
Oxidative Stress in Sepsis: A Focus on Cardiac Pathology
 , , , , and
, , , , and
Abstract
:1. Introduction
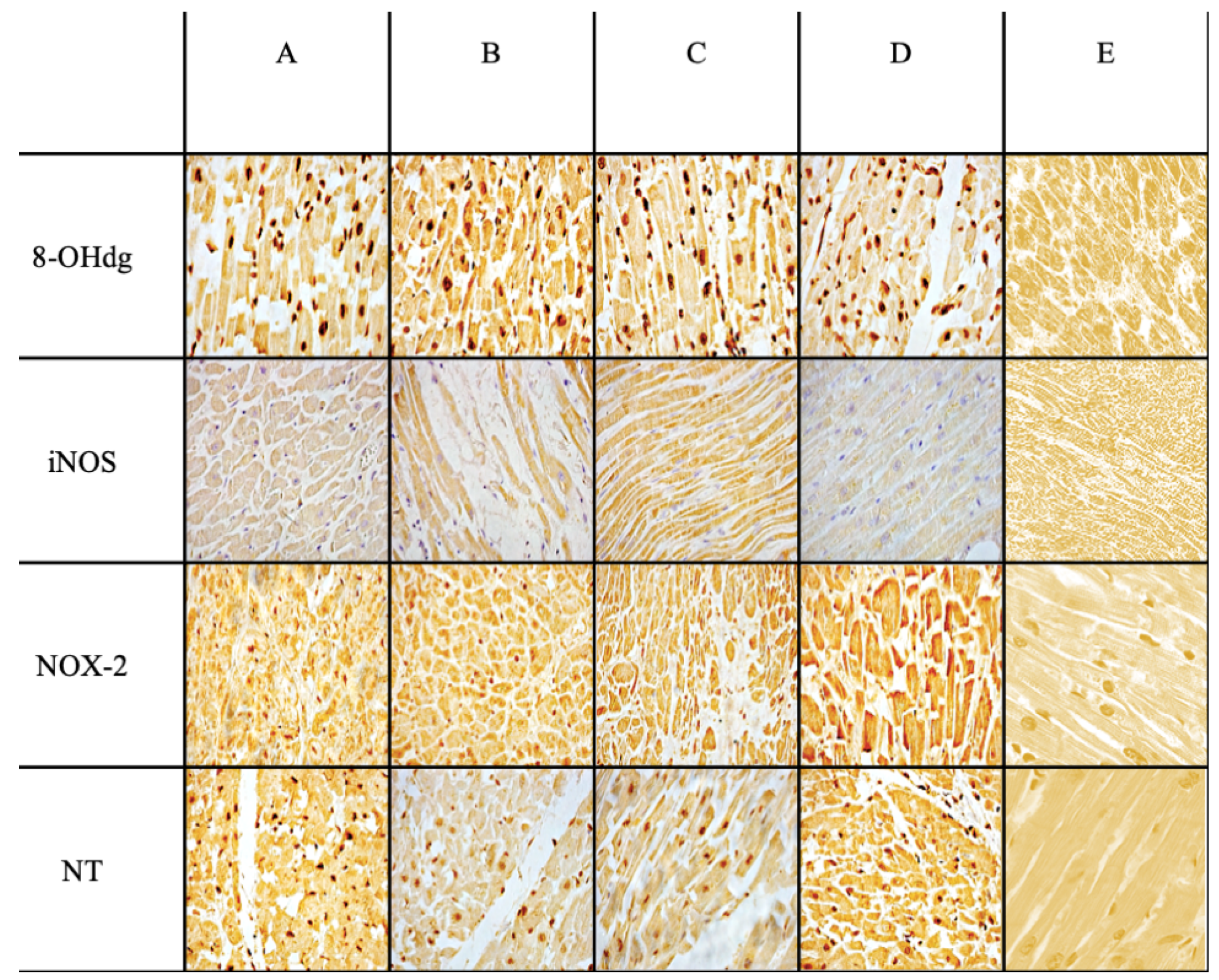
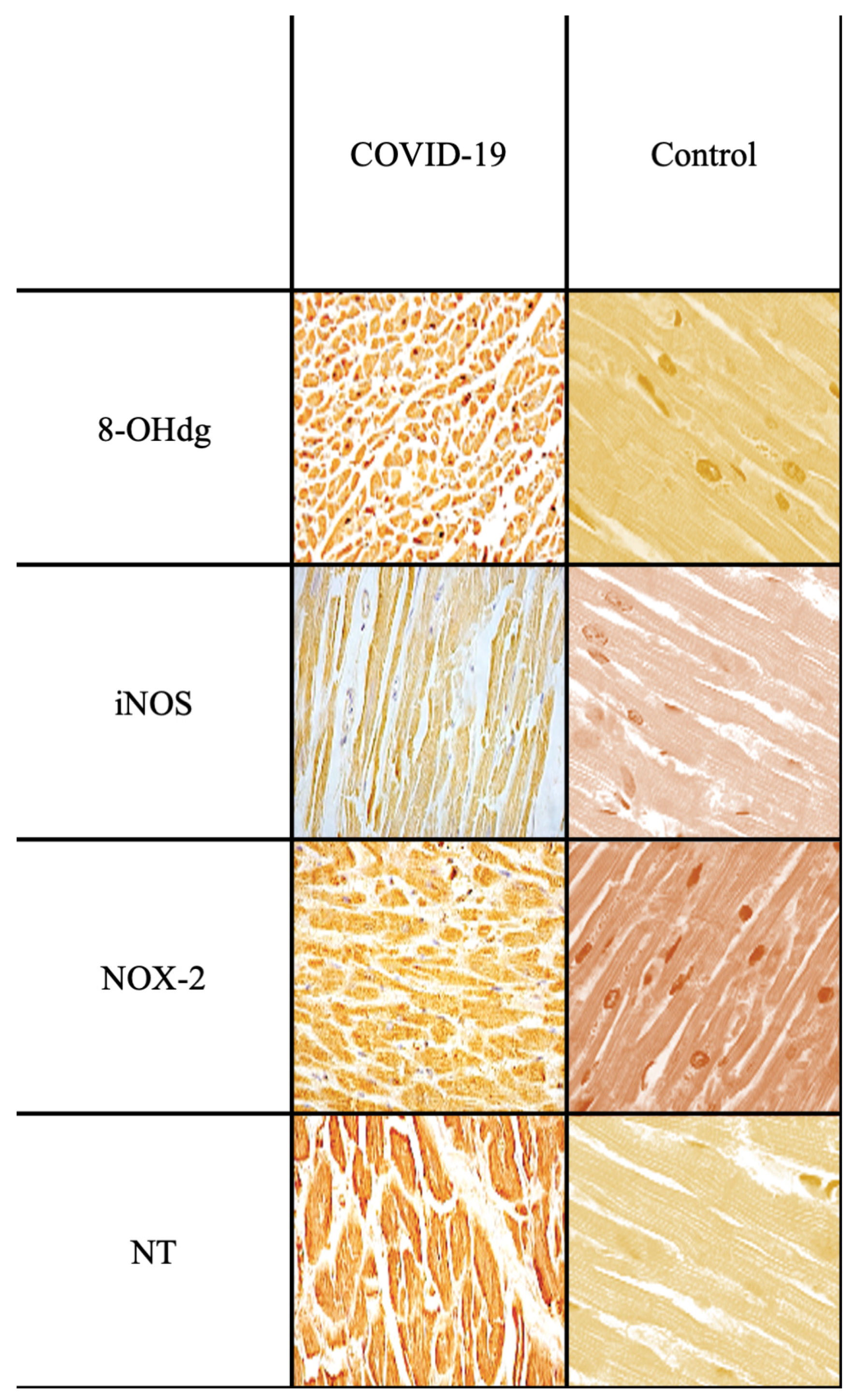
2. Results
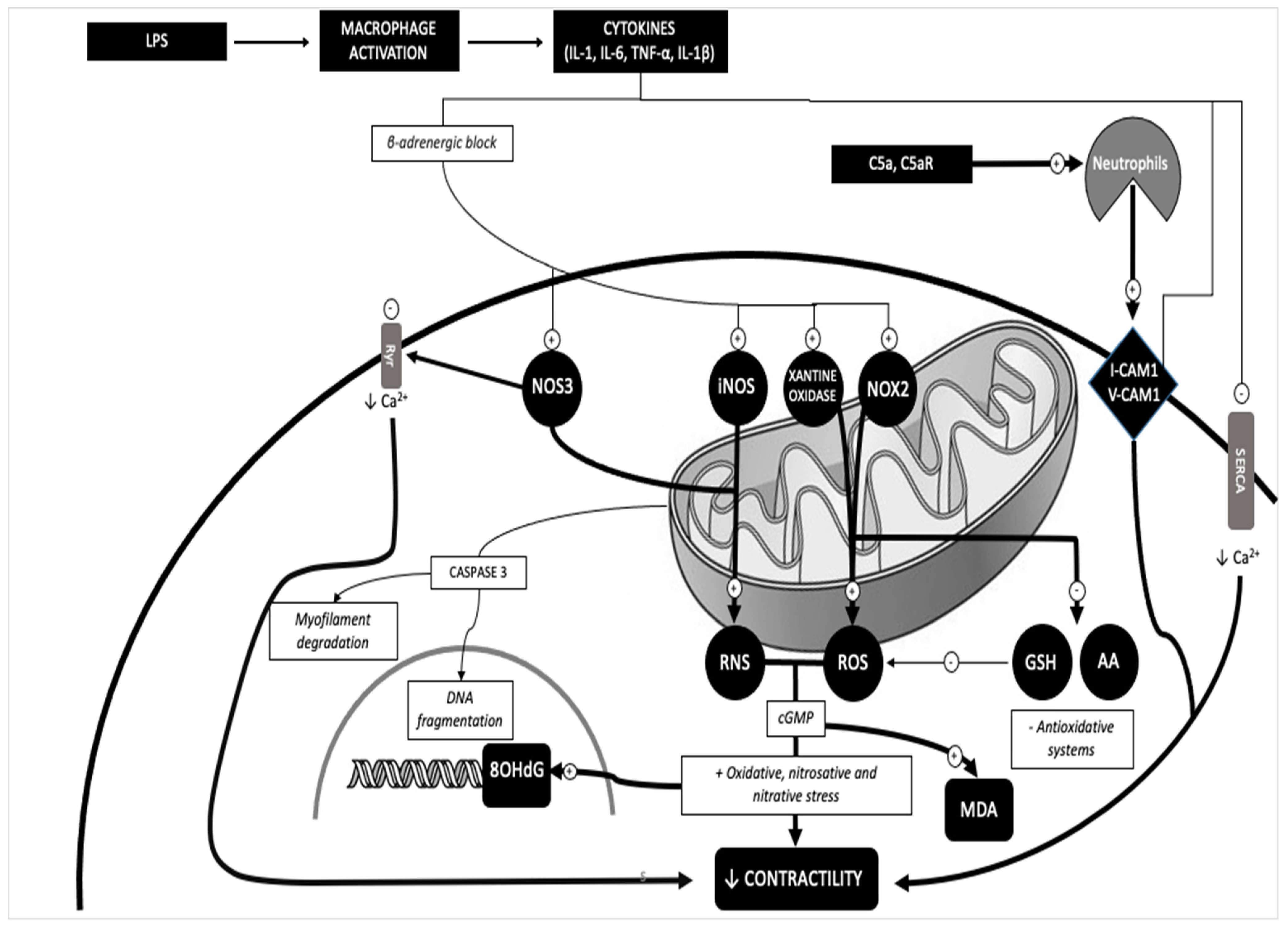
3. Discussion
4. Materials and Methods
4.1. Case Selection
4.2. IHC
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801. [Google Scholar] [CrossRef]
- WHO. Current evidence, identifying gaps and future directions. In Global Report on the Epidemiology and Burden of Sepsis; WHO: Geneva, Switzerland, 2020. [Google Scholar]
- Guarino, M.; Perna, B.; Cesaro, A.E.; Maritati, M.; Spampinato, M.D.; Contini, C.; De Giorgio, R. 2023 Update on Sepsis and Septic Shock in Adult Patients: Management in the Emergency Department. J. Clin. Med. 2023, 12, 3188. [Google Scholar] [CrossRef]
- Chiu, C.; Legrand, M. Epidemiology of sepsis and septic shock. Curr. Opin. Anaesthesiol. 2021, 34, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Bosmann, M.; Ward, P.A. The Inflammatory Response in Sepsis. Trends Immunol. 2013, 34, 129. [Google Scholar] [CrossRef] [PubMed]
- Sprung, C.L.; Annane, D.; Keh, D.; Moreno, R.; Singer, M.; Freivogel, K.; Weiss, Y.G.; Benbenishty, J.; Kalenka, A.; Forst, H.; et al. Hydrocortisone Therapy for Patients with Septic Shock. N. Engl. J. Med. 2008, 358, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Galley, H.F. Oxidative stress and mitochondrial dysfunction in sepsis. Br. J. Anaesth. 2011, 107, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Andrades, M.T.; Morina, A.; Spasić, S.; Spasojević, I. Bench-to-bedside review: Sepsis—From the redox point of view. Crit. Care 2011, 15, 230. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Solé, C.; Pinilla-González, V.; Lillo-Moya, J.; González-Fernández, T.; Saso, L.; Rodrigo, R. Integrated approach to reducing polypharmacy in older people: Exploring the role of oxidative stress and antioxidant potential therapy. Redox Rep. 2024, 29, 2289740. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Lee, Y.; Kellum, J.A. A new perspective on NO pathway in sepsis and ADMA lowering as a potential ther-apeutic approach. Crit. Care. 2022, 26, 246. [Google Scholar] [CrossRef] [PubMed]
- Cerretani, D.; Bello, S.; Cantatore, S.; Fiaschi, A.I.; Montefrancesco, G.; Neri, M.; Pomara, C.; Riezzo, I.; Fiore, C.; Bonsignore, A.; et al. Acute administration of 3,4-methylenedioxymethamphetamine (MDMA) induces oxidative stress, lipoperoxidation and TNFα-mediated apoptosis in rat liver. Pharmacol. Res. 2011, 64, 517–527. [Google Scholar] [CrossRef]
- Sikora, J.P.; Karawani, J.; Sobczak, J. Neutrophils and the Systemic Inflammatory Response Syndrome (SIRS). Int. J. Mol. Sci. 2023, 24, 13469. [Google Scholar] [CrossRef]
- Karapetsa, M.; Pitsika, M.; Goutzourelas, N.; Stagos, D.; Tousia Becker, A.; Zakynthinos, E. Oxidative status in ICU patients with septic shock. Food Chem. Toxicol. 2013, 61, 106–111. [Google Scholar] [CrossRef]
- Hong, Y.; Chen, P.; Gao, J.; Lin, Y.; Chen, L.; Shang, X. Sepsis-associated encephalopathy: From pathophysiology to clinical management. Int. Immunopharmacol. 2023, 124 Pt A, 110800. [Google Scholar] [CrossRef]
- Abelli, J.; Méndez-Valdés, G.; Gómez-Hevia, F.; Bragato, M.C.; Chichiarelli, S.; Saso, L.; Rodrigo, R. Potential Antiox-idant Multitherapy against Complications Occurring in Sepsis. Biomedicines 2022, 10, 3088. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R.; Nyska, A. Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [PubMed]
- Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, J.M. Lipid peroxidation and antioxidants as bio-markers of tissue damage. Clin. Chem. 1995, 41, 1819–1828. [Google Scholar] [CrossRef] [PubMed]
- Nedeva, C. Inflammation and cell death of the innate and adaptive immune system during sepsis. Biomolecules 2021, 11, 1011. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Calvani, R.; Coelho Junior, H.J.; Marzetti, E. Cell death and inflammation: The role of mitochondria in health and disease. Cells 2021, 10, 537. [Google Scholar] [CrossRef]
- Yu, Y.; Yan, Y.; Niu, F.; Wang, Y.; Chen, X.; Su, G.; Liu, Y.; Zhao, X.; Qian, L.; Liu, P.; et al. Ferroptosis: A cell death connecting oxidative stress, inflammation and cardiovascular diseases. Cell Death Discov. 2021, 7, 193. [Google Scholar] [CrossRef]
- Aki, T.; Unuma, K.; Uemura, K. The Role of Peroxiredoxins in the Regulation of Sepsis. Antioxidants 2022, 11, 126. [Google Scholar] [CrossRef]
- Pravda, J. Hydrogen peroxide and disease: Towards a unified system of pathogenesis and therapeutics. Mol. Med. 2020, 26, 41. [Google Scholar] [CrossRef]
- Robinson, N.; Ganesan, R.; Hegedűs, C.; Kovács, K.; Kufer, T.A.; Virág, L. Programmed necrotic cell death of macro-phages: Focus on pyroptosis, necroptosis, and parthanatos. Redox Biol. 2019, 26, 101239. [Google Scholar] [CrossRef]
- Turillazzi, E.; Cerretani, D.; Cantatore, S.; Fiaschi, A.I.; Frati, P.; Micheli, L.; Neri, M.; Cipolloni, L.; Di Paolo, M.; Pinchi, E.; et al. Myocardial oxidative damage is induced by cardiac Fas-dependent and mitochondria-dependent apoptotic pathways in human cocaine-related overdose. Sci. Rep. 2017, 7, 44262. [Google Scholar] [CrossRef]
- Brown, G.C.; Borutaite, V. Inhibition of mitochondrial respiratory complex I by nitric oxide, peroxynitrite and S-nitrosothiols. Biochim. Biophys. Acta 2004, 1658, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Cassina, A.; Radi, R. Differential inhibitory action of nitric oxide and peroxynitrite on mitochondrial electrontransport. Arch. Biochem. Biophys. 1996, 328, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Standiford, T.J.; Rahman, A.; Newstead, M.; Holland, S.M.; Dinauer, M.C.; Liu, Q.; Malik, A.B. Role of NADPH oxidase in the mechanism of lung neutrophil sequestration and microvessel injury induced by Gram-negative sepsis: Studies in p47phox-/- and gp91phox-/- mice. J. Immunol. 2002, 168, 3974–3982. [Google Scholar] [CrossRef] [PubMed]
- Russo, I.; Barale, C.; Melchionda, E.; Penna, C.; Pagliaro, P. Platelets and Cardioprotection: The Role of Nitric Oxide and Carbon Oxide. Int. J. Mol. Sci. 2023, 24, 6107. [Google Scholar] [CrossRef]
- Pall, M.L. The NO/ONOO-cycle as the central cause of heart failure. Int. J. Mol. Sci. 2013, 14, 22274–22330. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, E.; Fiaccadori, E.; Donadello, K.; Taccone, F.S.; Franchi, F.; Scolletta, S. Myocardial depression in sepsis: From pathogenesis to clinical manifestations and treatment. J. Crit. Care. 2014, 29, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Landesberg, G.; Gilon, D.; Meroz, Y.; Georgieva, M.; Levin, P.D.; Goodman, S.; Avidan, A.; Beeri, R.; Weissman, C.; Jaffe, A.S.; et al. Diastolic dysfunction and mortality in severe sepsis and septic shock. Eur. Heart J. 2012, 33, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Zanotti-Cavazzoni, S.L.; Hollenberg, S.M. Cardiac dysfunction in severe sepsis and septic shock. Curr. Opin. Crit. Care 2009, 15, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Ozer, E.K.; Goktas, M.T.; Kilinc, I.; Toker, A.; Bariskaner, H.; Ugurluoglu, C.; Iskit, A.B. Infliximab alleviates the mortality, mesenteric hypoperfusion, aortic dysfunction, and multiple organ damage in septic rats. Can. J. Physiol. Pharmacol. 2017, 95, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Xianchu, L.; Lan, P.Z.; Qiufang, L.; Yi, L.; Xiangcheng, R.; Wenqi, H.; Yang, D. Naringin protects against lipopolysaccha-ride-induced cardiac injury in mice. Environ. Toxicol. Pharmacol. 2016, 48, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Wang, B.; He, B.; Feng, M.; Yu, Y. LPS induces cardiomyocyte necroptosis through the Ripk3/Pgam5 signaling pathway. J. Recept. Signal Transduct. Res. 2021, 41, 32–37. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Misra, M.K.; Sarwat, M.; Bhakuni, P.; Tuteja, R.; Tuteja, N. Oxidative stress and ischemic myocardial syndromes. Med. Sci. Monit. 2009, 15, RA209–RA219. [Google Scholar]
- Gauthier, L.D.; Greenstin, J.L.; Cortassa, S.; O’Rourke, B.; Winslow, R.L. A computational model of reactive oxygen species and redox balance in cardiac mitochondria. Biophys. J. 2013, 105, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Nagar, H.; Piao, S.; Kim, C.S. Role of mitochondrial oxidative stress in sepsis. Acute Crit. Care 2018, 33, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Huet, O.; Dupic, L.; Harrois, A.; Duranteau, J. Oxidative stress and endothelial dysfunction during sepsis. Front. Biosci. 2011, 16, 1986–1995. [Google Scholar] [CrossRef]
- Kayar, S.R.; Banchero, N. Volume density and distribution of mitochondria in myocardial growth and hypertrophy. Respir. Physiol. 1987, 70, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Beretta, M.; Santos, C.X.; Molenaar, C.; Hafstad, A.D.; Miller, C.C.; Revazian, A.; Betteridge, K.; Schröder, K.; Streckfuß Bömeke, K.; Doroshow, J.H. Nox4 regulates InsP3 receptor dependent Ca2+ release into mitochondria to promote cell survival. EMBO J. 2020, 39, e103530. [Google Scholar] [CrossRef]
- Kim, J.; Kwon, J.; Kim, M.; Do, J.; Lee, D.; Han, H. Low dielectric constant polyimide aerogel composite films with low water uptake. Polym. J. 2016, 48, 829–834. [Google Scholar] [CrossRef]
- Weiss, J.N.; Korge, P.; Honda, H.M.; Ping, P. Role of the mitochondrial permeability transition in myocardial disease. Circ. Res. 2003, 93, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Larche, J.; Lancel, S.; Hassoun, S.M.; Favory, R.; Decoster, B.; Marchetti, P.; Chopin, C.; Neviere, R. Inhibition of mi-tochondrial permeability transition prevents sepsis induced myocardial dysfunction and mortality. J. Am. Coll. Cardiol. 2006, 48, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Zou, R.; Tao, J.; Qiu, J.; Lu, H.; Wu, J.; Zhu, H.; Li, R.; Mui, D.; Toan, S.; Chang, X.; et al. DNA-PKcs promotes sepsis-induced multiple organ failure by triggering mitochondrial dysfunction. J. Adv. Res. 2022, 41, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Ravingerová, T.; Kindernay, L.; Barteková, M.; Ferko, M.; Adameová, A.; Zohdi, V.; Bernátová, I.; Ferenczyová, K.; Lazou, A. The molecular mechanisms of iron metabolism and its role in cardiac dysfunction and cardioprotection. Int. J. Mol. Sci. 2020, 21, 7889. [Google Scholar] [CrossRef]
- Li, N.; Wang, W.; Zhou, H.; Wu, Q.; Duan, M.; Liu, C.; Wu, H.; Deng, W.; Shen, D.; Tang, Q. Ferritinophagy-mediated ferroptosis is involved in sepsis-induced cardiac injury. Free Radic. Biol. Med. 2020, 160, 303–318. [Google Scholar] [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Andrews, N.C. Balancing acts: Molecular control of mammalian iron metabolism. Cell 2004, 117, 285–297. [Google Scholar] [CrossRef]
- Tang, D.; Chen, X.; Kang, R.; Kroemer, G. Ferroptosis: Molecular mechanisms and health implications. Cell Res. 2020, 31, 107–125. [Google Scholar] [CrossRef]
- Han, C.; Liu, Y.; Dai, R.; Ismail, N.; Su, W.; Li, B. Ferroptosis and its potential role in human diseases. Front. Pharmacol. 2020, 11, 239. [Google Scholar] [CrossRef]
- Xiao, Z.; Kong, B.; Fang, J.; Qin, T.; Dai, C.; Shuai, W.; Huang, H. Ferrostatin-1 alleviates lipopolysaccharide-induced cardiac dysfunction. Bioengineered 2021, 12, 9367–9376. [Google Scholar] [CrossRef]
- Liu, V.W.; Zhang, C.; Nagley, P. Mutations in mitochondrial DNA accumulate differentially in three different human tissues during ageing. Nucleic Acids Res. 1998, 26, 1268–1275. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.A.; Hanke, T.; Erasmi, A.W.; Bechtel, M.J.; Scharfschwerdt, M.; Meissner, C.; Sievers, H.H.; Gosslau, A. Mitochondrial DNA deletions and the aging heart. Exp. Gerontol. 2006, 41, 508–517. [Google Scholar] [CrossRef]
- Barja, G.; Herrero, A. Oxidative damage to mitochondrial DNA is inversely related to maximum life span in the heart and brain of mammals. FASEB J. 2000, 14, 312–318. [Google Scholar] [CrossRef]
- Riezzo, I.; Cerretani, D.; Fiore, C.; Bello, S.; Centini, F.; D’Errico, S.; Fiaschi, A.I.; Giorgi, G.; Neri, M.; Pomara, C.; et al. Enzymatic-nonenzymatic cellular antioxidant defense systems response and immunohisto-chemical detection of MDMA, VMAT2, HSP70, and apoptosis as biomarkers for MDMA (Ecstasy) neurotoxicity. J Neurosci. Res. 2010, 88, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Tsolaki, V.; Makris, D.; Mantzarlis, K.; Zakynthinos, E. Sepsis-Induced Cardiomyopathy: Oxidative Implications in the Initiation and Resolution of the Damage. Oxid. Med. Cell. Longev. 2017, 2017, 7393525. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M.; Lambeth, J.D.; Nauseef, W. The neutrophil NADPH oxidase. Arch. Biochem. Biophys. 2002, 397, 342–344. [Google Scholar] [CrossRef]
- Lambeth, J.D.; Cheng, D.; Arnold, R.S.; Edens, W.A. Novel homologs of gp91phox. Trends Biochem. Sci. 2000, 25, 459–461. [Google Scholar] [CrossRef]
- Hernansanz-Agustín, P.; Enríquez, J.A. Generation of Reactive Oxygen Species by Mitochondria. Antioxidants 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Lu, X.; Feng, Q. Pivotal Role of gp91phox-Containing NADH Oxidase in Lipopolysaccharide-Induced Tumor Necrosis Factor-α Expression and Myocardial Depression. Circulation 2005, 111, 1637–1644. [Google Scholar] [CrossRef] [PubMed]
- Rongjin, H.; Feng, C.; Jun, K.; Shirong, L. Oxidative Stress-Induced Protein of SESTRIN2 in Cardioprotection Effect. Dis. Markers 2022, 2022, 7439878. [Google Scholar] [CrossRef] [PubMed]
- Vanden Berghe, T.; Kaiser, W.J.; Bertrand, M.J.; Vandenabeele, P. Molecular crosstalk between apoptosis, necroptosis, and survival signaling. Mol. Cell Oncol. 2015, 2, e975093. [Google Scholar] [CrossRef] [PubMed]
- Lafont, E.; Hartwig, T.; Walczak, H. Paving trail’s path with ubiquitin. Trends Biochem. Sci. 2018, 43, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Vucur, M.; Roderburg, C.; Kaiser, L.; Schneider, A.T.; Roy, S.; Loosen, S.H.; Luedde, M.; Trautwein, C.; Koch, A.; Tacke, F.; et al. Elevated serum levels of mixed lineage kinase domain like protein predict survival of patients during intensive care unit treatment. Dis. Markers 2018, 2018, 1983421. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Xu, J.; Ruan, W.; Li, S.; Xiao, F. PPAR γ activation prevents septic cardiac dysfunction via inhibition of apoptosis and necroptosis. Oxid. Med. Cell. Longev. 2017, 2017, 8326749. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, V.V.S.; Reis, J.F.; de Souza Gomes, R.; Navegantes, K.C.; Monteiro, M.C. Dual Behavior of Exosomes in Septic Cardiomyopathy. Adv. Exp. Med. Biol. 2017, 998, 101–112. [Google Scholar] [CrossRef]
- Maiese, A.; Frati, P.; Del Duca, F.; Santoro, P.; Manetti, A.C.; La Russa, R.; Di Paolo, M.; Turillazzi, E.; Fineschi, V. Myocardial Pathology in COVID-19-Associated Cardiac Injury: A Systematic Review. Diagnostics 2021, 11, 1647. [Google Scholar] [CrossRef]
- Maccio, U.; Zinkernagel, A.S.; Shambat, S.M.; Zeng, X.; Cathomas, G.; Ruschitzka, F.; Schuepbach, R.A.; Moch, H.; Varga, Z. SARS-CoV-2 leads to a small vessel endotheliitis in the heart. EBioMedicine 2021, 63, 103182. [Google Scholar] [CrossRef]
- Shi, S.; Qin, M.; Shen, B.; Cai, Y.; Liu, T.; Yang, F.; Gong, W.; Liu, X.; Liang, J.; Zhao, Q.; et al. Association of Cardiac Injury with Mortality in Hospitalized Patients With COVID-19 in Wuhan, China. JAMA Cardiol. 2020, 5, 802–810. [Google Scholar] [CrossRef]
- Guo, T.; Fan, Y.; Chen, M.; Wu, X.; Zhang, L.; He, T.; Wang, H.; Wan, J.; Wang, X.; Lu, Z. Cardiovascular Implications of Fatal Outcomes of Patients with Coronavirus Disease 2019 (COVID-19). JAMA Cardiol. 2020, 5, 811–818. [Google Scholar] [CrossRef]
- Wendel Garcia, P.D.; Fumeaux, T.; Guerci, P.; Heuberger, D.M.; Montomoli, J.; Roche-Campo, F.; Schuepbach, R.A.; Hilty, M.P.; RISC-19-ICU Investigators. Prognostic factors associated with mortality risk and disease progression in 639 critically ill patients with COVID-19 in Europe: Initial report of the international RISC-19-ICU prospective observational cohort. EClinicalMedicine 2020, 25, 100449. [Google Scholar] [CrossRef]
- Wenzel, P.; Kopp, S.; Göbel, S.; Jansen, T.; Geyer, M.; Hahn, F.; Kreitner, K.F.; Escher, F.; Schultheiss, H.P.; Münzel, T. Evidence of SARS-CoV-2 mRNA in endomyocardial biopsies of patients with clinically suspected myocarditis tested negative for COVID-19 in nasopharyngeal swab. Cardiovasc. Res. 2020, 116, 1661–1663. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S.; O’brien, J.; Tollefsen, K.E.; Kim, Y.; Chauhan, V.; Yauk, C.; Huliganga, E.; Rudel, R.A.; Kay, J.E.; Helm, J.S.; et al. Reactive Oxygen Species in the Adverse Outcome Pathway Framework: Toward Creation of Harmonized Consensus Key Events. Front. Toxicol. 2022, 4, 887135. [Google Scholar] [CrossRef]
- Piccioni, A.; Franza, L.; Rosa, F.; Candelli, M.; Covino, M.; Ferrara, M.; Volonnino, G.; Bertozzi, G.; Zamponi, M.V.; Maiese, A.; et al. The role of SARS-CoV-2 infection in promoting abnormal immune response and sepsis: A comparison between SARS-CoV-2-related sepsis and sepsis from other causes. Infect. Med. 2023, 2, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xu, Y.; Fang, Y.; Wang, C.; Xue, Y.; Wang, F.; Cheng, J.; Ren, H.; Wang, J.; Guo, W.; et al. Pathogenetic mechanisms of septic cardiomyopathy. J. Cell. Physiol. 2022, 237, 49–58. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, Y.; Liu, W.; Liu, W.; Dong, J.; Liu, Q.; Hao, H.; Ren, H. Research progress on the activation mechanism of NLRP3 inflammasome in septic car-diomyopathy. Immun. Inflamm. Dis. 2023, 11, e1039. [Google Scholar] [CrossRef]
- Hollenberg, S.M.; Singer, M. Pathophysiology of sepsis-induced cardiomyopathy. Nat. Rev. Cardiol. 2021, 18, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Maiese, A.; Scatena, A.; Costantino, A.; Chiti, E.; Occhipinti, C.; La Russa, R.; Di Paolo, M.; Turillazzi, E.; Frati, P.; Fineschi, V. Expression of MicroRNAs in Sepsis-Related Organ Dysfunction: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 9354. [Google Scholar] [CrossRef] [PubMed]
- La Russa, R.; Maiese, A.; Viola, R.V.; De Matteis, A.; Pinchi, E.; Frati, P.; Fineschi, V. Searching for highly sensitive and specific biomarkers for sepsis: State-of-the-art in post-mortem diagnosis of sepsis through immunohistochemical analysis. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419855226. [Google Scholar] [CrossRef] [PubMed]
- Galassi, A.; Turatello, L.; De Salvia, A.; Neri, M.; Turillazzi, E.; La Russa, R.; Viola, R.V.; Frati, P.; Fineschi, V. Septic cardiomyopathy: The value of lactoferrin and CD15 as specific markers to corroborate a definitive diagnosis. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418776526. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ab | Control Group | Sepsis Group | Statistical Value | Sepsis vs. Control |
|---|---|---|---|---|
| Anti-iNOS | +/− | +++ | *** | *** |
| Anti-NOX-2 | +/− | +++ | *** | *** |
| Anti-Nitrotyrosine | + | +++ | *** | *** |
| Anti-8-OHdG | +/− | +++ | *** | *** |
| Case | Sex, Age | Cause of Death Attributed |
|---|---|---|
| CASE 1 | F, 57 y.o. | Septic shock secondary to pneumonia |
| CASE 2 | M, 62 y.o. | Septic shock secondary to Fournier’s gangrene |
| CASE 3 | M, 68 y.o. | Septic shock secondary to peritonitis due to bowel perforation |
| CASE 4 | F, 75 y.o. | Septic shock secondary to peritonitis due to VAP |
| CASE 5 | M, 27 y.o. | Septic shock secondary to pneumonia |
| CASE 6 | M, 45 y.o. | Septic shock secondary to COVID-19 pneumonia |
| CASE 7 | M, 68 y.o. | Septic shock secondary to cholecystitis |
| CASE 8 | F, 85 y.o. | Septic shock secondary to pneumonia |
| CASE 9 | M, 62 y.o. | Septic shock secondary to bowel perforation |
| CASE 10 | F, 56 y.o. | Septic shock secondary to peritonitis due to enterocolitis |
| NC | M, 22 y.o | Head trauma from a shotgun |
| NC | F, 30 y.o. | Traffic accident resulting in the immediate death |
| NC | M, 44 y.o. | Traffic accident resulting in the immediate death |
| NOX-2 | gp91-phox, goat polyclonal antibody, sc-5826 | Santa Cruz Biotechnology, Inc., Dallas, TX, USA |
| NT | nitrotyrosine, mouse monoclonal antibody, sc-32757 | Santa Cruz Biotechnology, Inc., Dallas, TX, USA |
| iNOS | nitric oxide sinthases-2, mouse monoclonal antibody, sc-7271 | Santa Cruz Biotechnology, Inc., Dallas, TX, USA |
| 8-OHdG | 8-hidroxy-2′-deoxyguanosine, mouse monoclonal antibody, N45.1 | JaICA, Nikken SEIL Co., Fukuroi, Shizuoka, Japan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertozzi, G.; Ferrara, M.; Di Fazio, A.; Maiese, A.; Delogu, G.; Di Fazio, N.; Tortorella, V.; La Russa, R.; Fineschi, V. Oxidative Stress in Sepsis: A Focus on Cardiac Pathology. Int. J. Mol. Sci. 2024, 25, 2912. https://doi.org/10.3390/ijms25052912
Bertozzi G, Ferrara M, Di Fazio A, Maiese A, Delogu G, Di Fazio N, Tortorella V, La Russa R, Fineschi V. Oxidative Stress in Sepsis: A Focus on Cardiac Pathology. International Journal of Molecular Sciences. 2024; 25(5):2912. https://doi.org/10.3390/ijms25052912
Chicago/Turabian StyleBertozzi, Giuseppe, Michela Ferrara, Aldo Di Fazio, Aniello Maiese, Giuseppe Delogu, Nicola Di Fazio, Vittoria Tortorella, Raffaele La Russa, and Vittorio Fineschi. 2024. "Oxidative Stress in Sepsis: A Focus on Cardiac Pathology" International Journal of Molecular Sciences 25, no. 5: 2912. https://doi.org/10.3390/ijms25052912