K.-K.; Lee, K.; Sze, K.-H.; Leung, K.-F.; Yuen, K.-Y. NMR-Based Metabolomic Urinalysis: A Rapid Screening Test for Urinary Tract Infection. Clin. Chim. Acta 2014, 436, 217–223. [Google Scholar] [CrossRef]
- Hryckowian, A.J.; Baisa, G.A.; Schwartz, K.J.; Welch, R.A. dsdA Does Not Affect Colonization of the Murine Urinary Tract by Escherichia coli CFT073. PLoS ONE 2015, 10, e0138121. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, G.A. Genetics of pentose-phosphate pathway enzymes of Escherichia coli K-12. Arch. Microbiol. 1995, 164, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Alteri, C.J.; Smith, S.N.; Mobley, H.L.T. Fitness of Escherichia coli during Urinary Tract Infection Requires Gluconeogenesis and the TCA Cycle. PLoS Pathog. 2009, 5, e1000448. [Google Scholar] [CrossRef] [PubMed]
- Peekhaus, N.; Conway, T. What’s for Dinner?: Entner-Doudoroff Metabolism in Escherichia coli. J. Bacteriol. 1998, 180, 3495–3502. [Google Scholar] [CrossRef]
- Henderson, J.P.; Crowley, J.R.; Pinkner, J.S.; Walker, J.N.; Tsukayama, P.; Stamm, W.E.; Hooton, T.M.; Hultgren, S.J. Quantitative Metabolomics Reveals an Epigenetic Blueprint for Iron Acquisition in Uropathogenic Escherichia coli. PLoS Pathog. 2009, 5, e1000305. [Google Scholar] [CrossRef]
- Su, Q.; Guan, T.; Lv, H. Siderophore Biosynthesis Coordinately Modulated the Virulence-Associated Interactive Metabolome of Uropathogenic Escherichia coli and Human Urine. Sci. Rep. 2016, 6, 24099. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Nie, W.; Lv, H. Metabolic Phenotyping of the Yersinia High-Pathogenicity Island That Regulates Central Carbon Metabolism. Analyst 2015, 140, 3356–3361. [Google Scholar] [CrossRef] [PubMed]
- 1H NMR Spectroscopy in the Diagnosis of Pseudomonas Aeruginosa-Induced Urinary Tract Infection—Gupta—2005—NMR in Biomedicine—Wiley Online Library. Available online: https://analyticalsciencejournals.onlinelibrary.wiley.com/doi/10.1002/nbm.957 (accessed on 24 October 2023).
- Gupta, A.; Dwivedi, M.; Gowda, G.A.N.; Mahdi, A.A.; Jain, A.; Ayyagari, A.; Roy, R.; Bhandari, M.; Khetrapal, C.L. 1H NMR Spectroscopy in the Diagnosis of Klebsiella Pneumoniae-Induced Urinary Tract Infection. NMR Biomed. 2006, 19, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Dwivedi, M.; Mahdi, A.A.; Gowda, G.A.N.; Khetrapal, C.L.; Bhandari, M. 1H-Nuclear Magnetic Resonance Spectroscopy for Identifying and Quantifying Common Uropathogens: A Metabolic Approach to the Urinary Tract Infection. BJU Int. 2009, 104, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Ahrodia, T.; Das, S.; Bakshi, S.; Das, B. Structure, Functions, and Diversity of the Healthy Human Microbiome. Prog. Mol. Biol. Transl. Sci. 2022, 191, 53–82. [Google Scholar] [CrossRef]
- Sintsova, A.; Frick-Cheng, A.E.; Smith, S.; Pirani, A.; Subashchandrabose, S.; Snitkin, E.S.; Mobley, H. Genetically Diverse Uropathogenic Escherichia Coli Adopt a Common Transcriptional Program in Patients with UTIs. eLife 2019, 8, e49748. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.C.Y.; Lewis, I.A. Role of metabolism in uropathogenic Escherichia coli. Trends Microbiol. 2022, 30, 1174–1204. [Google Scholar] [CrossRef]
- A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing|Nature. Available online: https://www.nature.com/articles/nature08821 (accessed on 26 October 2023).
- Shannon, M.B.; Limeira, R.; Johansen, D.; Gao, X.; Lin, H.; Dong, Q.; Wolfe, A.J.; Mueller, E.R. Bladder Urinary Oxygen Tension Is Correlated with Urinary Microbiota Composition. Int. Urogynecology J. 2019, 30, 1261–1267. [Google Scholar] [CrossRef]
- Hull, R.A.; Hull, S.I. Nutritional Requirements for Growth of Uropathogenic Escherichia coli in Human Urine. Infect. Immun. 1997, 65, 1960–1961. [Google Scholar] [CrossRef]
- Forsyth, V.S.; Armbruster, C.E.; Smith, S.N.; Pirani, A.; Springman, A.C.; Walters, M.S.; Nielubowicz, G.R.; Himpsl, S.D.; Snitkin, E.S.; Mobley, H.L.T. Rapid Growth of Uropathogenic Escherichia coli during Human Urinary Tract Infection. mBio 2018, 9, e00186-18. [Google Scholar] [CrossRef] [PubMed]
- Olukoya, D.K. Nutritional Variation in Escherichia coli. Microbiology 1986, 132, 3231–3234. [Google Scholar] [CrossRef] [PubMed]
- Hynes, W.L.; Walton, S.L. Hyaluronidases of Gram-positive bacteria. FEMS Microbiol. Lett. 2000, 183, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Kamochi, R.; Oiki, S.; Murata, K.; Hashimoto, W. Probiotics in human gut microbiota can degrade host glycosaminoglycans. Sci. Rep. 2018, 8, 10674. [Google Scholar] [CrossRef]
- Zúñiga, M.; Monedero, V.; Yebra, M.J. Utilization of Host-Derived Glycans by Intestinal Lactobacillus and Bifidobacterium Species. Front. Microbiol. 2018, 9, 1917. [Google Scholar] [CrossRef]
- Dunstan, R.H.; Sparkes, D.L.; Macdonald, M.M.; De Jonge, X.J.; Dascombe, B.J.; Gottfries, J.; Gottfries, C.-G.; Roberts, T.K. Diverse Characteristics of the Urinary Excretion of Amino Acids in Humans and the Use of Amino Acid Supplementation to Reduce Fatigue and Sub-Health in Adults. Nutr. J. 2017, 16, 19. [Google Scholar] [CrossRef]
- Alteri, C.J.; Mobley, H.L.T. Metabolism and Fitness of Urinary Tract Pathogens. Microbiol. Spectr. 2015, 3, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Justice, S.S.; Hung, C.; Theriot, J.A.; Fletcher, D.A.; Anderson, G.G.; Footer, M.J.; Hultgren, S.J. Differentiation and Developmental Pathways of Uropathogenic Escherichia coli in Urinary Tract Pathogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 1333–1338. [Google Scholar] [CrossRef]
- Anderson, G.; Dodson, K.; Hooton, T.; Hultgren, S. Intracellular Bacterial Communities of Uropathogenic Escherichia coli in Urinary Tract Pathogenesis. Trends Microbiol. 2004, 12, 424–430. [Google Scholar] [CrossRef]
- Conover, M.S.; Hadjifrangiskou, M.; Palermo, J.J.; Hibbing, M.E.; Dodson, K.W.; Hultgren, S.J. Metabolic Requirements of Escherichia coli in Intracellular Bacterial Communities during Urinary Tract Infection Pathogenesis. mBio 2016, 7, e00104-16. [Google Scholar] [CrossRef]
- Ma, J.; Cai, X.; Bao, Y.; Yao, H.; Li, G. Uropathogenic F Preferentially Utilize Metabolites in Urine for Nucleotide Biosynthesis through Salvage Pathways. Int. J. Med. Microbiol. 2018, 308, 990–999. [Google Scholar] [CrossRef]
- Wick, L.M.; Egli, T. Molecular Components of Physiological Stress Responses in Escherichia coli. Adv. Biochem. Eng. Biotechnol. 2004, 89, 1–45. [Google Scholar] [CrossRef]
- Anfora, A.T.; Halladin, D.K.; Haugen, B.J.; Welch, R.A. Uropathogenic Escherichia coli CFT073 Is Adapted to Acetatogenic Growth but Does Not Require Acetate during Murine Urinary Tract Infection. Infect. Immun. 2008, 76, 5760–5767. [Google Scholar] [CrossRef]
- Enjalbert, B.; Millard, P.; Dinclaux, M.; Portais, J.-C.; Létisse, F. Acetate Fluxes in Escherichia coli Are Determined by the Thermodynamic Control of the Pta-AckA Pathway. Sci. Rep. 2017, 7, 42135. [Google Scholar] [CrossRef] [PubMed]
- Salazar, A.M.; Neugent, M.L.; De Nisco, N.J.; Mysorekar, I.U. Gut-Bladder Axis Enters the Stage: Implication for Recurrent Urinary Tract Infections. Cell Host Microbe 2022, 30, 1066–1069. [Google Scholar] [CrossRef]
- Hagan, E.C.; Lloyd, A.L.; Rasko, D.A.; Faerber, G.J.; Mobley, H.L.T. Escherichia coli Global Gene Expression in Urine from Women with Urinary Tract Infection. PLoS Pathog. 2010, 6, e1001187. [Google Scholar] [CrossRef] [PubMed]
- Neugent, M.L.; Kumar, A.; Hulyalkar, N.V.; Lutz, K.C.; Nguyen, V.H.; Fuentes, J.L.; Zhang, C.; Nguyen, A.; Sharon, B.M.; Kuprasertkul, A.; et al. Recurrent Urinary Tract Infection and Estrogen Shape the Taxonomic Ecology and Function of the Postmenopausal Urogenital Microbiome. Cell Rep. Med. 2022, 3, 100753. [Google Scholar] [CrossRef]
- Grobeisen-Duque, O.; Mora-Vargas, C.D.; Aguilera-Arreola, M.G.; Helguera-Repetto, A.C. Cycle Biodynamics of Women’s Microbiome in the Urinary and Reproductive Systems. J. Clin. Med. 2023, 12, 4003. [Google Scholar] [CrossRef] [PubMed]
- Kwok, M.; McGeorge, S.; Mayer-Coverdale, J.; Graves, B.; Paterson, D.L.; Harris, P.N.A.; Esler, R.; Dowling, C.; Britton, S.; Roberts, M.J. Guideline of Guidelines: Management of Recurrent Urinary Tract Infections in Women. BJU Int. 2022, 130, 11–22. [Google Scholar] [CrossRef]
- Nicolle, L.E. Urinary Tract Infections in Special Populations: Diabetes, Renal Transplant, HIV Infection, and Spinal Cord Injury. Infect. Dis. Clin. N. Am. 2014, 28, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Sanford, J.P. Urinary Tract Symptoms and Infections. Annu. Rev. Med. 1975, 26, 485–498. [Google Scholar] [CrossRef]
- Hunt, B.C.; Brix, V.; Vath, J.; Guterman, B.L.; Taddei, S.M.; Learman, B.S.; Brauer, A.L.; Shen, S.; Qu, J.; Armbruster, C.E. Metabolic Interplay between Proteus mirabilis and Enterococcus faecalis Facilitates Polymicrobial Biofilm Formation and Invasive Disease. bioRxiv 2023. [Google Scholar] [CrossRef]
- Siddiqui, H.; Lagesen, K.; Nederbragt, A.J.; Jeansson, S.L.; Jakobsen, K.S. Alterations of Microbiota in Urine from Women with Interstitial Cystitis. BMC Microbiol. 2012, 12, 205. [Google Scholar] [CrossRef]
- Vejborg, R.M.; De Evgrafov, M.R.; Phan, M.D.; Totsika, M.; Schembri, M.A.; Hancock, V. Identification of Genes Important for Growth of Asymptomatic Bacteriuria Escherichia coli in Urine. Infect. Immun. 2012, 80, 3179–3188. [Google Scholar] [CrossRef]
- Kenneally, C.; Murphy, C.P.; Sleator, R.D.; Culligan, E.P. The Urinary Microbiome and Biological Therapeutics: Novel Therapies for Urinary Tract Infections. Microbiol. Res. 2022, 259, 127010. [Google Scholar] [CrossRef]
- Jutley, G.S.; Young, S.P. Metabolomics to Identify Biomarkers and as a Predictive Tool in Inflammatory Diseases. Best Pract. Res. Clin. Rheumatol. 2015, 29, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Lu, H.; Lee, Y.H. Challenges and Emergent Solutions for LC-MS/MS Based Untargeted Metabolomics in Diseases. Mass Spectrom. Rev. 2018, 37, 772–792. [Google Scholar] [CrossRef]
- Yarbrough, M.L.; Briden, K.E.; Mitsios, J.V.; Weindel, A.L.; Terrill, C.M.; Hunstad, D.A.; Dietzen, D.J. Mass Spectrometric Measurement of Urinary Kynurenine-to-Tryptophan Ratio in Children with and without Urinary Tract Infection. Clin. Biochem. 2018, 56, 83–88. [Google Scholar] [CrossRef]
- Satink, H.P.W.M.; Hessels, J.; Kingma, A.W.; Van Den Berg, G.A.; Muskiet, F.A.J.; Halie, M.R. Microbial Influences on Urinary Polyamine Excretion. Clin. Chim. Acta 1989, 179, 305–314. [Google Scholar] [CrossRef]
- Gregson, D.B.; Wildman, S.D.; Chan, C.C.Y.; Bihan, D.G.; Groves, R.A.; Rydzak, T.; Pittman, K.; Lewis, I.A. Metabolomics Strategy for Diagnosing Urinary Tract Infections; Infectious Diseases (except HIV/AIDS). medRxiv 2021. [Google Scholar] [CrossRef]
- Ceprnja, M.; Oros, D.; Melvan, E.; Svetlicic, E.; Skrlin, J.; Barisic, K.; Starcevic, L.; Zucko, J.; Starcevic, A. Modeling of Urinary Microbiota Associated with Cystitis. Front. Cell. Infect. Microbiol. 2021, 11, 643638. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, C.; Hultgren, S. Adhesive Pili in UTI Pathogenesis and Drug Development. Pathogens 2016, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, J.; Berglund, J.; Schembri, M.; De Genst, E.; Cools, L.; Wuhrer, M.; Hung, C.; Pinkner, J.; Slättegård, R.; Zavialov, A.; et al. Receptor Binding Studies Disclose a Novel Class of High-Affinity Inhibitors of the Escherichia coli FimH Adhesin. Mol. Microbiol. 2005, 55, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, C.N.; Klein, R.D.; Ruer, S.; Kau, A.L.; Schreiber, H.L.; Cusumano, Z.T.; Dodson, K.W.; Pinkner, J.S.; Fremont, D.H.; Janetka, J.W.; et al. Selective Depletion of Uropathogenic E. coli from the Gut by a FimH Antagonist. Nature 2017, 546, 528–532. [Google Scholar] [CrossRef]
- Mydock-McGrane, L.K.; Cusumano, Z.T.; Janetka, J.W. Mannose-Derived FimH Antagonists: A Promising Anti-Virulence Therapeutic Strategy for Urinary Tract Infections and Crohn’s Disease. Expert Opin. Ther. Pat. 2016, 26, 175–197. [Google Scholar] [CrossRef]
- Jarvis, C.; Han, Z.; Kalas, V.; Klein, R.; Pinkner, J.S.; Ford, B.; Binkley, J.; Cusumano, C.K.; Cusumano, Z.; Mydock-McGrane, L.; et al. Antivirulence Isoquinolone Mannosides: Optimization of the Biaryl Aglycone for FimH Lectin Binding Affinity and Efficacy in the Treatment of Chronic UTI. ChemMedChem 2016, 11, 367–373. [Google Scholar] [CrossRef]
- Sihra, N.; Goodman, A.; Zakri, R.; Sahai, A.; Malde, S. Nonantibiotic Prevention and Management of Recurrent Urinary Tract Infection. Nat. Rev. Urol. 2018, 15, 750–776. [Google Scholar] [CrossRef]
- Grosen, A.K.; Povlsen, J.V.; Lemming, L.E.; Jørgensen, S.M.D.; Dahlerup, J.F.; Hvas, C.L. Faecal Microbiota Transplantation Eradicated Extended-Spectrum Beta-Lactamase-Producing Klebsiella Pneumoniae from a Renal Transplant Recipient with Recurrent Urinary Tract Infections. Case Rep. Nephrol. Dial. 2019, 9, 102–107. [Google Scholar] [CrossRef]
- Smith, A.L.; Brown, J.; Wyman, J.F.; Berry, A.; Newman, D.K.; Stapleton, A.E. Treatment and Prevention of Recurrent Lower Urinary Tract Infections in Women: A Rapid Review with Practice Recommendations. J. Urol. 2018, 200, 1174–1191. [Google Scholar] [CrossRef]
- Caretto, M.; Giannini, A.; Russo, E.; Simoncini, T. Preventing Urinary Tract Infections after Menopause without Antibiotics. Maturitas 2017, 99, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Murphy, C.P.; Sleator, R.D.; Culligan, E.P. The Urobiome, Urinary Tract Infections, and the Need for Alternative Therapeutics. Microb. Pathog. 2021, 161, 105295. [Google Scholar] [CrossRef] [PubMed]
- Van Nood, E.; Vrieze, A.; Nieuwdorp, M.; Fuentes, S.; Zoetendal, E.G.; De Vos, W.M.; Visser, C.E.; Kuijper, E.J.; Bartelsman, J.F.W.M.; Tijssen, J.G.P.; et al. Duodenal Infusion of Donor Feces for Recurrent Clostridium Difficile. N. Engl. J. Med. 2013, 368, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Lawley, T.D.; Walker, A.W. Intestinal Colonization Resistance. Immunology 2013, 138, 1–11. [Google Scholar] [CrossRef]
- Xu, R.; Deebel, N.; Casals, R.; Dutta, R.; Mirzazadeh, M. A New Gold Rush: A Review of Current and Developing Diagnostic Tools for Urinary Tract Infections. Diagnostics 2021, 11, 479. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Main Virulence Factors | Function |
|---|---|---|
| Enterococcus | Ebp pili | Fibrinogen-binding adhesin of EbpA binds to fibrinogen of encapsulated catheters |
| SprE | Secreted proteases (GelE and SprE) to cleave fibrinogen to enhance biofilm formation | |
| GelE | ||
| Staphylococcus | ClfB | Binding fibrinogen, thereby promoting bladder and duct colonization |
| Proteus | MR/P fimbriae | Adhesion to the bladder establishes infection |
| Urease | Hydrolysis of urea to ammonia, formation of crystalline biofilms, provision of protective ecological niches and promotion of relapse | |
| Escherichia | FimH | FimH adhesin binds fibrinogen covering the duct |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Wang, C.; Yu, X.; Zheng, W.; An, Y.; Zhang, J.; Zhang, Y.; Wang, G.; Qi, M.; Lin, H.; et al. The Role of Metabolomics and Microbiology in Urinary Tract Infection. Int. J. Mol. Sci. 2024, 25, 3134. https://doi.org/10.3390/ijms25063134
Zheng H, Wang C, Yu X, Zheng W, An Y, Zhang J, Zhang Y, Wang G, Qi M, Lin H, et al. The Role of Metabolomics and Microbiology in Urinary Tract Infection. International Journal of Molecular Sciences. 2024; 25(6):3134. https://doi.org/10.3390/ijms25063134
Chicago/Turabian StyleZheng, Haoyu, Chao Wang, Xiao Yu, Wenxue Zheng, Yiming An, Jiaqi Zhang, Yuhan Zhang, Guoqiang Wang, Mingran Qi, Hongqiang Lin, and et al. 2024. "The Role of Metabolomics and Microbiology in Urinary Tract Infection" International Journal of Molecular Sciences 25, no. 6: 3134. https://doi.org/10.3390/ijms25063134