
Genome-Wide Association Analysis of Effective Tillers in Rice under Different Nitrogen Gradients
 and
and
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Variation
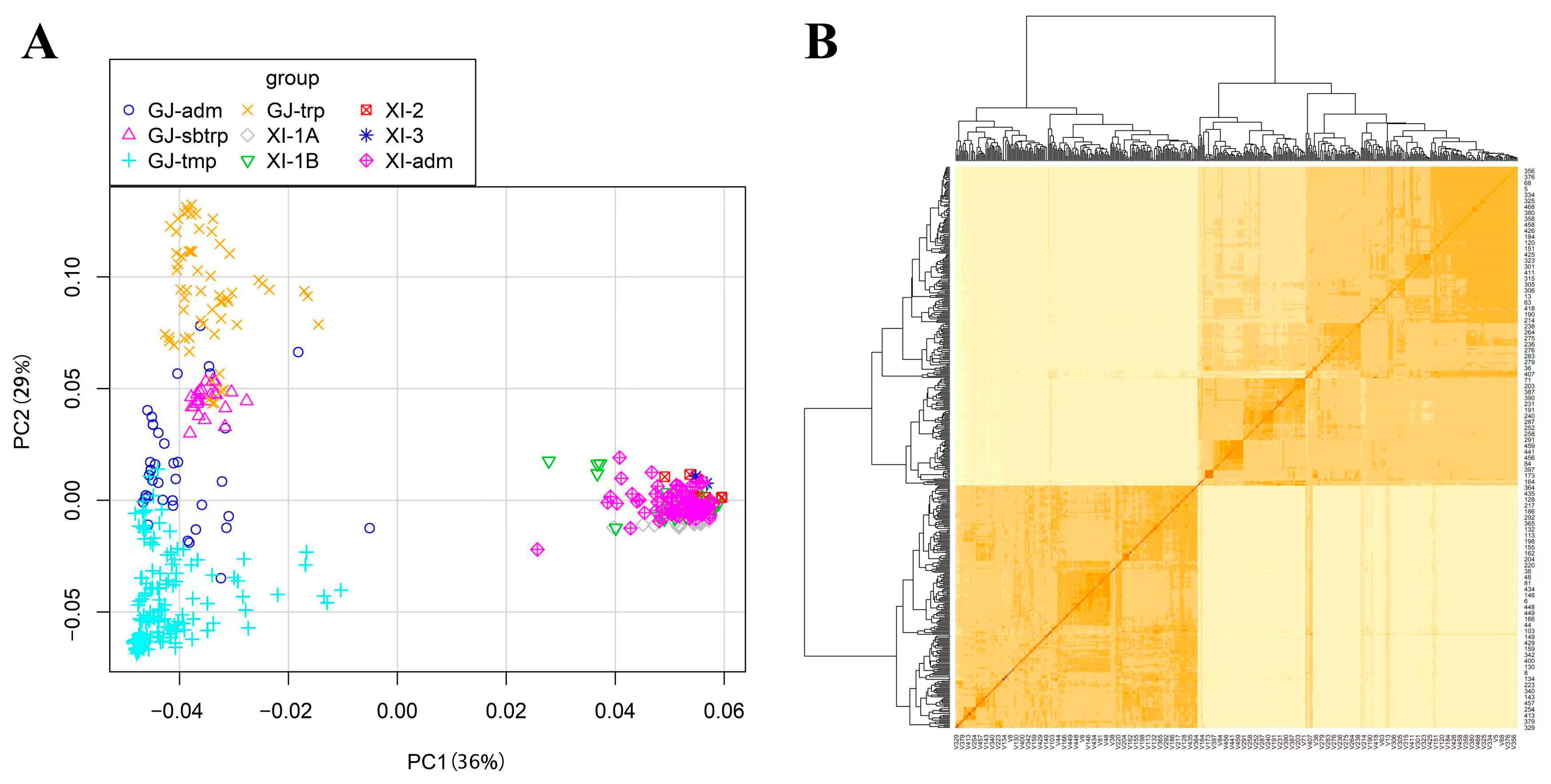
2.2. SNP Validation and Population Structure Analysis
2.3. ETN-Related QTL Detected by GWAS
2.4. ETN-Related QTL Co-Localized with Previously Reported Rice Tillering Genes
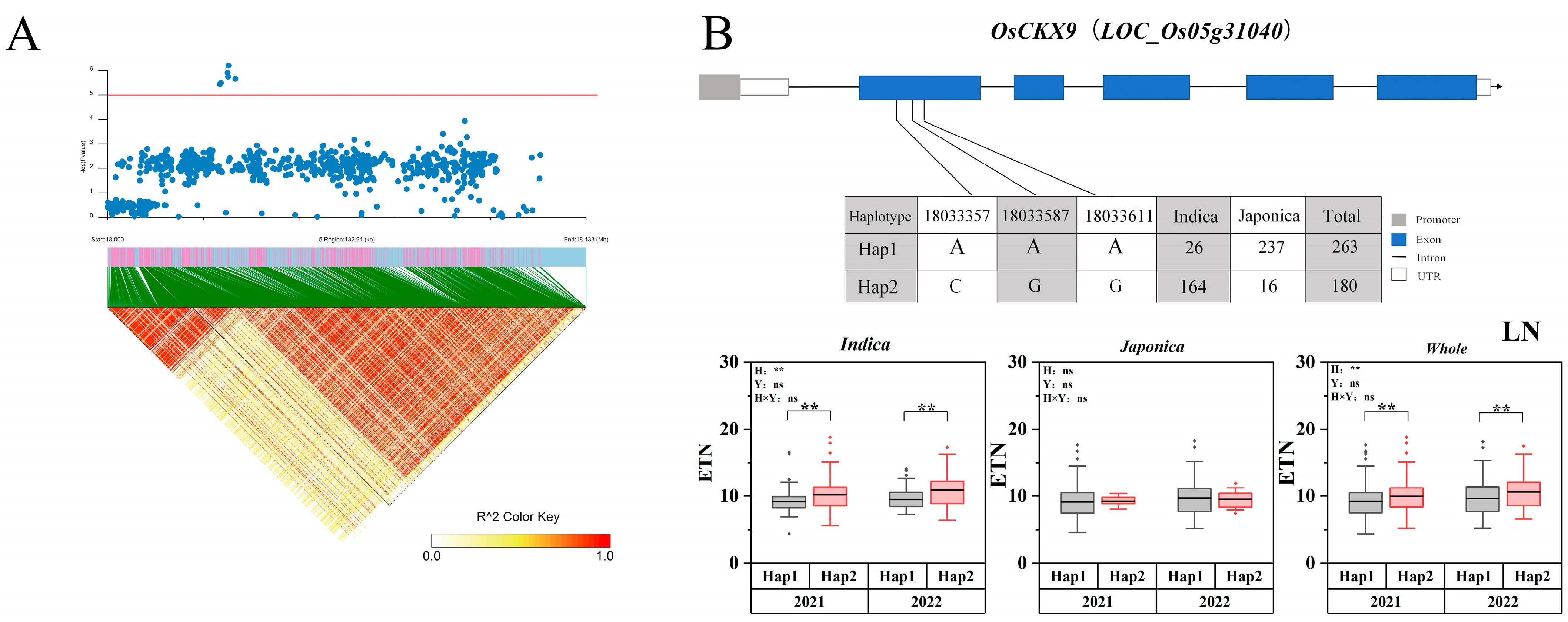
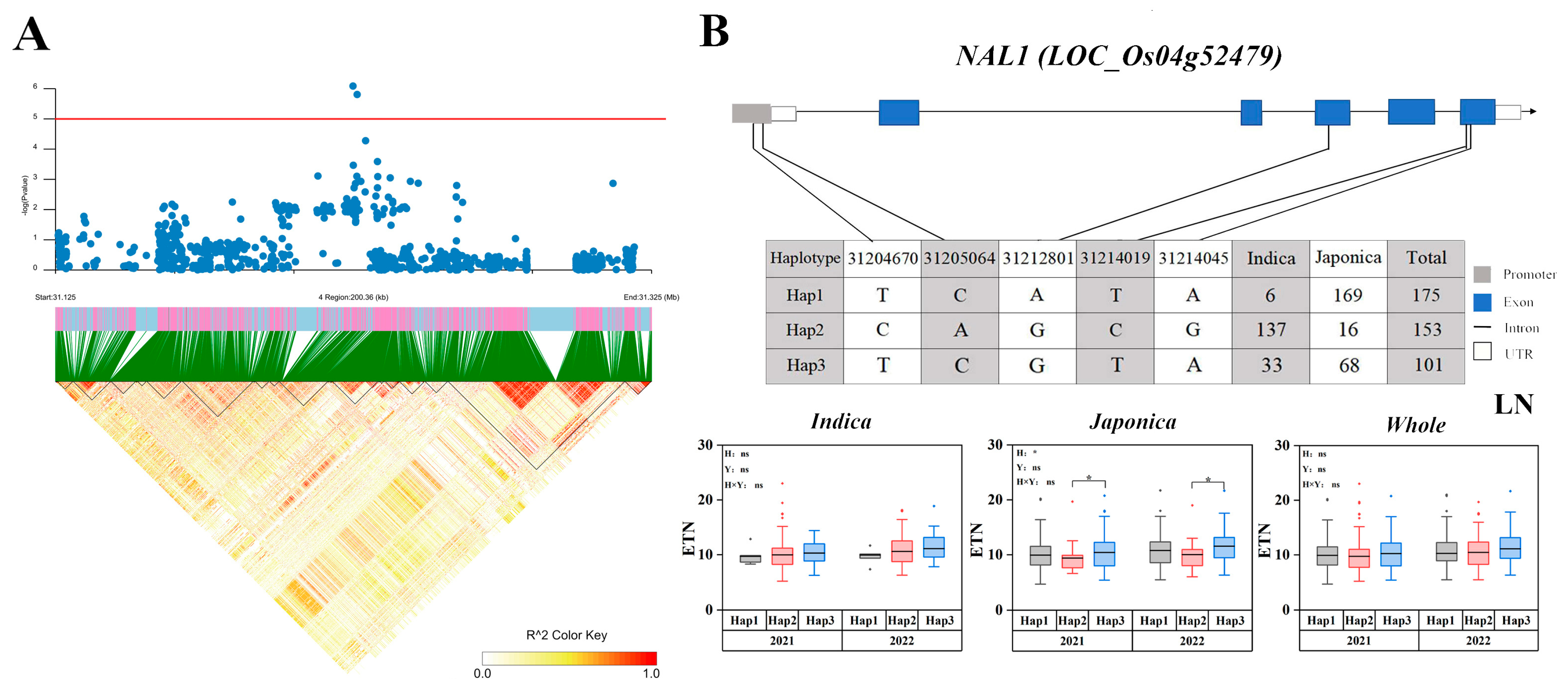
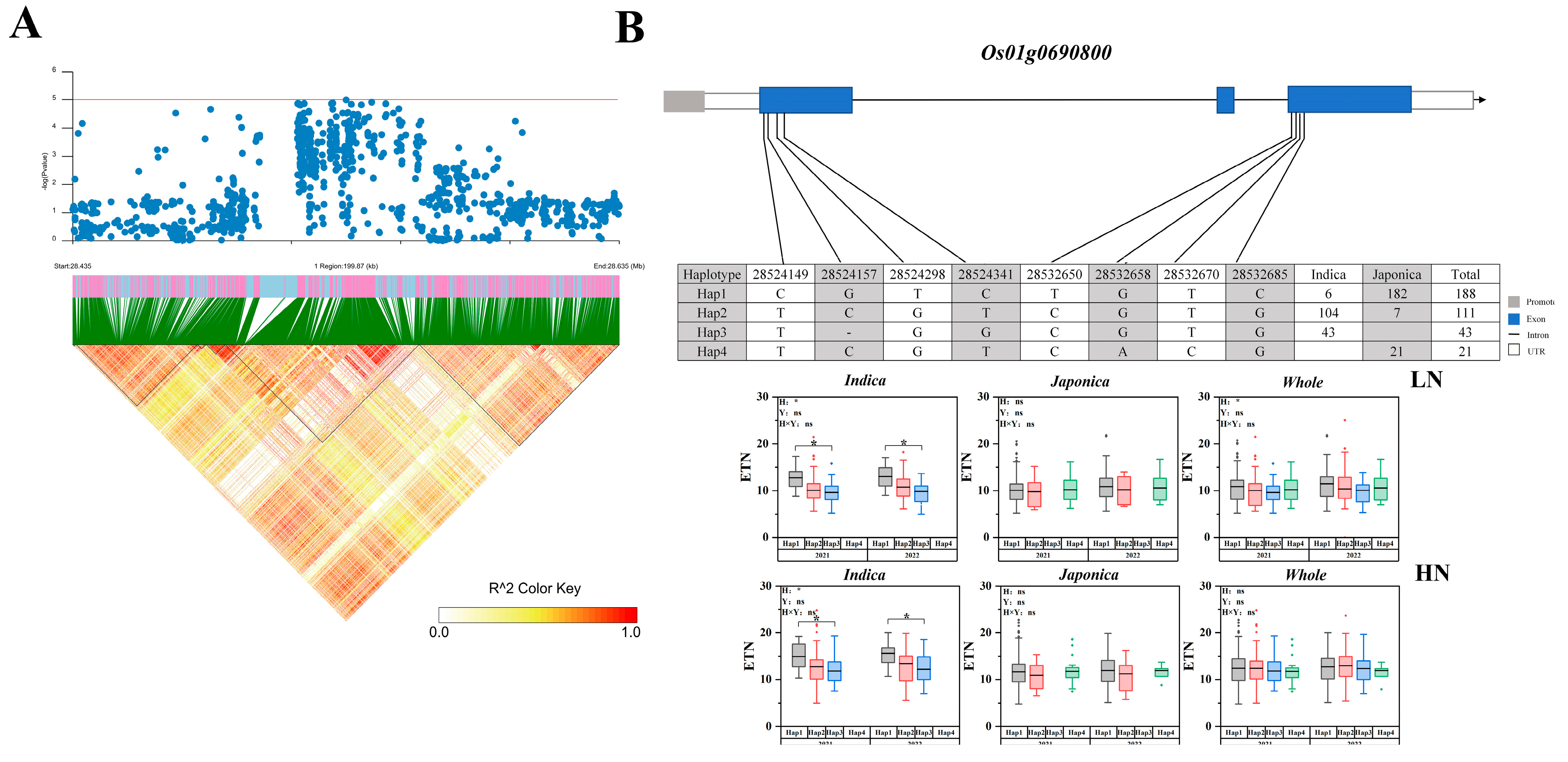
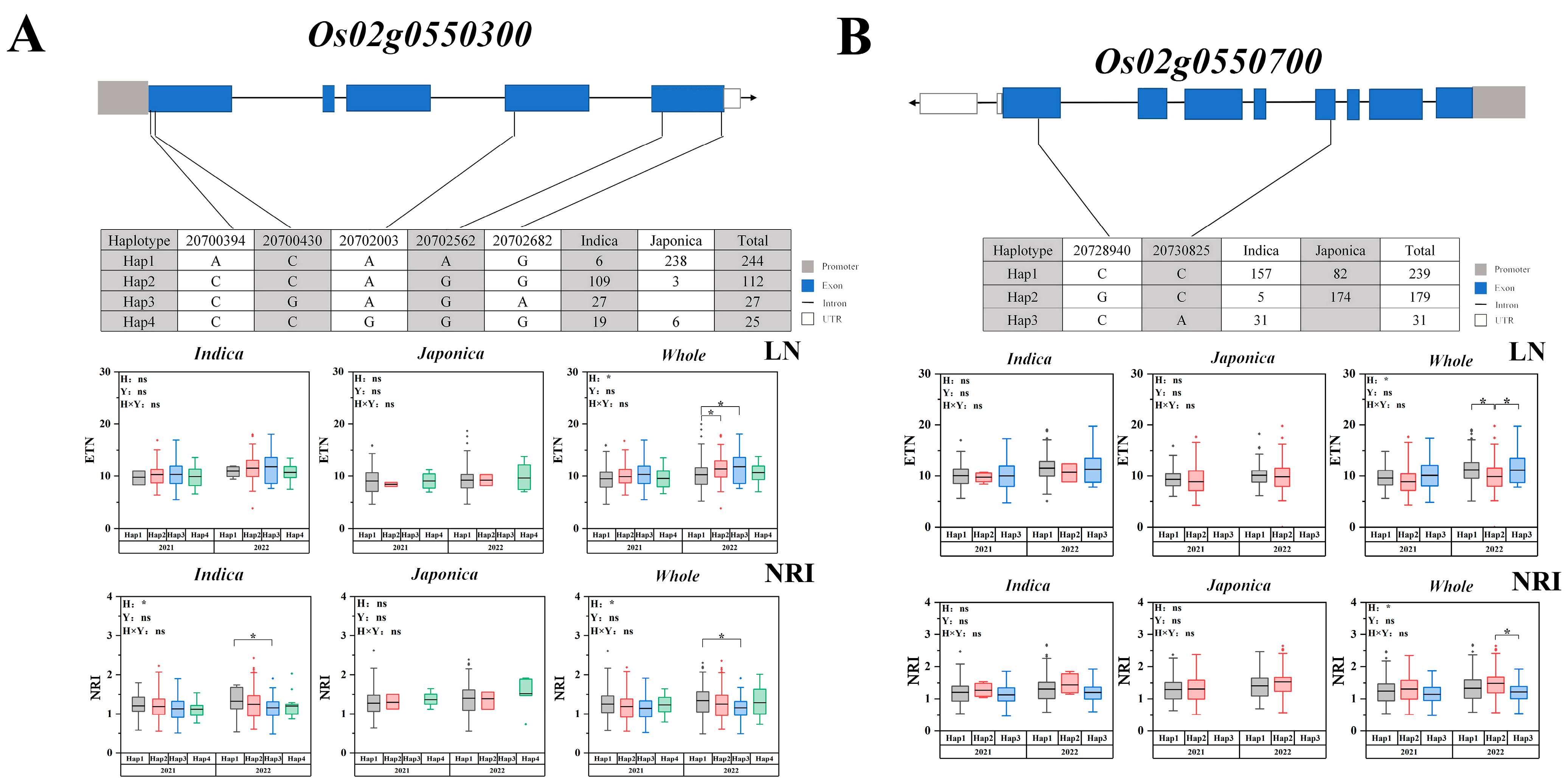
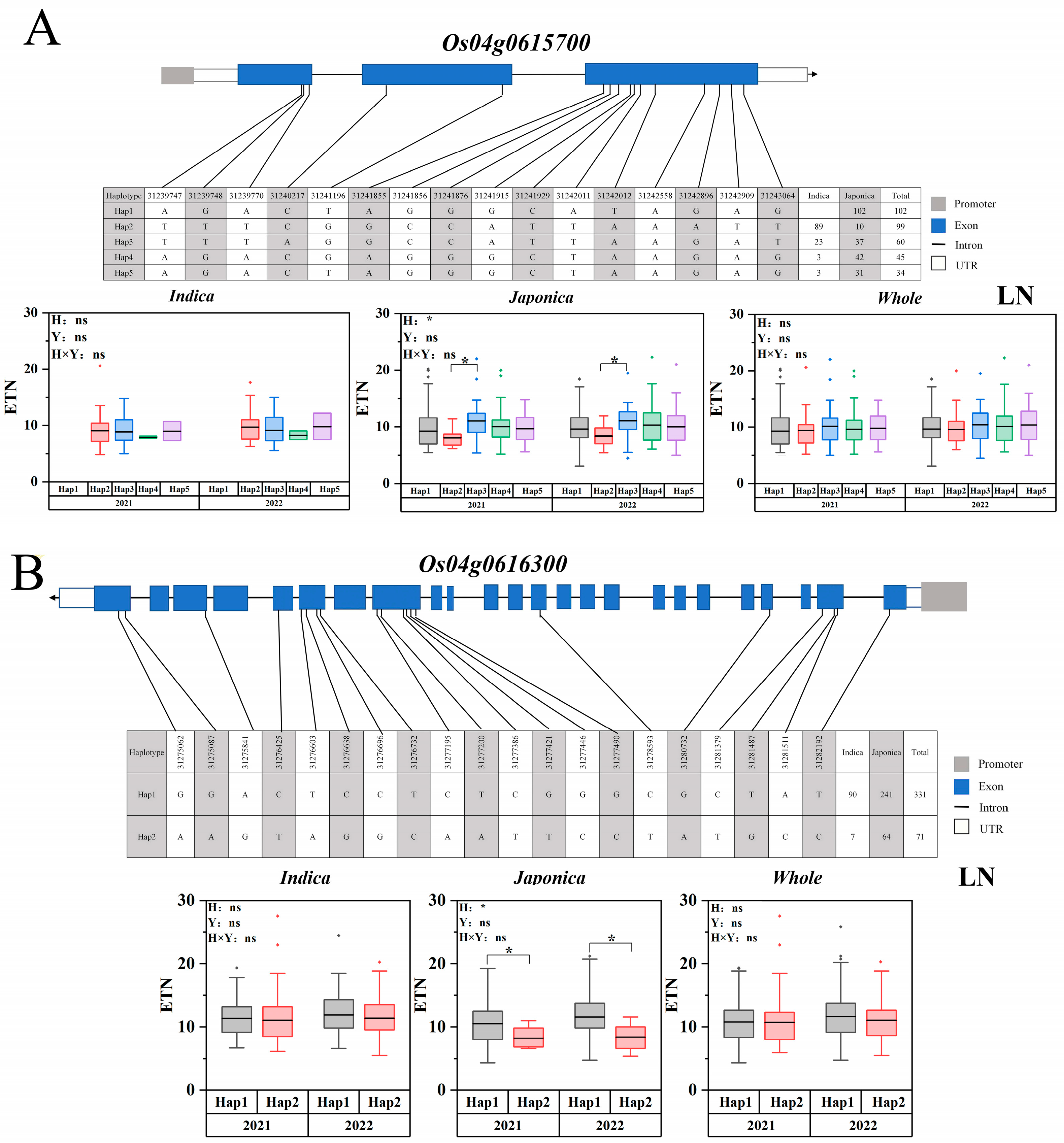
2.5. Candidate Gene Haplotype Analysis
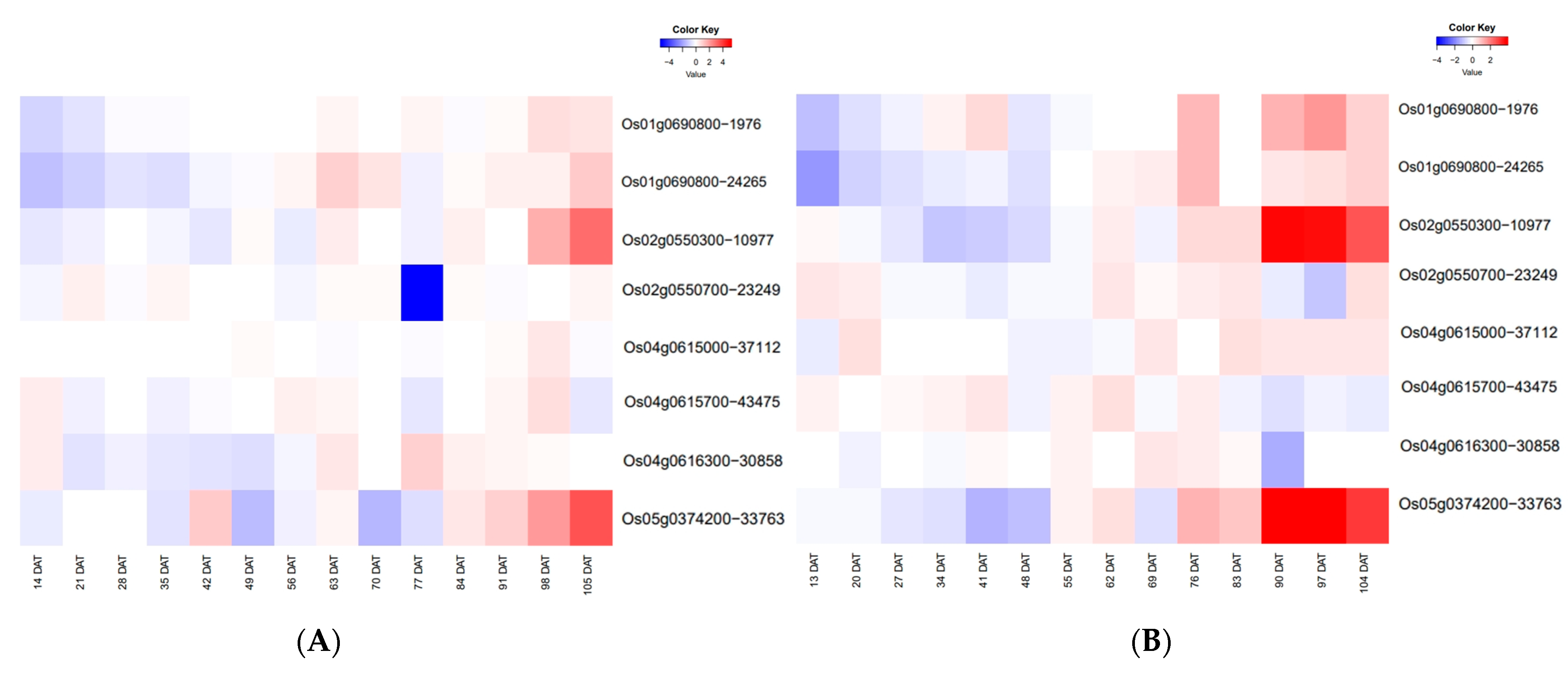
2.6. Temporal Expression Patterns of ETN-Related Genes
3. Discussion
3.1. Phenotypic Variation in ETN of Rice
3.2. Comparisons of QTL Detected in This Study with Previously Reported Genes
3.3. Candidate Gene Identification for Important QTLs
4. Materials and Methods
4.1. Plant Materials
4.2. Phenotype Determination
4.3. Phenotypic Data Analysis
4.4. Genotyping Data Analysis
4.5. Genome-Wide Association Analysis
4.6. Candidate Gene Haplotype Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cimini, A. Evolution of the Global Scientific Research on the Environmental Impact of Food Production from 1970 to 2020. Sustainability 2021, 13, 11633. [Google Scholar] [CrossRef]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate Trends and Global Crop Production Since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef]
- Huang, L.; Li, X.-X.; Zhang, Y.-B.; Fahad, S.; Wang, F. dep1 improves rice grain yield and nitrogen use efficiency simultaneously by enhancing nitrogen and dry matter translocation. J. Integr. Agric. 2022, 21, 2021. [Google Scholar] [CrossRef]
- Feng, H.; Li, B.; Zhi, Y.; Chen, J.; Li, R.; Xia, X.; Xu, G.; Fan, X. Overexpression of the nitrate transporter, OsNRT2.3b, improves rice phosphorus uptake and translocation. Plant Cell Rep. 2017, 36, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Zhang, Z.; Chai, X.; Wang, H.; Wang, Y.; et al. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wu, K.; Xu, R.; Yu, J.; Wang, J.; Zhao, Y.; Wang, Y.; Song, W.; Wang, S.; Gao, Z.; et al. Pyramiding of the dep1-1 and NAL1(NJ6) alleles achieves sustainable improvements in nitrogen-use efficiency and grain yield in japonica rice breeding. J. Genet. Genom. = Yi Chuan Xue Bao 2019, 46, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Wang, S.; Song, W.; Zhang, J.; Wang, Y.; Liu, Q.; Yu, J.; Ye, Y.; Li, S.; Chen, J.; et al. Enhanced sustainable green revolution yield via nitrogen-responsive chromatin modulation in rice. Science 2020, 367, eaaz2046. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Y.; Li, K.; Yan, M.; Zhang, J.; Yu, M.; Tang, S.; Wang, L.; Qu, H.; Luo, L.; et al. Nitrogen Mediates Flowering Time and Nitrogen Use Efficiency via Floral Regulators in Rice. Curr. Biol. CB 2021, 31, 671–683.e5. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, D.; Tang, M.; Li, D.; Zhu, Y.; Zhu, L.; Chen, C. THIS1 is a putative lipase that regulates tillering, plant height, and spikelet fertility in rice. J. Exp. Bot. 2013, 64, 4389–4402. [Google Scholar] [CrossRef]
- Lu, Z.; Shao, G.; Xiong, J.; Jiao, Y.; Wang, J.; Liu, G.; Meng, X.; Liang, Y.; Xiong, G.; Wang, Y.; et al. MONOCULM 3, an ortholog of WUSCHEL in rice, is required for tiller bud formation. J. Genet. Genom. = Yi Chuan Xue Bao 2015, 42, 71–78. [Google Scholar] [CrossRef]
- Luo, L.; Li, W.; Miura, K.; Ashikari, M.; Kyozuka, J. Control of tiller growth of rice by OsSPL14 and Strigolactones, which work in two independent pathways. Plant Cell Physiol. 2012, 53, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shang, L.; Yu, H.; Zeng, L.; Hu, J.; Ni, S.; Rao, Y.; Li, S.; Chu, J.; Meng, X.; et al. A Strigolactone Biosynthesis Gene Contributed to the Green Revolution in Rice. Mol. Plant 2020, 13, 923–932. [Google Scholar] [CrossRef]
- Arite, T.; Iwata, H.; Ohshima, K.; Maekawa, M.; Nakajima, M.; Kojima, M.; Sakakibara, H.; Kyozuka, J. DWARF10, an RMS1/MAX4/DAD1 ortholog, controls lateral bud outgrowth in rice. Plant J. Cell Mol. Biol. 2007, 51, 1019–1029. [Google Scholar] [CrossRef]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef]
- Liu, W.; Wu, C.; Fu, Y.; Hu, G.; Si, H.; Zhu, L.; Luan, W.; He, Z.; Sun, Z. Identification and characterization of HTD2: A novel gene negatively regulating tiller bud outgrowth in rice. Planta 2009, 230, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Guo, H.; Xiong, H.; Xie, Y.; Gu, J.; Zhao, L.; Zhao, S.; Ding, Y.; Liu, L. Strigolactone and abscisic acid synthesis and signaling pathways are enhanced in the wheat oligo-tillering mutant ot1. Mol. Breed. New Strateg. Plant Improv. 2024, 44, 12. [Google Scholar] [CrossRef]
- Yu, J.; Xuan, W.; Tian, Y.; Fan, L.; Sun, J.; Tang, W.; Chen, G.; Wang, B.; Liu, Y.; Wu, W.; et al. Enhanced OsNLP4-OsNiR cascade confers nitrogen use efficiency by promoting tiller number in rice. Plant Biotechnol. J. 2021, 19, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xia, K.; Liang, Z.; Chen, K.; Gao, C.; Zhang, M. MicroRNA393 is involved in nitrogen-promoted rice tillering through regulation of auxin signal transduction in axillary buds. Sci. Rep. 2016, 6, 32158. [Google Scholar] [CrossRef]
- Lv, Y.; Ma, J.; Wang, Y.; Wang, Q.; Lu, X.; Hu, H.; Qian, Q.; Guo, L.; Shang, L. Loci and Natural Alleles for Low-Nitrogen-Induced Growth Response Revealed by the Genome-Wide Association Study Analysis in Rice (Oryza sativa L.). Front. Plant Sci. 2021, 12, 770736. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, Y.; Chen, G.; Zhang, A.; Yang, S.; Shang, L.; Wang, D.; Ruan, B.; Liu, C.; Jiang, H.; et al. The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat. Commun. 2019, 10, 5207. [Google Scholar] [CrossRef]
- McCouch, S.R.; Wright, M.H.; Tung, C.W.; Maron, L.G.; McNally, K.L.; Fitzgerald, M.; Singh, N.; DeClerck, G.; Agosto-Perez, F.; Korniliev, P.; et al. Open access resources for genome-wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar] [CrossRef]
- Olsen, K.M.; Halldorsdottir, S.S.; Stinchcombe, J.R.; Weinig, C.; Schmitt, J.; Purugganan, M.D. Linkage disequilibrium mapping of Arabidopsis CRY2 flowering time alleles. Genetics 2004, 167, 1361–1369. [Google Scholar] [CrossRef]
- Sadhukhan, A.; Agrahari, R.K.; Wu, L.; Watanabe, T.; Nakano, Y.; Panda, S.K.; Koyama, H.; Kobayashi, Y. Expression genome-wide association study identifies that phosphatidylinositol-derived signalling regulates ALUMINIUM SENSITIVE3 expression under aluminium stress in the shoots of Arabidopsis thaliana. Plant Sci. Int. J. Exp. Plant Biol. 2021, 302, 110711. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Liu, S.; Ferjani, A.; Li, J.; Yan, J.; Yang, X.; Qin, F. Genetic variation in ZmVPP1 contributes to drought tolerance in maize seedlings. Nat. Genet. 2016, 48, 1233–1241. [Google Scholar] [CrossRef]
- Hinze, L.L.; Hulse-Kemp, A.M.; Wilson, I.W.; Zhu, Q.H.; Llewellyn, D.J.; Taylor, J.M.; Spriggs, A.; Fang, D.D.; Ulloa, M.; Burke, J.J.; et al. Diversity analysis of cotton (Gossypium hirsutum L.) germplasm using the CottonSNP63K Array. BMC Plant Biol. 2017, 17, 37. [Google Scholar] [CrossRef]
- Ma, X.; Feng, F.; Zhang, Y.; Elesawi, I.E.; Xu, K.; Li, T.; Mei, H.; Liu, H.; Gao, N.; Chen, C.; et al. A novel rice grain size gene OsSNB was identified by genome-wide association study in natural population. PLoS Genet. 2019, 15, e1008191. [Google Scholar] [CrossRef]
- Yuan, H.; Qin, P.; Hu, L.; Zhan, S.; Wang, S.; Gao, P.; Li, J.; Jin, M.; Xu, Z.; Gao, Q.; et al. OsSPL18 controls grain weight and grain number in rice. J. Genet. Genom. = Yi Chuan Xue Bao 2019, 46, 41–51. [Google Scholar] [CrossRef]
- Peng, Y.; Gao, Z.; Zhang, B.; Liu, C.; Xu, J.; Ruan, B.; Hu, J.; Dong, G.; Guo, L.; Liang, G.; et al. Fine mapping and candidate gene analysis of a major QTL for panicle structure in rice. Plant Cell Rep. 2014, 33, 1843–1850. [Google Scholar] [CrossRef]
- Wang, Q.; Su, Q.; Nian, J.; Zhang, J.; Guo, M.; Dong, G.; Hu, J.; Wang, R.; Wei, C.; Li, G.; et al. The Ghd7 transcription factor represses ARE1 expression to enhance nitrogen utilization and grain yield in rice. Mol. Plant 2021, 14, 1012–1023. [Google Scholar] [CrossRef]
- Fujita, D.; Trijatmiko, K.R.; Tagle, A.G.; Sapasap, M.V.; Koide, Y.; Sasaki, K.; Tsakirpaloglou, N.; Gannaban, R.B.; Nishimura, T.; Yanagihara, S.; et al. NAL1 allele from a rice landrace greatly increases yield in modern indica cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 20431–20436. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, S.; Liang, Y.; Zhang, R.; Liu, L.; Qin, P.; Zhang, Z.; Wang, Y.; Zhou, J.; Tang, X.; et al. Loss-function mutants of OsCKX gene family based on CRISPR-Cas systems revealed their diversified roles in rice. Plant Genome 2023, 16, e20283. [Google Scholar] [CrossRef]
- Duan, J.; Yu, H.; Yuan, K.; Liao, Z.; Meng, X.; Jing, Y.; Liu, G.; Chu, J.; Li, J. Strigolactone promotes cytokinin degradation through transcriptional activation of CYTOKININ OXIDASE/DEHYDROGENASE 9 in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 14319–14324. [Google Scholar] [CrossRef]
- Takai, T.; Adachi, S.; Taguchi-Shiobara, F.; Sanoh-Arai, Y.; Iwasawa, N.; Yoshinaga, S.; Hirose, S.; Taniguchi, Y.; Yamanouchi, U.; Wu, J.; et al. A natural variant of NAL1, selected in high-yield rice breeding programs, pleiotropically increases photosynthesis rate. Sci. Rep. 2013, 3, 2149. [Google Scholar] [CrossRef]
- Ma, X.; Li, F.; Zhang, Q.; Wang, X.; Guo, H.; Xie, J.; Zhu, X.; Ullah Khan, N.; Zhang, Z.; Li, J.; et al. Genetic architecture to cause dynamic change in tiller and panicle numbers revealed by genome-wide association study and transcriptome profile in rice. Plant J. Cell Mol. Biol. 2020, 104, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ren, M.; Peng, P.; Chun, Y.; Li, L.; Zhao, J.; Fang, J.; Peng, L.; Yan, J.; Chu, J.; et al. MIT1, encoding a 15-cis-ζ-carotene isomerase, regulates tiller number and stature in rice. J. Genet. Genom. = Yi Chuan Xue Bao 2021, 48, 88–91. [Google Scholar] [CrossRef]
- Ren, M.; Huang, M.; Qiu, H.; Chun, Y.; Li, L.; Kumar, A.; Fang, J.; Zhao, J.; He, H.; Li, X. Genome-Wide Association Study of the Genetic Basis of Effective Tiller Number in Rice. Rice 2021, 14, 56. [Google Scholar] [CrossRef]
- Huang, L.Y.; Sun, F.; Yuan, S.; Peng, S.B.; Wang, F. Different mechanisms underlying the yield advantage of ordinary hybrid and super hybrid rice over inbred rice under low and moderate N input conditions. Field Crops Res. 2018, 216, 150–157. [Google Scholar] [CrossRef]
- Sun, T.; Yang, X.; Tan, X.L.; Han, K.F.; Tang, S.; Tong, W.M.; Zhu, S.Y.; Hu, Z.P.; Wu, L.H. Comparison of Agronomic Performance between Japonica/Indica Hybrid and Japonica Cultivars of Rice Based on Different Nitrogen Rates. Agronomy 2020, 10, 171. [Google Scholar] [CrossRef]
- Deng, X.; Wang, J.; Liu, X.; Yang, J.; Zhou, M.; Kong, W.; Jiang, Y.; Ke, S.; Sun, T.; Li, Y. QTL Analysis and Heterosis Loci of Effective Tiller Using Three Genetic Populations Derived from Indica-Japonica Crosses in Rice. Agronomy 2022, 12, 2171. [Google Scholar] [CrossRef]
- Zhao, D.-D.; Park, J.-R.; Jang, Y.-H.; Kim, E.-G.; Du, X.-X.; Farooq, M.; Yun, B.-J.; Kim, K.-M. Identification of One Major QTL and a Novel Gene OsIAA17q5 Associated with Tiller Number in Rice Using QTL Analysis. Plants 2022, 11, 538. [Google Scholar] [CrossRef]
- Lanceras, J.C.; Pantuwan, G.; Jongdee, B.; Toojinda, T. Quantitative Trait Loci Associated with Drought Tolerance at Reproductive Stage in Rice. Plant Physiol. 2004, 135, 384–399. [Google Scholar] [CrossRef]
- Lin, L.; Zhao, Y.; Liu, F.; Chen, Q.; Qi, J. Narrow leaf 1 (NAL1) regulates leaf shape by affecting cell expansion in rice (Oryza sativa L.). Biochem. Biophys. Res. Commun. 2019, 516, 957–962. [Google Scholar] [CrossRef]
- Qi, J.; Qian, Q.; Bu, Q.; Li, S.; Chen, Q.; Sun, J.; Liang, W.; Zhou, Y.; Chu, C.; Li, X.; et al. Mutation of the Rice Narrow leaf1 Gene, Which Encodes a Novel Protein, Affects Vein Patterning and Polar Auxin Transport. Plant Physiol. 2008, 147, 1947–1959. [Google Scholar] [CrossRef]
- Zhai, L.; Zheng, T.; Wang, X.; Wang, Y.; Chen, K.; Wang, S.; Wang, Y.; Xu, J.; Li, Z. QTL mapping and candidate gene analysis of peduncle vascular bundle related traits in rice by genome-wide association study. Rice 2018, 11, 13. [Google Scholar] [CrossRef]
- Xu, J.L.; Wang, Y.; Zhang, F.; Wu, Y.; Zheng, T.Q.; Wang, Y.H.; Zhao, X.Q.; Cui, Y.R.; Chen, K.; Zhang, Q.; et al. SS1 (NAL1)- and SS2-Mediated Genetic Networks Underlying Source-Sink and Yield Traits in Rice (Oryza sativa L.). PLoS ONE 2015, 10, e0132060. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Li, W.W.; Liu, X.Y.; Chu, C.C. Molecular mechanism of tillering response to nitrogen in rice. Yi Chuan = Hereditas 2023, 45, 367–378. [Google Scholar]
- Jiang, L.; Liu, X.; Xiong, G.; Liu, H.; Chen, F.; Wang, L.; Meng, X.; Liu, G.; Yu, H.; Yuan, Y.; et al. DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature 2013, 504, 401–405. [Google Scholar] [CrossRef]
- Fang, Z.; Wu, B.; Ji, Y. The Amino Acid Transporter OsAAP4 Contributes to Rice Tillering and Grain Yield by Regulating Neutral Amino Acid Allocation through Two Splicing Variants. Rice 2021, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Bai, G.; Huang, W.; Wang, Z.; Wang, X.; Zhang, M. The Rice Peptide Transporter OsNPF7.3 Is Induced by Organic Nitrogen, and Contributes to Nitrogen Allocation and Grain Yield. Front. Plant Sci. 2017, 8, 1338. [Google Scholar] [CrossRef]
- van der Knaap, E.; Song, W.Y.; Ruan, D.L.; Sauter, M.; Ronald, P.C.; Kende, H. Expression of a gibberellin-induced leucine-rich repeat receptor-like protein kinase in deepwater rice and its interaction with kinase-associated protein phosphatase. Plant Physiol. 1999, 120, 559–570. [Google Scholar] [CrossRef]
- Tabangin, M.E.; Woo, J.G.; Martin, L.J. The effect of minor allele frequency on the likelihood of obtaining false positives. BMC Proc. 2009, 3 (Suppl. S7), S41. [Google Scholar] [CrossRef]
- Kang, H.M.; Sul, J.H.; Service, S.K.; Zaitlen, N.A.; Kong, S.-y.; Freimer, N.B.; Sabatti, C.; Eskin, E. Variance component model to account for sample structure in genome-wide association studies. Nat. Genet. 2010, 42, 348–354. [Google Scholar] [CrossRef]
- Lu, K.; Wei, L.; Li, X.; Wang, Y.; Wu, J.; Liu, M.; Zhang, C.; Chen, Z.; Xiao, Z.; Jian, H.; et al. Whole-genome resequencing reveals Brassica napus origin and genetic loci involved in its improvement. Nat. Commun. 2019, 10, 1154. [Google Scholar] [CrossRef]
- Slatkin, M. Linkage disequilibrium—Understanding the evolutionary past and mapping the medical future. Nat. Rev. Genet. 2008, 9, 477–485. [Google Scholar] [CrossRef]
- Kim, M.S.; Ko, S.R.; Le, V.T.; Jee, M.G.; Jung, Y.J.; Kang, K.K.; Cho, Y.G. Development of SNP Markers from GWAS for Selecting Seed Coat and Aleurone Layers in Brown Rice (Oryza sativa L.). Genes 2022, 13, 1805. [Google Scholar] [CrossRef] [PubMed]
- Hellenthal, G. Population Structure, Demography and Recent Admixture. In Handbook of Statistical Genomics; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 247–274. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Nitrogen | Subpopulation | Average | SD | Skewness | Kurtosis | Min | Max | CV% |
|---|---|---|---|---|---|---|---|---|---|
| 2021 | LN | Indica | 10.60 | 3.33 | 1.41 | 1.11 | 5.20 | 22.00 | 31.44 |
| Japonica | 8.24 | 2.47 | 1.29 | 0.94 | 3.33 | 18.40 | 29.92 | ||
| Whole | 9.35 | 2.42 | 0.74 | 0.78 | 3.00 | 17.25 | 29.07 | ||
| HN | Indica | 12.05 | 3.76 | 2.37 | 1.31 | 5.80 | 26.80 | 31.24 | |
| Japonica | 9.96 | 3.00 | 0.77 | 0.77 | 4.25 | 20.00 | 30.12 | ||
| Whole | 11.14 | 3.60 | 2.39 | 1.20 | 4.25 | 26.80 | 32.30 | ||
| 2022 | LN | Indica | 11.21 | 3.36 | 4.59 | 1.58 | 5.17 | 27.50 | 29.99 |
| Japonica | 10.12 | 3.27 | 0.10 | 0.80 | 4.33 | 19.25 | 32.30 | ||
| Whole | 10.63 | 3.35 | 2.46 | 1.15 | 4.33 | 27.50 | 31.52 | ||
| HN | Indica | 13.48 | 3.91 | 3.09 | 1.14 | 6.20 | 33.00 | 29.01 | |
| Japonica | 11.96 | 4.20 | 1.08 | 1.02 | 4.83 | 26.67 | 35.15 | ||
| Whole | 12.70 | 4.13 | 1.72 | 0.98 | 4.83 | 33.00 | 32.53 |
| Chr | Start | End | Year | Subpopulation | QTL | p-Value | Candidate/Known Gene |
|---|---|---|---|---|---|---|---|
| 1 | 28312507 | 28635235 | 2021 | Indica | qLETN1-2 | 9.7639 × 10−6 | Os01g0690800 |
| 2021 | Indica | qHETN1-2 | 4.96655 × 10−6 | ||||
| 2022 | Indica, Whole | qHETN1-2 | 3.05122 × 10−6 | ||||
| 2 | 1925017 | 2258654 | 2021 | Indica | qLETN2-1 | 3.77201 × 10−6 | |
| Indica, Whole | qHETN2-1, qHETN2-3 | 8.0282 × 10−7 | |||||
| 2022 | Whole | qLETN2-1 | 7.92919 × 10−6 | ||||
| 2 | 20316453 | 20806338 | 2022 | Indica, Whole | qLETN2-4, qLETN2-5, qLETN2-6, qLETN2-7 | 8.01316 × 10−7 | Os02t0550300 Os02t0550700 |
| Indica, Whole | qNRI2-4, qNRI2-5, qNRI2-6 | 5.38504 × 10−7 | |||||
| 3 | 28202203 | 28408332 | 2021 | Indica | qLETN3-1 | 1.25578 × 10−6 | |
| 2022 | Indica | qLETN3-1 | 7.8412 × 10−6 | ||||
| 4 | 31125957 | 31325965 | 2021 | Japonica, Whole | qLETN4-3 | 4.1416 × 10−6 | NAL1 [30] Os04g0615700 Os04t0616300 |
| 2022 | Japonica, Whole | qLETN4-3 | 3.52586 × 10−6 | ||||
| 5 | 18033644 | 18260106 | 2021 | Indica | qLETN5-1 | 3.84225 × 10−6 | OsCKX9 [31] |
| 2022 | Indica | qLETN5-1 | 5.65649 × 10−7 | ||||
| 5 | 20672878 | 20956221 | 2021 | Whole | qNRI5-3 | 8.01541 × 10−6 | |
| 2022 | Whole | qNRI5-3 | 8.29562 × 10−6 | ||||
| 7 | 9167844 | 9368439 | 2021 | Japonica | qLETN7-1 | 4.6369 × 10−6 | |
| 2022 | Japonica | qLETN7-1 | 7.8421 × 10−6 | ||||
| 7 | 27836562 | 28048531 | 2021 | Indica | qNRI7-2 | 6.56564 × 10−6 | |
| 2022 | Indica | qNRI7-2 | 5.6232 × 10−6 | ||||
| 8 | 20505376 | 20723565 | 2021 | Whole | qLETN8-1 | 8.65652 × 10−6 | |
| 2022 | Whole | qLETN8-1 | 7.51026 × 10−6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Xin, W.; Chen, L.; Liu, Y.; Wang, X.; Ma, C.; Zhai, L.; Feng, Y.; Gao, J.; Zhang, W. Genome-Wide Association Analysis of Effective Tillers in Rice under Different Nitrogen Gradients. Int. J. Mol. Sci. 2024, 25, 2969. https://doi.org/10.3390/ijms25052969
Liu Y, Xin W, Chen L, Liu Y, Wang X, Ma C, Zhai L, Feng Y, Gao J, Zhang W. Genome-Wide Association Analysis of Effective Tillers in Rice under Different Nitrogen Gradients. International Journal of Molecular Sciences. 2024; 25(5):2969. https://doi.org/10.3390/ijms25052969
Chicago/Turabian StyleLiu, Yuzhuo, Wei Xin, Liqiang Chen, Yuqi Liu, Xue Wang, Cheng Ma, Laiyuan Zhai, Yingying Feng, Jiping Gao, and Wenzhong Zhang. 2024. "Genome-Wide Association Analysis of Effective Tillers in Rice under Different Nitrogen Gradients" International Journal of Molecular Sciences 25, no. 5: 2969. https://doi.org/10.3390/ijms25052969