
Fibroblast Growth Factor Receptor Inhibitors Decrease Proliferation of Melanoma Cell Lines and Their Activity Is Modulated by Vitamin D
 , ,
, ,
Abstract
:1. Introduction
2. Results
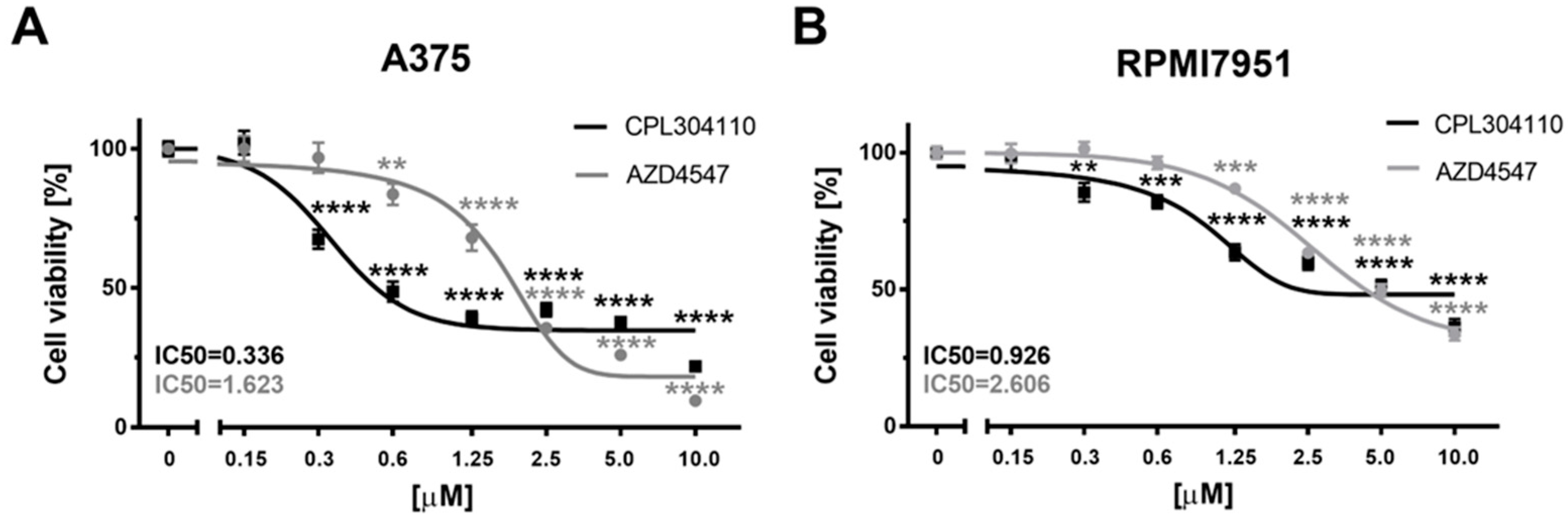
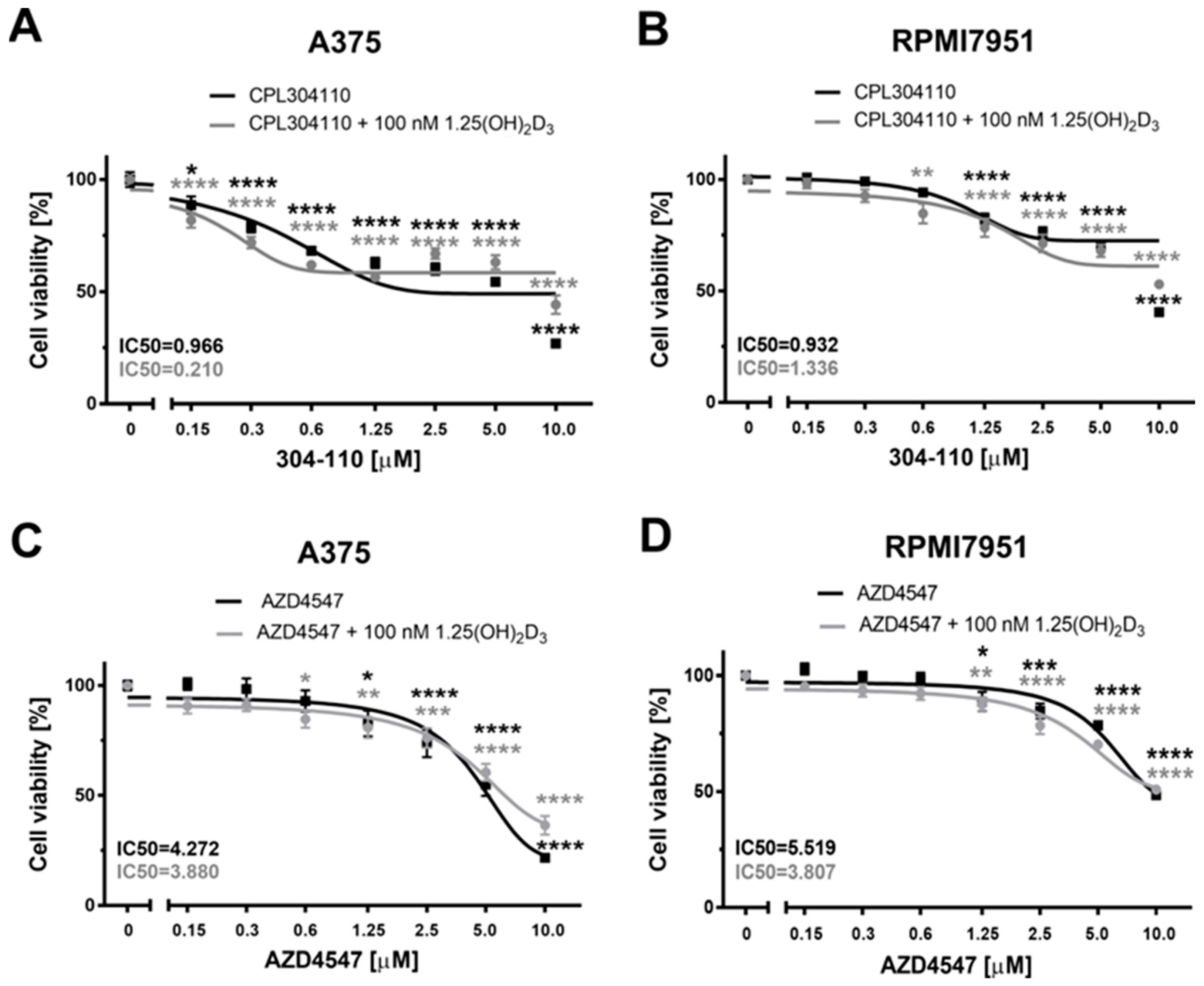
2.1. FGFR Inhibitors Efficiently Decrease Melanoma Cells’ Viability—The Efficacy of 304-110 Is Modulated by Vitamin D in A375 Melanoma Cells
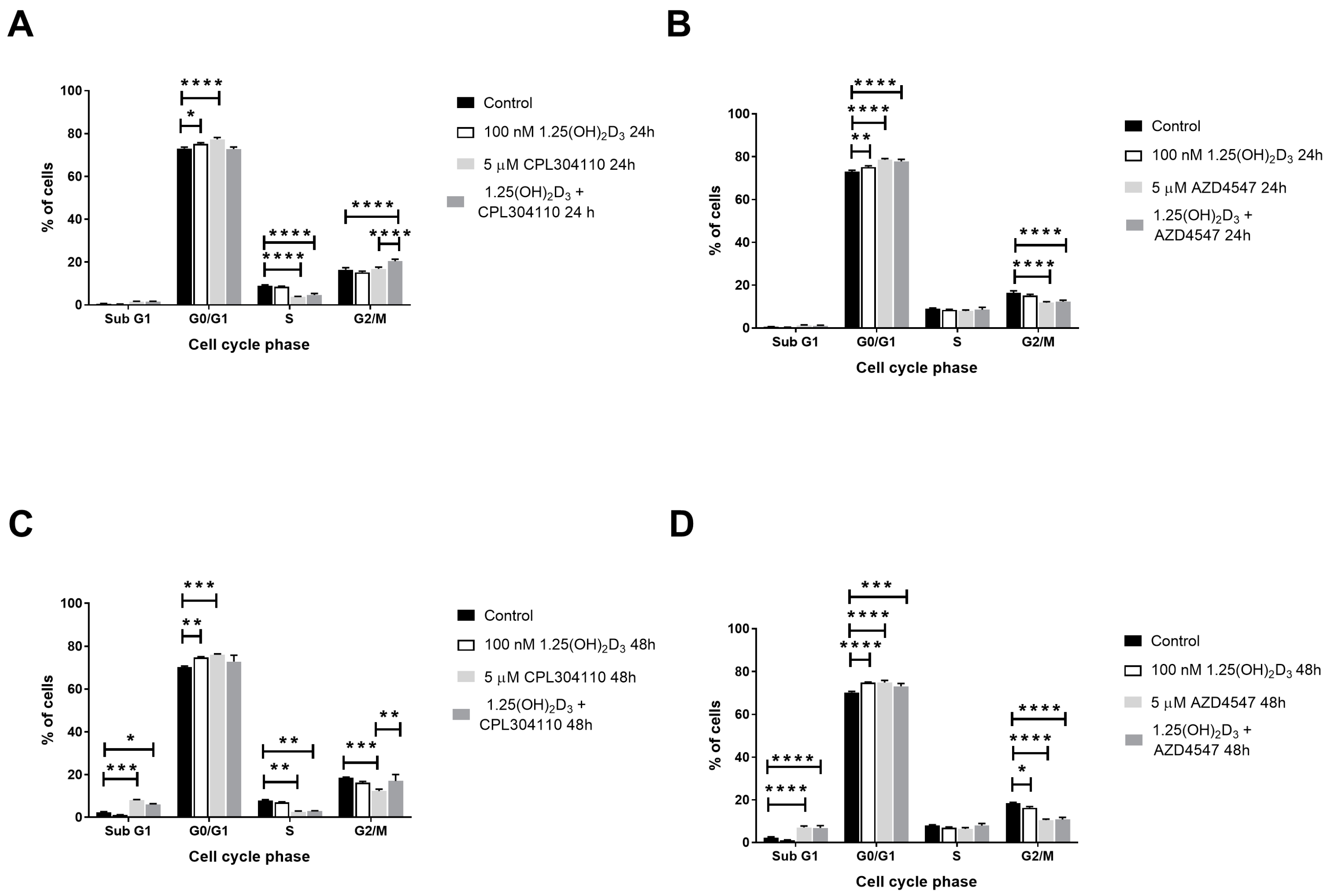
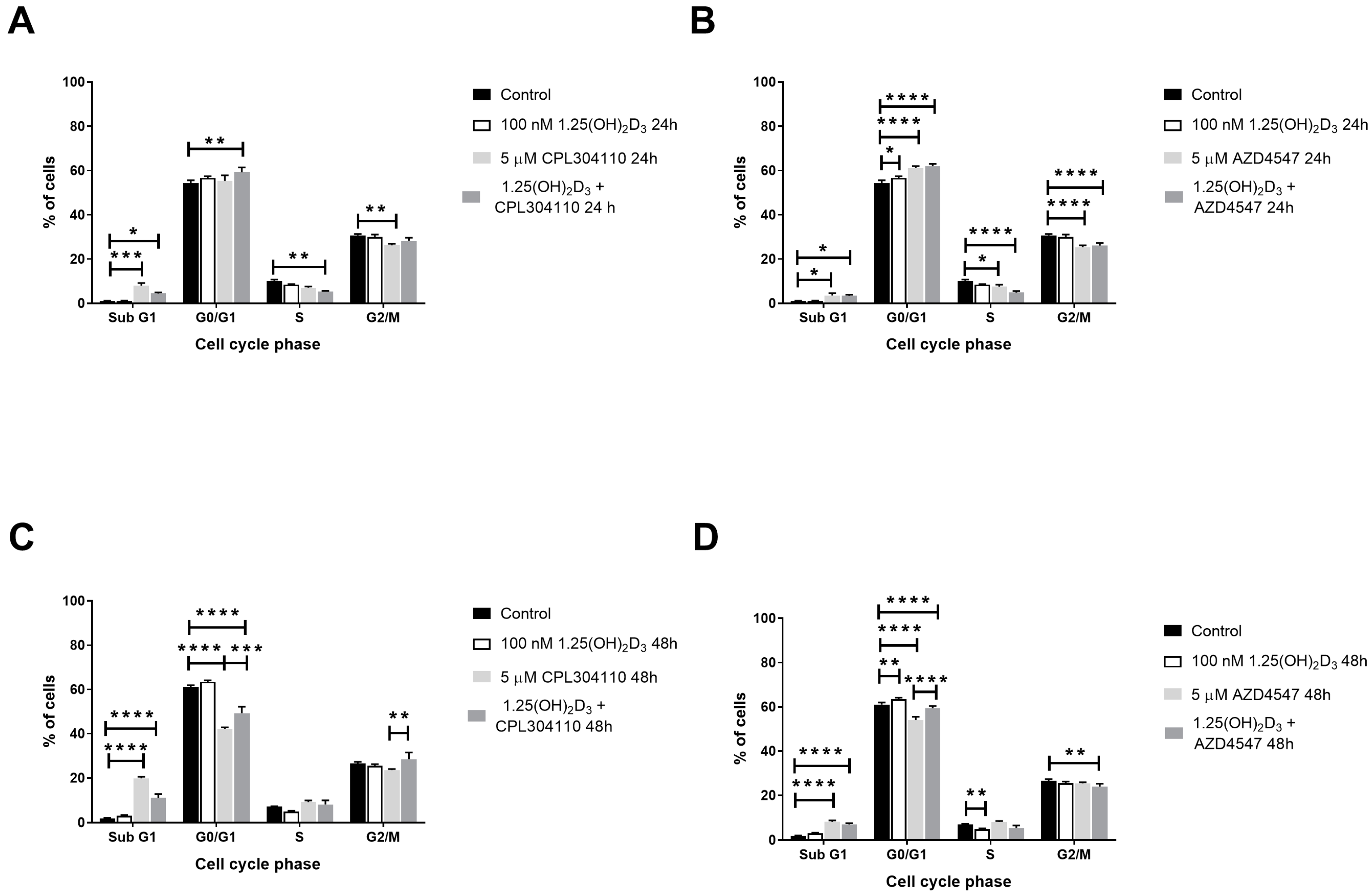
2.2. FGFR Inhibitors Trigger G0/G1 Cell Cycle Arrest in A375 Melanoma Cells and Increase SubG1 Fraction in RPMI7951 Melanoma Cells, and Their Activity Is Modulated by Vitamin D
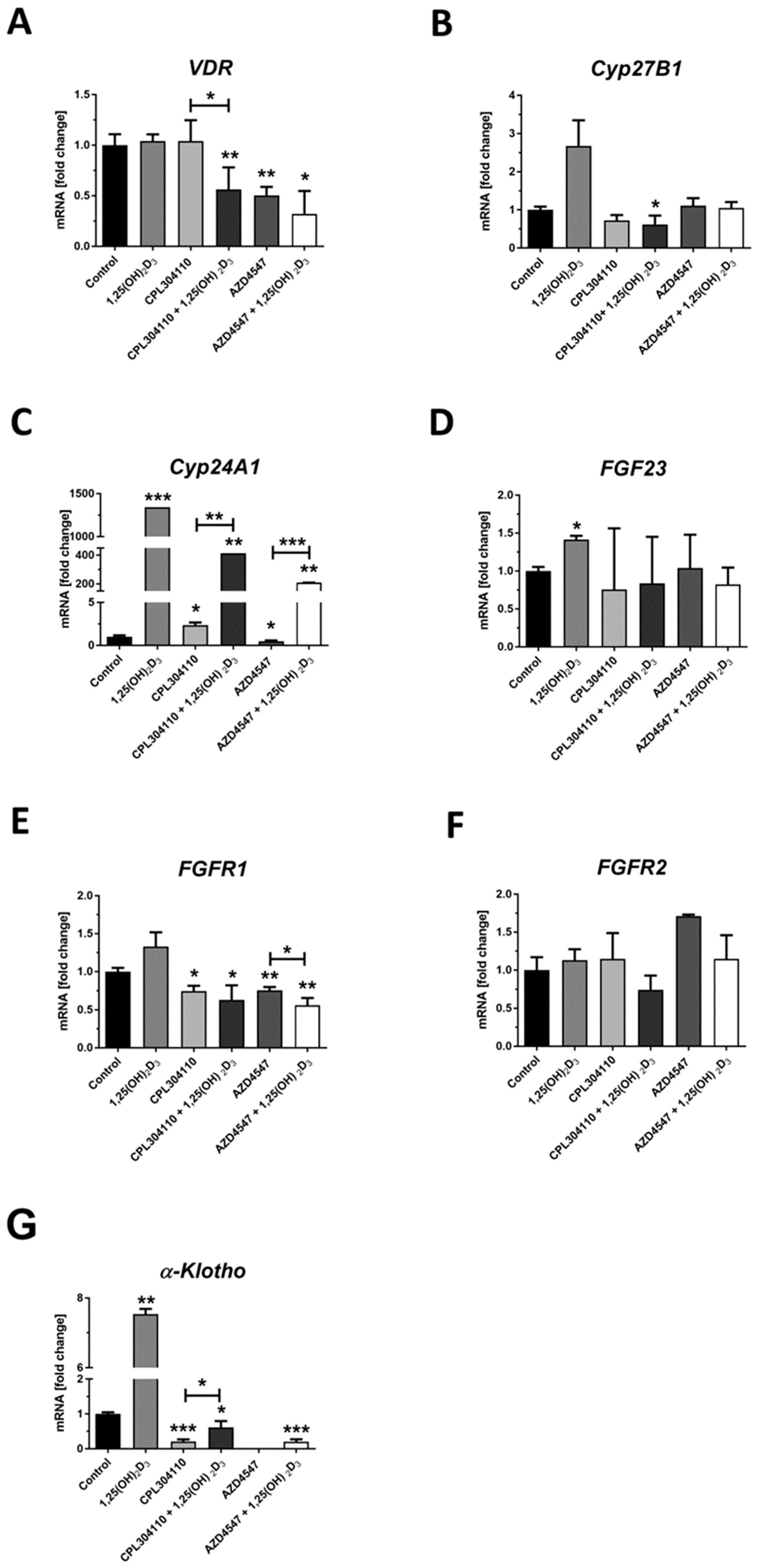
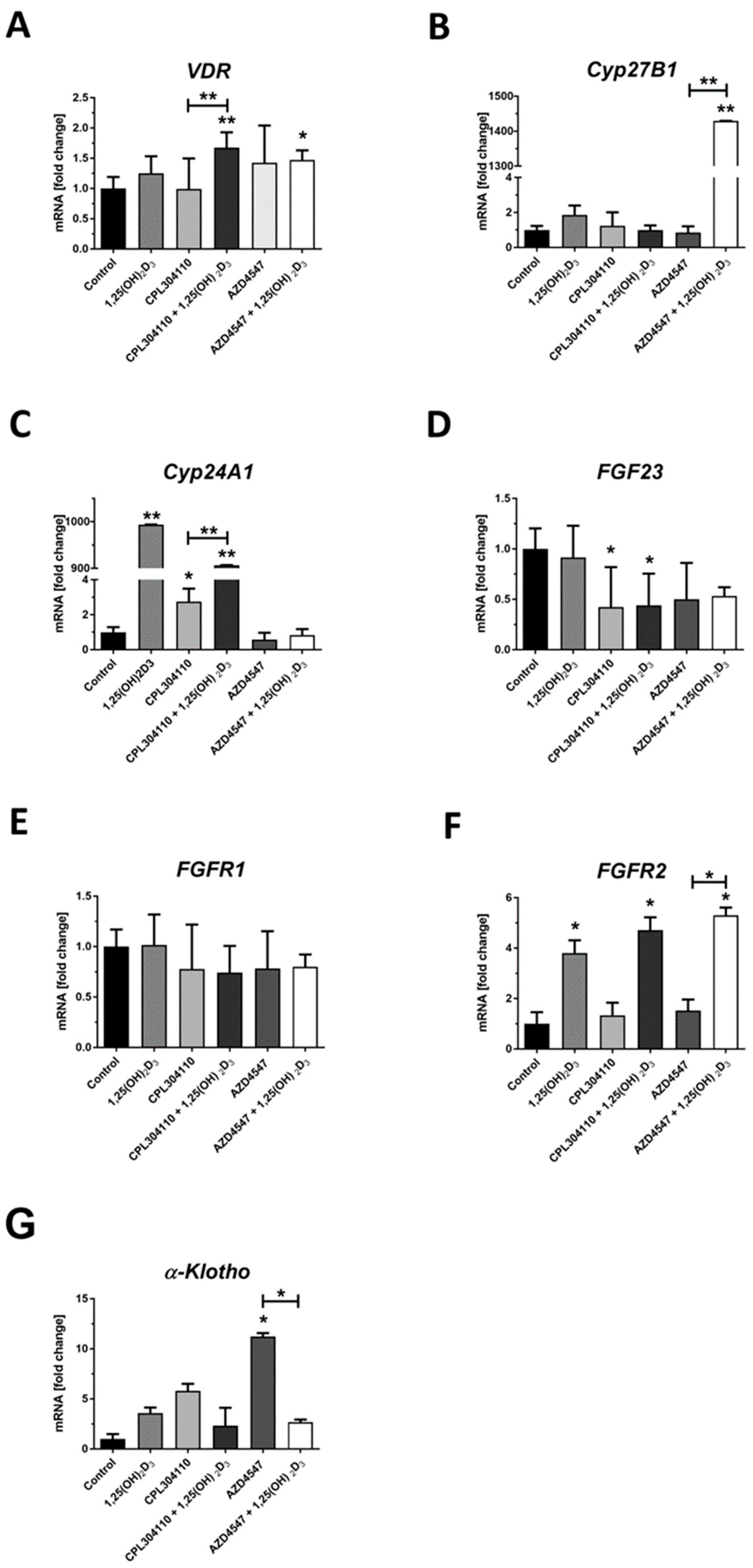
2.3. FGFR Inhibitors Modulate the Expression of FGFs, FGF23, α-KLOTHO and Vitamin D-Related Genes at mRNA Level
2.4. 1,25(OH)2D3 Regulates Levels of FGFR1 and 2 Receptors, as Well as Activation of FGFRs and ERK1/2 in A375 Melanoma Cells Treated with FGFR Inhibitors
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Viability Assay
4.4. Cell Cycle Analysis
4.5. RT-PCR
4.6. Immunoblotting
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garbe, C.; Amaral, T.; Peris, K.; Hauschild, A.; Arenberger, P.; Basset-Seguin, N.; Bastholt, L.; Bataille, V.; Del Marmol, V.; Dréno, B.; et al. European consensus-based interdisciplinary guideline for melanoma. Part 1: Diagnostics: Update 2022. Eur. J. Cancer 2022, 170, 236–255. [Google Scholar] [CrossRef]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Ascierto, P.A.; Puzanov, I.; Agarwala, S.S.; Blank, C.; Carvajal, R.D.; Demaria, S.; Dummer, R.; Ernstoff, M.; Ferrone, S.; Fox, B.A.; et al. Perspectives in melanoma: Meeting report from the “Melanoma Bridge” (December 5th–7th, 2019, Naples, Italy). J. Transl. Med. 2020, 18, 346. [Google Scholar] [CrossRef]
- Patton, E.E.; Mueller, K.L.; Adams, D.J.; Anandasabapathy, N.; Aplin, A.E.; Bertolotto, C.; Bosenberg, M.; Ceol, C.J.; Burd, C.E.; Chi, P.; et al. Melanoma models for the next generation of therapies. Cancer Cell 2021, 39, 610–631. [Google Scholar] [CrossRef] [PubMed]
- Trotta, A.P.; Gelles, J.D.; Serasinghe, M.N.; Loi, P.; Arbiser, J.L.; Chipuk, J.E. Disruption of mitochondrial electron transport chain function potentiates the pro-apoptotic effects of MAPK inhibition. J. Biol. Chem. 2017, 292, 11727–11739. [Google Scholar] [CrossRef]
- Imbert, C.; Montfort, A.; Fraisse, M.; Marcheteau, E.; Gilhodes, J.; Martin, E.; Bertrand, F.; Marcellin, M.; Burlet-Schiltz, O.; Peredo, A.G.; et al. Resistance of melanoma to immune checkpoint inhibitors is overcome by targeting the sphingosine kinase-1. Nat. Commun. 2020, 11, 437. [Google Scholar] [CrossRef] [PubMed]
- Trojaniello, C.; Vitale, M.G.; Scarpato, L.; Esposito, A.; Ascierto, P.A. Melanoma immunotherapy: Strategies to overcome pharmacological resistance. Expert Rev. Anticancer. Ther. 2020, 20, 289–304. [Google Scholar] [CrossRef]
- Czyz, M. Fibroblast Growth Factor Receptor Signaling in Skin Cancers. Cells 2019, 8, 540. [Google Scholar] [CrossRef] [PubMed]
- Krook, M.A.; Reeser, J.W.; Ernst, G.; Barker, H.; Wilberding, M.; Li, G.; Chen, H.Z.; Roychowdhury, S. Fibroblast growth factor receptors in cancer: Genetic alterations, diagnostics, therapeutic targets and mechanisms of resistance. Br. J. Cancer 2021, 124, 880–892. [Google Scholar] [CrossRef]
- Zarczynska, I.; Gorska-Arcisz, M.; Cortez, A.J.; Kujawa, K.A.; Wilk, A.M.; Skladanowski, A.C.; Stanczak, A.; Skupinska, M.; Wieczorek, M.; Lisowska, K.M.; et al. p38 Mediates Resistance to FGFR Inhibition in Non-Small Cell Lung Cancer. Cells 2021, 10, 3363. [Google Scholar] [CrossRef] [PubMed]
- Shan, K.S.; Dalal, S.; Thaw Dar, N.N.; McLish, O.; Salzberg, M.; Pico, B.A. Molecular Targeting of the Fibroblast Growth Factor Receptor Pathway across Various Cancers. Int. J. Mol. Sci. 2024, 25, 849. [Google Scholar] [CrossRef] [PubMed]
- Gartside, M.G.; Chen, H.; Ibrahimi, O.A.; Byron, S.A.; Curtis, A.V.; Wellens, C.L.; Bengston, A.; Yudt, L.M.; Eliseenkova, A.V.; Ma, J.; et al. Loss-of-function fibroblast growth factor receptor-2 mutations in melanoma. Mol. Cancer Res. 2009, 7, 41–54. [Google Scholar] [CrossRef]
- de Aguiar, R.B.; Parise, C.B.; Souza, C.R.; Braggion, C.; Quintilio, W.; Moro, A.M.; Navarro Marques, F.L.; Buchpiguel, C.A.; Chammas, R.; de Moraes, J.Z. Blocking FGF2 with a new specific monoclonal antibody impairs angiogenesis and experimental metastatic melanoma, suggesting a potential role in adjuvant settings. Cancer Lett. 2016, 371, 151–160. [Google Scholar] [CrossRef]
- Ronca, R.; Di Salle, E.; Giacomini, A.; Leali, D.; Alessi, P.; Coltrini, D.; Ravelli, C.; Matarazzo, S.; Ribatti, D.; Vermi, W.; et al. Long pentraxin-3 inhibits epithelial-mesenchymal transition in melanoma cells. Mol. Cancer Ther. 2013, 12, 2760–2771. [Google Scholar] [CrossRef]
- Giehl, K.A.; Nägele, U.; Volkenandt, M.; Berking, C. Protein expression of melanocyte growth factors (bFGF, SCF) and their receptors (FGFR-1, c-kit) in nevi and melanoma. J. Cutan. Pathol. 2007, 34, 7–14. [Google Scholar] [CrossRef]
- Li, L.; Zhang, S.; Li, H.; Chou, H. FGFR3 promotes the growth and malignancy of melanoma by influencing EMT and the phosphorylation of ERK, AKT, and EGFR. BMC Cancer 2019, 19, 963. [Google Scholar] [CrossRef]
- Yamani, A.; Zdżalik-Bielecka, D.; Lipner, J.; Stańczak, A.; Piórkowska, N.; Stańczak, P.S.; Olejkowska, P.; Hucz-Kalitowska, J.; Magdycz, M.; Dzwonek, K.; et al. Discovery and optimization of novel pyrazole-benzimidazole CPL304110, as a potent and selective inhibitor of fibroblast growth factor receptors FGFR (1–3). Eur. J. Med. Chem. 2020, 210, 112990. [Google Scholar] [CrossRef]
- Piotrowska, A.; Wierzbicka, J.; Rybarczyk, A.; Tuckey, R.C.; Slominski, A.T.; Zmijewski, M.A. Vitamin D and its low calcemic analogs modulate the anticancer properties of cisplatin and dacarbazine in the human melanoma A375 cell line. Int. J. Oncol. 2019, 54, 1481–1495. [Google Scholar] [CrossRef]
- Piotrowska, A.; Beserra, F.P.; Wierzbicka, J.M.; Nowak, J.I.; Żmijewski, M.A. Vitamin D Enhances Anticancer Properties of Cediranib, a VEGFR Inhibitor, by Modulation of VEGFR2 Expression in Melanoma Cells. Front. Oncol. 2021, 11, 763895. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, A.; Zaucha, R.; Król, O.; Żmijewski, M.A. Vitamin D Modulates the Response of Patient-Derived Metastatic Melanoma Cells to Anticancer Drugs. Int. J. Mol. Sci. 2023, 24, 8037. [Google Scholar] [CrossRef]
- Wöhrle, S.; Bonny, O.; Beluch, N.; Gaulis, S.; Stamm, C.; Scheibler, M.; Müller, M.; Kinzel, B.; Thuery, A.; Brueggen, J.; et al. FGF receptors control vitamin D and phosphate homeostasis by mediating renal FGF-23 signaling and regulating FGF-23 expression in bone. J. Bone Miner. Res. 2011, 26, 2486–2497. [Google Scholar] [CrossRef]
- Jeon, S.M.; Shin, E.A. Exploring vitamin D metabolism and function in cancer. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, A.; Grant, W.B. Vitamin D and Cancer: An Historical Overview of the Epidemiology and Mechanisms. Nutrients 2022, 14, 1448. [Google Scholar] [CrossRef]
- Donati, S.; Palmini, G.; Aurilia, C.; Falsetti, I.; Miglietta, F.; Iantomasi, T.; Brandi, M.L. Rapid Nontranscriptional Effects of Calcifediol and Calcitriol. Nutrients 2022, 14, 1291. [Google Scholar] [CrossRef]
- Kamiya, S.; Nakamori, Y.; Takasawa, A.; Takasawa, K.; Kyuno, D.; Ono, Y.; Magara, K.; Osanai, M. Vitamin D metabolism in cancer: Potential feasibility of vitamin D metabolism blocking therapy. Med Mol. Morphol. 2023, 56, 85–93. [Google Scholar] [CrossRef]
- Gibot, L.; Galbraith, T.; Huot, J.; Auger, F.A. Development of a tridimensional microvascularized human skin substitute to study melanoma biology. Clin. Exp. Metastasis 2013, 30, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Wasiewicz, T.; Piotrowska, A.; Wierzbicka, J.; Slominski, A.T.; Zmijewski, M.A. Antiproliferative Activity of Non-Calcemic Vitamin D Analogs on Human Melanoma Lines in Relation to VDR and PDIA3 Receptors. Int. J. Mol. Sci. 2018, 19, 2583. [Google Scholar] [CrossRef]
- Kommalapati, A.; Tella, S.H.; Borad, M.; Javle, M.; Mahipal, A. FGFR Inhibitors in Oncology: Insight on the Management of Toxicities in Clinical Practice. Cancers 2021, 13, 2968. [Google Scholar] [CrossRef]
- Katoh, M. FGFR inhibitors: Effects on cancer cells, tumor microenvironment and whole-body homeostasis (Review). Int. J. Mol. Med. 2016, 38, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, A.; Wierzbicka, J.; Slebioda, T.; Wozniak, M.; Tuckey, R.C.; Slominski, A.T.; Zmijewski, M.A. Vitamin D derivatives enhance cytotoxic effects of H2O2 or cisplatin on human keratinocytes. Steroids 2016, 110, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Kuro, O.M. The Klotho proteins in health and disease. Nat. Rev. Nephrol. 2019, 15, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Sarabipour, S.; Hristova, K. Mechanism of FGF receptor dimerization and activation. Nat. Commun. 2016, 7, 10262. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- Morales-Guadarrama, G.; Méndez-Pérez, E.A.; García-Quiroz, J.; Avila, E.; Larrea, F.; Díaz, L. AZD4547 and calcitriol synergistically inhibited BT-474 cell proliferation while modified stemness and tumorsphere formation. J. Steroid Biochem. Mol. Biol. 2022, 223, 106132. [Google Scholar] [CrossRef]
- Giacomini, A.; Taranto, S.; Rezzola, S.; Matarazzo, S.; Grillo, E.; Bugatti, M.; Scotuzzi, A.; Guerra, J.; Di Trani, M.; Presta, M.; et al. Inhibition of the FGF/FGFR System Induces Apoptosis in Lung Cancer Cells via c-Myc Downregulation and Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 9376. [Google Scholar] [CrossRef]
- Göke, A.; Göke, R.; Ofner, A.; Herbst, A.; Lankat-Buttgereit, B. The FGFR Inhibitor NVP-BGJ398 Induces NSCLC Cell Death by Activating Caspase-dependent Pathways as well as Caspase-independent Apoptosis. Anticancer Res. 2015, 35, 5873–5879. [Google Scholar]
- Sun, S.; Jiang, Y.; Zhang, G.; Song, H.; Zhang, X.; Zhang, Y.; Liang, X.; Sun, Q.; Pang, D. Increased expression of fibroblastic growth factor receptor 2 is correlated with poor prognosis in patients with breast cancer. J. Surg. Oncol. 2012, 105, 773–779. [Google Scholar] [CrossRef]
- Huang, T.; Liu, D.; Wang, Y.; Li, P.; Sun, L.; Xiong, H.; Dai, Y.; Zou, M.; Yuan, X.; Qiu, H. FGFR2 Promotes Gastric Cancer Progression by Inhibiting the Expression of Thrombospondin4 via PI3K-Akt-Mtor Pathway. Cell. Physiol. Biochem. 2018, 50, 1332–1345. [Google Scholar] [CrossRef]
- Schrumpf, T.; Behrens, H.M.; Haag, J.; Krüger, S.; Röcken, C. FGFR2 overexpression and compromised survival in diffuse-type gastric cancer in a large central European cohort. PLoS ONE 2022, 17, e0264011. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, T.; Matsuda, Y.; Yamamoto, T.; Uchida, E.; Korc, M.; Naito, Z. Enhanced expression of fibroblast growth factor receptor 2 IIIc promotes human pancreatic cancer cell proliferation. Am. J. Pathol. 2012, 180, 1928–1941. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, R.; Matsuda, Y.; Ishiwata, T.; Naito, Z. Downregulation of fibroblast growth factor receptor 2 and its isoforms correlates with a high proliferation rate and poor prognosis in high-grade glioma. Oncol. Rep. 2014, 32, 1163–1169. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SubG1 | G1 | S | G2/M | |
|---|---|---|---|---|
| A375 24 h | ||||
| Control | 0.57 ± 0.11 | 73.01 ± 0.64 | 9.03 ± 0.33 | 16.43 ± 0.94 |
| 1,25(OH)2D3 | 0.37 ± 0.04 | 75.14 ± 0.60 | 8.47 ± 0.28 | 15.18 ± 0.60 |
| CPL304110 | 1.57 ± 0.15 | 77.26 ± 0.91 | 3.73 ± 0.24 | 16.80 ± 0.81 |
| CPL304110 + 1,25(OH)2D3 | 1.56 ± 0.20 | 72.78 ± 0.93 | 4.69 ± 0.64 | 20.41 ± 0.93 |
| AZD4547 | 1.31 ± 0.18 | 78.56 ± 0.57 | 7.93 ± 0.50 | 11.86 ± 0.46 |
| AZD4547 + 1,25(OH)2D3 | 1.09 ± 0.29 | 77.77 ± 0.96 | 8.55 ± 1.10 | 12.38 ± 0.60 |
| A375 48 h | ||||
| Control | 2.30 ± 0.34 | 70.23 ± 0.48 | 7.00 ± 0.30 | 16.33 ± 0.56 |
| 1,25(OH)2D3 | 1.03 ± 0.17 | 74.79 ± 0.33 | 11.99 ± 1.17 | 20.13 ± 1.34 |
| CPL304110 | 8.03 ± 0.35 | 75.97 ± 0.38 | 2.61 ± 0.34 | 12.37 ± 0.89 |
| CPL304110 + 1,25(OH)2D3 | 6.06 ± 0.38 | 72.79 ± 3.02 | 3.07 ± 0.06 | 17.10 ± 2.97 |
| AZD4547 | 7.07 ± 0.66 | 74.81 ± 1.06 | 6.39 ± 0.61 | 10.48 ± 0.53 |
| AZD4547 + 1,25(OH)2D3 | 6.87 ± 1.05 | 73.11 ± 1.03 | 7.90 ± 1.04 | 10.84 ± 0.97 |
| RPMI7951 24 h | ||||
| Control | 1.06 ± 0.13 | 54.28 ± 1.30 | 10.09 ± 0.64 | 30.74 ± 0.59 |
| 1,25(OH)2D3 | 1.06 ± 0.19 | 56.60 ± 0.86 | 8.48 ± 0.19 | 29.89 ±1.20 |
| CPL304110 | 8.00 ± 1.20 | 55.32 ± 2.45 | 7.03 ± 0.58 | 26.20 ± 0.71 |
| CPL304110 + 1,25(OH)2D3 | 4.41 ± 0.48 | 59.27 ± 2.15 | 5.22 ± 0.37 | 28.23 ± 1.44 |
| AZD4547 | 3.53 ± 1.01 | 61.18 ± 0.81 | 7.46 ± 1.00 | 25.32 ± 0.89 |
| AZD4547 + 1,25(OH)2D3 | 3.41 ± 0.50 | 61.91 ± 1.07 | 4.85 ± 0.72 | 26.07 ± 1.24 |
| RPMI7951 48 h | ||||
| Control | 1.94 ± 0.18 | 61.07 ± 0.87 | 7.09 ± 0.20 | 26.52 ± 0.78 |
| 1,25(OH)2D3 | 3.09 ± 0.29 | 63.41 ± 0.74 | 4.89 ± 0.40 | 25.49 ± 0.74 |
| CPL304110 | 19.81 ± 0.70 | 42.07 ± 0.81 | 9.20 ± 0.67 | 23.52 ± 0.50 |
| CPL304110 + 1,25(OH)2D3 | 11.07 ± 1.67 | 49.30 ± 2.86 | 8.11 ± 1.79 | 28.47 ± 2.99 |
| AZD4547 | 8.17 ± 0.63 | 53.98 ± 1.53 | 8.06 ± 0.47 | 25.37 ± 0.56 |
| AZD4547 + 1,25(OH)2D3 | 7.03 ± 0.58 | 59.35 ± 1.04 | 5.42 ± 1.15 | 23.87 ± 1.33 |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| RPL37A | CCATTTCAGGCGGCGGTAGTCTT | ACGGTGTCTTTCTCGTTCAC |
| VDR | AGTAAGTGTGCTTGACCTCC | GAGAAGTCTGTACTACCTGAGC |
| CYP27B1 | AGACTCGTTTACGTTTGT | GACATACTCGAGAGGGCC |
| CYP24A1 | TTTAGACGTGATCCGACG | GTTGTCAAGACCCACTTA |
| FGF23 | CGACACCGACACATGTCCAC | TAGTCTCCTACGACCGAAAC |
| FGFR1 | CATCAGCTACACCACTTACAGG | TCTCACTACACACCAGAAAGC |
| FGFR2 | GAAACAGTTAAGGGTGACGAAG | GGAAAGACTAGACCACAGTCTC |
| α-KLOTHO | CAATAAGAAGTACGAGAGCCCT | GAAACCTGGGTGGAACTCAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piotrowska, A.; Nowak, J.I.; Wierzbicka, J.M.; Domżalski, P.; Górska-Arcisz, M.; Sądej, R.; Popiel, D.; Wieczorek, M.; Żmijewski, M.A. Fibroblast Growth Factor Receptor Inhibitors Decrease Proliferation of Melanoma Cell Lines and Their Activity Is Modulated by Vitamin D. Int. J. Mol. Sci. 2024, 25, 2505. https://doi.org/10.3390/ijms25052505
Piotrowska A, Nowak JI, Wierzbicka JM, Domżalski P, Górska-Arcisz M, Sądej R, Popiel D, Wieczorek M, Żmijewski MA. Fibroblast Growth Factor Receptor Inhibitors Decrease Proliferation of Melanoma Cell Lines and Their Activity Is Modulated by Vitamin D. International Journal of Molecular Sciences. 2024; 25(5):2505. https://doi.org/10.3390/ijms25052505
Chicago/Turabian StylePiotrowska, Anna, Joanna I. Nowak, Justyna M. Wierzbicka, Paweł Domżalski, Monika Górska-Arcisz, Rafał Sądej, Delfina Popiel, Maciej Wieczorek, and Michał A. Żmijewski. 2024. "Fibroblast Growth Factor Receptor Inhibitors Decrease Proliferation of Melanoma Cell Lines and Their Activity Is Modulated by Vitamin D" International Journal of Molecular Sciences 25, no. 5: 2505. https://doi.org/10.3390/ijms25052505