
Pulmonary Adenocarcinoma In Situ and Minimally Invasive Adenocarcinomas in European Patients Have Less KRAS and More EGFR Mutations Compared to Advanced Adenocarcinomas

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
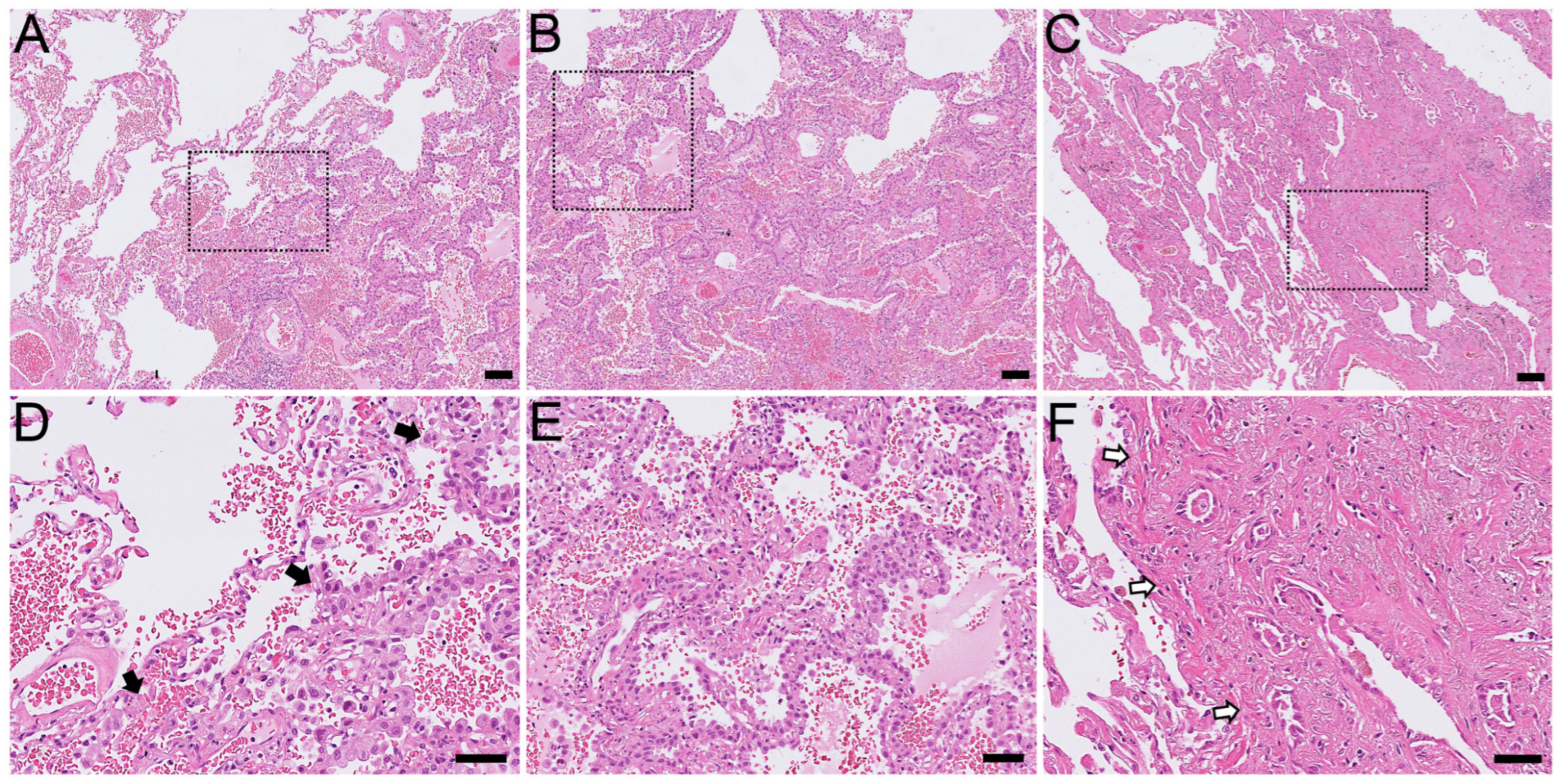
2.1. Histological Findings
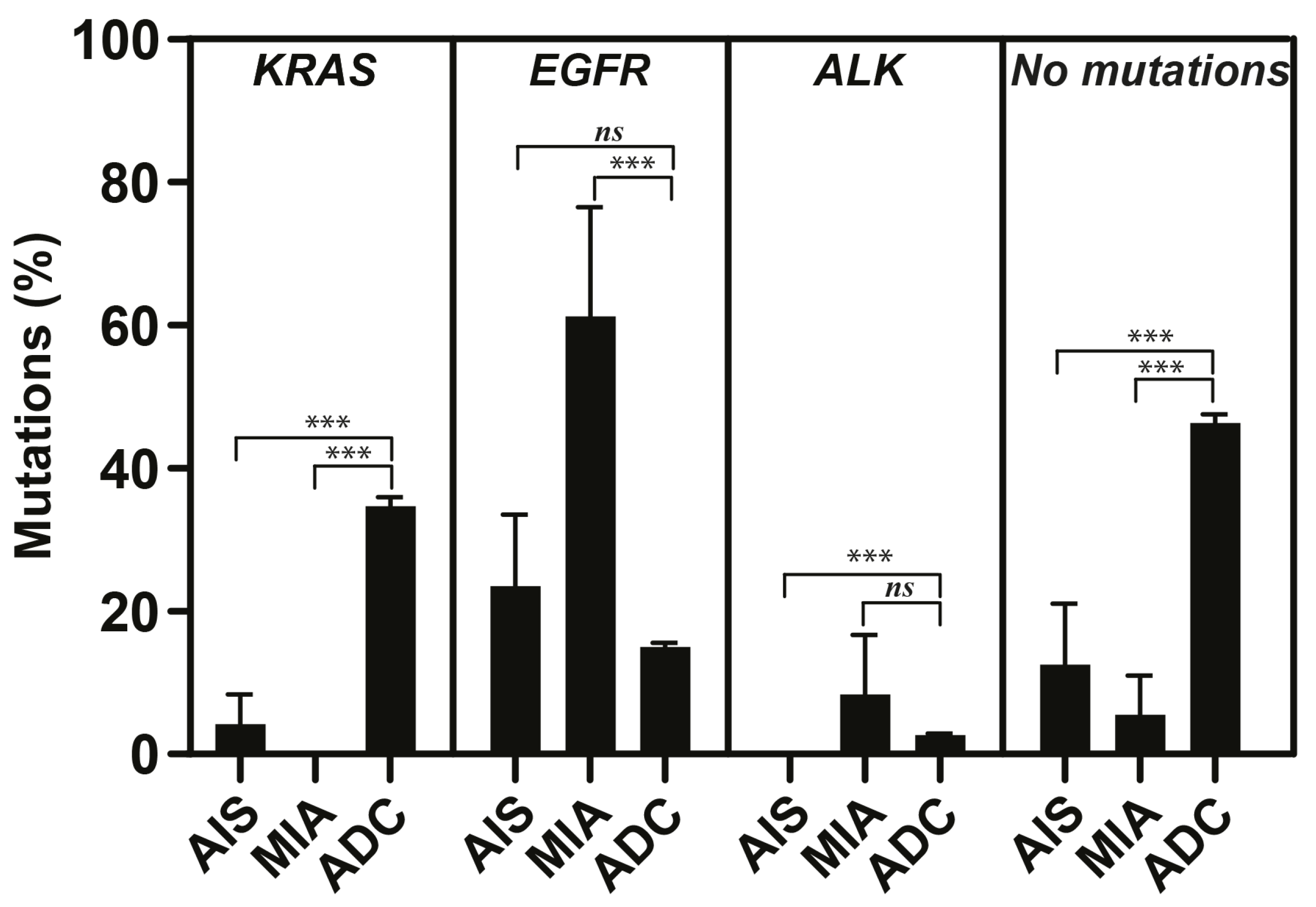
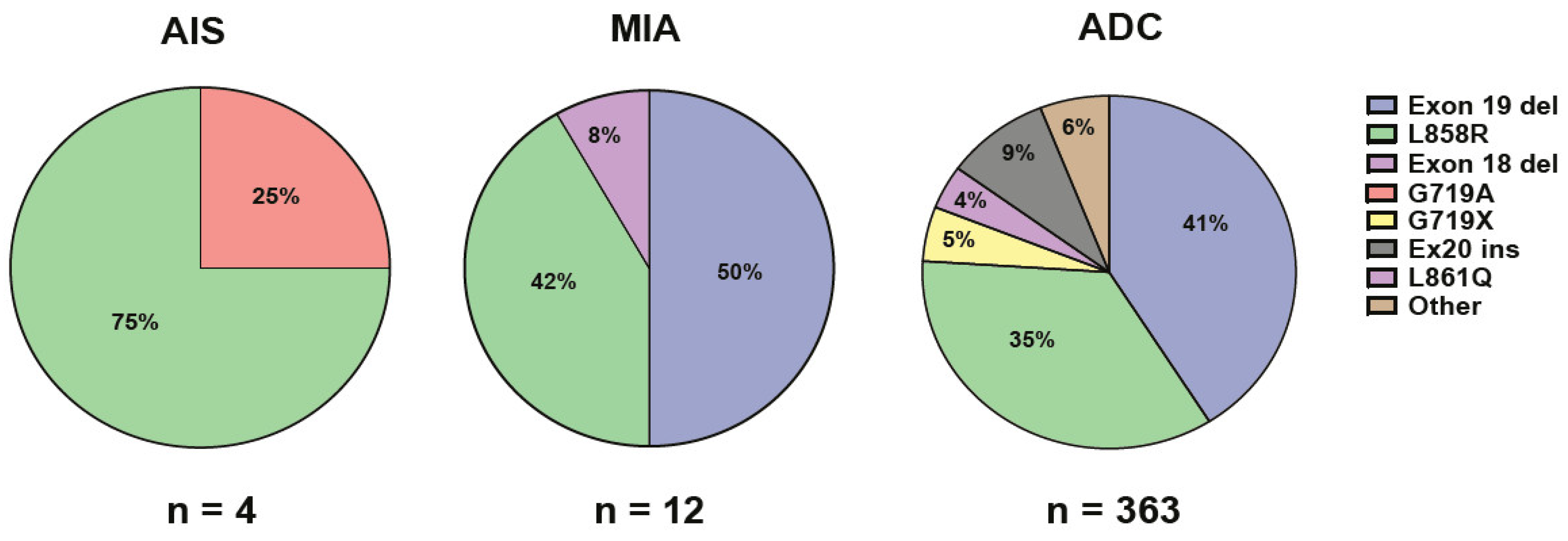
2.2. Genetic Findings
3. Discussion
4. Material and Methods
4.1. Patient Material
4.2. Histomorphological Review
4.3. Next-Generation Sequencing
4.4. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gazdar, A.F.; Bunn, P.A.; Minna, J.D. Small-cell lung cancer: What we know, what we need to know and the path forward. Nat. Rev. Cancer 2017, 17, 765. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, K.D.; Berns, A. Cell of origin of lung cancer. Mol. Oncol. 2010, 4, 397–403. [Google Scholar] [CrossRef]
- Jackson, E.L.; Willis, N.; Mercer, K.; Bronson, R.T.; Crowley, D.; Montoya, R.; Jacks, T.; Tuveson, D.A. Analysis of lung tumor initiation and progression using conditional expression of oncogenic K-ras. Genes. Dev. 2001, 15, 3243–3248. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.F.; Jackson, E.L.; Woolfenden, A.E.; Lawrence, S.; Babar, I.; Vogel, S.; Crowley, D.; Bronson, R.T.; Jacks, T. Identification of bronchioalveolar stem cells in normal lung and lung cancer. Cell 2005, 121, 823–835. [Google Scholar] [CrossRef]
- WHO Classification of Tumors Editorial Board. Thoracic Tumors, 5th ed.; WHO Classification of Tumors Editorial Board: Lyon, France, 2021; Volume 5, pp. 57–84. [Google Scholar]
- Ricciuti, B.; Leonardi, G.C.; Metro, G.; Grignani, F.; Paglialunga, L.; Bellezza, G.; Baglivo, S.; Mencaroni, C.; Baldi, A.; Zicari, D.; et al. Targeting the KRAS variant for treatment of non-small cell lung cancer: Potential therapeutic applications. Expert. Rev. Respir. Med. 2016, 10, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.T.; Vyse, S.; Huang, P.H. Rare epidermal growth factor receptor (EGFR) mutations in non-small cell lung cancer. Semin. Cancer Biol. 2020, 61, 167–179. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Yuan, J.Q.; Wang, K.F.; Fu, X.H.; Han, X.R.; Threapleton, D.; Yang, Z.Y.; Mao, C.; Tang, J.L. The prevalence of EGFR mutation in patients with non-small cell lung cancer: A systematic review and meta-analysis. Oncotarget 2016, 7, 78985–78993. [Google Scholar] [CrossRef]
- Altorki, N.; Wang, X.; Kozono, D.; Watt, C.; Landrenau, R.; Wigle, D.; Port, J.; Jones, D.R.; Conti, M.; Ashrafi, A.S.; et al. Lobar or sublobar resection for peripheral stage IA non-small-cell lung cancer. N. Engl. J. Med. 2023, 388, 489–498. [Google Scholar] [CrossRef]
- Aramini, B.; Banchelli, F.; Bettelli, S.; Manfredini, S.; D’Amico, R.; Masciale, V.; Pinelli, M.; Moretti, M.; Stefani, A.; Bertolini, F.; et al. Overall survival in patients with lung adenocarcinoma harboring “niche” mutations: An observational study. Oncotarget 2020, 11, 550–559. [Google Scholar] [CrossRef]
- Inamura, K. Clinicopathological Characteristics and mutations driving development of early lung adenocarcinoma: Tumor initiation and progression. Int. J. Mol. Sci. 2018, 19, 1259. [Google Scholar] [CrossRef]
- Pino, M.S.; Chung, D.C. The chromosomal instability pathway in colon cancer. Gastroenterology 2010, 138, 2059–2072. [Google Scholar] [CrossRef]
- Smith, G.; Carey, F.A.; Beattie, J.; Wilkie, M.J.; Lightfoot, T.J.; Coxhead, J.; Garner, R.C.; Steele, R.J.; Wolf, C.R. Mutations in APC, Kirsten-ras, and p53—Alternative genetic pathways to colorectal cancer. Proc. Natl. Acad. Sci. USA 2002, 99, 9433–9438. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Petricoin, E.F.; Maitra, A.; Rajapakse, V.; King, C.; Jacobetz, M.A.; Ross, S.; Conrads, T.P.; Veenstra, T.D.; Hitt, B.A.; et al. Preinvasive and invasive ductal pancreatic cancer and its early detection in the mouse. Cancer Cell 2003, 4, 437–450. [Google Scholar] [CrossRef]
- Kent, D.G.; Green, A.R. Order Matters: The order of somatic mutations influences cancer evolution. Cold Spring Harb. Perspect. Med. 2017, 7, a027060. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Carrot-Zhang, J.; Zhao, Y.; Hu, H.; Freeman, S.S.; Yu, S.; Ha, G.; Taylor, A.M.; Berger, A.C.; Westlake, L.; et al. Genomic and immune profiling of pre-invasive lung adenocarcinoma. Nat. Commun. 2019, 10, 5472. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, A.; Sumiyoshi, S.; Sonobe, M.; Kobayashi, M.; Fujimoto, M.; Kawakami, F.; Tsuruyama, T.; Travis, W.D.; Date, H.; Haga, H. Validation of the IASLC/ATS/ERS lung adenocarcinoma classification for prognosis and association with EGFR and KRAS gene mutations: Analysis of 440 Japanese patients. J. Thorac. Oncol. 2013, 8, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Koizumi, H.; Kimura, H.; Marushima, H.; Saji, H.; Takagi, M. Epidermal growth factor receptor mutations in adenocarcinoma in situ and minimally invasive adenocarcinoma detected using mutation-specific monoclonal antibodies. Lung Cancer 2016, 99, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Beau-Faller, M.; Texier, M.; Blons, H.; Richard, N.; Escande, F.; Melaabi, S.; Lizard, S.; De Fraipont, F.; Longchampt, E.; Morin, F.; et al. Clinical relevance of EGFR- or KRAS-mutated subclones in patients with advanced non-small-cell lung cancer receiving Erlotinib in a French prospective cohort (IFCT ERMETIC2 Cohort—Part 2). Clin. Lung Cancer 2019, 20, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Jaromi, L.; Csongei, V.; Vesel, M.; Abdelwahab, E.M.M.; Soltani, A.; Torok, Z.; Smuk, G.; Sarosi, V.; Pongracz, J.E. KRAS and EGFR mutations differentially alter ABC drug transporter expression in Cisplatin-resistant non-small cell lung cancer. Int. J. Mol. Sci. 2021, 22, 5384. [Google Scholar] [CrossRef]
- Wang, X.; Ricciuti, B.; Nguyen, T.; Li, X.; Rabin, M.S.; Awad, M.M.; Lin, X.; Johnson, B.E.; Christiani, D.C. Association between smoking history and tumor mutation burden in advanced non-small cell lung cancer. Cancer Res. 2021, 81, 2566–2573. [Google Scholar] [CrossRef]
- Skoulidis, F.; Heymach, J.V. Co-occurring genomic alterations in non-small-cell lung cancer biology and therapy. Nat. Rev. Cancer 2019, 19, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, H.; Pan, Y.; Wang, R.; Li, Y.; Shen, L.; Yu, Y.; Li, H.; Cai, D.; Sun, Y.; et al. PIK3CA mutations frequently coexist with EGFR/KRAS mutations in non-small cell lung cancer and suggest poor prognosis in EGFR/KRAS wildtype subgroup. PLoS ONE 2014, 9, e88291. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.A.; Suzawa, K.; Jordan, E.; Zehir, A.; Ni, A.; Kim, R.; Kris, M.G.; Hellmann, M.D.; Li, B.T.; Somwar, R.; et al. Concurrent alterations in EGFR-mutant lung cancers associated with resistance to EGFR kinase inhibitors and characterization of MTOR as a mediator of resistance. Clin. Cancer Res. 2018, 24, 3108–3118. [Google Scholar] [CrossRef] [PubMed]
- Arbour, K.C.; Jordan, E.; Kim, H.R.; Dienstag, J.; Yu, H.A.; Sanchez-Vega, F.; Lito, P.; Berger, M.; Solit, D.B.; Hellmann, M.; et al. Effects of Co-occurring genomic alterations on outcomes in patients with KRAS-mutant non-small cell lung cancer. Clin. Cancer Res. 2018, 24, 334–340. [Google Scholar] [CrossRef]
- Benesova, L.; Minarik, M.; Jancarikova, D.; Belsanova, B.; Pesek, M. Multiplicity of EGFR and KRAS mutations in non-small cell lung cancer (NSCLC) patients treated with tyrosine kinase inhibitors. Anticancer Res. 2010, 30, 1667–1671. [Google Scholar]
- Qu, Y.; Che, N.; Zhao, D.; Zhang, C.; Su, D.; Zhou, L.; Zhang, L.; Wang, C.; Zhang, H.; Wei, L. The clinicopathological significance of ALK rearrangements and KRAS and EGFR mutations in primary pulmonary mucinous adenocarcinoma. Tumour Biol. 2015, 36, 6417–6424. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Lyu, Z.; Cai, J.; Zheng, N.; Li, Y.; Xu, T.; Zeng, H. The co-mutation of EGFR and tumor-related genes leads to a worse prognosis and a higher level of tumor mutational burden in Chinese non-small cell lung cancer patients. J. Thorac. Dis. 2022, 14, 185–193. [Google Scholar] [CrossRef]
- Tsukumo, Y.; Naito, M.; Suzuki, T. Influence of EGFR-activating mutations on sensitivity to tyrosine kinase inhibitors in a KRAS mutant non-small cell lung cancer cell line. PLoS ONE 2020, 15, e0229712. [Google Scholar] [CrossRef]
- Ito, M.; Miyata, Y.; Kushitani, K.; Ueda, D.; Takeshima, Y.; Okada, M. Distribution and prognostic impact of EGFR and KRAS mutations according to histological subtype and tumor invasion status in pTis-3N0M0 lung adenocarcinoma. BMC Cancer 2023, 23, 248. [Google Scholar] [CrossRef]
- Adib, E.; Nassar, A.H.; Abou Alaiwi, S.; Groha, S.; Akl, E.W.; Sholl, L.M.; Michael, K.S.; Awad, M.M.; Jӓnne, P.A.; Gusev, A.; et al. Variation in targetable genomic alterations in non-small cell lung cancer by genetic ancestry, sex, smoking history, and histology. Genome Med. 2022, 14, 39. [Google Scholar] [CrossRef]
- Thulin, A.; Andersson, C.; Werner Ronnerman, E.; De Lara, S.; Chamalidou, C.; Schoenfeld, A.; Kovács, A.; Fagman, H.; Enlund, F.; Linderholm, B.K. Discordance of PIK3CA and TP53 mutations between breast cancer brain metastases and matched primary tumors. Sci. Rep. 2021, 11, 23548. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| AIS + MIA (n = 28) | AIS (n = 10) | MIA (n = 18) | |
|---|---|---|---|
| Gender | |||
| Men | 17.0% (5) | 10.0% (1) | 22.0% (4) |
| Women | 83.0% (23) | 90.0% (9) | 78.0% (14) |
| Age (year) | |||
| Men | 68.0 ± 3.4 | 66.0 ± 0.0 | 69.0 ± 4.4 |
| Women | 69.0 ± 2.0 | 71.1 ± 3.1 | 67.7 ± 2.4 |
| Mutation | |||
| EGFR | 57.1% (16) | 40.0% (4) | 66.0% (12) |
| KRAS | 3.5% (1) | 10.0% (1) | 0.0% (0) |
| Tumor size | |||
| <10 mm | 32.0% (9) | 40.0% (4) | 28.0% (5) |
| 10–19 mm | 54.0% (15) | 40.0% (4) | 61.0% (11) |
| 20–30 mm | 14.0% (4) | 20.0% (2) | 11.0% (2) |
| Lobe | |||
| Left upper | 24.0% (7) | 20.0% (2) | 28.0% (5) |
| Left lower | 18.0% (5) | 30.0% (3) | 11.0% (2) |
| Right upper | 36.0% (10) | 30.0% (3) | 40.0% (7) |
| Right middle | 4.0% (1) | 0.0% (0) | 6.0% (1) |
| Right lower | 18.0% (5) | 20.0% (2) | 17.0% (3) |
| Smoking history | |||
| Active | 3.5% (1) | 0.0% (0) | 6.0% (1) |
| Prior | 43.0% (12) | 80.0% (8) | 22.0% (4) |
| Never | 53.5% (15) | 20.0% (2) | 72.0% (13) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petterson, J.; Mustafa, D.; Bandaru, S.; Eklund, E.Ä.; Hallqvist, A.; Sayin, V.I.; Gagné, A.; Fagman, H.; Akyürek, L.M. Pulmonary Adenocarcinoma In Situ and Minimally Invasive Adenocarcinomas in European Patients Have Less KRAS and More EGFR Mutations Compared to Advanced Adenocarcinomas. Int. J. Mol. Sci. 2024, 25, 2959. https://doi.org/10.3390/ijms25052959
Petterson J, Mustafa D, Bandaru S, Eklund EÄ, Hallqvist A, Sayin VI, Gagné A, Fagman H, Akyürek LM. Pulmonary Adenocarcinoma In Situ and Minimally Invasive Adenocarcinomas in European Patients Have Less KRAS and More EGFR Mutations Compared to Advanced Adenocarcinomas. International Journal of Molecular Sciences. 2024; 25(5):2959. https://doi.org/10.3390/ijms25052959
Chicago/Turabian StylePetterson, Jennie, Dyar Mustafa, Sashidar Bandaru, Ella Äng Eklund, Andreas Hallqvist, Volkan I. Sayin, Andréanne Gagné, Henrik Fagman, and Levent M. Akyürek. 2024. "Pulmonary Adenocarcinoma In Situ and Minimally Invasive Adenocarcinomas in European Patients Have Less KRAS and More EGFR Mutations Compared to Advanced Adenocarcinomas" International Journal of Molecular Sciences 25, no. 5: 2959. https://doi.org/10.3390/ijms25052959